
Abstract
Worm-like endosymbionts were found in the hepatic region of the digestive tract of the deep-sea acorn worm Quatuoralisia malakhovi Ezhova et Lukinykh, 2022 (family Torquaratoridae) from the Bering Sea. The symbionts were assigned to the taxon Nemertodermatida on the basis of histological examination. Torquaratoridae are similar in feeding type to holothuroids, which have also been found to have Xenacoelomorpha endosymbionts.
Similar content being viewed by others

The deep-sea acorn worms Torquaratoridae have recently been discovered as a group of marine invertebrates [1–5]. These animals live at depths of 350–8800 m [3, 4, 6] and do not bury in the seafloor in contrast to shallow-water acorn worms. A high population density of Torquaratoridae has been observed in certain oceanic regions. For example, their population density reaches 12 acorn worms per square meter in the Bering Sea at depths of approximately 2000 m [7]. Given this population density, Torquaratoridae certainly play an important role in the function of bathyal and abyssal communities.
The biology of Torquaratoridae is poorly understood. In particular, little is known about their symbionts. Acoelic flatworms and parasitic copepods have been mentioned as gut inhabitants of Torquarator bullocki Holland, Clague, Gordon, Gebruk, Pawson and Vecchione, 2005, in the first publication about Torquaratoridae, but without any description or illustration [1]. A white process on the outer side of the genital wings has been observed in an underwater image of Allapasus isidis Priede, Osborn, Gebruk, Jones, Shale, Rogacheva and Holland, 2012, and is possibly a leech attached to the worm [4]. Trematode metacercariae have been found in all divisions of the coelom in Quatuoralisia malakhovi Ezhova et Lukinykh, 2022 [8].
Here we describe the Nemertodermatida sym-bionts found in the deep-see acorn worms Torquaratoridae.
Material for the study was collected on June 18, 2018, during the 82nd expedition onboard the Akademik M.A. Lavrent’ev research ship. Trawling was performed at station LV 82-9 (55.3451–55.3466° N, 167.2750–167.2752° E) in the Komandorsky Basin (the Volcanologists Massif) of the Bering Sea at depths of 1957–1933 m. Quatuoralisia malakhovi individuals were fixed with 8% formalin in seawater for histological examination. Specimens were washed to remove the fixative and dehydrated with increasing ethanol concentrations by standard methods. Fragments prepared for histological examination were embedded in Paraplast, and the resulting blocks were used to obtain series of transverse and sagittal 10-µm histological sections with a Leica RM 2125 microtome. The sections were stained with Carracci hematoxylin and eosin (an ethanol solution). Images of the sections were obtained using a Micmed-6 microscope (LOMO, St. Petersburg, Russia, 2018) with a MS-12 digital camera. Five worms were examined, and symbionts were found in two of them.
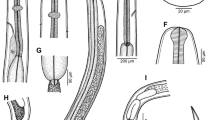
The infected Q. malakhovi specimens were females and carried a single symbiont each. The symbionts were found in the gut lumen in the anterior part of the Q. malakhovi hepatic region. The symbionts appeared elongate leaf-shaped in sections (Figs. 1a, 1b). One symbiont was approximately 900 µm long and 400 µm wide in the widest part of its body; the dimensions of the other symbiont were 1 mm in length and approximately 870 µm in width. The symbionts were in contact with folds of the Q. malakhovi gut epithelium.

Histological sections of the symbionts within the hepatic region of Quatuoralisia malakhovi: (a) a longitudinal section with the mouth (mo) and the pharynx (ph), (b) a longitudinal section with gonia (square frames), (c) part of a transverse section of the anterior end with a presumable statocyst (circular frame), and (d) part of a longitudinal section with the genital atrium (ga). Bar, (a, b, d) 100 or (c) 20 μm. Designations: cp, central parenchyma of the symbiont; ep, epidermal epithelium of the symbiont; ga, genital atrium; gd, gastrodermal epithelium of the host; lum, gut lumen of the host; mo, mouth; ph, pharynx; sel, subepidermal layer of the symbiont; sfg, secret of the frontal glands.
A vacuolated ciliated epithelium of 20 µm in height coats the symbionts (Figs. 1a, 1b; ep). A basal lamina underlying the coating epithelium was not detected. A subepidermal layer occurs under the coating epithelium and consists of small cells (Figs. 1a, 1b; sel). The height of the subepidermal layer varies from 50 to 80 µm. The central part of the body is occupied by large vacuolated cells of the central parenchyma (Figs. 1a, 1b; cp). A mouth opening is 100 µm wide, occurs in the middle of the body (Fig. 1a, mo), and leads to a cylindrical pharynx, which is 130 µm deep (Fig. 1a, ph). A statocyst is found at the putative anterior end of the symbiont (Fig. 1c, circle framed). The statocyst is an oval body of 15 µm in diameter and hangs on radial ligaments. Statoliths were not preserved. However, two flattened nuclei are found in the center of the statocyst, presumably corresponding to the nuclei of two lithocytes. Secretion of frontal glands is detectable at the anterior end (Fig. 1c, sfg). A genital atrium opens subterminally at the opposite, posterior end of the body (Fig. 1d, ga). A dark bundle extends from its opening into the inside of the body and is adjacent to the central parenchyma. Separate large cells with a large nucleus and a dark cytoplasm are found at the boundary of the subepidermal layer and the central parenchyma (Fig. 1b, square framed). The cells can be assumed to be gonia, which are precursors to gametes. Structures that could be assigned to the excretory system were not found in the histological sections of the symbionts. Likewise, nerve and muscle fibers were not distinguished in the sections stained with hematoxylin and eosin.
The main organization features (lack of a basal lamina of the coating epithelium, the positions of the mouth and genital atrium, lack of an excretory system, lack of structured gonads, the oogonia occurring at the boundary the subepidermal layer and the central parenchyma, and a statocyst with two lithocytes) allow us to assign the symbionts to the group Nemertodermatida, which belongs to Xenacoelomorpha together with Acoela and Xenoturbellida. Acoela and Nemertodermatida are considered to be sister groups that form the clade Acoelomorpha [9–12].
Various holothurians mostly predominate in the macrozoobenthos community of the slopes of the Volcanologists Massif at depths of 1370–4278 m [7]. However, the torquaratorid Q. malakhovi displaces holothurians from their dominant position at depths of 1830−2290 m [7, 13]. The ecological niches of Torquaratoridae and Holothuroidea overlap because both are detritus feeders [7, 14–16]. It is of interest that shallow-water holothurians are also known to have symbionts that belong to Acoelomorpha and inhabit their digestive tract [17, 18]. For example, Aechmalotus pyrula Beklemischev, 1915, which represents Acoela, lives in the digestive tract and respiratory trees of the holothurian Eupyrgus scaber Lütken, 1857, from the Barents Sea. The taxon Acoela also includes the members of the genus Aphanostoma Ørsted, 1845, that have been found in the digestive tract of the holothurians Myriotrochus rinkii Steenstrup, 1851 and Chiridota laevis (O. Fabricius, 1780) from the Barents Sea [18]. Meara stichopi Westblad, 1949, which belongs to Nemertodermatida, has been found in the digestive tract and coelomic cavities of the holothurians Mesothuria intestinalis (Ascanius, 1805) and Parastichopus tremulus (Gunnerus, 1767) from the North Sea [17, 19]. Thus, the presence of endosymbionts belonging to related groups reflects the similarity in ecological niches between Holothuroidea and Torquaratoridae.
REFERENCES
Holland, N.D., Clague, D.A., Gordon, D.P., et al., “Lophenteropneust” hypothesis refuted by collection and photos of new deep-sea hemichordates, Nature, 2005, vol. 434, pp. 374–376.
Osborn, K.J., Kuhnz, L.A., Priede, I.G., et al., Diversification of acorn worms (Hemichordata, Enteropneusta) revealed in the deep sea, Proc. R. Soc. B, 2012, vol. 279, pp. 1646–1654.
Osborn, K.J., Gebruk, A.V., Rogacheva, A., et al., An externally brooding acorn worm (Hemichordata, Enteropneusta, Torquaratoridae) from the Russian Arctic, Biol. Bull., 2013, vol. 225. pp. 113–123.
Priede, I.G., Osborn, K.J., Gebruk, A.V., et al., Observations on torquaratorid acorn worms (Hemichordata, Enteropneusta) from the North Atlantic with descriptions of a new genus and three new species, Invertebr. Biol., 2012, vol. 131, no. 3, pp. 244–257.
Holland, N.D., Hiley, A.S., and Rouse, G.W., A new species of deep-sea torquaratorid enteropneust (Hemichordata): A sequential hermaphrodite with exceptionally wide lips, Invertebr. Biol., 2022, vol. 141, p. e12379.
Smith, K.L.J., Holland, N.D., and Ruhl, H.A., Enteropneust production of spiral fecal trails on the deep-sea floor observed with time-lapse photography, Deep Sea Res., Part I, 2005, vol. 52, pp. 1228–1240.
Rybakova, E., Galkin, S., Gebruk, A., et al., Vertical distribution of megafauna on the Bering Sea slope based on ROV survey, PeerJ, 2020, vol. 8, p. e8628.
Ezhova, O.V., Belolyubskaya, K.I., Krupenko, D.Yu., et al., The first discovery of Trematodes (Digenea) in the deep-sea acorn worms Torquaratoridae (Hemichordata, Enteropneusta), Dokl. Biol. Sci., 2022, vol. 503, pp. 53–71.
Ehlers, U., Comparative morphology of statocysts in the Plathelminthes and the Xenoturbellida, Hydrobiologia, 1991, vol. 227, pp. 263–271.
Zhang, Z.-Q., Hooper, J., Soest, R., et al., Animal biodiversity: An outline of higher-level classification and taxonomic richness, Zootaxa, 2011, vol. 3148, pp. 24–25.
Brusca, R.C., Moore, W., and Shustes, S.M., Invertebrates. Introduction to the Bilateria and the Phylum Xenacoelomorpha: Triploblasty and Bilateral Symmetry Provide New Avenues for Animal Radiation, Sinauer Associates, 2016, part 9, pp. 345–372.
Haszprunar, G., Review of data for a morphological look on Xenacoelomorpha (Bilateria incertae sedis), Org. Diversity Evol., 2016, vol. 16, pp. 363–389.
Ezhova, O.V., Lukinykh, A.I., Galkin, S.V., et al., Deep-sea acorn worms (Enteropneusta) from the Bering Sea with the description of a new genus and a new species of Torquaratoridae dominating soft-bottom communities, Deep Sea Res., Part II, 2022, vol. 195, p. 105014.
Ezhova, O.V., Trukhan, M.A., Lukinykh, A.I., et al., Digestive system and feeding of deep-sea acorn worm Quatuoralisia malakhovi (Enteropneusta: Torquaratoridae) from the Bering Sea, Deep Sea Res., Part I, 2023 (In press).
Rodkina, S.A., The fatty acid composition of the deep-sea acorn worm Quatuoralisia malakhovi Ezhova et Lukinykh, 2022 (Hemichordata, Enteropneusta), Russ. J. Mar. Biol., 2023, vol. 49, no. 4, pp. 326–329.
Rodkina, S.A., Kiyashko, S.I., and Mordukhovich, V.V., Diet of deep-sea holothurians in the Volcanologists Massif, Bering Sea, as inferred from stable isotope and fatty acid analyses, Deep Sea Res., Part II, 2023, p. 105266.
Jangoux, M., Diseases of Echinodermata. II. Agents metazoans (Mesozoa to Bryozoa), Dis. Aquat. Org., 1987, vol. 2, pp. 205–234.
Beklemishev, V.N., On parasitic turbellarians from the Murmansk coast. I. Acoela, Tr. Petrogr. O-va. Estestvoispyt., 1916, pp. 103–172.
Westblad, E., On Meara stichopi (Bock) Westblad, a new representative of Turbellaria Archoophora, Ark. Zool., 1949, vol. 1, pp. 43–57.
ACKNOWLEDGMENTS
We are grateful to staff of the Zhirmunskii National Scientific Center of Marine Biology, Far Eastern Branch of Russian Academy of Sciences (NSCMB FEB RAS, Vladivostok) for providing the material for the study and E.M. Krylova, S.V. Galkin, and A.V. Gebruk (Shirshov Institute of Oceanology, RAS) for help in transferring the study material. Histology was performed at the Student Laboratory of Evolutionary Morphology of Animals (www.evolmorphan.ru) of the Invertebrate Zoology Department (Biological Faculty, Moscow State University).
Funding
This work was supported by the Russian Science Foundation (project no. 23-24-00066).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. In accordance with Directive 2010/63/EU of September 22, 2010 on the protection of animals used for scientific purposes, Chapter 1, Paragraph 3, the requirements of bioethics do not apply to the object of this study.
CONFLICT OF INTEREST
The authors of this work declare that they have no conflicts of interest.
Additional information
Translated by T. Tkacheva
Publisher’s Note.
Pleiades Publishing remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Ezhova, O.V., Lukinykh, A.I. & Malakhov, V.V. Nemertodermatida—Endosymbionts of Deep-Sea Acorn Worms (Hemichordata, Torquaratoridae). Dokl Biol Sci 515, 11–14 (2024). https://doi.org/10.1134/S001249662360015X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S001249662360015X