
Abstract
A pyramid strategy combining the Cry1A and Cry2A toxins in Bt crops has been widely used throughout the world to delay pest adaption to transgenic crops and broaden the insecticidal spectrum. Midgut membrane-bound cadherin (CAD), aminopeptidase-N (APN) and alkaline phosphatase (ALP) are important for Cry1A toxicity in some lepidopteran larvae, but the proteins that bind Cry2A in the midgut of target insects and their role in the Cry2A mechanism of action are still unclear. In this study, we found that heterologously expressed CAD, APN4 and ALP2 peptides from the midgut of Helicoverpa armigera could bind to the Cry2Aa toxin with a high affinity. Additionally, the efficiency of Cry2Aa insecticidal activity against H. armigera larvae was obviously reduced after the genes encoding these proteins were silenced with specific siRNAs: CAD- and ALP2-silenced larvae showed significantly similar reductions in mortality due to the Cry2Aa toxin (41.67% and 43.06%, respectively), whereas a larger reduction in mortality was observed in APN4-silenced larvae (61.11%) than in controls. These results suggest that CAD, APN4 and ALP2 are involved in the mechanism of action of Cry2Aa in H. armigera and may play important functional roles in the toxicity of the Cry2Aa toxin.
Similar content being viewed by others

Introduction
Selected cry genes encoding insecticidal crystal proteins from the soil bacterium Bacillus thuringiensis (Bt) have been successfully transferred to genetically modified (GM) crops to produce toxic proteins because these proteins show specific toxicity to certain insect pests, such as Lepidoptera, Diptera, and Coleoptera, but do not harm non-target organisms1,2,3,4,5. In addition to their economic benefits, GM crops also have clearly delivered substantial agricultural and environmental benefits to farmers and society. Therefore, the global cultivated acreage of GM crops increased rapidly following their first commercialization in 1996, covering 1.7 million hectares, to 179.7 million hectares in 2015, and Bt crops, such as Bt cotton and Bt maize, lead the world in the adoption of GM crops, covering more than 78 million hectares6,7,8,9.
However, similar to the history of synthetic pesticides, long-term application of Bt crops and Bt spray products exerts high selection pressure on their target insects, resulting in some insects evolving resistance to Bt insecticidal proteins. As a result, the durability and effectiveness of Bt crops have diminished10,11,12. Thus, a pyramid strategy in which at least two sufficiently different Bt toxins are expressed in a single plant was proposed for use in resistance management programs. Two-toxin crops are generally expected to improve the efficacy of Bt, broaden the insecticidal spectrum, delay the evolution of target pest resistance and increase the duration of benefits from transgenic crops13. For example, two-toxin Bt cotton, in which both the Cry1A and Cry2A toxins are produced, has been widely used in many countries to replace first-generation Bt cotton, producing only the Cry1A toxin. However, only first-generation Bt cotton has been allowed for commercial production in China since 199614,15.
The cotton bollworm, Helicoverpa armigera (Hübner; Lepidoptera:Noctuidae), is a major polyphagous pest of cotton and other crops in China, and the introduction of Bt cotton expressing Cry1Ac has effectively suppressed regional outbreaks of H. armigera, not only on cotton but also on other host crops16,17. However, the discovery of resistant H. armigera populations under laboratory and field conditions may signal an early warning for more serious problems, such as control failure of Bt cotton18,19,20,21. Cry1Ac and Cry2Ab are considered to represent a good combination of Bt toxins because they show low amino acid homology and might ideally target different binding sites in the larval midgut22,23. In contrast with Cry1A proteins, Cry2A toxins can also effectively control lepidopteran pests other than H. armigera, such as Spodoptera exigua and Agrotis ypsilon24. Consequently, introduction of Cry1A/Cry2A Bt cotton in China is expected to provide more effective control activity against many target lepidopteran pests and assist in delaying the development of insect resistance to Bt toxic proteins25,26.
After being ingested by susceptible lepidopteran larvae, the Cry protoxin is solubilized and activated in the midgut. It then goes through a complex series of interactions with midgut membrane proteins, finally leading to cell death and killing the larvae27,28. Several membrane proteins have been characterized as functional receptors for the Cry1A toxin, including cadherin (CAD), aminopeptidase-N (APN), alkaline phosphatase (ALP) and ATP-binding cassette (ABC) transporter protein29,30,31, whereas the binding receptors of the Cry2A toxin and their potential roles in the mode of action of the Cry2A protein remain incompletely characterized. To determine the roles of CAD, APN and ALP from the midgut of H. armigera in the Cry2Aa toxin mechanism of action, we studied the binding characteristics of the Cry2Aa toxin with recombinant H. armigera CAD, APN4 (a cluster of lepidopteran APNs) and ALP2 (a cluster of lepidopteran ALPs) peptides32,33. Then, we investigated their roles in Cry2Aa toxicity by suppressing their transcription with siRNAs. Based on our results, we hypothesize that CAD, APN4 and ALP2 may function as receptors for the Cry2Aa toxin in H. armigera and play important functional roles in the mechanism of action of the Cry2Aa toxin.
Results
Toxicity of Cry1Ac and Cry2Aa to the susceptible H. armigera strain
The results of insect assays showed that the Cry1Ac and Cry2Aa toxins exhibited similar toxicity against first-instar H. armigera larvae. The LC50 values of these two toxins were 1.20 μg/cm2 (95% fiducial limits, 0.79–1.92) and 1.05 μg/cm2(95% fiducial limits, 0.59–1.91), respectively.
Expression and purification of HaCAD, HaAPN4 and HaALP2 and determination of polyclonal antibody specificity
To further characterize the binding of CAD, APN4 and ALP2 from H. armigera to the Cry2Aa toxin, three fragments of these proteins were expressed in E. coli BL21 (DE3) cells and purified from the total crude proteins. The expressed peptides were evaluated through SDS-PAGE, and the results demonstrated that the expressed CAD, APN4 and ALP2 proteins exhibited the expected sizes of 47 kDa, 40 kDa and 51 kDa, respectively, and that there was only one main visible band for these three proteins after purification (Fig. 1, lane 1, lane 2 and lane 3). The purified proteins were detected with the corresponding antibodies against HaCAD, HaAPN4 and HaALP2, and the results indicated that all three purified recombinant proteins could strongly hybridize with the corresponding polyclonal antibodies (Fig. 1, lane 4, lane 5 and lane 6).

Lane M: protein marker; lanes 1–3: Purified recombinant CAD, APN4 and ALP2 proteins; lanes 4–6: The purified CAD, APN4 and ALP2 fragments were detected with the corresponding polyclonal antibodies.
Western and ligand blotting detection
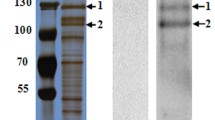
The CAD, APN4 and ALP2 proteins in brush border membrane vesicle (BBMV) samples obtained from the midguts of H. armigera were detected by western blotting using anti-HaCAD, anti-HaAPN4 and anti-HaALP2, and the results revealed an apparent size for the CAD protein of 210 kDa, for the APN4 protein of 120 kDa and for the ALP2 protein of 65 kDa in the BBMV samples (Fig. 2A, lane 1, lane 2 and lane 3). To determine the binding of the Cry2Aa toxin to the CAD, APN4 and ALP2 proteins from the midguts of H. armigera, ligand blotting analysis of the biotinylated Cry2Aa toxin was performed using BBMV protein or heterologously expressed recombinant HaCAD, HaAPN4 and HaALP2 proteins. The results showed that ligand blot could detect several binding bands, included binding proteins of 210, 120 and 65 kDa for Cry2Aa toxin in the BBMV proteins from susceptible cotton bollworm (Fig. 2B, lane 7), and all the three purified recombinant proteins could also bind to the Cry2Aa toxin (Fig. 2B, lane 4, lane 5 and lane 6).

Western blot analysis of anti-CAD, anti-APN4 and anti-ALP2 antibodies using BBMV proteins from H. armigera larvae (A) and ligand blot analysis of Cry2Aa with the recombinant CAD, APN4 and ALP2 proteins or BBMV proteins (B). Lane M: protein marker; lanes 1–3: CAD, APN4 and ALP2 antibody binding of BBMV from fifth-instar H. armigera larvae; lanes 4–6: Cry2Aa toxin binding by the recombinant CAD, APN4 and ALP2 proteins; lane 7: Cry2Aa binding with BBMV proteins.
ELISA binding assays of Cry2Aa with HaCAD, HaAPN4 and HaALP2
To further determine the specific binding characteristics of the Cry2Aa toxin in relation to the recombinantly expressed HaCAD, HaAPN4 and HaALP2 proteins, we tested their binding affinities through ELISA binding assays. The results showed that the Cry2Aa toxin bound to the recombinant HaCAD, HaAPN4 and HaALP2 proteins with high affinities of 12.28 nM, 36.82 nM and 22.91 nM, respectively (Fig. 3A–C).

Binding ELISAs were performed by fixing 1 μg of the CAD (A), APN4 (B) and ALP2 (C) fragments per well in ELISA plates, followed by incubation with different concentrations of the biotinylated Cry2Aa toxin. Kd values obtained through SigmaPlot 12.5 analysis are indicated within the graphs.
Effects of CAD, APN4 and ALP2 knockdown on their transcript levels and Cry2Aa-induced larval mortality in H. armigera
RNAi efficiency was investigated via qPCR, and the results showed that the transcription of HaCAD, HaAPN4 and HaALP2 in larvae fed diets containing HaCAD-, HaAPN4-or HaALP2-siRNA was significantly reduced by 48.33% (P = 0.002), 63.67% (P = 0.013) and 61.33% (P = 0.002), respectively, compared with that in larvae fed control diets with only RNase-free H2O or GFP-siRNA (Fig. 4A–C). The CAD- and ALP2-silenced larvae showed significantly similar reductions of the Cry2Aa toxin-induced mortality rate (41.67% (P = 0.004) and 43.06% (P < 0.001), respectively), whereas a greater reduction in mortality was observed in the APN4-silenced larvae (61.11%, P = 0.003) compared with the controls (Fig. 5A). The Cry1Ac toxin was used as a positive control, and the results showed that the most significant reduction of mortality was observed in APN4-silenced H. armigera larvae, which exhibited an 87.50% reduction (P < 0.001), followed by ALP2-silenced larvae (59.72% mortality reduction, P < 0.001) and CAD-silenced larvae (40.28% reduction, P < 0.001; Fig. 5B).

Transcript abundance was determined via qPCR. Bars represent the means and standard errors for 3 individual samples (A–C).

CAD-silenced, APN4-silenced, ALP2-silenced and control larvae were exposed to diets containing 8 μg/cm2 of the Cry2Aa (A) or Cry1Ac toxin (B), and mortality was recorded after 7 days. Bars represent the means and standard errors determined from 3 replicates. Different letters indicate a significant difference at a P value < 0.05.
Discussion
In the present study, the Cry2Aa and Cry1Ac toxins showed similar toxicity to the susceptible strain of H. armigera (for Cry2Aa, LC50 = 1.05 μg/cm2; for Cry1Ac, LC50 = 1.20 μg/cm2). In addition to H. armigera, Cry2A toxins (Cry2Aa and Cry2Ab) also have been shown to exhibit high efficiency against several other lepidopteran pests34,35,36,37. Therefore, the introduction of Cry1A/Cry2A Bt cotton in China is expected to be more effective against target pests and to broaden the insecticidal spectrum compared with first-generation Bt cotton.
A key step in the mechanism of action of Cry toxins in lepidopteran pests is the interaction of activated Cry proteins with specific binding sites in the BBMVs of the larval midgut, and reduced binding of Cry toxins to these membrane binding receptors is generally related to resistance of target pests to Bt toxins11,26,28,38,39,40. The insect midgut membrane protein CAD and the highly abundant glycosyl phosphatidylinositol (GPI)-anchored APN and ALP proteins have been characterized as functional binding receptors for Cry1A toxins in several lepidopteran insects28,41,42,43. However, limited studies have focused on defining the role of the insect membrane proteins CAD, APN and ALP in relation to Cry2A toxins to date44,45.
In lepidopteran pests, insect midgut CAD, APN and ALP proteins are important binding receptors for several classes of Cry1A toxins2,26,43,46,47,48. GPI-anchored APN and ALP proteins first bind to protease-activated Cry1A toxins with a low affinity (101 nM for APN, 267 nM for ALP) and subsequently carry the toxins to the microvilli membrane of the larval midgut, where the toxins bind to CAD with a high affinity (1 nM). Then, CAD promotes oligomerization of Cry toxins, and APN and ALP interact with the oligomeric Cry structure with a high affinity (0.6 nM for APN, 0.5 nM for ALP), which causes membrane insertion and pore formation, ultimately leading to osmotic lysis of midgut cells28,41,42. The results of ligand blot and western blot assays conducted in this study indicated that the 210, 120 and 65 KDa proteins from H. armigera larval midgut BBMV could recognize by Cry2Aa toxin and the antibodies that against CAD, APN and ALP, respectively. In addition, the Cry2Aa toxin could bind to all the three expressed recombinant proteins (HaCAD, HaAPN4 and HaALP2; Fig. 2B). The binding affinities of HaCAD and HaALP2 with Cry2Aa were 3.00-fold and 1.61-fold higher, respectively, than that of HaAPN4 (Fig. 3). However, the binding affinity results indicated much lower affinities than reported previously28,49,50. This difference may be due to the different toxin proteins examined or to the fact that the CAD, APN and ALP proteins used in this study were heterologously expressed as recombinant proteins. Previous reports of findings in H. armigera and H.virescens revealed that the binding of Cry proteins to APN and ALP relies on an N-acetylgalactosamine interaction51,52. However, in the present study, the expressed recombinant HaAPN4 and HaALP2 proteins from E. coli were not glycosylated, which explains the low binding affinity of the Cry2Aa toxin for the expressed recombinant APN and ALP proteins.
Cry1A and Cry2A are the two most widely used Cry toxins in pyramided transgenic cotton based on reports indicating that Cry1A and Cry2A exhibit different binding receptor proteins and different modes of action22,23,53,54. However, further research indicated that Domain II of Cry2A shows similar binding sites to Cry1A23,55, and some cross-resistance between these two toxin types has been documented. This finding implies that they may share a common receptor, although cross-resistance can be independent of toxins sharing receptors19,44,56,57,58. In the current study, after silencing the HaCAD, HaAPN4 and HaALP2 genes using specific siRNAs in vivo, CAD, APN4 and ALP2 transcript levels were found to be suppressed (Fig. 4). Compared with a positive control using the Cry1Ac toxin, a similar increase in the tolerance of H. armigera to Cry2Aa was observed (Fig. 5). Therefore, these three membrane-bound midgut proteins may play similar significant roles in the mechanism of action of the Cry2Aa toxin in H. armigera.
Recently, CAD was confirmed to be involved in the action of the Cry2Aa toxin in S. exigua, serving as a functional receptor for both the Cry2Aa and Cry1Ac toxins. These two toxins did not compete for the same binding sites, which suggests that they may bind to diverse CAD protein epitopes in this species45,59. It has also been shown that ALP2 plays a dominant role in the susceptibility of S. exigua to the Cry2Aa toxin60. Among the several clusters of APNs present in lepidopteran insects, APN1 has been reported as an important functional receptor for Cry1Ac in several lepidopteran species42,44,61, and APN3 and APN6 serve as functional candidate receptors for Cry1Ca in S. exigua62. However, there is no direct or conflict evidence of the roles of other APNs in the mechanisms of Cry1A, Cry2A or other Cry toxins46. In the present study, we also found that CAD and ALP played significant roles in the action of the Cry2Aa toxin, similar to those associated with the Cry1Ac toxin. Silencing of APN4 in H. armigera had less of an effect on the toxicity of Cry2Aa (61.11% reduction in larval mortality) than on the Cry1Ac toxin (87.50%), which might indicate that APN exerts a greater effect on the toxicity of Cry1Ac than Cry2Aa (Fig. 5). Our results suggested that CAD, APN4 and ALP2 are involved in the action of the Cry2Aa toxin in H. armigera and play important functional roles in the toxicity of Cry2Aa. Therefore, we hypothesize that CAD, APN4 and ALP2 may function as receptors for Cry2Aa in H. armigera. However, to determine whether the Cry1Ac and Cry2Aa toxins exhibit the same binding sites on CAD, APN or ALP, more compete binding experiments are needed.
Materials and Methods
Insect rearing
The susceptible H. armigera 96S strain was initiated from 20 pairs of adults collected in Xinxiang County, Henan Province, China, in 1996. After collection, the moths were maintained in the laboratory at 27 ± 1 °C, under 70 ± 10% relative humidity (RH) and a photoperiod of 14: 10 light:dark (L:D) h. The larvae of the laboratory colony were reared on an artificial diet, and adults were provided with a 10% sugar solution63. Both larvae and adults were maintained in the laboratory without exposure to any insecticides.
Bt toxins and insect bioassays
The Bt toxins Cry1Ac and Cry2Aa used in this study were purchased from Envirotest-China (an agent of Envirologix Inc., Portland, ME, USA; www.envirotest-china.com); the details of their production and purification were described by Li et al.64. The susceptibility of 96S larvae to Bt proteins was evaluated through diet overlay bioassays65. The Cry1Ac or Cry2Aa protein was dissolved and diluted with 50 mM Na2CO3 (pH 10.5) to obtain various concentrations of either protein, after which 75 μL of solutions corresponding to each dose was overlaid on the diet surface in each well of 24-well plates (TianJin Xiangyushun Co., TianJin, China) and allowed to air-dry. For controls, the same volume of 50 mM Na2CO3 (pH 10.5) was poured onto the surface of the diet. A total of 72 neonates of the 96S strain were then individually placed on the dried diet surface in 24-well plates for each treatment in three replicates. When assessed 7 days later, larvae that were dead or did not grow to the third instar were considered to be dead. Larval mortality was recorded to calculate the LC50.
Midgut preparation, total RNA extraction and cDNA synthesis
The midguts of 20 fifth-instar H. armigera larvae were dissected and washed with an ice-cold 0.7% NaCl solution, then immediately frozen in liquid nitrogen and stored at −80 °C for RNA extraction. Total RNA was extracted from the midguts using the TRIzol reagent (Invitrogen, CA, USA) according to the manufacturer’s instructions. The purity of the total RNA was evaluated by measuring the 260/280 and 260/230 ratios with a NanoVue spectrophotometer (GE Healthcare, CT, USA), and RNA integrity was evaluated through 1% agarose gel electrophoresis. Then, genomic DNA was removed the RNA using DNase I (TakaRa, Dalian, China), and first-strand cDNA was immediately synthesized from 4 μg of total RNA using the SuperScriptTM IIIFirst-Strand Synthesis Kit (Invitrogen, CA, USA).
Expression and purification of recombinant proteins and polyclonal antibody production
According to the nucleotide sequences of the cad gene (GenBank accession no. AF519180), the apn4 gene (GenBank accession no. AY181026) and the alp2 gene (GenBank accession no. EU729323) of H. armigera, three pairs of primers containing BamHI and HindIII restriction sites at the 5′ends (Table S1) were designed to amplify partial fragments of CAD, APN4, and ALP2 corresponding to bases 3649–4383, 1084–1980 and 532–1377, respectively, which encode amino acid residues 1217–1461, 361–660, and 177–459. The conditions for PCR amplification included a pre-denaturing step at 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 1 min, annealing at 55 °C for 30 s and extension at 72 °C for 1.5 min, with a final extension at 72 °C for 10 min. Subsequently, the PCR products were gel purified with the AxyPrep DNA gel extraction kit (Axygen Scientific, CA, USA) and subcloned into the pEASY-T3 vector (TransGen, Beijing, China), then transformed into Escherichia coli Trans-T1 cells (TransGen, Beijing, China). Next, the recombinant T3 vectors were digested with BamHI and HindIII (TaKaRa Bio Inc., Dalian, China) at 37 °C for 2 h, and the gene fragments were then subcloned into the His-tag expression vector pET-30a (+) (Addgene, MA, USA), which had previously been digested with the same restriction enzymes to generate the pET-CAD, pET-APN4 and pET-ALP2 recombinant plasmids. The coding sequences and clone orientations were confirmed by sequencing.
The recombinant plasmids described above were transformed into E.coli BL21 (DE3) (Tiangen, Beijing, China) to express the recombinant HaCAD, HaAPN4 and HaALP2 proteins. Positive transformed clones were selected and cultured at 37 °C in LB medium supplemented with 100 mg/ml ampicillin. The three recombinant proteins were induced by adding isopropyl β-D-1-thiogalactopyranoside (IPTG) to a final concentration of 1 mM. Then, the cultured cells were harvested via centrifugation at 7,000 rpm at 4 °C for 30 min, re-suspended and lysed in phosphate-buffered saline (NaCl 137 mM, KCl 2.7 mM, Na2HPO4 4.3 mM, KH2PO4 1.4 mM, pH 7.4). After sonication on ice for 15 min, total crude protein was isolated from the lysate through centrifugation at 18,000 rpm at 4 °C for 30 min. For protein purification, the expressed recombinant proteins were purified with a nickel-nitrilotriacetic acid (Ni-NTA) affinity column (Genscript Biology Co., NJ, USA) and eluted from the column with elution buffer (8 M urea, 100 mM NaH2PO4, 10 mM Tris-HCl, 500 mM imidazole, pH 8.0), then refolded through a gradient of decreasing concentrations of urea (8 M, 6 M, 4 M, 2 M, 1 M) and desalted with a dialysis bag (molecular weight cutoff 10,000–14,000; Sigma, MO, USA) against 10 mM PBS (pH 7.4) overnight at 4 °C. The purity of these three recombinant proteins was verified in 4–20% gradient SDS-PAGE gels (Genscript Biology Co., NJ, USA), and total protein concentrations were estimated via the Bradford method using bovine serum albumin (BSA, Sigma, MO, USA) as the standard protein66. The purified HaCAD, HaAPN4 and HaALP2 proteins were used as antigens in rabbits, and the sera from immunized rabbits were collected and purified to generate polyclonal antibodies.
Western and ligand blotting assays
The midguts of fifth-instar H. armigera larvae were prepared as described previously to isolate brush border membrane vesicles (BBMVs) through the differential magnesium precipitation method67, and the protein concentration was measured. To detect the CAD, APN4 and ALP2 proteins, total BBMV proteins (20 μg) or purified recombinant proteins (10 μg) were dissolved in loading buffer (LaBest, Beijing, China), then heat denatured and separated in 4–20% gradient SDS-PAGE gels (Genscript Biology Co.,China). Next, the proteins were transferred to polyvinylidene fluoride (PVDF) filters (Millipore, MA, USA) in transfer buffer (39 mM Glycine, 48 mM Tris, 0.037% SDS, and 20% methanol), and the filters were subsequently blocked overnight in PBS (135 mM NaCl, 2 mM KCl, 10 mM Na2HPO4, and 1.7 mM KH2PO4, pH 7.4) containing 0.05% Tween-20 and 5% skim milk with constant agitation at 120 rpm. After being washed three times (10 min each time) with PBST (PBS containing 0.05% Tween-20), the filters were probed for 1 h at room temperature with polyclonal antibodies (1:4,000 dilution) against HaCAD, HaAPN4 and HaALP2 (separately), followed by another three washes. Then, the filters were probed with horseradish peroxidase (HRP)-conjugated anti-rabbit IgG (ZSGB-BIO, Beijing, China) at a 1:10,000 dilution for 1 h at room temperature. After another series of washes, the filters were developed using the EasySee Western Blot Kit (TransGen, Beijing, China) and photographed with an ImageQuant LAS4000mini system (GE Healthcare, Japan).
Prior to ligand blotting assays, the purified Cry2Aa toxin was labeled using a Biotin Labeling Kit (Elabscience, Wuhan, China) according to the manufacturer’s instructions. Then, the BBMV proteins from H. armigera larval midgut or purified HaCAD, HaAPN4 and HaALP2 proteins were separated, transferred and blocked following the same methods described above for western blotting. After being washed three times with PBST, the filters were incubated for 1 h in blocking buffer with biotinylated Cry2Aa toxin (5 μg/mL), then washed again and then probed with horseradish peroxidase (HRP)-conjugated streptavidin (Thermo Fisher Scientific, MA, USA) in blocking buffer (1: 10,000 dilution) for 1 h at room temperature. The subsequent procedures for detection were as same as those described for western blotting.
Binding ELISAs
The binding of the Cry2Aa toxin to the recombinant purified HaCAD, HaAPN4 and HaALP2 proteins was tested via enzyme-linked immunosorbent assays (ELISA). Individual wells of 96-well ELISA plates (Costar 9018, Sigma, USA) were coated overnight at 4 °C with 1 μg of the purified HaCAD, HaAPN4 or HaALP2 protein in a final volume of 100 μl of PBS for each well. On the following day, the plates were washed three times (10 min each) with 200 μl of PBST in each well to remove unbound fragments and then blocked with 200 μl of blocking buffer (PBST containing 1% BSA) for 2 h at room temperature, with shaking at 80 rpm. Next, each well was washed three times with 200 μl of PBST, followed by the addition of different concentrations of the biotinylated Cry2Aa toxin to each well in a final volume of 100 μl of blocking buffer. The plates were subsequently incubated at room temperature for 1 h with constant shaking at 80 rpm. After incubation, unbound toxins in each well were removed through washing steps, and the wells were then incubated with HRP-conjugated streptavidin (Thermo Fisher Scientific, MA, USA) in blocking buffer (1:10,000 dilution) for 1 h at room temperature. The plates were subsequently washed again, and 100 μl of freshly prepared 3,3′,5,5′-tetramethylbenzidine (TMB) ELISA substrate was added to each well. After incubation for 30 min at room temperature, the reaction was terminated by adding 50 μl of 0.5 M H2SO4 and the optical density (OD) values were read with a multi-mode microplate reader (FlexStation 3, Molecular Devices, USA). Nonspecific binding was evaluated in the presence of excess unlabeled Cry2Aa toxin, and specific binding was calculated by subtracting nonspecific binding from total binding. The binding data were analyzed using SigmaPlot v.12.5software (Systat Software, USA).
Silencing of CAD, APN and ALP in H. armigera through RNA interference
The roles of HaCAD, HaAPN4 and HaALP2 in the susceptibility of H. armigera to Cry2Aa were tested with an RNA interference (RNAi) technique using sequence-specific small interfering RNAs (siRNA) synthesized by Invitrogen (USA). The siRNA sequences are listed in Table S1. Lyophilized siRNAs targeting HaCAD, HaAPN4, HaALP2 and GFP (negative control) were dissolved with RNase-free H2O to a final concentration of 0.015 nM. Then, 75 μl of the suspensions or RNase-free H2O (positive control) was poured into the wells of 24-well plates into which 1 ml of the solidified artificial diet had previously been added and allowed to air-dry. Subsequently, early second-instar 96S larvae that had been starved for 12 h were separately transferred into each well of the 24-well plates and allowed to feed for 2 days. The larvae from the control and siRNA-fed treatments were then randomly divided into two groups. One group was collected to assess the transcript levels of HaCAD, HaAPN4 and HaALP2 using quantitative real-time PCR (qPCR) as described below, and the other group was individually transferred to a newly prepared artificial diet containing Cry1Ac (8 μg/cm2) or Cry2Aa (8 μg/cm2) to evaluate the effect of knockdown of HaCAD, HaAPN4 and HaALP2 on the toxicity of the Cry1Ac and Cry2Aa toxins. The larval mortality of the latter group was recorded 7 days later. Each treatment included three replicates, and each replicate included 24 H. armigera larvae.
qPCR analysis of the suppression of HaCAD, HaAPN4 and HaALP2
The suppression of HaCAD, HaAPN4 and HaALP2 transcript levels was analyzed via qPCR to evaluate the effectiveness of the silencing method. Midgut isolation, total RNA extraction and cDNA synthesis were performed as described previously. Each treatment included three biological samples, and each sample included 20 pooled midguts of H. armigera larvae. qPCR was conducted using the cDNA template from each biological sample, and each sample was measured in triplicate in anABI 7500 fast RT-PCR system (Applied Biosystems, Foster City, CA, USA) with the SuperReal PreMix Probe RT-PCR Kit (Tiangen, Beijing, China). qPCR amplifications were carried out in a final volume of 20 μl, containing 1 μl of cDNA (200 ng), 0.6 μl of each primer (10 μM) listed in Table S1, 10 μl of 2 × SuperReal PreMix (probe), 0.4 μl of Probe, 0.4 μl of 50 × RO × Reference Dye and 7.0 μl of RNase-Free ddH2O. The conditions for qPCR involved an initial denaturation at 95 °C for 15 min, followed by 40 cycles of 95 °C for 3 s and 60 °C for 30 s. The relative transcript levels of the target genes were first normalized to the reference genes β-actin (Accession no. EU527017.1) and glyceraldehydes-3-phosphate dehydrogenase (GAPDH; Accession no. JF417983.1), and then calculated using the comparative Ct (2−ΔΔCt) method68.
Data analysis
The concentration of the Cry1Ac or Cry2Aa toxin causing 50% mortality (LC50) and the 95% fiducial limits were calculated via Probit analysis (Polo-PC; LeOra Software, Petaluma, CA, USA). Comparisons to determine significance in the RNAi assays, including assessment of the relative transcription of HaCAD, HaAPN4 and HaALP2 and larval mortality on diets treated with or without Bt toxins, were performed (the larval mortality was arcsine transformed before comparison) via one-way analysis of variance (ANOVA) with Tukey’s test for multiple comparisons using SPSS v.18.0 (SPSS Inc., Chicago, IL, USA).
Additional Information
How to cite this article: Zhao, M. et al. Functional roles of cadherin, aminopeptidase-N and alkaline phosphatase from Helicoverpa armigera (Hübner) in the action mechanism of Bacillus thuringiensis Cry2Aa. Sci. Rep. 7, 46555; doi: 10.1038/srep46555 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Guo, Z. J. et al. MAPK signaling pathway alters expression of midgut ALP and ABCC genes and causes resistance to Bacillus thuringiensis Cry1Ac toxin in diamondback moth. PLoS genetics. 11, e1005124 (2015).
Chen, W. B. et al. A toxin-binding alkaline phosphatase fragment synergizes Bt toxin Cry1Ac against susceptible and resistant Helicoverpa armigera . PloS one 10, e0126288 (2015).
Comas, C., Lumbierres, B., Pons, X. & Albajes, R. No effects of Bacillus thuringiensis maize on nontarget organisms in the field in southern Europe: a meta-analysis of 26 arthropod taxa. Transgenic Res. 23, 135–143 (2014).
Hua, G., Zhang, R., Bayyareddy, K. & Adang, M. J. Anopheles gambiae alkaline phosphatase is a functional receptor of Bacillus thuringiensis jegathesan Cry11Ba toxin. Biochemistry. 48, 9785–9793 (2009).
Frankenhuyzen, K. V. Insecticidal activity of Bacillus thuringiensis crystal proteins. J Invertebr Pathol. 101, 1–16 (2009).
James, C. Global status of commercialized biotech/GM Crops: 2015. ISAAABriefs 49 (ISAAA, Ithaca, NY, 2015) (2015).
Höfte H. & Whiteley H. R. Insecticidal crystal proteins of Bacillus thurin giensis . Microbiological reviews. 53, 242–255 (1989).
Schnepf, E. et al. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiology and molecular biology reviews. 62, 775–806 (1998).
Senthil-Nathan, S. A review of biopesticides and their mode of action against insect pests. Environmental Sustainability. Springer: India, 49–63 (2015).
Gassmann, A. J. et al. Field-evolved resistance by western corn rootworm to multiple Bacillus thuringiensis toxins in transgenic maize. Proc Natl Acad Sci USA 111, 5141–5146 (2014).
Jakka, S. R. et al. Field-evolved mode 1 resistance of the fall armyworm to transgenic Cry1Fa-expressing corn associated with reduced Cry1Fa toxin binding and midgut alkaline phosphatase expression. Appl Environ Microbiol. 82, 1023–1034 (2015).
Tabashnik, B. E. et al. Efficacy of genetically modified Bt toxins alone and in combinations against pink bollworm resistant to Cry1Ac and Cry2Ab. PloS one 8, e80496 (2013).
Carriere, Y., Crickmore, N. & Tabashnik, B. E. Optimizing pyramided transgenic Bt crops for sustainable pest management. Nat Biotechnol. 33, 161–168 (2015).
Fabrick, J. A. et al. Alternative splicing and highly variable cadherin transcripts associated with field-evolved resistance of pink bollworm to Bt Cotton in India. PloS one 9, e97900 (2014).
Tabashnik, B. E., Brevault, T. & Carriere, Y. Insect resistance to Bt crops: lessons from the first billion acres. Nat Biotechnol. 31, 510–521 (2013).
Lu, Y., Wu, K., Jiang, Y., Guo, Y. & Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nature. 487, 362–365 (2012).
Wu, K., Lu, Y., Feng, H., Jiang, Y. & Zhao, J. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin-containing cotton. Science. 321, 1676–1678 (2008).
Zhang, H. et al. Early warning of cotton bollworm resistance associated with intensive planting of Bt cotton in China. PloS one 6, e22874 (2011).
Wei, J. et al. Cross-resistance and interactions between Bt toxins Cry1Ac and Cry2Ab against the cotton bollworm. Sci Rep 5, doi: 10.1038/srep07714 (2015).
Xiao, Y. et al. Resistance to Bacillus thuringiensis mediated by an ABC transporter mutation increases susceptibility to toxins from other bacteria in an invasive insect. PLoS pathogens 12, e1005450 (2016).
Liu, C. et al. Cis-mediated down-regulation of a trypsin gene associated with Bt resistance in cotton bollworm. Sci Rep 4, doi: 10.1038/srep07219 (2014).
Caccia, S. et al. Binding site alteration is responsible for field-isolated resistanceto Bacillus thuringiensis Cry2A insecticidal proteins in two Helicoverpa species. PloS one 5, e9975 (2010).
Hernandez-Rodriguez, C. S., Van Vliet, A., Bautsoens, N., Van Rie, J. & Ferre, J. Specific binding of Bacillus thuringiensis Cry2A insecticidal proteins to a common site in the midgut of Helicoverpa species. Appl Environ Microbiol. 74, 7654–7659 (2008).
Adamczyk, J. J. et al. Evalutions of Bollgard (R), Bollgard II (R), and Widestrike (R) technologies against beet and fall armyworm larvae (Lepidoptera: Noctuidae). Florida Entomologist. 91, 531–536 (2008).
Jurat‐Fuentes J. L. & Adang M. J. Characterization of a Cry1Ac‐receptor alkaline phosphatase in susceptible and resistant Heliothis virescens larvae. European Journal of Biochemistry. 271, 3127–3135 (2004).
Jurat‐Fuentes J. L. & Adang M. J. Cry toxin mode of action in susceptible and resistant Heliothis virescens larvae. J Invertebr Pathol. 92, 166–171 (2006).
Adang, M. J., Crickmore, N. & Jurat-Fuentes, J. L. Chapter Two-Diversity of Bacillus thuringiensis crystal toxins and mechanism of action. Advance in Insect Physiology. 47, 39–87 (2014).
Pardo-Lopez, L., Soberon, M. & Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: mode of action, insect resistance and consequences for crop protection. FEMS Microbiology Rev. 37, 3–22 (2013).
Gahan, L. J., Pauchet, Y., Vogel, H. & Heckel, D. G. An ABC transporter mutation is correlated with insect resistance to Bacillus thuringiensis Cry1Ac toxin. PLoS genetics 6, e1001248 (2010).
Ferre, J. & Van Rie, J. Biochemistry and genetics of insect resistance to Bacillusthuringiensis . Annu Rev Entomol 501–533 (2002).
Tabashnik, B. E. et al. Efficacy of genetically modified Bt toxins against insects with different genetic mechanisms of resistance. Nat Biotechnol. 29, 1128–1131 (2011).
Wang, G., L., G., Wu, K. & Guo, Y. Cloning and seuencing of a gene encoding Aminopetidase N in the midgut of Helicoverpa armigera (Hübner). J Integra Agricul. 2, 760–767 (2003).
Ning, C. et al. Characterization of a Cry1Ac toxin-binding alkaline phosphatase in the midgut from Helicoverpa armigera (Hubner) larvae. J Insect Physiol. 56, 666–672 (2010).
Tabashnik, B. E. et al. Efficacy of genetically modified Bt toxins alone and in combinations against pink bollworm resistant to Cry1Ac and Cry2Ab. PloS one 8, e80496 (2013).
Hu, X. et al. PirB-Cry2Aa hybrid protein exhibits enhanced insecticidal activity against Spodoptera exigua larvae. J Invertebr Pathol. 120, 40–42 (2014).
Ibargutxi, M. A., Muñoz, D. & Escudero, I. R. d. & Caballero, P. Interactions between Cry1Ac, Cry2Ab, and Cry1Fa Bacillus thuringiensis toxins in the cotton pests Helicoverpa armigera (Hübner) and Earias insulana (Boisduval). Biological Control. 47, 89–96 (2008).
Gayen, S., Mandal, C. C., Samanta, M. K., Dey, A. & Sen, S. K. Expression of an engineered synthetic cry2Aa (D42/K63F/K64P) gene of Bacillus thuringiensis in marker free transgenic tobacco facilitated full-protection from cotton leaf worm (S. littoralis) at very low concentration. World journal of microbiology & biotechnology. 32, 1–12 (2016).
Vachon, V., Laprade, R. & Schwartz, J. L. Current models of the mode of action of Bacillus thuringiensis insecticidal crystal proteins: a critical review. J Invertebr Pathol. 111, 1–12 (2012).
Kaur, R., Sharma, A., Gupta, D., Kalita, M. & Bhatnagar, R. K. Bacillus thuringiensis toxin, Cry1C interacts with 128HLHFHLP134 region of aminopeptidase N of agricultural pest, Spodoptera litura . Process Biochem. 49, 688–696 (2014).
Park, Y. & Kim, Y. RNA interference of cadherin gene expression in Spodoptera exigua reveals its significance as a specific Bt target. J Invertebr Pathol. 114, 285–291 (2013).
Herrero, S., Bel, Y., Hernandez-Martinez, P. & Ferre, J. Susceptibility, mechanisms of response and resistance to Bacillus thuringiensis toxins in Spodoptera spp. Current opinion in insect science. 15, 89–96 (2016).
Flores-Escobar, B., Rodriguez-Magadan, H., Bravo, A., Soberon, M. & Gomez, I. Differential role of Manduca sexta aminopeptidase-N and alkaline phosphatase in the mode of action of Cry1Aa, Cry1Ab, and Cry1Ac toxins from Bacillus thuringiensis . Appl Environ Microbiol. 79, 4543–4550 (2013).
Wang, J. et al. Functional validation of cadherin as a receptor of Bt toxin Cry1Ac in Helicoverpa armigera utilizing the CRISPR/Cas9 system. Insect Bioche Molecular Biol. 76, 11–17 (2016).
Wei, J. et al. APN1 is a functional receptor of Cry1Ac but not Cry2Ab in Helicoverpa zea . Sci Rep 6, doi: 10.1038/srep19179 (2016).
Qiu, L. et al. Cadherin is involved in the action of Bacillus thuringiensis toxins Cry1Ac and Cry2Aa in the beet armyworm, Spodoptera exigua . J Invertebr Pathol. 127, 47–53 (2015).
Tiewsiri, K. & Wang, P. Differential alteration of two aminopeptidases N associated with resistance to Bacillus thuringiensis toxin Cry1Ac in cabbage looper. Proc Natl Acad Sci USA 108, 14037–14042 (2011).
Senthil-Nathan, S., Chung, P. G. & Murugan, K. Effect of botanical insecticides and bacterial toxins on the gut enzyme of the rice leaffolder Cnaphalocrocis medinalis . Phytoparasitica. 32, 433–443 (2004).
Senthil-Nathan, S., Chung, P. G. & Murugan, K. Combined effect of biopesticides on the digestive enzymatic profiles of Cnaphalocrocis medinalis (Guenée) (the rice leaffolder) (Insecta: Lepidoptera: Pyralidae). Ecotoxicology & environmental safety. 64, 382–389 (2006).
Arenas, I., Bravo, A., Soberon, M. & Gomez, I. Role of alkaline phosphatase from Manduca sexta in the mechanism of action of Bacillus thuringiensis Cry1Ab toxin. J BiologicChem. 285, 12497–12503 (2010).
Pacheco, S. et al. Domain II loop 3 of Bacillus thuringiensis Cry1Ab toxin is involved in a “ping pong” binding mechanism with Manduca sexta aminopeptidase-N and cadherin receptors. J BiologicChem. 284, 32750–32757 (2009).
Perera, O. P., Willis, J. D., Adang, M. J. & Jurat-Fuentes, J. L. Cloning and characterization of the Cry1Ac-binding alkaline phosphatase (HvALP) from Heliothis virescens . Insect Biochem Mol Biol. 39, 294–302 (2009).
Sengupta, A., Sarkar, A., Priya, P., Ghosh Dastidar, S. & Das, S. New insight to structure-function relationship of GalNAc mediated primary interaction between insecticidal Cry1Ac toxin and HaALP receptor of Helicoverpa armigera . PloS one 8, e78249 (2013).
English, L. et al. Mode of action of CryIIA: a Bacillus thuringiensis delta-endotoxin. Insect Biochem. Mol. Biol. 24, 1025–1035 (1994).
Estela, A., Escriche, B. & Ferre, J. Interaction of Bacillus thuringiensis toxins with larval midgut binding sites of Helicoverpa armigera (Lepidoptera: Noctuidae). Appl Environ Microbiol. 70, 1378–1384 (2004).
Bravo, A., Gill, S. S. & Soberón, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon. 49, 423–435 (2007).
Tabashnik, B. E. et al. Asymmetrical cross-resistance between Bacillus thuringiensis toxins Cry1Ac and Cry2Ab in pink bollworm. Proc Natl Acad Sci USA 106, 11889–11894 (2009).
Welch, K. L. et al. Cross-resistance to toxins used in pyramided Bt crops and resistance to Bt sprays in Helicoverpa zea . J Invertebr Pathol. 132, 149–156 (2015).
Brévault, T. et al. Potential shortfall of pyramided transgenic cottonfor insect resistance management. Proc Natl Acad Sci USA 110, 5806–5811 (2013).
Chen, R. R. et al. A cadherin-like protein from the beet armywormSpodoptera exigua (Lepidoptera: Noctuidae) is a putative Cry1Ac receptor. Archives of Insect Biochem & Physiol. 86, 58–71 (2014).
Yuan, X. D. et al. New insights on the role of alkaline phosphatase 2 from Spodoptera exigua (Hübner) in the action mechanism of Bt toxin Cry2Aa. J Insect Physiol. 98, 101–107 (2017).
Valaitis, A. P. Localization of Bacillus thuringiensis Cry1A toxin-binding molecules in gypsy moth larval gut sections using fluorescence microscopy. J Invertebr Pathol. 108, 69–75 (2011).
Ren, X. L., Ma, Y., Cui, J. J. & Li, G. Q. RNA interference-mediated knockdown of three putative aminopeptidases N affects susceptibility of Spodoptera exigua larvae to Bacillus thuringiensis Cry1Ca. J Insect Physiol. 67, 28–36 (2014).
Liang, G. M., Tan, W. J. & Guo, Y. Y. An improvement in the technique of artificial rearing cotton bollworm. Plant Protection 15–17 (1999).
Li, Y. et al. Consumption of Bt rice pollen containing Cry1C or Cry2A does not pose a risk to Propylea japonica (Thunberg) (Coleoptera: Coccinellidae). Sci Rep 5, doi: 10.1038/srep07679 (2015).
Hernández-Rodríguez, C. S., Vliet, A. V., Bautsoens, N., Rie, J. V. & Ferré, J. Specific binding of Bacillus thuringiensis Cry2A insecticidal proteins to a common site in the midgut of Helicoverpa Species. Appl Environ Microbiol. 74, 7654–7659 (2008).
Bradford, M. M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254 (1976).
Wolfersberger, M. et al. Preparation and partial characterization of amino acid transporting brush border membrane vesicles from the larval midgut of the cabbage butterfly (Pieris brassicae). Comparative Biochemistry & Physiology Part A Physiology. 86, 301–308 (1987).
Livak, K. J. & Schmittgen, T. D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔ C T Method. Methods. 25, 402–408 (2001).
Acknowledgements
This research was supported by the Key Project for Breeding Genetically Modified Organisms (grant number 2016ZX08011–002) and the National Natural Science Foundation of China (grant number 31621064).
Author information
Authors and Affiliations
Contributions
G.L., M.Z. and X.Y. designed the study. M.Z. and X.Y. performed the experiments. G.L. and M.Z analyzed the data and wrote the manuscript. J.W., W.Z., B.W. and M.K. provided help for performing the experiments. G.L. provided all experimental materials. M.Z. and G.L. shared in the scoping and writing responsibilities. All authors have read and approved the manuscript for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhao, M., Yuan, X., Wei, J. et al. Functional roles of cadherin, aminopeptidase-N and alkaline phosphatase from Helicoverpa armigera (Hübner) in the action mechanism of Bacillus thuringiensis Cry2Aa. Sci Rep 7, 46555 (2017). https://doi.org/10.1038/srep46555
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep46555
- Springer Nature Limited
This article is cited by
-
Molecular profiling of resistance alleles in Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae) collected from different locations
Egyptian Journal of Biological Pest Control (2023)
-
Identifying the Epitopes of Bacillus thuringiensis Cry2Aa Toxin Involved in Cadherin Interaction by a Monoclonal Antibody
Applied Biochemistry and Biotechnology (2023)
-
Molecular stacking of two codon-modified genes encoding Bt insecticidal proteins, Cry1AcF and Cry2Aa for management of resistance development in Helicoverpa armigera
Journal of Plant Biochemistry and Biotechnology (2020)
-
Midgut de novo transcriptome analysis and gene expression profiling of Spodoptera exigua larvae exposed with sublethal concentrations of Cry1Ca protein
3 Biotech (2020)
-
Search for Cry proteins expressed by Bacillus spp. genomes, using hidden Markov model profiles
3 Biotech (2019)