
Abstract
Cadmium is a highly neurotoxic heavy metal that interferes with DNA repair mechanisms via generation of reactive oxygen species. The potentials of polyphenols and antioxidants as effective protective agents following heavy metal-induced neurotoxicity are emerging. We therefore explored the neuroprotective potentials of gallic and ascorbic acids in CdCl2-induced neurotoxicity. Seventy-two Wistar rats were divided into six groups. Group A received distilled water, B: 3 mg/kg CdCl2, C: 3 mg/kg CdCl2 + 20 mg/kg gallic acid (GA), D: 3 mg/kg CdCl2 + 10 mg/kg ascorbic acid (AA), E: 20 mg/kg GA and F: 10 mg/kg AA orally for 21 days. Depression, anxiety, locomotion, learning and memory were assessed using a battery of tests. Neuronal structure and myelin expression were assessed with histological staining and immunofluorescence. The Morris Water Maze test revealed significant increase in escape latency in CdCl2 group relative to rats concurrently treated with GA or AA. Similarly, time spent in the target quadrant was reduced significantly in CdCl2 group relative to other groups. Concomitant administration of gallic acid led to significant reduction in the durations of immobility and freezing that were elevated in CdCl2 group during forced swim and open field tests respectively. Furthermore, GA and AA restored myelin integrity and neuronal loss observed in the CdCl2 group. We conclude that gallic and ascorbic acids enhance learning and memory, decrease anxiety and depressive-like behavior in CdCl2-induced neurotoxicity with accompanying myelin-protective ability.
Similar content being viewed by others

Introduction
Naturally occurring elements with atomic weight and density at least 5 times greater than that of water have been described as heavy metals1. Globally, heavy metal toxicity is a major health threat affecting plants, animals and humans2. Cadmium is a heavy metal that currently has no known physiological function and poses health risks for animals and humans3,4. It is carcinogenic and ranked seventh most toxic occupational and environmental heavy metal5,6. Aside from its use as alloys in electroplating and manufacturing of pigments, it is also combined with nickel to make batteries and a by-product during the production of other heavy metals such as lead, zinc and copper1,7,8.
Cadmium exposure in humans usually occurs as a result of ingestion of contaminated food and water, cigarette smoking and cigarette smoke inhalation2,4. Its rate of excretion in the body is extremely low causing it to accumulate in soft tissues leading to neurotoxicity, nephrotoxicity, hepatotoxicity, endocrine and reproductive toxicities9,10. Exposure can lead to acute or chronic intoxications2 causing reactive oxygen species generation, initiation of apoptosis and interferences with DNA repair7.
Cadmium-induced neurotoxicity has been documented to result in decreased equilibrium, vasomotor dysfunction, peripheral neuropathy, learning and memory deficits11. Cadmium neurotoxicity has also been associated with onset of several neurodegenerative diseases such as Parkinson’s and Alzheimer’s that are characterized by alteration in the permeability of the blood–brain barrier12.
Gallic acid (3, 4, 5-trihydroxybenzoic acid) is one of the most abundant phenolic acids present in various plants. Its derivatives can be found in various fruits such as lemon, bananas, pineapples and some plants13,14. It is known to have antioxidant, anti-inflammatory, antimicrobial, anticancer, gastroprotective, cardioprotective and neuroprotective properties15. It exerts its neuroprotective effects by preventing the formation of free radicals, inhibiting the production of cytokines and maintenance of calcium homeostasis thereby helping to maintain neuronal plasticity16. It has been reported to improve cognitive and motor impairments in Wistar rats16,17.
Similarly, ascorbic acid is an antioxidant that protects the body from various adverse effects of free radicals and toxins18. Furthermore, it has been documented to decrease oxidative stress and formation of protein aggregates in neurodegenerative disorders19. Despite several studies reporting the protective effects of these two antioxidants in Cd-induced nephrotoxicity, hepatotoxicity and reproductive toxicity in animals20,21,22, to the best of our knowledge, their roles in the protection of Cd-induced behavioral alterations and neuropathology are yet to be assessed. In this study, we therefore evaluated the neuro-protective effects of gallic and ascorbic acids against cadmium-induced neurotoxicity and behavioral alterations in rodent model.
Materials and methods
Experimental animals
A total of seventy-two Wistar rats (150–180 g), aged 8–10 weeks of both sexes were used for this study. The rats were housed under standard housing condition, fed regular pelletized diet and water ad libitum under a 12/12 h light/dark cycle.
Chemicals
Gallic acid (#20110905, Tianjin Kermel Chemical Reagents Limited, China), ascorbic acid (#2520, ChemoPharma Nigeria) and cadmium chloride (#02,416 LOBA-Chemie, India) and all other chemicals used in this study were of analytical grade.
Experimental design
The 72 experimental rats were randomly divided into six groups (A-F; n = 12/group; 6 males and 6 females/group). Group A (control) received distilled water only; Group B (CdCl2) received 3 mg/kg of cadmium chloride; Group C (CdCl2 + GA) 3 mg/kg cadmium chloride and 20 mg/kg of gallic acid; Group D (CdCl2 + AA) 3 mg/kg cadmium chloride and 10 mg/kg ascorbic acid; Group E (GA alone) 20 mg/kg gallic acid and Group F (AA alone) 10 mg/kg body weight ascorbic acid. All chemicals were dissolved in distilled water and treatments were administered in a single daily dose via oral gavage for 21 consecutive days (Fig. 1A).

(A) Schematic diagram of experimental groups (B) Outline of behavioral assessments.
Ethical statement
The National Institute of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80–23 revised, 1996) was strictly adhered to during the study. Efforts were also made to reduce pain and stress in the experimental animals. The University of Ibadan Animal Care and Use Ethics Committee approved the study (UI-ACUREC/18/0026) in line with ARRIVE guidelines.
Daily body weight change
The body weights of the rats were measured daily throughout the study. The weight on the first day of the study was regarded as the baseline weight. We thereafter calculated the percentage of initial body weight daily from the baseline weight of the animals as:
Morris water maze (MWM) test
The maze was a rounded water pool with concealed spherical hidden escape platform placed about two inches below the water surface23. The water was made opaque with powdered non-fat milk. The pool was marked north, south, east and west with the escape platform fixed at a specific location. Each rat was placed into the pool from different start points randomly from the 4 locations and allowed to search for the escape stage. The time spent to locate the escape platform was recorded. If the platform was not found after 120 s, the animals were directed towards it and allowed to stay on it for at least 15 s. Individual animals underwent four tests (corresponding to the four different start locations) per day for five uninterrupted days. The animals were allowed inter-trial rest of up to one hour. This represents the acquisition phase of the test. On the 6th day, a single probe test was carried out to assess spatial memory. During this phase, the escape platform was removed. The retention of the former position of the platform was measured by evaluating the period the animal spent in the target quadrant and the number of crosses within it. The time spent at the opposite of the target quadrant was also recorded and the test documented manually by two independent viewers blinded to the treatment protocol. The MWM commenced fifteen days after the onset of drug administration (Fig. 1B).
Forced swim test (FST)
The forced swim test (FST) is one of the most commonly used tests for depressive-like behavior in rodents24. Rats were placed in a container filled with clean water (24 ± 1 °C) and were monitored for six minutes25. The onset and duration of immobility were recorded using a timer. This test was performed on experimental day 18 (Fig. 1B).
Open field maze (OFM) test
The open field was used to assess anxiety and locomotion. The apparatus was a box measuring 72 × 72 cm × 36 cm high. One of the walls was covered with a plexiglass so the rats could be visible in the apparatus. The floor area was separated into 25 squares (20 × 20 cm), defined as 9 central and 16 peripheral squares. At the beginning of the test, each animal was placed in the center of the apparatus and then allowed to freely explore it for 5 min. The parameters recorded were line crossing which is the total number of squares visited (a measure of anxiety and locomotion), rearing (the number of times rat stood on its hind legs), grooming (number of times the animal spent licking or scratching itself while stationary), freezing (the duration with which the animal was completely stationary), center square duration (time spent in the central squares of the box), Stretched-Attend Posture (frequency with which the animal displayed forward elongation of the head and the shoulders followed by retraction to the original position) and defecation (the number of fecal boli). The OFM test was performed twenty (20) days after the onset of drug administration.
The elevated plus maze test
The elevated plus maze test was made of two open arms (50 × 10 cm) crossed at right angles with two opposing closed arms of equal sizes. Each animal was placed at the center of the maze, facing one of the closed arms, and allowed to freely explore the arena for 5 min. The anxiety-related measures taken include the frequency and duration with which the animals visited the open arms and the closed arms.
Forelimb support (hanging wire test)
A 2 mm thick metallic wire was tightly attached to a frame to avoid vibration or unwanted displacement of the wire during the test. Individual rat was placed on the 2 mm diameter hanging wire with its forelimb and monitored for a period of 2 min. The period of time it took the animal to stay on the wire before falling was recorded. Rats were tested twice and allowed about 2 h of rest in between. The hanging wire was performed twenty-one days after the commencement of drug administration.
In between testing, all the apparatus were cleaned with 70% ethyl alcohol to eliminate odor cues of previously tested rat. All behavioral tests were carried out by an experimenter blinded to the treatment groups.
Animal sacrifice
Twenty-four hours after the behavioral tests, each rat was deeply anesthetized with sodium pentobarbital (10 mg/100 g body weight) and transcardially perfused with physiological saline, followed by 4% paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.4) at 4 °C. Brains were removed and placed in 10% formaldehyde and 30% NBF solution for histopathology and immunohistochemistry respectively.
Haematoxylin and Eosin Staining
The brains were processed routinely for paraffin embedding. 5 μm sections were obtained with rotary microtome and processed for Haematoxylin and Eosin staining as previously documented26. Sections were observed with light microscope and photomicrograph taken with a Leica LAS-EZ Microscope installed with Leica software application suite version 3.4.
Luxol fast blue (LFB) staining
Brain sections (5 μm thick) were deparaffinized in xylene and thereafter hydrated to 5% ethanol. The sections were then incubated in LFB (#26681-01, Electron Microscopy Sciences, USA) solution (0.1% LFB in 95% ethanol, 5% methanol with 0.05% glacial acetic acid) at 56 °C for 16 h. Thereafter, the sections were differentiated with 0.05% lithium carbonate solution (#26681-04, Electron Microscopy Sciences, USA) for 30 s followed by 70% ethanol. The sections were further differentiated in 95% ethanol and rehydrated before mounting with Eukitt® Quick-hardening mounting medium (#03989, Sigma, USA).
Immunofluorescence
Immunofluorescence was performed by de-paraffinizing the brain sections in xylene and rehydration in graded percentages of alcohol. Antigen retrieval was done by placing the slides in 10 mM citrate buffer at 95–100 °C for 15 min. The slides were thereafter blocked in blocking buffer (2X casein/10% goat serum/0.1% Triton X-100 pH 7.4) and later incubated with the following primary antibodies: anti-NeuN polyclonal antibody (1:500; #MAB377, Millipore®), anti-MBP rabbit polyclonal antibody (1:500; #PA5-78397, Invitrogen®). The sections were rinsed 3 times (5 min each) in PBS and then incubated with secondary antibodies conjugated with Alexa dyes (AF-488, #A11001, Invitrogen and AF-647, #A21245, Life Technologies) at room temperature for 1 h. Sections were then washed 3 times (5 min each) followed by the addition of 2 drops of mounting media with DAPI (Invitrogen #P36935) before sections were cover slipped. The pixel intensities for MBP and NeuN images were analyzed using ImageJ software.
Statistical analysis
Data were analyzed using a one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test for comparison between groups. All tests were used with significance set at p < 0.05. The data were presented as mean ± standard deviation.
Data analysis indicated no sex differences in any of the parameters investigated above hence we combined data from males and females experimental rats.
Results
CdCl2 administration led to reduction in body weight
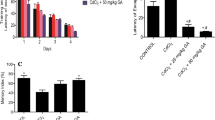
We observed a gradual increase in body weight across all the groups in the first nine days of this study. However, by day 10, 11 and 14 there was a decline in body weight gain in the CdCl2 + AA, CdCl2 + GA and CdCl2 groups respectively. The decrease in body weight gain in these groups was initially not significant but by day 16 till the end of the experiment this decrease in weight had become statistically significant relative to the GA alone and AA alone groups (Fig. 2).

The body weight gain of rats expressed as percent of initial body weight exposed to Cadmium (CdCl2) only, CdCl2 and gallic acid (GA), CdCl2 and ascorbic acid (AA), GA alone and AA alone compared with the control (n = 12/group).
Gallic acid improved the learning pattern of rats following impairment by CdCl2
During the acquisition trial of the Morris water maze we assessed the learning of the rats as the time it took the rat to find and climb onto the escape platform (escape latency). In all the experimental groups, the escape latency diminished over the 5-day training period (Fig. 3A). Repeated-measures one-way ANOVA showed a significant reduction in escape latency with training (F (6.031, 63.63) = 2.514) p = 0.0316. On the 6th day when the escape platform had been removed we observed the CdCl2 group had a significant decrease in time spent at the target quadrant when compared to the other groups. Remarkably, the time spent in the target quadrant by the CdCl2 + GA group was similar to control, which was significantly (p = 0.029) increased when compared to the other groups (Fig. 3B). On the contrary, the CdCl2 group spent a significantly (p = 0.0001) increased time in the opposite quadrant while the control and the CdCl2 + GA group had similar significantly (p = 0.041) reduced duration in the opposite quadrant (Fig. 3C).

(A) Learning curve during the acquisition phase of the Morris water maze test (B) The probe trial of the Morris water maze showing time spent in the target quadrant and (C) The time spent in the opposite quadrant. Values are presented as mean ± Standard Deviation n = 12/group; aindicates significant difference at p < 0.05 compared with control; bIndicates significant difference at p < 0.05 compared with CdCl2(one-way ANOVA; n = 12). GA gallic acid, AA ascorbic acid.
The depressive-like behavior exhibited in the CdCl2 group was ameliorated following concurrent administration of gallic acid
During FST, we observed that the onset of immobility was significantly higher in the CdCl2 + GA and GA alone groups relative to other groups. Similarly, the duration of immobility was significantly increased in the GA alone group, while the CdCl2 + GA and CdCl2 + AA groups had significantly lower duration of immobility relative to GA alone and control groups. Overall, our results showed that the concurrent administration of GA or AA with CdCl2increased struggling behavior in the experimental rats (Fig. 4).

The effects of Gallic acid (GA) and ascorbic acid (AA) on CdCl2-induced depression. (A) is the onset of immobility. (B) is the duration of immobility. Values are presented as mean ± SD n = 12/group. asignificant difference at p < 0.05 versus control, bsignificant difference at p < 0.05 versus CdCl2.
Since we observed that CdCl2 + GA and CdCl2 + AA groups had increased escape or struggling-directed behavior (as depicted by significant decreased duration of immobility) compared to the CdCl2 alone group we subsequently sought to explore their locomotor activity since most antidepressants are known to reduce locomotion. We consequently investigated their actions on locomotion using the open field maze (OFM). We used the OFM as a positive control to further analyze the effect seen in the FST. Our results indicated similarity in line crossing across all the groups except in the AA only group where number of lines crossed was significantly reduced. Interestingly, we also observed a significant reduction in freezing time in the CdCl2 + GA and CdCl2 + AA groups relative to the CdCl2 alone group. In addition, stretched attend posture, number of fecal boli, peripheral square and grooming duration were similar across all the experimental groups (Fig. 5).

The effects of gallic acid and ascorbic acid on CdCl2-induced anxiety and locomotion in the open field test showing (A) freezing time (B) line crossings (C) rearing frequency(D) grooming duration (E) duration of stretched attend posture (F) center square duration (G) peripheral square duration and (H) number of fecal boli. Values are presented as mean + SD. aindicates significant difference at p < 0.05 compared with control bindicates significant difference at p < 0.05 compared with CdCl2 only group. GA gallic acid, AA ascorbic acid.
In the elevated plus Maze test, One-way analysis of variance (ANOVA) revealed a significant decrease in the frequency of entry into the open arms in the CdCl2 + AA group when compared with the control (p = 0.0014) and CdCl2 alone (p = 0.0006) groups. In addition, we observed a significant decrease in the frequency of entry into the close arms in all groups administered CdCl2, whether alone or concurrently with GA or AA. However, the time spent in the open and closed arms of the maze were similar across all the groups (Fig. 6).

The effects of gallic and ascorbic acids on CdCl2-induced anxiety in the elevated plus maze test. (A) Number of entries into the open arms (B) time spent in the open arms (C) number of entries into the closed arms and (D) time spent in the closed arms. n = 12/group, Values are presented as mean + SEM. aindicates significant difference at p < 0.05 compared with control bindicates significant difference at p < 0.05 compared with CdCl2 only group. GA gallic acid, AA ascorbic acid.
CdCl2 did not alter hanging latency on the forelimb suspension test
The mean hanging latency was 71.45 ± 11.52, 72.45 ± 13.19, 80.35 ± 11.92, 84.05 ± 11.18, 107.4 ± 5.026 and 105.0 ± 6.407 in the control, CdCl2 alone, CdCl2 + GA, CdCl2 + AA, GA alone and AA alone groups respectively. Our results indicate similarity in mean hanging latency across all groups (Fig. 7).

The hanging latency in experimental rats following administration of gallic acid, ascorbic acid and CdCl2 Values are presented as mean + SEM (n = 12/group).
Gallic acid rescued myelin disruption and reduced astrocytic activation following CdCl2-induced neurotoxicity
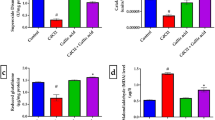
We further investigated the effects of CdCl2 on integrity of myelin sheaths using anti-MBP antibodies and Luxol Fast Blue (LFB) staining. Anti-MBP immunofluorescence staining showed a reduction in staining intensity in the cortex of CdCl2 and CdCl2 + AA rats (Fig. 8A). We observed a significant decrease in expression of neuronal nuclei (NeuN) protein across all groups relative to control (Fig. 8B).Quantitative examination of staining intensity indicated a significant reduction (p < 0.05) in the CdCl2 and CdCl2 + AA rats compared with control and CdCl2 + GA rats (Fig. 8C). Furthermore, we observed that LFB revealed the white matter tracts appeared thinner with vacuolations and stained pale in rats administered with CdCl2 (arrows, Fig. 9). Conversely, deeply stained myelinated fibers were detected traversing through the white matter in control and CdCl2 + GA groups (Fig. 9). Similarly, the CdCl2 group had less intensely stained, disorganized cerebellar white matter with significant decrease in number of positive MBP cells. Likewise, our results showed that cells positive for GFAP appear to be enlarged and activated in the CdCl2 rats (Fig. 10A). In addition, quantification of the expression of GFAP revealed a significant elevation in the CdCl2 group relative to the other test groups (Fig. 10B).

(A) Representative double immunofluorescence stainingphotomicrograph showing neuronal nulei (NeuN; red), myelin basic protein (MBP; green) and cell nuclei (DAPI; blue) in the brain cortex in experimental rats. Quantification ofstaining intensity of (B) NeuN and (C) MBP was performed using ImageJ software. GA gallic acid, AA ascorbic acid.

Representative images of Luxol fast blue staining of subcortical white matter of experimental rat showing the myelin density of the tract. Red arrowheads indicate vacuolation within the white matter. Note: the pale staining intensity in the CdCl2 and AA groups compared to other groups. GA gallic acid, AA ascorbic acid (scale bar = 50 µm).

(A) Immunofluorescence showing Glial fibrillary astrocytic protein (GFAP) positive cells following the administration of CdCl2, GA and AA. (B) Quantification of mean astrocyte count. GA gallic acid, AA ascorbic acid scale bar = 50 µm).
CdCl2-induced Purkinje cell degeneration was ameliorated by gallic and ascorbic acids
In Wistar rats, the cerebellum is made up of molecular, Purkinje and granular cell layers (Fig. 11). Our results showed a disruption with lack of clear organization of the Purkinje cell layer in the CdCl2 group. Additionally, the remaining Purkinje cells appeared disorganized with distorted cytoplasms. We further investigated this apparent neuronal loss, by carrying out Purkinje cell counts in the different treatment groups. Our results revealed a significant decrease in entire Purkinje cell number in the CdCl2 group (9% decrease in total number; p = 0.032). However, the total number of Purkinje neurons were similar in the control and groups administered gallic acid.

The cerebellum of experimental rats showing the molecular, Purkinje and granule cell layers with (A) H&E staining and (B) Luxol fast blue stain. Control, GA and AA groups had organized molecular cell layer with discrete stellate and basket cells. Large pyriform-shaped cells were also observed in the Purkinje cell layer with small deeply stained granular cell layer. Note the disorganization in the cells of the Purkinje layer in the CdCl2 group. Molecular cell layer (MCL), Purkinje cell (black arrowheads) and granular cell layers (GCL). GA gallic acid, AA ascorbic acid.
Discussion
Cadmium has been reported by several studies to induce behavioral deficits such as memory impairment, anxiety and depression27,28,29. It does this primarily through the generation of reactive oxygen species (ROS)9. Gallic acid is a phenolic acid with antioxidant activity which has been reported over the past years to be effective against nervous system disorders such as depression, anxiety, ischaemia, Alzheimer’s and Parkinson’s diseases14. Oral supplementation with ascorbic acid has also been used to mitigate heavy metal toxicity as well as protect against cadmium induced toxicity18. The present study was carried out to evaluate the effects of gallic acid on cadmium-induced neurotoxicity with focus on neuro-behavioral deficits such as memory loss, anxiety, depression and loss of motor coordination; and compare its effects with that of ascorbic acid, a common antioxidant.
The decrease in body weight in the CdCl2 group reported in this study is in consonance with the reports of El-Demerdash et al.30 and Prabu et al.31. This observed weight loss may be as a result of the association of CdCl2 administration with disruption in glucose regulation resulting in impaired protein, carbohydrate and lipid metabolism as previously highlighted32. Conversely, Young et al.33 reported an increase in body weight following CdCl2 administration. This variability in results may be due to the different exposure routes and dosage of CdCl2 in the studies. While in the previous study, CdCl2 was administered in water, our study utilized oral gavage to ensure that the full dose of CdCl2 was delivered directly to the stomach. However, the association of CdCl2 administration with weight alteration is outside the aim of this study.
The Morris water maze in rodents is a test for spatial learning and memory that relies on external or extra-maze cues to locate a submerged escape platform34. It tests for hippocampal dependent learning, which includes acquisition and long-term memory and plays an important role in the validation of cognitive studies in rodent models35. In the present study, learning occurred across all the groups but there was a significant decrease in time to locate the escape platform in the CdCl2 alone group when compared with the control which indicates that CdCl2 caused a learning deficit. In the probe phase, CdCl2 alone group showed a significant reduction in the time spent in the target quadrant when compared with the other experimental groups which is indicative of an impairment in spatial memory. It is anticipated that rats would use the visual stimuli from the cues within the testing room to form a “spatial orientation map” in their brain leading to a decrease in escape latency over the course of the test. However, this was not so in the CdCl2 only, this corroborates the findings of Halder et al.28 that cadmium causes a decrease in retention memory. Remarkably, CdCl2 rats treated with gallic and ascorbic acid spent similar time at the target quadrant as the control group in the probe trial of the MWM test, indicating a recovery in memory retention. Although CdCl2 caused a significant impairment in memory retention, spatial learning was still evident during the acquisition phase of the test as the animals spent reduced time in finding the escape platform with increasing number of trials. Interestingly, a study by Wang et al.36 aligns with this finding that cadmium-treated mice exhibit a deficit in this spatial memory and did not show improvements even after nine weeks of the cessation of cadmium.
Increased cadmium levels have also been linked to the presence of depressive-like behavior29. The forced swim test is a widely used test to evaluate depressive disorders24. The increase in the duration of immobility that is seen in the cadmium alone group is indicative of anxiety as earlier documented25,37. Remarkably, gallic acid and ascorbic acid treated groups reduced significantly the duration of immobility indicating their anti-depressive like potential in cadmium-induced toxicity.
The fear-related behaviors ranging from locomotion, freezing duration, exploratory activity and emotional state examined in our study revealed a dramatic decline in anxiety-like behaviors following administration of gallic and ascorbic acids. This outcome further supports that ascorbic acid promotes anxiolytic effects in diverse animal models as previously documented in zebrafish38, mice39 and rats40. Although, the exact mechanisms of anxiolytic activity of these compounds remains to be decisively established, reports have suggested anti-oxidant related decrease of brain oxidative stress and NMDA antagonism41,42. On the other hand, some authors have associated this activity to their ability in reducing circulating levels of cortisol, a stress hormone43.
The hanging wire test is a test of muscle strength44. Cadmium has been previously linked with motor activity impairment45. Our study nevertheless showed no significant difference in hanging latency across all the groups. This incongruence in hanging latency across the groups in this study may be due to the shorter duration of administration of the metal in our study compared to earlier reports14,45 that had a 28-day of administration of cadmium.
Demyelination is due to several pathologies in the brain such as inflammation, viral infection, focal compression, metabolic derangements and ischaemia46. Accumulation of cadmium has been linked to loss of myelin and reduced expression of myelin basic protein (MBP) in the cerebral and cerebellar cortices of rats47. Our study showed a reduced myelin expression in the cadmium treated group which was restored in both the ascorbic and gallic acid treated groups. The loss of cerebral neurons observed in LFB and demyelination in both MBP and LFB may be responsible for the impaired memory that was observed in the Morris water maze test. This study suggests that the demyelination could be due to an inflammatory process.
Astrocytes are cells that produce intermediate filaments such as glial fibrillary acidic protein (GFAP). Astrocytic activation (or astrocytosis) results from almost all insults or injury to the brain. The increased GFAP expression could also indicate an ongoing inflammation in cadmium toxicity leading to astrogliosis48. This supports the claim of astrocytic involvement in heavy metal neurotoxicity49. This astrogliosis was however reversed in the groups treated with gallic acid and ascorbic acid. Plant metabolites such as soybean supplementation in diet has also been shown to reduce arterial and cardiac injuries induced by cadmium50. Hence we propose that gallic acid being a secondary polyphenol with antioxidant potential could be beneficial against damages caused by cadmium chloride-induced experimental neurotoxicity and behavioral alterations. These effects can both be linked to their antioxidant activities.
Conclusion
Cadmium chloride caused a deficit in learning and memory with accompanying evidences of anxiety and depressive-like behavior which are indicative of neurotoxicity. Concurrent administration of gallic acid and ascorbic acid ameliorate the neurotoxic effects induced by cadmium. Although gallic acid showed a better neuroprotective effects over ascorbic acid; these compounds should be explored in population that are environmentally or occupationally exposed to toxic levels of cadmium.
Data availability
The supporting data of this study are available from the corresponding author upon reasonable request.
Abbreviations
- EPM:
-
Elevated plus maze
- GFAP:
-
Glial fibrillary acidic protein
- MBP:
-
Myelin basic protein
- MWM:
-
Morris water maze
- NeuN:
-
Neuronal nuclei
References
Tchounwou, P. B., Yedjou, C. G., Patlolla, A. K. & Sutton, D. J. Heavy metal toxicity and the environment. Exp. Suppl. 101, 133–164. https://doi.org/10.1007/978-3-7643-8340-4_6 (2012).
Jaishankar, M., Tseten, T., Anbalagan, N., Mathew, B. B. & Beeregowda, K. N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol 7(2), 60–72. https://doi.org/10.2478/intox-2014-0009 (2014).
Bernhoft, R. A. Cadmium toxicity and treatment. Sci. World J. 2013, 394652. https://doi.org/10.1155/2013/394652 (2013).
Genchi, G., Sinicropi, M. S., Lauria, G., Carocci, A. & Catalano, A. The effects of cadmium toxicity. Int. J. Environ. Res. Public Health. 17(11), 1–24. https://doi.org/10.3390/ijerph17113782 (2020).
Agency for Toxic Substance and Disease Registry (ATSDR). Toxicological Profile for Cadmium. Department of Health and Humans Services, Public Health Service, Centers for Disease Control, Atlanta, GA, USA. https://www.atsdr.cdc.gov/ToxProfiles/tp5.pdf (2017) accessed 28 January 2022.
Cui, Z. G., Ahmed, K., Zaidi, S. F. & Muhammad, J. S. Ins and outs of cadmium-induced carcinogenesis: Mechanism and prevention. Cancer Treat Res. Commun. 27, 100372. https://doi.org/10.1016/j.ctarc.2021.100372 (2021).
Rahimzadeh, M. R., Rahimzadeh, M. R., Kazemi, S. & Moghadamnia, A. A. Cadmium toxicity and treatment: An update. Casp. J. Intern. Med. 8(3), 135–145. https://doi.org/10.22088/cjim.8.3.135 (2017).
Aminzadeh, A. & Salarinejad, A. Effects of myricetin against cadmium-induced neurotoxicity in PC12 cells. Toxicol. Res. 10, 84–90. https://doi.org/10.1093/toxres/tfaa104 (2021).
López, E., Arce, C., Oset-Gasque, M. J., Cañadas, S. & González, M. P. Cadmium induces reactive oxygen species generation and lipid peroxidation in cortical neurons in culture. Free Radic. Biol. Med. 40(6), 940–951. https://doi.org/10.1016/j.freeradbiomed.2005.10.062 (2006).
Wallin, M., Sallsten, G., Lundh, T. & Barregard, L. Low-level cadmium exposure and effects on kidney function. Occup. Environ. Med. 71(12), 848–854. https://doi.org/10.1136/oemed-2014-102279 (2014).
Wang, B. & Du, Y. Cadmium and its neurotoxic effects. Oxid. Med. Cell Longev. 2013, 898034. https://doi.org/10.1155/2013/898034 (2013).
Ali, T. et al. Cadmium, an environmental contaminant, exacerbates alzheimer’s pathology in the aged mice’s brain. Front. Aging Neurosci. 13, 1–15. https://doi.org/10.3389/fnagi.2021.650930 (2021).
Ola-Davies, O. E. & Olukole, S. G. Gallic acid protects against bisphenol A-induced alterations in the cardio-renal system of Wistar rats through the antioxidant defense mechanism. Biomed. Pharmacother. 107, 1786–1794. https://doi.org/10.1016/j.biopha.2018.08.108 (2018).
Shabani, S., Rabiei, Z. & Amini-Khoei, H. Exploring the multifaceted neuroprotective actions of gallic acid: A review. Int. J. Food Prop. 23(1), 736–752. https://doi.org/10.1080/10942912.2020.1753769 (2020).
Kahkeshani, N. et al. Pharmacological effects of gallic acid in health and disease: A mechanistic review. Iran J. Basic Med. Sci. 22(3), 225–237. https://doi.org/10.22038/ijbms.2019.32806.7897 (2019).
Maya, S., Prakash, T. & Madhu, K. Assessment of neuroprotective effects of Gallic acid against glutamate-induced neurotoxicity in primary rat cortex neuronal culture. Neurochem. Int. 121, 50–58. https://doi.org/10.1016/j.neuint.2018.10.011 (2018).
Mansouri, M. T. et al. Neuroprotective effects of oral gallic acid against oxidative stress induced by 6-hydroxydopamine in rats. Food Chem. 138(2–3), 1028–1033. https://doi.org/10.1016/j.foodchem.2012.11.022 (2013).
Chambial, S., Dwivedi, S., Shukla, K. K., John, P. J. & Sharma, P. Vitamin C in disease prevention and cure: An overview. Indian J. Clin. Biochem. 28(4), 314–328. https://doi.org/10.1007/s12291-013-0375-3 (2013).
Moretti, M. Fraga DB, Rodrigues ALS Preventive and therapeutic potential of ascorbic acid in neurodegenerative diseases. CNS Neurosci. Ther. 23(12), 921–929. https://doi.org/10.1111/cns.12767 (2017).
Chen, N., Su, P., Wang, M. & Li, Y. M. Ascorbic acid inhibits cadmium-induced disruption of the blood-testis barrier by regulating oxidative stress-mediated p38 MAPK pathways. Environ. Sci. Pollut. Res. 25(22), 21713–21720. https://doi.org/10.1007/s11356-018-2138-4 (2018).
Erdogan, Z., Erdogan, S., Celik, S. & Unlu, A. Effects of ascorbic acid on cadmium-induced oxidative stress and performance of broilers. Biol. Trace Elem. Res. 104(1), 19–31. https://doi.org/10.1385/bter:104:1:019 (2005).
Ojo, O. A. et al. Gallic acid protects against cadmium chloride-induced alterations in Wistar rats via the antioxidant defense mechanism. J. Pharm. Pharmacogn. Res. 9(5), 668–676 (2021).
Vorhees, C. V. & Williams, M. T. Morris water maze: procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 1(2), 848–858. https://doi.org/10.1038/nprot.2006.116 (2010).
Yankelevitch-Yahav, R., Franko, M., Huly, A. & Doron, R. The forced swim test as a model of depressive-like behavior. J. Vis. Exp. 97, 52587. https://doi.org/10.3791/52587 (2015).
Can, A. et al. The mouse forced swim test. J. Vis. Exps. 58, 4–8. https://doi.org/10.3791/3638 (2011).
Carson, F. & Cappellano, C. Histotechnology: A Self Instructional Text. 4th ed. [Chicago], 183–184 (ASCP Press, 2015).
Haider, S. et al. Short term cadmium administration dose dependently elicits immediate biochemical, neurochemical and neurobehavioral dysfunction in male rats. Metab. Brain Dis. 30(1), 83–92. https://doi.org/10.1007/s11011-014-9578-4 (2015).
Halder, S. et al. Cadmium level in brain correlates with memory impairment in F1 and F2 generation mice: improvement with quercetin. Environ. Sci. Pollut. Res. 26(10), 9632–9639. https://doi.org/10.1007/s11356-019-04283-2 (2019).
Buser, M. C. & Scinicariello, F. Cadmium, lead, and depressive symptoms: analysis of national health and nutrition examination survey 2011–2012. J. Clin. Psychiatry. 78(5), e515–e521. https://doi.org/10.4088/JCP.15m10383 (2017).
El-Demerdash, F. M., Yousef, M. I., Kedwany, F. S. & Baghdadi, H. H. Cadmium-induced changes in lipid peroxidation, blood hematology, biochemical parameters and semen quality of male rats: protective role of vitamin E and beta-carotene. Food Chem. Toxicol. 42(10), 1563–1571. https://doi.org/10.1016/J.FCT.2004.05.001 (2004).
Prabu, S. M., Muthumani, M. & Shagirtha, K. Protective effect of Piper betle leaf extract against cadmium-induced oxidative stress and hepatic dysfunction in rats. Saudi J. Biol. Sci. 19(2), 229–239. https://doi.org/10.1016/j.sjbs.2012.01.005 (2012).
Kaltreider, R. C., Davis, A. M., Lariviere, J. P. & Hamilton, J. W. Arsenic alters the function of the glucocorticoid receptor as a transcription factor. Environ. Health Perspect. 109(3), 245–251. https://doi.org/10.1289/EHP.01109245 (2001).
Young, J. L. et al. Cadmium and high-fat diet disrupt renal, cardiac and hepatic essential metals. Sci. Rep. 9, 14675. https://doi.org/10.1038/s41598-019-50771-3 (2019).
Nunez, J. Morris water maze experiment. J. Vis. Exp. 19, e897. https://doi.org/10.3791/897 (2008).
Bromley-Brits, K., Deng, Y. & Song, W. Morris Water Maze test for learning and memory deficits in Alzheimer’s disease model mice. J. Vis. Exp. 53, 2–6. https://doi.org/10.3791/2920 (2011).
Wang, H., Zhang, L., Abel, G. M., Storm, D. R. & Xia, Z. Cadmium exposure impairs cognition and olfactory memory in male C57BL/6 Mice. Toxicol. Sci. 161(1), 87–102. https://doi.org/10.1093/toxsci/kfx202 (2018).
Lamtai, M. et al. Effect of chronic administration of cadmium on anxiety-like, depression-like and memory deficits in male and female rats: possible involvement of oxidative stress mechanism. J. Behav. Brain Sci. 8(5), 240–268. https://doi.org/10.4236/jbbs.2018.85016 (2018).
Puty, B. et al. Ascorbic acid protects against anxiogenic-like effect induced by methylmercury in zebrafish: Action on the serotonergic system. Zebrafish 11(4), 365–370. https://doi.org/10.1089/ZEB.2013.0947 (2014).
Angrini, M. A. & Leslie, J. C. Vitamin C attenuates the physiological and behavioural changes induced by long-term exposure to noise. Behav. Pharmacol. 23, 119–125. https://doi.org/10.1097/FBP.0b013e32834f9f68 (2012).
Hughes, R. N., Lowther, C. L. & Van Nobelen, M. Prolonged treatment with vitamins C and E separately and together decreases anxiety-related open-field behavior and acoustic startle in hooded rats. Pharmacol. Biochem. Behav. 97(3), 494–499. https://doi.org/10.1016/J.PBB.2010.10.010 (2011).
Lee, L. et al. Effect of supplementation of vitamin E and vitamin C on brain acetylcholinesterase activity and neurotransmitter levels in rats treated with scopolamine, an inducer of dementia. J. Nutr. Sci. Vitaminol. 47, 323–337. https://doi.org/10.3177/jnsv.47.323 (2001).
Bouayed, J., Rammal, H. & Soulimani, R. Oxidative stress and anxiety: relationship and cellular pathways. Oxid. Med. Cell Longev. 2(2), 63–67. https://doi.org/10.4161/oxim.2.2.7944 (2009).
Barbadoro, P. et al. Fish oil supplementation reduces cortisol basal levels and perceived stress: A randomized, placebo-controlled trial in abstinent alcoholics. Mol. Nutr. Food Res. 57, 1110–1114. https://doi.org/10.1002/MNFR.201200676 (2013).
Deacon, R. M. J. Measuring the strength of mice. J. Vis. Exp. 76, 1–4. https://doi.org/10.3791/2610 (2013).
Maodaa, S. N. et al. Effect of parsley (Petroselinum crispum, Apiaceae) juice against cadmium neurotoxicity in albino mice (Mus Musculus). Behav. Brain Funct. 12(1), 1–16. https://doi.org/10.1186/s12993-016-0090-3 (2016).
Love, S. Demyelinating diseases. J. Clin. Pathol. 59(11), 1151–1159. https://doi.org/10.1136/jcp.2005.031195 (2006).
Rai, N. K. et al. Exposure to As, Cd and Pb-mixture impairs myelin and axon development in rat brain, optic nerve and retina. Toxicol. Appl. Pharmacol. 273(2), 242–258. https://doi.org/10.1016/j.taap.2013.05.003 (2013).
Refolo, V. & Stefanova, N. Neuroinflammation and glial phenotypic changes in alpha-synucleinopathies. Front. Cell Neurosci. 13, 1–17. https://doi.org/10.3389/fncel.2019.00263 (2019).
Li, B., Xia, M., Zorec, R., Parpura, V. & Verkhratsky, A. Astrocytes in heavy metal neurotoxicity and neurodegeneration. Brain Res. 1752(77), 147234. https://doi.org/10.1016/j.brainres.2020.147234 (2021).
Brochin, R. L. et al. The cellular effect of lead poisoning and its clinical picture. GUJHS. 5(2), 1–8 (2008).
Funding
This study was partly supported by the International Research Organisation (IBRO) Return Home Fellowship awarded Grant 2019 (IBRORHF-2019) to OA.
Author information
Authors and Affiliations
Contributions
O.A., K.A., and F.O. conceptualized and designed the study, O.A., K.A., P.D., and U.A. acquired, analyzed and interpreted the data, O.A. and K. A wrote the first draft of the manuscript, O.A. secured funding for the study. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Adebiyi, O., Adigun, K., David-Odewumi, P. et al. Gallic and ascorbic acids supplementation alleviate cognitive deficits and neuropathological damage exerted by cadmium chloride in Wistar rats. Sci Rep 12, 14426 (2022). https://doi.org/10.1038/s41598-022-18432-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-18432-0
- Springer Nature Limited
This article is cited by
-
Subchronic Exposure to Mixture of Cadmium, Copper, and Nickel Induces Neurobehavioral Deficits and Hippocampal Oxidative Stress of Wistar Rats
Biological Trace Element Research (2024)
-
Insight into the emerging and common experimental in-vivo models of Alzheimer’s disease
Laboratory Animal Research (2023)