
Abstract
In the Late Cretaceous, northern and southern hemispheres evolved distinct dinosaurian faunas. Titanosaurians and abelisaurids dominated the Gondwanan continents; hadrosaurids, ceratopsians and tyrannosaurs dominated North America and Asia. Recently, a lambeosaurine hadrosaurid, Ajnabia odysseus, was reported from the late Maastrichtian phosphates of the Oulad Abdoun Basin Morocco, suggesting dispersal between Laurasia and Gondwana. Here we report new fossils from the phosphates of Morocco showing lambeosaurines achieved high diversity in the late Maastrichtian of North Africa. A skull represents a new dwarf lambeosaurine, Minqaria bata. Minqaria resembles Ajnabia odysseus in size, but differs in the ventrally positioned jugal facet and sinusoidal toothrow. The animal is small, ~ 3.5 m long, but the fused braincase shows it was mature. A humerus and a femur belong to larger hadrosaurids, ~ 6 m long, implying at least three species coexisted. The diversity of hadrosaurids in Europe and Africa suggests a dispersal-driven radiation, with lambeosaurines diversifying to take advantage of low ornithischian diversity. African lambeosaurines are small compared to North American and Asia hadrosaurids however, perhaps due to competition with titanosaurians. Hadrosaurids are unknown from eastern Africa, suggesting Moroccan hadrosaurids may be part of a distinct insular fauna, and represent an island radiation.
Similar content being viewed by others


Introduction
Dinosaurs from the end of the Cretaceous remain poorly known in Africa and the Afro-Arabian plate, especially compared to the faunas of North America and Asia. Only a handful of localities, in Egypt1,2,3,4, Angola5, Oman6, and Morocco7,8,9,10 have produced Campanian—Maastrichtian dinosaurs. Initial finds suggested African dinosaurs resembled those of other Gondwanan landmasses, i.e. India, Madagascar, and South America, with faunas dominated by titanosaurians1,9 and abelisaurids8. However, the presence of hadrosauroids in the latest Cretaceous of Oman6 and Angola5 suggests a more complicated picture. More recently, a small hadrosaurid, Ajnabia odysseus7, was discovered in the late Maastrichtian phosphates of Morocco (Fig. 1).

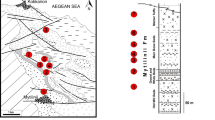
Map showing localities that produced the fossil specimens described in this paper. (A), location of the Oulad Abdoun Basin, central Morocco; (B), localities of hadrosaurids recovered from Sidi Daoui, Mrah Lahrach, and Sidi Chennane.
Duckbill dinosaurs, or hadrosaurids, were highly successful herbivores that staged a major radiation in the Late Cretaceous11,12,13. Hadrosaurids evolved in North America in the Turonian7, before dispersing into Asia and Europe7. The presence of a hadrosaurid in Africa is perplexing, because Africa had been isolated from Laurasia by deep seaways since the mid-Jurassic14, while hadrosaurids evolved in the Late Cretaceous. The resolution to the paradox appears to be that duckbills swam or rafted to Africa7.
Ajnabia is related to the European lambeosaurines, Arenysaurini7, suggesting overwater dispersal from Europe. Yet with Ajnabia known by only a pair of maxillae and a partial dentary, our knowledge of African hadrosaurids is tantalizingly incomplete. We report three new hadrosaurid specimens from the late Maastrichtian of Morocco (Fig. 1). One represents a new small lambeosaurine, distinct from Ajnabia. Two specimens represent larger lambeosaurines, suggesting at least one additional species.
Geographic and geological setting
The new hadrosaurid remains come from the marine phosphates of the Oulad Aboun Basin, in Khouribga Province. The new small lambeosaurine, Minqaria bata, comes from the mines at Sidi Chennane, the same locality that produced Ajnabia odysseus7, and is based on an associated skull. The other fossils come from Sidi Daoui and Mrah Lahrach (Fig. 1) and were found as isolated elements, as is typical of the phosphates.
The Phosphates consist of phosphatic sands, marls, and limestones (Fig. 2), deposited in an embayment along the eastern margin of the Atlantic from the late Cretaceous to the early Eocene15,16. The Phosphates are divided into beds or ‘couches’. Couche III is Cretaceous; Couche II, I and 0, are Paleogene (Danian—Ypresian). Couche III is Maastrichtian based on shark teeth17,18 and chemostratigraphy15, upper Couche III is latest Maastrichtian, dating to ~ 1 Ma or less before the K-Pg boundary.

Synthetic stratigraphic column of the Phosphates of the Oulad Abdoun Basin of Morocco, showing position of hadrosaurid fossils in Upper Couche III, late Maastrichtian. After Kocsis et al.15.
Couche III is dominated by abundant and diverse marine vertebrates, and represents a highly productive upwelling zone19. Shark teeth17,18 and fish remains17 are the most common fossils. Marine reptiles are exceptionally common, and include diverse mosasauroids17,20,21,22,23,24,25,26,27,28,29,30,31,32,33, sea turtles, elasmosaurid plesiosaurs34,35, and rare crocodilians36; pterosaurs are also common37,38.
Dinosaurs are very rare as a proportion of fossils, but the exceptional abundance of fossils means Morocco’s Phosphates currently have the best record of dinosaurs from the latest Cretaceous of Africa. Dinosaurs include titanosaurians9, the abelisaurid Chenanisaurus barbaricus8,10 and two smaller abelisaurids39, and the lambeosaurine hadrosaurid Ajnabia odysseus7. Dinosaurs typically preserve as isolated bones8,39 and more rarely as associated remains7,9, a pattern also seen in the marine reptiles. This preservational mode likely results from carcasses spending time afloat, then falling apart due to decomposition and the action of scavengers; shark feeding traces are common on bones in the phosphates29.
Systematic paleontology
Dinosauria Owen, 1842
Ornithischia Seeley, 1887
Iguanodontia, Baur, 1891
Hadrosauridae Cope, 1869
Lambeosaurinae Parks, 1923
Arenysaurini Longrich, Pereda-Suberbiola, Pyron et Jalil, 2021
Minqaria bata gen. et sp. nov.
Etymology. Arabic, ‘minqar’, beak; ‘bata’, duck.
Holotype. MHNM.KHG.1395 (Museum d'Histoire Naturelle de Marrakech, Marrakech, Morocco, Cadi Ayyad University), right maxilla with teeth, left dentary, and braincase (Figs. 3, 4, 5, 6 and 7). The remains found in association by locals working the mines; color and preservation of the bone and associated matrix, size, and lack of duplication of elements are consistent with an associated individual.

Skull elements of Minqaria bata nov. gen. et sp., MHNM.KHG.1395, holotype: right maxilla, braincase, left dentary. Sidi Chennane, Oulad Abdoun Basin, Upper Couche III, late Maastrichtian. Scale = 5 cm.
Locality and horizon. Sidi Chennane, Oulad Abdoun Basin, Morocco; upper Couche III, late Maastrichtian (Figs. 1, 2).
Diagnosis. Small lambeosaurine characterized by the following character combination, which also differentiates it (where the specimens overlap) from Ajnabia odysseus. Jugal articulation lies very low on maxilla (autapomorphic within lambeosaurinae); ectopterygoid ridge ends at jugal articulation. Ectopterygoid ridge concave in lateral and dorsal views, narrower posteriorly than anteriorly. Neurovascular foramina arranged in a row. Highly domed frontoparietal, with extensive contribution of parietal to the dome, and a triangular parietal table. Maxillary toothrow sinusoidal in lateral or ventral views, with a deep buccal fossa. Dentary short and deep, occlusal margin straight; symphyseal process strongly extended anteriorly, and with straight ventral margin. Alveolar ridges of maxilla and dentary poorly developed. Teeth small, with narrow apices, broad central ridges, and rugose enamel.
Description and comparisons.
Maxilla. The maxilla (Figs. 3, 4) is 195 mm long as preserved, but is incomplete anteriorly and dorsally. Alveoli become much smaller anteriorly (Fig. 4b), suggesting the maxilla did not extend much further; perhaps 15–20% of the bone’s length is missing anteriorly. There are 27 alveoli preserved and perhaps two or three on the missing part of the maxilla. This is low for Lambeosaurinae, which typically have more than thirty teeth40,41,42,43, but a low tooth count occurs in Ajnabia odysseus7 and Canardia garonnensis44. Because hadrosaurids added tooth positions as they grew40, this feature may result from the animal’s small size.

Minqaria bata nov. gen. et sp., MHNM.KHG.1395, holotype, Sidi Chennane, Oulad Abdoun Basin, Upper Couche III, late Maastrichtian. Right maxilla of in (a), lateral, (b), ventral, (c), medial, and (d), dorsal views. Abbreviations: buc, buccal fossa ec, ectopterygoid process, ecr, ectopterygoid ridge; ju, jugal articular facet, ne, neurovascular foramina, pmx, premaxillary process. Scale = 5 cm.
In lateral view (Fig. 4a), the maxilla is short anteroposteriorly and tall dorsoventrally, as in other hadrosaurids11 especially Lambeosaurinae7,45, and subrectangular. The maxilla’s anterodorsal margin extends up and forms a broad dorsal process, as in Saurolophinae11 and Arenysaurini including Canardia garonnensis and Pararhabdodon isonensis44; the maxilla is broken here in Ajnabia7. In Lambeosaurini46 and Parasaurolophini42,45, the maxilla is shallow anterior to the jugal articulation, with the premaxilla approaching the jugal articulation. The maxilla is narrow in dorsal (Fig. 4d) or ventral view (Fig. 4b), similar to Pararhabdodon isonensis; maxillae of Ajnabia and Canardia are broad in dorsal or ventral view.
The premaxillary process is downturned anteriorly (Fig. 4a), as in Ajnabia7 and other Arenysaurini44, and certain lambeosaurines such as Corythosaurus casuarius47, Amurosaurus riabinini41, Olorotitan arharensis42 and Magnapaulia laticaudus43. The maxilla’s anterolateral surface is convex; it is flatter in Ajnabia7, Pararhabdodon44, and Canardia44. The toothrow is sinusoidal in lateral view, curving inward anteriorly, outwards below the jugal facet, and curving in again posteriorly; the toothrow is straighter in lateral view in Ajnabia7 and the arenysaurins Pararhabdodon44 and Canardia44. The teeth are strongly inset medially (Fig. 4b), creating a deep buccal fossa absent in Ajnabia7 and other arenysaurins44.
The jugal facet lies very low on the maxilla (Fig. 4a), unlike all other Lambeosaurinae, where the jugal facet lies high on the maxilla7,45, an autapomorphy of the species. Here Minqaria resembles saurolophines such as Kritosaurus and Edmontosaurus45. The jugal articulation is convex anteroventrally, then forms a straight edge where it extends above the ectopterygoid ridge. Extension of the jugal articulation posterodorsally above the ectopterygoid ridge is an apomorphy of Lambeosaurinae7,45.
Five neurovascular foramina extend in a line from below the jugal articulation anteriorly onto the maxilla’s lateral surface (Figs. 4a, b). This differs from Ajnabia7, where a pair of neurovascular foramina lie below the jugal articulation. Neither is this condition seen in Pararhabdodon44, but it is similar to Canardia44.
The ectopterygoid ridge (Fig. 4a) extends from the jugal articulation back along the maxilla’s posterior dentigerous. It forms a low, broad ridge, similar to that of Ajnabia7; the ectopterygoid ridge in Canardia and Pararhabdodon is narrower and better defined44. The ridge has a distinctly concave ventral margin, similar to Canardia, those of Ajnabia and Pararhabdodon are straight in lateral view44. In dorsal view (Fig. 4d), the shelf formed by the ectopterygoid ridge narrows towards the back of the jaw; that of Ajnabia7 becomes broader, and is more convex.
In ventral view (Fig. 4b), the toothrow is sinusoidal. The toothrow curves inwards at the front of the jaw, outwards near the middle of the maxilla, then curves inwards again at the back. Ajnabia7 has a similar but less extreme curvature, the toothrow of Pararhabdodon is only weakly sinusoidal48. The toothrow in Canardia is strongly curved outward posteriorly44, but this feature may be exaggerated by crushing.
Maxillary dentition. Several unerupted teeth lie partially exposed in the maxillae (Figs. 4, 5). These teeth are typical of hadrosaurids in being much taller than wide11 and resemble lambeosaurines41,46,49,50,51 in their tall, narrow, lanceolate shape (Fig. 5). The tips of the crowns are narrower and more pointed than in Ajnabia odysseus7. As in other hadrosaurids, maxillary teeth bear enamel on the lateral surface of the tooth only11. The enamel has a distinctly coarse, roughened texture, Ajnabia7 has smoother enamel.

Minqaria bata nov. gen. et sp., MHNM.KHG.1395, holotype, Sidi Chennane, Oulad Abdoun Basin, Upper Couche III, late Maastrichtian. Dentition. (a), posterior maxillary dentition, (b), closeup of teeth. Abbreviations: cr, central ridge; dc, distal carina; mc, mesial carina. Scale = 1 cm.
The mesial and distal carinae have highly reduced denticles, a derived feature shared with other Arenysaurini7, but unlike most Lambeosaurinae11 where denticles are large and well-developed. The single central ridge is slightly offset towards the distal carina, as in Ajnabia7. The central ridge is low and rounded; in Ajnabia it is taller and narrower. The lateral, enamel-covered surface of the tooth is not planar, but instead curls onto the apex of the tooth, as in Ajnabia7.
Braincase. The braincase (Fig. 6) comprises the frontals, parietal, orbitosphenoids, laterosphenoids, prootics, supraoccipital, basisphenoid, basioccipital, and exoccipitals. Overall, the braincase closely resembles that of Arenysaurus11.

Minqaria bata nov. gen. et sp., MHNM.KHG.1395, holotype, Sidi Chennane, Oulad Abdoun Basin, Upper Couche III, late Maastrichtian. Braincase in (a), dorsal, (b), ventral, (c), anterior, (d), posterior, (e), right lateral, and (f), left lateral views. Abbreviations: Bo, basioccipital; bpt, basipterygoid processes; Bsp, basisphenoid; Ex, exoccipital, Fr, frontal; Lsp, laterosphenoid; Par, parietal; Pro, prootic; So, supraoccipital. Roman numerals (X, XII) = cranial nerve foramina. Scale = 5 cm.
The frontals contact each other medially, the parietal posteriorly (Fig. 6a), the laterosphenoid ventrally. Although the sutures are visible, the frontals are tightly joined to each other and the parietal by a strongly interdigitating contact, similar to that seen in other lambeosaurines52. The degree of interdigitation is stronger than in Arenysaurus50, but not developed to the same degree as seen in Corythosaurus52. In places the frontoparietal suture is well-developed, but in others is difficult to see, due either to tight knitting of bones or perhaps partial fusion of bones. The tight knitting of the bones suggests the animal is mature.
The frontals form a well-developed dome, as in Arenysaurus50, although the frontals of Minqaria are less strongly domed than in Arenysaurus, where the posterior margin of the frontals rises up almost vertically. A well-developed dome also occurs in Jaxartosaurus. The frontals of Lambeosaurini40 and Parasaurolophini53,54are less strongly domed than in Minqaria or Arenysaurus. This may be partly size-related, as juvenile lambeosaurines, e.g. Hypacrosaurus, show stronger frontal domes than adults40.
Laterally, frontal postorbital processes are slightly concave. Posteriorly, the frontals form a V-shaped notch to receive a triangular tab from the parietal.
The parietal contacts the frontals anteriorly (Fig. 6a), the laterosphenoids anteroventrally, and the supraoccipital posteriorly (Fig. 6e, f). Along the midline the parietal forms a broad, triangular anterior process projecting between the frontals, contributing to the doming of the skull roof. A similar morphology is seen in Arenysaurus50; in Jaxartosaurus the parietal has a broad, quadrangular contribution to the dome. A similar triangular process contributing to the frontoparietal dome occurs in juvenile Corythosaurus but is reduced in adults52. In the basal hadrosauroid Eotrachodon, a triangular process projects between the frontals55, but does not contribute to the dome; the saurolophines Edmontosaurus56 and Gryposaurus52 lack this triangular process.
The parietal’s anterolateral processes angle forward and wrap around the back of the frontals. A similar morphology occurs in Arenysaurus, Jaxartosaurus, the lambeosaurin Corythosaurus54 and the parasaurolophin Charonosaurus. The anterolateral processes project laterally in Saurolophinae52,56 and Eotrachodon55. The parietal has a small, triangular parietal table and behind this a parietal sagittal crest; in Arenysaurus50 the parietal lacks a parietal table and the sagittal crest extends further forward towards the frontals.
The parietal is broadly arched between the supratemporal fenestrae, with a narrow sagittal crest as in Hypacrosaurus40; the parietal of Arenysaurus is narrower50. The parietal is moderately elongate, resembling Arenysaurus, Lambeosaurini52 and Parasaurolophini54; the parietal of Tsintaosaurus is shorter; the parietal of Saurolophinae tends to be more elongate52,56. Ventrolaterally, the parietal curls down to contact the orbitosphenoids, laterosphenoids, and prootics.
Laterally (Fig. 6e, f), the laterosphenoid and prootic form the lateral walls of the braincase. Ventrally, the basisphenoid projects down forming a pair of long basipterygoid processes. The basipterygoid processes project caudally as in Arenysaurus50 and Olorotitan42; the basipterygoid processes project ventrally in Hypacrosaurus altispinus40 and the saurolophines Brachylophosaurus canadensis57, and Edmontosaurus regalis56 and anteroventrally in Parasaurolophus cyrtocristatus53.
Posterior to the basipterygoid processes, the basisphenoid is markedly concave, as in Arenysaurus11. The basisphenoid’s alar process is well-developed and posterolaterally projecting. It is similarly well-developed in Arenysaurus, but projects more laterally. The alar process is small in most hadrosaurids except brachylophosaurins57. Posteriorly, the supraoccipital is tall and triangular, with a midline crest not seen in other hadrosaurids40,54,56. The foramen magnum and occipital condyle are obscured by hard matrix. Two cranial nerves perforate the exoccipital, (Fig. 6f) forming the exits of the hypoglossal (XII) nerve posteriorly and of the glossopharyngeal and vagoaccessory nerves (IX-XI) anteriorly through what appears to be a single metotic foramen (Fig. 6f).
Dentary. The dentary (Fig. 7) is short and deep, similar to Blasisaurus canudoi49, Corythosaurus casuarius47 or Hypacrosaurus altispinus40, and unlike the elongate dentary of the arenysaurins Arenysaurus ardevoli50 and Koutalisaurus kohlerorum58, or lambeosaurines such as Amurosaurus riabinini41, Olorititan arharensis42, Sahaliyania elunchunorum59, and Parasaurolophus tubicen51. The dentary’s dorsal and ventral margins are roughly parallel in lateral view (Fig. 7a), giving it a rectangular shape. The dentary’s dentigerous margin is straight for most of its length, but turns downwards at the anterior and posterior end of the toothrow, similar to Blasisaurus49 and Koutalisaurus58; the dentigerous margin of Arenysaurus9 is more convex. The dentary’s ventral margin is nearly straight, with a slight inflection where the symphyseal process projects anteroventrally. This weak inflection and the anteriorly projecting symphyseal process are shared with Koutalisaurus58; in Arenysaurus50 and Blasisaurus49, the ventral margin of the jaw curves more and the symphyseal process projects more strongly downward. The posteroventral margin of the dentary is weakly convex. The coronoid process is strongly projected laterally as in other hadrosaurids.

Minqaria bata nov. gen. et sp., MHNM.KHG.1395, holotype, Sidi Chennane, Oulad Abdoun Basin, Upper Couche III, late Maastrichtian. Dentary in (a), lateral, (b), medial, and (c), dorsal views. Abbreviations: alv, alveolar slots; cp, coronoid process; mec, Meckelian canal; sym, symphysis. Scale = 5 cm.
In medial view (Fig. 7b), there are at least 25 alveoli, formed as long, narrow grooves to accommodate multiple replacement teeth, as in other Hadrosauridae11. Alveolar slots are shallower than in Ajnabia7. Alveoli extend three-quarters of the way down the dentary. Alveoli extend further ventrally in Blasisaurus49, Koutalisaurus58, and Hypacrosaurus40, and almost all the way down in Arenysaurus50. The Meckelian canal is visible medially as in Blasisaurus and Koutalisaurus58; it runs ventrally and is concealed in medial view in Arenysaurus50 and Hypacrosaurus40. In dorsal view (Fig. 7c), the dentary is narrow and the toothrow has a sinuous curve matching the curve of the maxillary toothrow.
Lambeosaurinae indet.
Material. MHNM.KHG.1483, left humerus (Fig. 8); MHNM.KHG.1484, right femur (Fig. 9).

Lambeosaurinae indet., MHNM.KHG.1483, Sidi Daoui, Oulad Abdoun Basin, Upper Couche III, late Maastrichtian. Left humerus in posterior (a), posterolateral (b) and lateral (c) views. Abbreviations: dpc, deltopectoral crest; of, olecranon fossa; rc, radial condyle; uc, ulnar condyle. Scale = 5 cm.

Lambeosaurinae indet., MHNM.KHG.1484, Mrah Lahrach, Oulad Abdoun Basin, Upper Couche III, late Maastrichtian. Right femur in anterior (a), medial (b), posterior (c), and lateral (d) views. Abbreviations: gt, greater trochanter; ig, intercondylar groove; lc, lateral condyle, lt, lesser trochanter; mc, medial condyle; tr4, fourth trochanter. Scale = 20 cm.
Locality and Horizon. MHNM.KHG.1483 comes from Sidi Daoui, and MHNM.KHG.1484 from Mrah Lahrach, in the Oulad Abdoun Basin of Morocco (Fig. 1). Both come from upper Couche III, corresponding to the late Maastrichtian (Fig. 2).
Description and comparisons
Humerus. MHNM.KHG.1483 is a left humerus missing its proximal end (Fig. 8). The bone measures 278 mm long, and likely measured around 435 mm when complete. Assuming proportions similar to Corythosaurus casuarius60, MHNM.KHG.1483 was around 5.9 m in length.
In lateral view (Fig. 8a), the humerus is long and gracile. It has a weak sigmoidal curvature, the shaft being bowed anteriorly at the level of the deltopectoral crest, and the distal shaft being bowed posteriorly. The distal shaft is weakly curved, as in other lambeosaurines54, including the arenysaurins Canardia garonnensis44 and cf. Pararhabdodon isonensis61; the distal shaft is more strongly curved in Saurolophinae62,63, but straight in some basal hadrosauroids64.
In posterior view (Fig. 8c), the humerus is narrow at midshaft and broadly flared distally. The radial and ulnar condyles are well-developed. A deep olecranon fossa separates the radial and ulnar condyles, and extends about halfway up the shaft.
The prominent deltopectoral crest (Fig. 8b) projects strongly anterolaterally. The distal margin is strongly concave as in other lambeosaurines44,54 and saurolophines62,63, but unlike Hadrosaurus foulkii65 or basal hadrosauroids64,66 where it is weakly concave, or straight.
Femur. MHNM.KH.1484 (Fig. 9) is a right femur. It measures 640 mm in length, with a midshaft circumference of 238 mm. Scaling a skeletal reconstruction of Corythosaurus60 to a femur length of 640 mm suggests a length of ~ 4.5 m.
The femur’s proximal end is crushed (Fig. 9a), and the bone is difficult to differentiate or physically separate from the consolidated matrix, complicating comparisons. The shaft is large in diameter despite the animal’s small size. It is anteroposteriorly crushed but even accounting for crushing, it is robust, even compared to large hadrosaurids like Magnapaulia laticaudus43 and Olorotitan arharensis42.
The femoral shaft appears to be relatively straight in anterior view (Fig. 9a), as in Lambeosaurinae43,67, Edmontosaurini63, and Hadrosaurini65. In basal hadrosauroids, the femoral shaft is bowed66,68 with the condyles more obliquely oriented relative to the femoral shaft, suggesting a less vertical femoral posture.
In medial or lateral view (Fig. 9b, d), the shaft has a gentle sigmoidal curvature, with the proximal shaft being bowed posteriorly, and the distal end bowed anteriorly. This sigmoid curvature occurs in the type of Orthomerus dolloi68, but not in other specimens69, and in Parasaurolophini54, but not in Lambeosaurini43. The femur of the arenysaurin Adynomosaurus arcanus is straight in lateral view67, but has also been reconstructed. The femoral shaft is also straight in some saurolophines, e.g. Secernosaurus koerneri70 and Saurolophus osborni71. Sigmoidal curvature of the femoral shaft occurs in some basal hadrosauroids, including Bactrosaurus johnsoni72 and Telmatosaurus transsylvanicus68, but it is straighter in others, such as Gobihadros mongoliensis64.
There is a convex fourth trochanter at midshaft (Fig. 9b), on the posteromedial surface of the shaft. The fourth trochanter is weakly projecting, shared with the arenysaurin Adynomosaurus arcanus67, the Basturs Poble arenysaur61, a femur from the Maastricht Formation of the Netherlands73, and Orthomerus dolloi68,69. The fourth trochanter is a prominent, winglike triangular structure in parasaurolophins42, lambeosaurins43, saurolophins74, and basal hadrosauroids64,68,72. A low fourth trochanter may be a diagnostic feature of Arenysaurini.
Medial and lateral condyles are well-developed and project anterior and posterior to the shaft as in other hadrosaurids. The cranial intercondylar groove is broadly open (Fig. 9c). There is a deep posterior intercondylar groove. Because body mass is tightly correlated with limb bone dimensions, particularly diameters and circumferences75, it is possible to make a mass estimate from the femur. Given a femur circumference of 238 mm and assuming a ratio of humerus circumference to femur circumference of 0.651, as seen in Corythosaurus76, the corresponding humeral circumference should be 154.9. A regression of humerus + femoral circumference suggests a mass of 1066 kg based on the equation for quadrupeds from Campione and Evans75 and 1099 kg for the equation for quadrupeds from Campione 201777 for this ~ 4.5 m long animal. The Sidi Daoui humerus suggests a larger, 5.9 m long animal, approximately twice this mass, but smaller than contemporary Laurasian hadrosaurids.
Discussion
Ontogeny. Skeletal fusion can be used as a proxy for maturity in dinosaurs. Although some elements (e.g. parietals) can fuse early in ontogeny, extensive skull fusion occurs as animals approach full size40,78,79. Extensive co-ossification of the braincase in the Minqaria holotype is therefore consistent with skeletal maturity. No clear exoccipital-prootic suture is visible, suggesting these elements are fused; fusion between these bones occurs late in ontogeny in lambeosaurines40. The prootic-laterosphenoid suture is also indistinct suggesting fusion; the laterosphenoid—parietal suture is visible anteriorly but not posteriorly, suggesting partial fusion. This extensive cranial fusion contrasts with juvenile and subadult lambeosaurines40,80, showing MHNM.KH.1484 is a mature adult.
Frontal-frontal and frontal-parietal sutures are also highly interdigitated and the midline of the parietal forms a sagittal crest, as in adult lambeosaurines40,81. Finally, surface texture of the bone suggests maturity. Juvenile lambeosaurines, like other juvenile dinosaurs, show striated bone textures associated with rapid growth82; but the bone of MHNM.KH.1484 lacks striated texture.
Systematics. Minqaria is distinguished from Ajnabia by the shape of the maxilla (Fig. 4), which has a more ventrally placed jugal facet, a curved ectopterygoid ridge, a more sinusoidal toothrow, and neurovascular foramina arranged in a line. However, Minqaria closely resembles Ajnabia and other arenysaurins in its small size, and many anatomical features. Derived features uniting Minqaria with arenysaurins include the reduced number of maxillary alveoli (shared with Ajnabia and Canardia), a posteriorly shifted dental midline ridge (shared with Ajnabia, Blasisaurus, and the Serrat del Rostiar lambeosaurine); a strongly domed frontal (shared with Arenysaurus, and the Basturs lambeosaurine). The dentary predentary process appears to resemble that of other arenysaurins in being triangular, with straight dorsal and ventral margins, although damage makes comparisons difficult. Accordingly, Minqaria is recovered within Arenysaurini, a lambeosaurine clade endemic to Europe and North Africa.
The humerus and femur appear to represent lambeosaurines. The humerus resembles Hadrosauridae, including Lambeosaurinae, in having a concave distal edge to the deltopectoral crest; the weak sigmoidal curvature of the shaft resembles Lambeosaurinae. Neither is unique to Lambeosaurinae, but this character combination seems to be unique to lambeosaurines. The femur resembles Hadrosauridae in having a straight shaft in anterior view. The sigmoidal curvature seen in lateral view is seen in a variety of taxa, but is consistent with referral to Lambeosaurinae. The long, low fourth trochanter suggests affinities with Arenysaurini. Neither the humerus nor femur are diagnostic, and are referred to Lambeosaurinae indet. Given their large size, they are likely distinct from either Ajnabia or Minqaria, but additional fossils are required to test this hypothesis.
Phylogeny and biogeography
Although the lambeosaurine affinities of the Moroccan hadrosaur Ajnabia have been questioned83, the new material corroborates the presence of Lambeosaurinae in Africa. Other analyses have recovered Ajnabia as a lambeosaurine84,85 but not a monophyletic clade of European and African species. The differences result from inclusion of novel characters (e.g. the triangular predentary process of the mandible) and recodings of characters of the jaws and teeth that were found to unite Arenysaurini. We argue the results found here- with small-bodied European and African lambeosaurines representing a single lineage- are more parsimonious than the alternatives, which require multiple overwater dispersals to colonize Iberia, long ghost-lineages, and repeated evolution of small body size. The simplest explanation of all the data is that European and African lambeosaurines represent one lineage. More complete remains of the African and European arenysaurins are needed to better resolve their systematics, however.
Our phylogenetic analysis suggests Lambeosaurinae initially diversified in Asia, then saw dispersals into North America by Parasaurolophini and Lambeosaurini. Dispersal from Asia into Europe was followed by dispersal into North Africa. Multiple dispersals from Europe to Africa seem less parsimonious than a single dispersal (Fig. 10), but are not impossible, particularly given the distinctive morphologies of Ajnabia and Minqaria.

Evolutionary tree of Lambeosaurinae. European and African lambeosaurines are placed in Arenysaurini, a basally-diverging group of lambeosaurines.
The precise number of dispersals in hadrosauroids remains unclear due to conflicting tree topologies. The existence of basal hadrosauroids and lambeosaurines in Europe suggests at least two dispersals7, followed by dispersal of lambeosaurines into Africa7. The discovery of basal hadrosauroids in South America suggests two dispersals into South America, one by basal hadrosauroids83 and one by kritosaurins86,87, which dispersed into Antarctica86. Hadrosaurines dispersed into Appalachia; reinterpretation of Lophorhothon as a basal hadrosauroid suggests saurolophines did not disperse into Appalachia88. Absent any strong evidence for land bridges, and given the limited number of dispersals of dinosaurs between these land masses, the simplest hypothesis is that hadrosauroids colonized Europe, Africa, Appalachia, South America and Antarctica via oceanic dispersal7.
Diversity of North African lambeosaurines. The discovery of Ajnabia was surprising given that, during the Cretaceous, hundreds of kilometers of water separated North Africa from Eurasia. The new lambeosaurine fossils not only confirm the existence of lambeosaurines in North Africa, but shows they were diverse. Although Minqaria is similar in size to Ajnabia, differences in the shape of the maxilla, the position of the jugal contact, and especially the tooth morphology suggest that it likely occupied a distinct ecological niche. Meanwhile, the relatively larger size of the Daoui and Mrah Lahrach lambeosaurines suggests they occupied a niche distinct from the smaller Ajnabia and Minqaria. The different morphologies and sizes (Fig. 11) suggest that we are sampling a diverse radiation (Fig. 10). The relatively robust femur of the Mrah Lahrach animal is also curious suggesting an unusual locomotor strategy and perhaps ecology.

Late Maastrichtian dinosaurs of the latest Cretaceous Phosphates of Morocco.
Given the long distance and deep ocean channels separating North Africa from Europe89, a single dispersal seems more likely than multiple dispersals. If so, Lambeosaurinae may have arrived in the Maastrichtian or Campanian, possibly during the Campanian–Maastrichtian lowstand event, then diversified over several million years. A possible hadrosaurid from the early Maastrichtian of Angola would potentially constrain lambeosaurine dispersal to the early Maastrichtian or late Campanian5, if it does represent a hadrosaurid. Given the rapid rates of speciation seen in colonizing lineages such as Galapagos finches90 and island anoles91, and high rates of turnover seen in herbivorous dinosaurs92,93 lambeosaurines could have produced a high diversity of species in just a few million years. Few other assemblages have a comparably high diversity of lambeosaurines, the exception being the Dinosaur Park Formation of Alberta, which for a brief interval contained the genera Lambeosaurus, Corythosaurus, and Parasaurolophus93, but what is remarkable about Morocco is that it shows high diversity despite far more limited sampling, suggesting that it draws from a highly diverse fauna.
Absence of competing ornithischians may have driven diversification. In contrast to North America or Asia, where hadrosaurids and ornithischians were highly diverse, no ornithischians other than lambeosaurines are known from the Maastrichtian of Morocco. Similarly, diversity of the European Lambeosaurinae may be high, with a wide range of jaw morphologies and species occurring, because hadrosaurids in Europe had limited competition, with relatively few other ornithischian lineages present. These included rhabdodontid iguanodontians94, basal hadrosauroids95,96,97, and struthiosaurine ankylosaurs98,99. North American lambeosaurines competed with multiple lineages of hadrosaurids as well as other ornithischians such as thescelosaurids, pachycephalosaurids, leptoceratopsids, nodosaurids and ankylosaurids, as well as herbivorous coelurosaurs such as ornithomimids, deinocheirids, caenagnathids, and possibly troodontids100.
By the latest Maastrichtian, lambeosaurines saw regional extinction in North America, disappearing from the northern Great Plains, but persisting in the south101. Meanwhile, lambeosaurines in Europe and especially the Ibero-Armorican landmass became highly diverse at the end of the Cretaceous44,102,103. A comparable diversification appears to have taken place in North Africa. These patterns emphasize the highly local nature of dinosaur diversity; local extinctions or radiations may not capture global patterns in dinosaur diversification and extinction.
The new fossils confirm that African hadrosaurids were small by hadrosaurid standards. The femur suggests an animal around 4.5 m in length and weighing ~ 1000 kg; the humerus suggests a substantially larger animal. This is large by the standards of extant mammals, but small by the standards of hadrosaurids. Minqaria was a smaller animal, around 3.5 m in length, which suggests a weight of around 250 kg, but the fusion of the braincase indicates it was somatically mature. Ajnabia was likely similar in size given the mature bone texture of the holotype.
Iberian arenysaurins resembled African forms in being relatively small. Small species include indeterminate species from Lleida61,104 falling within the size range of Minqaria and Ajnabia. In addition, very small lambeosaurine remains belonging to mature individuals of about 2 m body length have been described from latest Cretaceous sites of the Spanish Pyrenees105. The Basturs Poble lambeosaurine is somewhat larger, with a femur length of 645 mm, comparable to the larger hadrosaurids known from Morocco.
Meanwhile, in North America and Asia, hadrosaurids evolved large and giant forms among Lambeosaurini43, Parasaurolophini, Edmontosaurini106, and Kritosaurini74,107. The lambeosaurin Corythosaurus weighed ~ 3.5 tons and the giant Magnapaulia grew significantly larger; the parasaurolophin Parasaurolophus walkeri exceeded 5 tons76; Gryposaurus latidens reached 4 tons76, and a late Maastrichtian gryposaur from the Naashoibito shows some kritosaurs grew larger. Saurolophus osborni approached 7 tons76; a related Hell Creek form grew larger108. The edmontosaurin Edmontosaurus reached almost 8 tons76; in Asia, Shantungosaurus reached 17 tons76. Strikingly, small-bodied hadrosaurids (< 1000 kg) are unknown from latest Cretaceous North America and Asia.
While hadrosaurids in North America and Asia occupied megaherbivore niches, Moroccan and European lambeosaurines occupied small- and medium-sized herbivore niches. These patterns might result from the low diversity of competing small herbivores, and titanosaurian sauropods outcompeting lambeosaurines for megaherbivore niches.
Curiously, ornithischians occur in the latest Cretaceous of East Africa and Oman but do not appear to represent lambeosaurines. Ornithopods found in the Maastrichtian of Kenya represent non-hadrosaurid ornithopods (J. Sertich, pers. comm. 2023). Likewise, hadrosauroids from Oman6 represent non-hadrosaurids (D. Baastians, pers. comm. 2023). That lambeosaurines are (so far) unknown from East Africa or the Arabian plate could result from geographic barriers to dispersal. It is conceivable that the Trans-Saharan Seaway may have divided Africa into a series of smaller landmasses39, with endemic dinosaur faunas, but sampling remains and issue and more dinosaur fossils are needed from Africa to test this hypothesis.
Conclusions
A hadrosaurid from the latest Maastrichtian phosphates of Sidi Chennane, Morocco, is distinct from Ajnabia odysseus and represents a new arenysaurin, Minqaria bata. Minqaria differs from Ajnabia in jaw and tooth morphology, suggesting it occupied a distinct niche. Fusion of cranial elements shows that it was mature despite its small size (~ 3.5 m), confirming the existence of small hadrosaurids in North Africa. Similarities with the European Arenysaurus provide further evidence for dispersal of lambeosaurines between the Ibero-Armorican landmass and Africa. A humerus from Sidi Daoui and a femur from Mrah Lahrach belong to larger individuals, suggesting at least three hadrosaur species occur in the phosphates. Even as lambeosaurines declined in the Maastrichtian of North America, they diversified in Africa.
Methods
Phylogenetic analysis was conducted using a modified version of the character-taxon matrix from Longrich et al.7, which is derived from the matrix Kobayashi et al.63. Characters are from Xing et al.109 Kobayashi et al.63, and Longrich et al.7 and 14 new characters were added to help resolve lambeosaurine relationships, for a total of 380 morphological characters (see SI). Two characters were excluded from analysis (87 is made redundant by two new characters; 193 was added by a previous analysis but the character description was not specified). Geographic distribution was included in addition to morphology, because biogeography shows strong phylogenetic signal, with closely related species inhabiting the same landmass110. Biogeography was coded as a series of binary characters rather than a single, multi-state character7 since this allows implied weighting to assign each biogeographic character its own weight, rather than to assume all dispersals are equally probable. Four taxa were added; Minqaria bata, the Daoui hadrosaurid, the Sidi Chennane hadrosaurid, and Tlatolophus galorum.
Analyses were run in TNT111, using a new technology search algorithm and default settings for implied weighting (K = 3), and a strict consensus was estimated; analysis was repeated in PAUP 4.0 b10 in implied weighting and the same K = 3 under a heuristic search until 100,000 trees were recovered; this produced an identical strict consensus. TNT returned 7 trees with a score of 120.23435. The resulting strict consensus (Fig. 10) has a CI of 0.3669, an RI of 0.8244, and a rescaled consistency index of 0.3025 (for full tree, see SI).
Data availability
All data generated or analyzed during this study are available in the paper or as part of the associated supplementary information.
References
Sallam, H. M. et al. New Egyptian sauropod reveals Late Cretaceous dinosaur dispersal between Europe and Africa. Nat. Ecol. Evol. 2, 445 (2018).
Gemmellaro, M. Rettili Maëstrichtiani di Egitto (Arti graf. G. Castiglia, 1921).
Sallam, H. M. et al. Vertebrate paleontological exploration of the Upper Cretaceous succession in the Dakhla and Kharga Oases, Western Desert, Egypt. J. Afr. Earth Sci. 117, 223–234 (2016).
Salem, B. S. et al. Dinosaur remains from the Upper Cretaceous (Campanian) of the Western Desert, Egypt. Cretaceous Res. 123, 104783 (2021).
Mateus, O. et al. In V Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno 71–105 (Actas, Salas de los Infantes, 2012).
Buffetaut, E., Hartman, A.-F., Al-Kindi, M. & Schulp, A. S. Hadrosauroid Dinosaurs from the Late Cretaceous of the Sultanate of Oman. PLoS ONE 10, e0142692 (2015).
Longrich, N. R., Suberbiola, X. P., Pyron, R. A. & Jalil, N.-E. The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography. Cretac. Res. 120, 104678 (2021).
Longrich, N. R., Pereda-Suberbiola, X., Jalil, N.-E., Khaldoune, F. & Jourani, E. An abelisaurid from the latest Cretaceous (late Maastrichtian) of Morocco, North Africa. Cretaceous Res. 76, 40–52 (2017).
Pereda-Suberbiola, X., Bardet, N., Iarochène, M., Bouya, B. & Amaghzaz, M. The first record of a sauropod dinosaur from the Late Cretaceous phosphates of Morocco. J. Afr. Earth Sc. 40, 81–88 (2004).
Buffetaut, E., Escuillié, F. & Pohl, B. First theropod dinosaur from the Maastrichtian phosphates of Morocco. Kaupia 14, 3–8 (2005).
Horner, J. R., Weishampel, D. B. & Forster, C. A. In The Dinosauria (eds. David, B. W. et al.) 438–463 (University of California Press, 2004).
Prieto-Márquez, A. Global historical biogeography of hadrosaurid dinosaurs. Zool. J. Linnean Soc. 159, 503–525 (2010).
Eberth, D. A. & Evans, D. C. Hadrosaurs (Indiana University Press, 2014).
Blakey, R. C., Fielding, C., Frank, T. & Isbell, J. Gondwana paleogeography from assembly to breakup—a 500 my odyssey. Geol. Soc. Am. Spec. Pap. 441, 1–28 (2008).
Kocsis, L. et al. Comprehensive stable isotope investigation of marine biogenic apatite from the Late Cretaceous-Early Eocene phosphate series of Morocco. Palaeogeogr. Palaeoclimatol. Palaeoecol. 394, 74–88 (2014).
Yans, J. et al. First carbon isotope chemostratigraphy of the Ouled Abdoun phosphate Basin, Morocco; implications for dating and evolution of earliest African placental mammals. Gondwana Res. 25, 257–269 (2014).
Arambourg, C. Les vertébrés fossiles des gisements de phosphates (Maroc-Algérie-Tunisie). Notes Mémoires du Serv. Géol. Maroc. 92, 1–372 (1952).
Noubhani, A. & Cappetta, H. Les Orectolobiformes, Carcharhiniformes et Myliobatiformes (Elasmobranchii, Neoselachii) des Bassins à phosphate du Maroc (Maastrichtien-Lutétien basal): Systématique, biostratigraphie, évolution et dynamique des faunes. Palaeo Ichthyol. 8, 1–327 (1997).
Martin, J. E. et al. Calcium isotopic evidence for vulnerable marine ecosystem structure prior to the K/Pg extinction. Curr. Biol. 27, 1641–1644 (2017).
Bardet, N., Pereda-Suberbiola, X., Iarochène, M., Amalik, M. & Bouya, B. Durophagous Mosasauridae (Squamata) from the Upper Cretaceous phosphates of Morocco, with description of a new species of Globidens. Netherl. J. Geosci. 84, 167–175 (2005).
Bardet, N., Pereda-Suberbiola, X., Iarochène, M., Bouya, B. & Amaghzaz, M. A new species of Halisaurus from the Late Cretaceous phosphates of Morocco, and the phylogenetical relationships of the Halisaurinae (Squamata: Mosasauridae). Zool. J. Linnean Soc. 143, 447–472 (2005).
Schulp, A. S., Bardet, N. & Bouya, B. A new species of the durophagous mosasaur Carinodens (Squamata, Mosasauridae) and additional material of Carinodens belgicus from the Maastrichtian phosphates of Morocco. Netherl. J. Geosci. 88, 161–167 (2009).
Strong, C. R., Caldwell, M. W., Konishi, T. & Palci, A. A new species of longirostrine plioplatecarpine mosasaur (Squamata: Mosasauridae) from the Late Cretaceous of Morocco, with a re-evaluation of the problematic taxon ‘Platecarpus’ ptychodon. J. Syst. Palaeontol. 2020, 1–36 (2020).
LeBlanc, A. R. H., Caldwell, M. W. & Bardet, N. A new mosasaurine from the Maastrichtian (Upper Cretaceous) phosphates of Morocco and its implications for mosasaurine systematics. J. Vertebr. Paleontol. 32, 82–104 (2012).
Bardet, N. et al. Mosasaurids (Squamata) from the Maastrichtian phosphates of Morocco: Biodiversity, palaeobiogeography and palaeoecology based on tooth morphoguilds. Gondwana Res. 27, 1068–1078 (2015).
Bardet, N. et al. Mosasaurus beaugei Arambourg, 1952 (Squamata, Mosasauridae) from the Late Cretaceous phosphates of Morocco. Geobios 37, 315–324 (2004).
Longrich, N. R., Bardet, N., Schulp, A. S. & Jalil, N.-E. Xenodens calminechari gen. et. sp. nov., a bizarre mosasaurid (Mosasauridae, Squamata) with shark-like cutting teeth from the upper Maastrichtian of Morocco, North Africa. Cretaceous Res. 2021, 104764 (2021).
Longrich, N. R., Bardet, N., Khaldoune, F., Yazami, O. K. & Jalil, N.-E. Pluridens serpentis, a new mosasaurid (Mosasauridae: Halisaurinae) from the Maastrichtian of Morocco and implications for mosasaur diversity. Cretaceous Res. 2021, 104882 (2021).
Longrich, N. R. et al. Thalassotitan atrox, a giant predatory mosasaurid (Squamata) from the Upper Maastrichtian Phosphates of Morocco. Cretaceous Res. 140, 105315 (2022).
Rempert, T. H., Martens, B. P. & Melchers, A. P. V. First record of a tylosaurine mosasaur from the latest cretaceous phosphates of Morocco. Open J. Geol. 12, 883–906 (2022).
Bardet, N. et al. A giant chelonioid turtle from the late cretaceous of Morocco with a suction feeding apparatus unique among tetrapods. PLOS One 8, e63586. https://doi.org/10.1371/journal.pone.0063586 (2013).
Lapparent-de-Broin, F. D., Bardet, N., Amaghzaz, M. & Meslouh, S. A strange new chelonioid turtle from the Latest Cretaceous phosphates of Morocco. Compt. Rend. Palevol. 13, 87–95. https://doi.org/10.1016/j.crpv.2013.07.008 (2013).
Houssaye, A. et al. A review of Pachyvaranus crassispondylus Arambourg, 1952, a pachyostotic marine squamate from the latest Cretaceous phosphates of Morocco and Syria. Geol. Mag. 148, 237–249 (2011).
Vincent, P., Bardet, N., Houssaye, A., Amaghzaz, M. & Meslouh, S. New plesiosaur specimens from the Maastrichtian Phosphates of Morocco and their implications for the ecology of the latest Cretaceous marine apex predators. Gondwana Res. 24, 796–805 (2013).
Vincent, P. et al. Zarafasaura oceanis, a new elasmosaurid (Reptilia: Sauropterygia) from the Maastrichtian Phosphates of Morocco and the palaeobiogeography of latest Cretaceous plesiosaurs. Gondwana Res. 19, 1062–1073 (2011).
Jouve, S. et al. The oldest African crocodylian: phylogeny, paleobiogeography, and differential survivorship of marine reptiles through the Cretaceous-Tertiary boundary. J. Vertebr. Paleontol. 28, 409–421 (2008).
Longrich, N. R., Martill, D. M. & Andres, B. Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. PLOS Biol. 16, e2001663 (2018).
Pereda-Suberbiola, X. et al. A new azhdarchid pterosaur from the Late Cretaceous phosphates of Morocco. Geol. Soc. Lond. Spec. Publ. 217, 79–90 (2003).
Longrich, N. R., Isasmendi, E., Pereda-Suberbiola, X. & Jalil, N.-E. New fossils of Abelisauridae (Dinosauria: Theropoda) from the upper Maastrichtian of Morocco, North Africa. Cretaceous Res. 152, 105677 (2023).
Evans, D. C. Cranial anatomy and systematics of Hypacrosaurus altispinus, and a comparative analysis of skull growth in lambeosaurine hadrosaurids (Dinosauria: Ornithischia). Zool. J. Linnean Soc. 159, 398–434 (2010).
Godefroit, P., Bolotsky, Y. L. & Itterbeeck, J. V. The lambeosaurine dinosaur Amurosaurus riabinini from the Maastrichtian of Far Eastern Russia. Acta Palaeontol. Polon. 49, 585–618 (2004).
Godefroit, P., Bolotsky, Y. L. & Bolotsky, I. Y. Osteology and relationships of Olorotitan arharensis, a hollow-crested hadrosaurid dinosaur from the latest Cretaceous of Far Eastern Russia. Acta Palaeontol. Polon. 57, 527–561 (2012).
Prieto-Márquez, A., Chiappe, L. M. & Joshi, S. H. The lambeosaurine dinosaur Magnapaulia laticaudus from the Late Cretaceous of Baja California, Northwestern Mexico. PLOS One 7, e38207 (2012).
Prieto-Márquez, A., Dalla Vecchia, F. M., Gaete, R. & Galobart, À. Diversity, relationships, and biogeography of the lambeosaurine dinosaurs from the European archipelago, with description of the new aralosaurin Canardia garonnensis. PLOS One 8, e69835 (2013).
Wagner, J. R. & Lehman, T. M. An enigmatic new lambeosaurine hadrosaur (Reptilia: Dinosauria) from the Upper Shale Member of the Campanian Aguja Formation of Trans-Pecos Texas. J. Vertebr. Paleontol. 29, 605–611 (2009).
Takasaki, R. et al. Description of the first definitive Corythosaurus (Dinosauria, Hadrosauridae) specimens from the Judith River Formation in Montana, USA and their paleobiogeographical significance. Anatom. Record 2022, 895 (2022).
Brown, B. Corythosaurus casuarius, a new crested dinosaur from the Belly River Cretaceous; with Provisional classification of the family Trachodontidae. Bull. AMNH 33, 35 (1914).
Prieto-Márquez, A. & Wagner, J. R. Pararhabdodon isonensis and Tsintaosaurus spinorhinus: A new clade of lambeosaurine hadrosaurids from Eurasia. Cretaceous Res. 30, 1238–1246 (2009).
Cruzado-Caballero, P., Pereda-Suberbiola, X. & Ruiz-Omeñaca, J. I. Blasisaurus canudoi gen. et sp nov., a new lambeosaurine dinosaur (Hadrosauridae) from the Latest Cretaceous of Arén (Huesca, Spain). Can. J. Earth Sci. 47, 1507–1517 (2010).
Pereda-Suberbiola, X. et al. The last hadrosaurid dinosaurs of Europe: A new lambeosaurine from the uppermost Cretaceous of Arén (Huesca, Spain). Compt. Rend. Palevol. 8, 559–572 (2009).
Sullivan, R. M. & Williamson, T. E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a Revision of the Genus. Compt. Rend. Palevol. 15, 1–52 (1999).
Dudgeon, T. W. & Evans, D. C. Calvarial suture interdigitation in hadrosaurids (Ornithischia: Ornithopoda): Perspectives through ontogeny and evolution. Evol. Dev. 25, 209–225 (2023).
Gates, T. A., Evans, D. C. & Sertich, J. J. Description and rediagnosis of the crested hadrosaurid (Ornithopoda) dinosaur Parasaurolophus cyrtocristatus on the basis of new cranial remains. PeerJ 9, e10669 (2021).
Godefroit, P., Zan, S. & Jin, L. Charonosaurus jiayinensis ng, n. sp., a lambeosaurine dinosaur from the Late Maastrichtian of northeastern China. Compt. Rend. Acad. Sci.-Ser. IIA-Earth Planet. Sci. 330, 875–882 (2000).
Prieto-Márquez, A., Erickson, G. M. & Ebersole, J. A. Anatomy and osteohistology of the basal hadrosaurid dinosaur Eotrachodon from the uppermost Santonian (Cretaceous) of southern Appalachia. PeerJ 4, e1872 (2016).
Xing, H., Mallon, J. C. & Currie, M. L. Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae. PloS one 12, e0175253 (2017).
Prieto-Marquez, A. New information on the cranium of Brachylophosaurus canadensis (Dinosauria, Hadrosauridae), with a revision of its phylogenetic position. J. Vertebr. Paleontol. 25, 144–156 (2005).
Prieto-Marquez, A., Gaete, R., Rivas, G., Galobart, Á. & Boada, M. Hadrosauroid dinosaurs from the Late Cretaceous of Spain: Pararhabdodon isonensis revisited and Koutalisaurus kohlerorum, gen. et sp. nov.. J. Vertebr. Paleontol. 26, 929–943 (2006).
Godefroit, P., Shulin, H., Tingxiang, Y. & Lauters, P. New hadrosaurid dinosaurs from the uppermost Cretaceous of northeastern China. Acta Palaeontol. Polon. 53, 47–74 (2008).
Paul, G. S. Dinosaur models: The good, the bad, and using them to estimate the mass of dinosaurs. In DinoFest International: Proceedings of a Symposium Sponsored by Arizona State University. Academy of Natural Sciences, Philadelphia 129–154 (1997).
Fondevilla, V. et al. Ontogeny and taxonomy of the hadrosaur (Dinosauria, Ornithopoda) remains from Basturs Poble bonebed (late early Maastrichtian, Tremp Syncline, Spain). PLOS One 13, e0206287 (2018).
Maryanska, T. & Osmólska, H. Postcranial anatomy of Saurolophus angustirostris with comments on other hadrosaurs. Palaeontol. Polon. 46, 119–141 (1984).
Kobayashi, Y. et al. A new hadrosaurine (Dinosauria: Hadrosauridae) from the marine deposits of the late cretaceous Hakobuchi Formation, Yezo Group, Japan. Sci. Rep. 9, 1–14 (2019).
Tsogtbaatar, K., Weishampel, D. B., Evans, D. C. & Watabe, M. A new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous Baynshire Formation of the Gobi Desert (Mongolia). PLOS One 14, e0208480 (2019).
Prieto-Marquez, A., Weishampel, D. B. & Horner, J. R. The dinosaur Hadrosaurus foulkii, from the Campanian of the East Coast of North America, with a reevaluation of the genus. Acta Palaeontol. Polon. 51, 77–98 (2006).
Prieto-Márquez, A. & Norell, M. A. Anatomy and relationships of Gilmoreosaurus mongoliensis (Dinosauria: Hadrosauroidea) from the Late Cretaceous of Central Asia. Am. Museum Novit. 2010, 1–49 (2010).
Prieto-Márquez, A., Fondevilla, V., Sellés, A. G., Wagner, J. R. & Galobart, À. Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European Archipelago. Cretaceous Res. 2018, 859 (2018).
Dalla Vecchia, F. M. Telmatosaurus and the other hadrosaurids of the Cretaceous European Archipelago. An overview. Nat. Nascosta 32, 1–55 (2006).
Madzia, D., Jagt, J. W. & Mulder, E. W. Osteology, phylogenetic affinities and taxonomic status of the enigmatic late Maastrichtian ornithopod taxon Orthomerus dolloi (Dinosauria, Ornithischia). Cretaceous Res. 108, 104334 (2020).
Prieto-Márquez, A. A re-evaluation of Secernosaurus koerneri and Kritosaurus australis (Dinosauria, Hadrosauridae) from the Late Cretaceous of Argentina. J. Vertebr. Paleontol. 30, 813–837 (2010).
Brown, B. The skeleton of Saurolophus, a crested duck-billed dinosaur from the Edmonton Cretaceous. Bull. AMNH 32, 19 (1913).
Prieto-Márquez, A. Cranial and appendicular ontogeny of Bactrosaurus johnsoni, a hadrosauroid dinosaur from the Late Cretaceous of northern China. Palaeontology 54, 773–792 (2011).
Jagt, J. W., Mulder, E. W., Schulp, A. S., Dortangs, R. W. & Fraaije, R. H. Dinosaurs from the Maastrichtian-type area (southeastern Netherlands, northeastern Belgium). Compt. Rend. Palevol. 2, 67–76 (2003).
Kirkland, J. I. et al. Large hadrosaurine dinosaurs from the latest Campanian of Coahuila, Mexico. New Mexico Museum Nat. Hist. Sci. Bull. 35, 299–315 (2006).
Campione, N. E. & Evans, D. C. A universal scaling relationship between body mass and proximal limb bone dimensions in quadrupedal terrestrial tetrapods. BMC Biol. 10, 1–22 (2012).
Benson, R. B. et al. Rates of dinosaur body mass evolution indicate 170 million years of sustained ecological innovation on the avian stem lineage. PLOS Biol. 12, 856 (2014).
Campione, N. E. Extrapolating body masses in large terrestrial vertebrates. Paleobiology 43, 693–699 (2017).
Longrich, N. R. & Field, D. Torosaurus is not Triceratops: Ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy. PLOS One 7, e32623326. https://doi.org/10.31371/journal.pone.0032623 (2012).
Longrich, N. R. & Saitta, E. T. Taxonomic status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—a distinct taxon of small-bodied tyrannosaur. Fossil Stud. 2, 1–65 (2024).
Gates, T. A. et al. Velafrons coahuilensis, a new lambeosaurine hadrosaurid (Dinosauria: Ornithopoda) from the Late Campanian Cerro Del Pueblo Formation, Coahila, Mexico. J. Vertebr. Paleontol. 27, 917–930 (2007).
Weishampel, D. B. & Horner, J. R. In The Dinosauria Vol. 534–561 (eds. WEishampel, D. B. et al.) (University of California Press, 1990).
Evans, D. C., Forster, C. A. & Reisz, R. R. In Dinosaur Provincial Park, A Spectacular Ancient Ecosystem Revealed (eds. Currie, P. J. & Koppelhus, E. B.) 349–366 (2005).
Rozadilla, S., Agnolín, F., Manabe, M., Tsuihiji, T. & Novas, F. E. Ornithischian remains from the Chorrillo formation (Upper Cretaceous), southern Patagonia, Argentina, and their implications on ornithischian paleobiogeography in the Southern Hemisphere. Cretaceous Res. 125, 104881 (2021).
Ramírez-Velasco, A. A. Phylogenetic and biogeography analysis of Mexican hadrosauroids. Cretaceous Res. 2022, 105267 (2022).
Serrano-Brañas, C. I. & Prieto-Márquez, A. Taphonomic attributes of the holotype of the lambeosaurine dinosaur Latirhinus uitstlani from the late Campanian of Mexico: Implications for its phylogenetic systematics. J. South Am. Earth Sci. 114, 103689 (2022).
Ibiricu, L. M. et al. New hadrosaurid (Dinosauria: Ornithopoda) fossils from the uppermost Cretaceous of central Patagonia and the influence of paleoenvironment on South American hadrosaur distribution. J. South Am. Earth Sci. 110, 103369 (2021).
Rozadilla, S., Brissón-Egli, F., Agnolín, F. L., Aranciaga-Rolando, A. M. & Novas, F. E. A new hadrosaurid (Dinosauria: Ornithischia) from the Late Cretaceous of northern Patagonia and the radiation of South American hadrosaurids. J. Syst. Palaeontol. 19, 1207–1235 (2021).
Gates, T. A. & Lamb, J. P. A redescription of Lophorhothon atopus (Ornithopoda: Dinosauria) from the Late Cretaceous of Alabama based on new material1. Can. J. Earth Sci. 58, 918–935 (2021).
Dercourt, J., Gaetani, M. & Vrielynck, B. Atlas Peri-Tethys Palaeogeographical Maps. (CCGM, 2000).
Al-Mosleh, S., Choi, G. P., Abzhanov, A. & Mahadevan, L. Geometry and dynamics link form, function, and evolution of finch beaks. Proc. Natl. Acad. Sci. 118, e2105957118 (2021).
Patton, A. H. et al. When adaptive radiations collide: Different evolutionary trajectories between and within island and mainland lizard clades. Proc. Natl. Acad. Sci. 118, e2024451118 (2021).
Sampson, S. D. et al. New horned dinosaurs from Utah provide evidence for intracontinental dinosaur endemism. PLOS One 5, e12292 (2010).
Mallon, J. C., Evans, D. C., Ryan, M. J. & Anderson, J. S. Megaherbivorous dinosaur turnover in the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 350–352, 124–138 (2012).
Weishampel, D. B., Jianu, C. M., Csiki, Z. & Norman, D. B. Osteology and phylogeny of Zalmoxes (ng), an unusual euornithopod dinosaur from the latest Cretaceous of Romania. J. Syst. Palaeontol. 1, 65–123 (2003).
Weishampel, D., Norman, D. & Grigorescu, D. Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: The most basal hadrosaurid dinosaur. Palaeontology 36, 361–385 (1993).
Dalla Vecchia, F. M. Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy. J. Vertebr. Paleontol. 29, 1100–1116 (2009).
Prieto-Márquez, A. & Farias, C. A new late-surviving early diverging Ibero-Armorican duck-billed dinosaur and the role of the Late Cretaceous European Archipelago in hadrosauroid biogeography. Acta Palaeontol. Polon. 66, 8596 (2021).
Garcia, G. & Pereda-Suberbiola, X. A new species of Struthiosaurus (Dinosauria: Ankylosauria) from the Upper Cretaceous of Villeveyrac (southern France). J. Vertebr. Paleontol. 23, 156–165 (2003).
Nopcsa, F. Die Dinosaurier der siebenbürgischen Landesteile Ungarns: Von Franz Baron Nopcsa. Mit Tafel I-IV und 3 Figuren im Texte.[Umschlagtitel.] (Buchdruckerei des Franklin-Vereins, 1915).
Weishampel, D. B. et al. In The Dinosauria (edss David B. W. et al.) 517–606 (University of California Press, 2004).
Sullivan, R. M., Jasinski, S. E., Guenther, M. F. & Lucas, S. G. The first lambeosaurin (Dinosauria, Hadrosauridae, Lambeosaurinae) from the Upper Cretaceous Ojo Alamo formation (Naashoibito Member), San Juan Basin, New Mexico. New Mexico Museum Nat. Hist. Sci. Bull. 53, 405–417 (2011).
Vila, B., Sellés, A. G. & Brusatte, S. L. Diversity and faunal changes in the latest Cretaceous dinosaur communities of southwestern Europe. Cretaceous Res. 57, 552–564 (2016).
Blanco, A., Prieto-Márquez, A. & De Esteban-Trivigno, S. Diversity of hadrosauroid dinosaurs from the Late Cretaceous Ibero-Armorican Island (European Archipelago) assessed from dentary morphology. Cretaceous Res. 56, 447–457 (2015).
Conti, S. et al. The oldest lambeosaurine dinosaur from Europe: Insights into the arrival of Tsintaosaurini. Cretaceous Res. 107, 104286 (2020).
Cruzado-Caballero, P. & Canudo, J. Presence of diminutive hadrosaurids (Dinosauria: Ornithopoda) in the Maastrichtian of the south-central Pyrenees (Spain). J. Iber. Geol. 41, 71–81 (2015).
Xing, H. et al. Comparative osteology and phylogenetic relationship of Edmontosaurus and Shantungosaurus (Dinosauria: Hadrosauridae) from the Upper Cretaceous of North America and East Asia. Acta Geol. Sin. (Engl. Ed.) 88, 1623–1652 (2014).
Prieto-Márquez, A. Skeletal morphology of Kritosaurus navajovius (Dinosauria: Hadrosauridae) from the Late Cretaceous of the North American south-west, with an evaluation of the phylogenetic systematics and biogeography of Kritosaurini. J. Syst. Palaeontol. 12, 133–175 (2014).
Evangelista, A., Campione, N. & Scannella, J. In Society of Vertebrate Paleontology Annual Meeting 160 (2023).
Xing, H. et al. A new basal hadrosauroid dinosaur (Dinosauria: Ornithopoda) with transitional features from the Late Cretaceous of Henan Province, China. PLOS One 9, e98821 (2014).
Darwin, C. R. The Origin of Species (John Murray, 1859).
Goloboff, P. A., Farris, J. S. & Nixon, K. C. TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786 (2008).
Acknowledgements
Thanks to Mustapha Meharich for assistance to NL with translation and logistics in Oulad Bou Ali. Research of XPS is financed by the Spanish Ministry of Science and Innovation (MCIN) and the European Regional Development Fund (FEDER) (research project PID2021-122612OB-I00), and by the Basque Country Government (research group IT1485-22). Thanks to the reviewers.
Author information
Authors and Affiliations
Contributions
N.R.L., X.P.S., N.B. and N.E.J. wrote the manuscript. N.L. prepared the figures and performed the phylogenetic analysis. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Longrich, N.R., Pereda-Suberbiola, X., Bardet, N. et al. A new small duckbilled dinosaur (Hadrosauridae: Lambeosaurinae) from Morocco and dinosaur diversity in the late Maastrichtian of North Africa. Sci Rep 14, 3665 (2024). https://doi.org/10.1038/s41598-024-53447-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-53447-9
- Springer Nature Limited