
Abstract—The mechanisms of the ketamine antidepressant effects observed in humans and laboratory animals are not fully understood. To further clarify the role of brain serotonergic (5-HT) activity in the action of antidepressant drugs, optogenetic inhibition of 5-HT neurons in the rat dorsal raphe nucleus (DRN) was applied. In control animals, a subanesthetic dose of ketamine alleviated their depressive-like behavior in the tail suspension test. Inhibition of 5-HT neurons abolished the drug effect and moreover, a sedative response to ketamine was found under these conditions. Furthermore, optogenetic suppression of the activity of 5-HT neurons prevented the increase in c-Fos expression induced by ketamine both in light-sensitive neurons and in other DRN neurons. The data emphasize the key role of 5-HT neuron activity in the rapid ketamine antidepressant effect.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
Major depressive disorder is a multifactorial disease characterized by dysfunctions of neurotransmitter systems in the brain [1, 2]. Ketamine is one of the fast-acting antidepressants that is effective in the clinic, even in patients suffering from depression that is resistant to treatment with other drugs [3, 4]. The antidepressant effect of ketamine primarily manifests itself through antagonism to glutamate receptors of the NMDAR type [5, 6]. Two possible routes of action have been suggested for NMDAR-related effects of ketamine: preferential inhibition of NMDAR on GABAergic interneurons and thereby disinhibition of glutamatergic neurons, or inhibition of NMDAR on pyramidal neurons, resulting in reduced downregulation of eukaryotic elongation factor 2 and increased synaptic plasticity and activity of these neurons [7–9].
In addition to glutamatergic effects, ketamine also modulates monoaminergic neurotransmission. It increases the activity of serotonergic (5-HT) neurons in the dorsal raphe nucleus (DRN) [9, 10] and in the prefrontal cortex (PFC) it increases the extracellular level of 5-HT and the excitation it causes [11, 12]. At the same time, 5-HT1A PFC receptors are stimulated, which activate the PI3K/Akt/mTORC1 signaling pathway [10]. Ketamine also modulates the binding of 5-HT to its type 1B receptors [13], whose activation is involved in the antidepressant action [14].
DRN activity is regulated by several glutamatergic afferent projections, including those coming from the PFC [10], which control active/passive behavioral state selection and social choice [11]. The ketamine-induced increase in the activity of the 5-HT system may be mediated by glutamatergic projections of the medial prefrontal cortex (mPFC) to the DRN [12]. This is supported by the fact that local injection of ketamine into the DRN has no antidepressant effect, in contrast to local injection of this drug into the mPFC [15]. Optogenetic stimulation of excitatory afferents from mPFC to DRN activates 5-HT DRN neurons [16]. However, such stimulation also activates GABAergic DRN neurons, which inhibit 5-HT neurons [17]. Ketamine also activates cholinergic neurons of the pedunculopontine nucleus of the tegmentum, giving projections to DRN neurons, and an increased level of acetylcholine stimulates presynaptic α4β2 nicotinic receptors localized on glutamatergic DRN neurons [18], which leads to an increase in local release of glutamate. Glutamate activation of AMPA receptors on 5-HT DRN neurons induces the release of 5-HT into mPFCs [15]. There is no doubt that ketamine affects the activity of 5-HT neurons of the DRN, and its antidepressant effects may be related to its action on the 5-HT system [12, 14–16]. Although pharmacological blockade of 5-HT synthesis attenuates or eliminates the persistent behavioral effects of ketamine [19, 20], lowering 5-HT levels does not affect the acute effects of ketamine, and only hinders the development of long-term antidepressant effects [21]. Therefore, it remains unclear whether the activation of 5-HT DRN neurons is an important link in the chain of rapid antidepressant action of ketamine.
In general, despite a significant amount of information about the interaction of glutamate and 5-HT in the mechanism of action of ketamine, the role of activation of 5-HT DRN neurons in the manifestation of the antidepressant effect of this drug remains unclear. To eliminate this gap, in this work, we studied the effect of optogenetic [22] rapid suppression of the activity of 5-HT DRN neurons, in the manifestation of the antidepressant effect of ketamine.
MATERIALS AND METHODS
For optogenetic suppression of the activity of 5-HT neurons, lentiviral particles were created based on plasmids obtained from Prof. Nishitani [23], using transfection of an HEK293 cell culture. The plasmid TPH2-eArchT3.0-eYFP-WPRE contains the sequence of the archearhodopsin (eArchT3.0) proton pump gene conjugated to yellow fluorescent protein (eYFP), whose expression is regulated by the TPH2 promoter. The TPH2-Venus-WPRE (Venus) plasmid encoding the Venus green fluorescent protein was used as a control. Cells were transfected with a mixture of plasmids for assembly of the viral capsid (pPAX2 and pMD2.G) and TPH2-eArchT3.0-eYFP-WPRE or TPH2-Venus-WPRE using polyethyleneimine [24]. Viral particles were isolated from cell culture using PEG 6000, precipitated on quaternary ammonium columns (DIAPAC-TA, ZAO BioKhimMak ST, Russia) and concentrated to a titer of 1012 particles/mL on Amicon Ultra-4 centrifuge concentrators (Merck, United States) [25].
Adult, 2.5–3 month male Wistar rats weighing 220–250 g were used in the experiments. The animals were kept in the vivarium of the Institute of Cytology and Genetics of the Siberian Branch of the Russian Academy of Sciences, 4 in cages (40 cm × 25 cm × 15 cm) with a 12-hour light/dark cycle and a temperature of 20–22°C. All procedures were carried out in accordance with the International European Bioethical Standards (86/609-EEC), Russian Rules for Working on Laboratory Animals (N 267 19.06.2003) and were approved by the Bioethical Committee of the Institute of Cytology and Genetics of the Siberian Branch of the Russian Academy of Sciences. All manipulations with animals were performed between 09:00 a.m. and 1:00 p.m.. The scheme of the experiment is shown in Fig. 1.

The scheme of the experiment.
Viral particles were injected into DRN stereotaxically along the coordinates AP = –7.7 mm, ML = 0 mm, DV = 7 mm [26]. Animals were anesthetized with Avertin, fixed in a stereotaxic frame, and 1 µL of viral particles was injected through a small hole in the skull with a neuros microsyringe (Hamilton, United States) for 7 min. The skin incision was sutured and treated with antibiotics.
A week later, fiber optic cannulas were stereotaxically implanted into the DRN area and fixed to the skull. For this, an enlarged hole in the skull, made for the introduction of the virus, was used, followed by postoperative processing.
One week after the implantation of fiber optic cannulas, four animals from each group were injected with ketamine at a subanesthetic dose (15 mg/kg body weight), and four others received an equivalent volume of saline (0.5 mL/100 g body weight) intraperitoneally. One hour after the injection, all animals were subjected to optical stimulation during the tail suspension test. To do this, a fiber optic cable was connected to the cannula, after which the animals were quickly hung with adhesive tape by the area of the base of the tail at a height of 60 cm from the floor for 5 min. At 1, 3, and 5 min of the test, DRN was irradiated with green light (560 nm, 1.2 mV/mm2, 20 Hz). In one rat of the eArchT3.0-eYFP × Ketamine group, the opto-cable detached from the cannula and the animal was excluded from the experiment. Behavior in the test was recorded on a video camera and the immobility time in the test was assessed by experts who did not know which group the animal belonged to.
Animals anesthetized with Avertin were transcardially perfused 1 h after the test with 1 × PBS and 4% PFA. The extracted brain was post-fixed for 12 h in 4% paraformaldehyde, then for 24 h in 30% sucrose, after which it was frozen in Richard-Allan Scientific Neg-50 gel (Thermo Fisher Scientific, United States) and stored at –70°C.
For immunohistochemical analysis, sections 20-μm thick were prepared on a cryotome, which were dried at 37°C for 1 h before staining and washed for 10 min in PBS with 0.2% Triton X-100 (PBST). Nonspecific binding was blocked by incubation in PBST containing 1.5% BSA for 1 h at room temperature. With primary antibodies (Table 1) sections were incubated overnight at +4°C. Antibodies were diluted in PBST containing 1.5% BSA at a dilution of 1 : 200. After incubation with primary antibodies, sections were washed twice for 15 min in PBST and incubated with secondary antibodies (Table 1) and hybridized 1 : 500 for 2 h at room temperature. They were then washed twice in PBST, once in PBS for 15 min, and then embedded in moveol containing the DAPI nuclear dye (4',6-diamidino-2-phenylindole) [27].
The preparations were photographed using an LSM780NLO confocal microscope (Carl Zeiss, Germany) at 20×. The photographs were analyzed using the QuPath software [28]. The number of c-Fos-, eArchT3.0-eYFP/Venus- and TPH2-expressing neurons per 1 mm2 was counted, as well as the number of neurons co-expressing eArchT3.0-eYFP/Venus + TPH2 to assess the specificity and efficiency of expression of viral vectors, as well as eArchT3.0-eYFP/Venus + c-Fos cells to assess the effect of light irradiation on neuronal activity. For this, the QuPath software used the detection of cells by nuclei stained with DAPI, followed by classification based on the counting of green and red pixels. The intensity threshold for pixel detection and classification was selected for each type of staining separately and applied to all photographs in the sample in the same way.
From the quantitative data, the mean value was calculated with a standard deviation from the mean. To assess c-Fos expression in vector-expressing neurons, the percentage of cells co-expressing eArchT3.0-eYFP/Venus + c-Fos of the total number of vector-expressing cells was calculated using the formula (number of co-expressing cells/number of c-Fos expressing cells) × 100%.
The Kolmogorov–Smirnov test was used to check the normality of the data. Behavioral test parameters were processed by two-way analysis of the variance for repeated measures (factor 1, vector; factor 2, ketamine), and the results of immunohistochemical analysis of optogenetic modulation of neuronal activity in sensitive and light-insensitive groups of animals were processed by two-way analysis of the variance (factor 1, vector; factor 2, ketamine) followed by the a posteriori Fisher comparison. The result was considered significant at p < 0.05. Statistical calculations were performed using the STATISTICA 10.0 software package.
RESULTS
Analysis of depression-like behavior in the tail suspension test revealed a significant effect of the vector quality (photosensitive or not) on the immobility time (Vector: F(1, 11) = 12.681, p < 0.004) and presence of interaction of the Vector × Ketamine factors: (F(1, 11) = 18.508, p < 0.001). In animals with the control vector, a ketamine-induced decrease in immobility time was observed, which was especially pronounced at 2 and 3 min of the test. Optogenetic suppression of the activity of 5-HT DRN neurons turned this antidepressant effect into a pro-depressant one: the time of immobility in the tail suspension test increased. The first minutes of the test were the most indicative, during minute 1 and 3 of which the DRN was illuminated with green light, and in animals of the eArchT3.0 × Ketamine group manifested the pro-depressant effect of the drug. During the second minute, when there was no illumination, this group did not differ from the control group. By the fourth and fifth minutes of the test, the antidepressant effect of ketamine in the Venus × Ketamine group was leveled, and in animals of the eArchT3.0 × Ketamine group the pro-depressant effect of optogenetic stimulation persisted (Fig. 2).

Changes in the immobility time during each of the 5 min tail suspension test during optogenetic suppression of the activity of 5-HT DRN neurons against the background of preliminary administration of ketamine. Ketamine (15 mg/kg) administered 1 h before the test reduced the immobility time in animals with the control vector, but increased it in animals with the eArchT3.0 expressing vector. During 1, 3, 4, and 5 min of the test, photosensitive animals of the eArchT3.0 × Ketamine group significantly outperformed the control Venus × Ketamine rats for the duration of immobility (n = 4 (3 in the eArchT3.0 × Ketamine group), n = number of animals in the group. *, p from < 0.05 to < 0.001).
The duration of immobility in each minute of the test increased with its duration (min of the test: F(4, 44) = 15.6739, p = 0.000). The effect of ketamine also depended on the duration of the test (Ketamine x Min F(4, 44) = 3.683, p = 0.011).
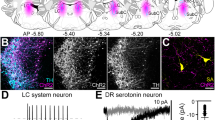
Two weeks after the injection of the vectors, 90.72 ± 1.13% of the neurons expressing Venus and 94.05 ± 1.03% of neurons expressing eArchT3.0-eYFP were also TPH2 immunopositive. Therefore, the vectors were specifically expressed in 5-HT neurons (Fig. 3). In addition, 42.46 ± 2.16% of TPH2 positive neurons expressed Venus and 41.64 ± 2.12% expressed eArchT3.0-eYFP.

Expression of c-Fos and vectors in tryptophan hydroxylase-2 (TPH2)-expressing neurons after optogenetic inhibition of 5-HT DRN neurons in the tail suspension test after pre-administration of a subanesthetic dose of ketamine. The vectors were specifically expressed in 5-HT neurons. Ketamine increased c-Fos expression in DRN neurons, and optogenetic suppression of 5-TH neuron activity prevented this increase. Scale, 100 µm.
In control vector animals, ketamine increased the percentage of neurons expressing c-Fos among cells expressing the Venus vector. However, in animals with a vector encoding a photosensitive protein, the expression of c-Fos in cells immunopositive for eArchT3.0-eYFP did not change after administration of ketamine and photostimulation (Fig. 4a).

Changes in the expression of the early response protein c-Fos in DRN neurons after the tail suspension test after a preliminary administration of a subanesthetic dose of ketamine: (a), changes in c-Fos expression in DRN neurons expressing the eArchT3.0-eYFP optogenetic vector or the Venus control vector; (b), changes in c-Fos expression in TPH2-expressing DRN neurons; (c), change in the number of DRN neurons expressing only c-Fos, but not the vector and/or TPH2. n = 4 (3 in the group eArchT3.0 × Ketamine; n = number of animals per group; *, p < 0.05 compared to the Venus × saline group).
The number of co-expressing c-Fos + TPH2 neurons was increased in animals that received ketamine and were not sensitive to light compared to all other groups. However, in animals in which the activity of 5‑HT DRN neurons was optogenetically suppressed, ketamine did not cause an increase in the number of co-expressing c-Fos + TPH2 neurons (Fig. 4b).
However, ketamine increased the number of neurons in DRN control animals that expressed only c-Fos but not the vector and/or TPH2. However, in animals in which the activity of 5-HT neurons was optogenetically suppressed, such an increase did not occur (Fig. 4c). At the same time, in animals injected with saline, optogenetic suppression of 5-HT neuron activity did not alter c-Fos expression in the DRN.
DISCUSSION
Over the past 20 years, ketamine has attracted much attention due to its rapid antidepressant action after a single subanesthetic dose in patients with treatment-resistant depression [2, 3]. In animal models, this drug has been shown to reduce depression-like behavior and increase AMPAR activity, phosphorylated mTOR levels, and BDNF expression [7]. In addition, many studies have emphasized the involvement of 5-HT neurotransmission in the mechanism of action of ketamine [12, 14–16].
In fact, in our experiment, a subanesthetic dose of ketamine was expected to reduce depression-like behavior in animals with functionally active 5-HT DRN neurons during the tail suspension test. In contrast, in animals with optogenetically inhibited activity of these neurons, an increase in the time of immobility was observed compared to control groups. In other studies, pharmacological blockade of 5-HT synthesis with parachlorophenylalanine [19, 20] prevented the antidepressant effect of ketamine in the forced swim test, but in our study, short-term suppression of the activity of serotonergic neurons using optogenetics led to the transformation of the effect of ketamine into a prodepressant and/or sedative effect. It is important to note that some cumulative effect of optogenetic suppression of 5-HT neuronal activity was observed with the duration of the tail suspension test in animals with preliminary administration of ketamine, which may be related to the kinetics of archearhodopsin, which is a proton pump [29], and/or the lighting regimen.
Systemic administration of ketamine activates glutamatergic projections from the PFC to the DRN [10]. AMPARs in DRNs are located on both 5-HT and GABAergic neurons [30]. 5-HT- and GABAergic neurons in the DRN receive signals both from each other and from other areas of the brain, both excitatory and inhibitory [30]. The final effect on neuronal activity obviously depends on the balance of excitatory and inhibitory signals. At the same time, it is not clear whether glutamatergic mPFC projections directly affect 5-HT neurons or GABAergic interneurons in the DRN. It is also possible that not all, but only a certain group of 5-HT neurons in the DRN, which have projections in the mPFC, are involved in response to ketamine [12]. This may be the reason that the indiscriminate inhibition of the activity of 5-HT neurons in the DRN in our experiments could lead to the conversion of the antidepressant effect of ketamine into a sedative effect.
In addition, the optogenetic effect prevented the ketamine-induced increase in expression of the early response protein c-Fos. It has already been shown that ketamine increases c-Fos expression in 5-HT DRN neurons [31, 32]; however, in our study, it was found that an increase in c-Fos expression after ketamine administration occurs not only in 5-HT neurons but also in neurons that do not express TPH2. Moreover, only about 42% of 5-HT neurons expressed the photosensitive protein eArchT3.0-eYFP, but suppression of the activity of this fraction of neurons was sufficient to prevent the ketamine-induced increase in c-Fos expression not only in vector-expressing cells, but also in other DRN neurons. Ketamine has previously been shown to have no effect on 5-HT release or 5-HT1A autoreceptor activity in the DRN [33]. At the same time, an increase in the number of c-Fos-positive DRN neurons that are not 5-HT neurons may indicate that interactions within the DRN between 5-HT and GABAergic neurons are involved in the mechanism of action of ketamine, which deserves attention and requires further study.
One important issue is that the behavior in the tail suspension test and the expression of c-Fos in the DRN do not change after optogenetic suppression of the activity of 5-HT neurons in this nucleus in animals injected with saline. The optogenetic effect that inhibited neuronal activity reduced the expression of c-Fos in eArchT3.0-eYFP-positive 5-HT DRN neurons in anesthetized animals [34], but this effect was not observed in actively awake animals.
Previously, another group of researchers found that inhibition of 5-HT DRN neurons with the same vector as in our study had no effect on immobility time in the forced swim test. Blockade of the activity of these neurons increased anxiety in rats in the elevated plus maze test but not in the open field test [24]. In contrast, optogenetic activation of 5-HT DRN neurons [24] or excitatory mPFC-DRN projections [16] induced an antidepressant-like effect in the tail suspension test. However, the inhibition of these projections also caused an antidepressant effect in the social injury model, which the authors associated with the involvement of GABAergic neurons in response [17]. Some inconsistency in the data is obviously associated with the peculiarities of behavioral patterns that cause avoidance or freezing reactions depending on the test conditions. Complex interactions in the network of glutamate, GABA, and 5-HT neurons involved in modulating the behavioral response to ketamine [35] may also contribute to the variability of results.
It is known that the discharge activity of DRN neurons varies depending on changes in behavior in the tail suspension test [36]. DRN neurons expressing the 5-HT transporter were active when an animal was in the home cage, whether moving or not, but during the tail suspension test the discharge activity of these neurons was high in mobile states and low in immobile states. At the same time, some parvalbumin-expressing GABAergic DRN neurons are active during periods of immobility, while the other part is inactive [37]. In other words, there are complex neural networks that involve subtypes of local inhibitory neurons that are involved in regulating the balance of excitation and inhibition in the DRN that are still poorly understood. Our results indicate that optogenetic downregulation of 5-HT DRN neuron activity alters the effect of ketamine on tail suspension behavior and c-Fos protein expression, but does not affect these measures in control animals, suggesting the likely fine regulation of complex interactions between DRN neurons in providing responses to situations that cause manifestations of struggle or despair.
CONCLUSIONS
This work showed the key role of 5-HT activity in DRN neurons in the rapid antidepressant effect of ketamine and the potential involvement of neurons of other DRN neurotransmitter systems in this effect for the first time. The reversal of the antidepressant action of a subanesthetic dose of the drug into a sedative one in blockade of the activity of 5-HT neurons qualitatively expands the range of possible effects of ketamine and requires further study. These findings add to the current understanding of the mechanism of action of ketamine, which is needed to develop the next generation of more effective antidepressants.
REFERENCES
Shishkina, G.T. and Dygalo, N.N., Zhurnal Vysshei Nervnoi Deiatelnosti Imeni I.P. Pavlova, 2010, vol. 60, no. 2, pp. 138–152.
Liu, B., Liu, J., Wang, M., Zhang, Y., and Li, L., Frontiers in Cellular Neuroscience, 2017, vol. 11, p. 305.
Pham, T.H. and Gardier, A.M., Pharmacology & Therapeutics, 2019, vol. 199, pp. 58–90.
Krystal, J.H., Abdallah, C.G., Sanacora, G., Charney, D.S., and Duman, R.S., Neuron, 2019, vol. 101, no. 5, pp. 774–778.
Yang, C., Yang, J., Luo, A., and Hashimoto, K., Translational Psychiatry, 2019, vol. 9, no. 1, p. 280.
Zanos, P., Moaddel, R., Morris, P.J., Riggs, L.M., Highland, J.N., Georgiou, P., Pereira, E.F.R., Albuquerque, E.X., Thomas, C.J., and Zarate, Jr.C.A., Pharmacological Reviews, 2018, vol. 70, no. 3, pp. 621—660.
Miller, O.H., Moran, J.T., and Hall, B.J., Neuropharmacology, 2016, vol. 100, pp. 17–26.
Nosyreva, E., Szabla, K., Autry, A.E., Ryazanov, A.G., Monteggia, L.M., and Kavalali, E.T., J. Neuroscience, 2013, vol. 33, no. 16, pp. 6990–7002.
Gerhard, D.M., Pothula, S., Liu, R.J., Wu, M., Li, X.Y., Girgenti, M.J., Taylor, S.R., Duman, C.H., Delpire, E., Picciotto, M., Eric, S., Wohleb, E.S., and Duman, R.S., The Journal of Clinical Investigation, 2020, vol. 130, no. 3, pp. 1336–1349.
Fukumoto, K., Iijima, M., Funakoshi, T., and Chaki, S., Neuropharmacology, 2018, vol. 137, pp. 96–103.
Ago, Y., Tanabe, W., Higuchi, M., Tsukada, S., Tanaka, T., Yamaguchi, T., Igarashi, H., Yokoyama, R., Seiriki, K., Kasai, A., Nakazawa, T., Nakagawa, S., Hashimoto, K., and Hashimoto, H., Int. J. Neuropsychopharmacology, 2019, vol. 22, no. 10, pp. 665–674.
Lopez-Gil, X., Jimenez-Sanchez, L., Campa, L., Castro, E., Frago, C., and Adell, A., ACS Chemical Neuroscience, 2019, vol. 10, no. 7, pp. 3318–3326.
Tiger, M., Veldman, E.R., Ekman, C.J., Halldin, C., Svenningsson, P., and Lundberg, J., Translational Psychiatry, 2020, vol. 10, no. 1, p. 159.
Du, JardinK.G., Liebenberg, N., Cajina, M., Muller, H.K., Elfving, B., Sanchez, C., and Wegener, G., Frontiers in Pharmacology, 2018, vol. 8, p. 978.
Nishitani, N., Nagayasu, K., Asaoka, N., Yamashiro, M., Shirakawa, H., Nakagawa, T., and Kaneko, S., Int. J. Neuropsychopharmacology, 2014, vol. 17, no. 8, pp. 1321–1326.
Warden, M.R., Selimbeyoglu, A., Mirzabekov, J.J., Lo, M., Thompson, K.R., Kim, S.Y., Adhikari, A., Tye, K.M., Frank, L.M., and Deisseroth, K., Nature, 2012, vol. 492, pp. 428–432.
Challis, C., Beck, S.G., and Berton, O., Frontiers in Behavioral Neuroscience, 2014, vol. 8, p. 43.
Kinoshita, H., Nishitani, N., Nagai, Y., Andoh, C., Asaoka, N., Kawai, H., Shibui, N., Nagayasu, K., Shirakawa, H., Nakagawa, T., and Kaneko, S., Int. J. Neuropsychopharmacology, 2018, vol. 21, no. 3, pp. 305–310.
Fukumoto, K., Iijima, M., and Chaki, S., Psychopharmacology, 2014, vol. 231, no. 11, pp. 2291–2298.
Du, JardinK.G., Liebenberg, N., Muller, H.K., Elfving, B., Sanchez, C., and Wegener, G., Psychopharmacology, 2016, vol. 233, pp. 2813–2825.
Gigliucci, V., O’Dowd, G., Casey, S., Egan, D., Gibney, S., and Harkin, A., Psychopharmacology, 2013, vol. 228, no. 1, pp. 157–166.
Fakhoury, M., Progress in Neuro-Psychopharmacology and Biological Psychiatry, 2021, vol. 106, p. 110094.
Nishitani, N., Nagayasu, K., Asaoka, N., Yamashiro, M., Andoh, C., Nagai, Y., Kinoshita, H., Kawai, H., Shibui, N., Liu, B., Hewinson, J., Shirakawa, H., Nakagawa, T., Hashimoto, H., Kasparov, S., and Kaneko, S., Neuropsychopharmacology, 2019, vol. 44, no. 4, p. 721.
McClure, C., Cole, K.L., Wulff, P., Klugmann, M., and Murray, A.J., JoVE (Journal of Visualized Experiments), 2011, no. 57.
Shaburova, E.V. and Lanshakov, D.A., Biotekhnologiya, 2020, vol. 36, no. 5, pp. 89–97.
Paxinos, G. and Watson, C., The Rat Brain in Stereotaxic Coordinates, Academic Press, 1998.
Lanshakov, D.A., Sukhareva, E.V., Kalinina, T.S., and Dygalo, N.N., Neurobiol. Dis., 2016, vol. 91, pp. 1–9.
Bankhead, P., Loughrey, M.B., Fernandez, J.A., Dombrowski, Y., McArt, D.G., Dunne, P.D., McQuaid, S., Gray, R.T., Murray, L.J., Coleman, H.G., James, J.A., Salto-Tellez, M., and Hamilton, P.W., Sci. Rep., 2017, vol. 7, no. 1, pp. 1–7.
Krol, A., Lopez-Huerta, V.G., Corey, T.E., Deisseroth, K., Ting, J.T., and Feng, G., Frontiers in Neural Circuits, 2019, vol. 13, p. 4.
Pham, T.H., Mendez-David, I., Defaix, C., Guiard, B.P., Tritschler, L., David, D.J., and Gardier, A.M., Neuropharmacology, 2017, vol. 112, pp. 198–209.
Fukumoto, K., Iijima, M., and Chaki, S., Neuropsychopharmacology, 2016, vol. 41, no. 4, pp. 1046–1056.
Dolzani, S.D., Baratta, M.V., Moss, J.M., Leslie, N.L., Tilden, S.G., Sorensen, A.T., Watkins, L.R., Lin, Y., and Maier, S.F., Eneuro, 2018, vol. 5, no. 1.
Hernandez-Vazquez, F., Garduno, J., and Hernandez-Lopez, S., Reviews in the Neurosciences, 2019, vol. 30, no. 3, pp. 289—303.
Drozd, U.S. and Lanshakov, D.A., Integrativnaya Fiziologiya, 2020, vol. 1, no. 2, pp. 144–146.
Li, Y.F., Pharmacology & Therapeutics, 2020, vol. 208, p. 107494.
Seo, C., Guru, A., Jin, M., Ito, B., Sleezer, B.J., Ho, Y.Y., Wang, E., Boada, C., Krupa, N.A., Kullakanda, D.S., and Shen, C.X., Science, 2019, vol. 363, no. 6426, pp. 538–542.
Zhou, L., Liu, D., Xie, Z., Deng, D., Shi, G., Zhao, J., Bai, S., Yang, L., Zhang, R., and Shi, Y., Frontiers in Behavioral Neuroscience, 2022, vol. 16, p. 200.
ACKNOWLEDGMENTS
The authors are grateful to Professor Naoya Nishitani, Graduate School of Pharmaceutical Sciences, Kyoto, Japan, for kindly providing the TPH2-eArchT3.0-eYFP-WPRE and TPH2-Venus-WPRE plasmids.
Funding
The work was supported by the budget project FWNR-2022-0002.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest. The authors declare that they have no conflicts of interest.
Ethical approval. All procedures were carried out in accordance with the International European Bioethical Standards (86/609-EEC), Russian Rules for Working on Laboratory Animals (N 267 19.06.2003) and were approved by the Bioethical Committee of the Institute of Cytology and Genetics of the Siberian Branch of the Russian Academy of Sciences.
Additional information
Corresponding author; address: pr. Ak. Lavrentiev 10, Novosibirsk, 630090 Russia; e-mail: drozd@bionet.nsc.ru.
Rights and permissions
Open Access. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Drozd, U.S., Lanshakov, D.A. & Dygalo, N.N. Dorsal Raphe Nucleus Serotonergic Neuron Activity is Necessary for the Manifestation of the Antidepressant Effect of Ketamine. Neurochem. J. 17, 394–401 (2023). https://doi.org/10.1134/S181971242303008X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S181971242303008X