
Abstract
Although the availability of the measles vaccine, it is still epidemic in many countries globally, including Bangladesh. Eradication of measles needs to keep the basic reproduction number less than one \((\mathrm{i}.\mathrm{e}. \, \, {\mathrm{R}}_{0}<1)\). This paper investigates a modified (SVEIR) measles compartmental model with double dose vaccination in Bangladesh to simulate the measles prevalence. We perform a dynamical analysis of the resulting system and find that the model contains two equilibrium points: a disease-free equilibrium and an endemic equilibrium. The disease will be died out if the basic reproduction number is less than one \((\mathrm{i}.\mathrm{e}. \, \, {\mathrm{ R}}_{0}<1)\), and if greater than one \((\mathrm{i}.\mathrm{e}. \, \, {\mathrm{R}}_{0}>1)\) epidemic occurs. While using the Routh-Hurwitz criteria, the equilibria are found to be locally asymptotically stable under the former condition on \({\mathrm{R}}_{0}\). The partial rank correlation coefficients (PRCCs), a global sensitivity analysis method is used to compute \({\mathrm{R}}_{0}\) and measles prevalence \(\left({\mathrm{I}}^{*}\right)\) with respect to the estimated and fitted model parameters. We found that the transmission rate \((\upbeta )\) had the most significant influence on measles prevalence. Numerical simulations were carried out to commissions our analytical outcomes. These findings show that how progression rate, transmission rate and double dose vaccination rate affect the dynamics of measles prevalence. The information that we generate from this study may help government and public health professionals in making strategies to deal with the omissions of a measles outbreak and thus control and prevent an epidemic in Bangladesh.
Similar content being viewed by others


Introduction
Many people are being infected every year by serious respiratory infectious diseases, including measles. A significant number of them die or suffer severe illness and life-long complications1,2,3,4,5. The annual reports on the estimated number of measles cases and measles caused deaths worldwide are announced by the World Health Organization (WHO) and UNICEF based on the reported data of the member countries. According to WHO and UNICEF statistics for 2017, the total measles cases and measles-related deaths were recorded at 7,585,900 and 124,000, respectively. In 2018, there were about 9,769,400 recorded measles cases and 142,300 measles caused deaths6. The maximum cases were reported from Madagascar, Ukraine, Somalia and Liberia. Also, some developed countries, including the United Kingdom, Greece, Czechia, and Albania, lost the elimination status of measles in the latest year. Moreover, the United States counted the maximum number of cases, which was highest in 25 years, in 2018. In 2019, the maximum number of 207,500 people died due to measles, and the reported measles cases were 869,770 globally7. This year, Madagascar, Ukraine and Congo have reported the highest numbers of cases. Outbreaks are continuous in Angola, Cameroon, Kazakhstan, Chad, Nigeria, Thailand, Philippines, South Sudan and Sudan8. These continuous annually increments are indicating a matter of concerning issue in the world.
Measles is one of the most contagious respiratory infectious diseases caused by the measles virus that lives in an infected person’s nose and throat mucus. It is a virus of paramyxovirus family, genus morbilivirus, and this virus is found only in the human body among all animal species1,2. This virus can be spread directly from person to person through coughing and sneezing of the infected person. The clinical symptoms of measles are high fever, runny nose, cough, conjunctivitis, rhinitis, small white spots and a rash in the body of the infected people. This disease is more dangerous, especially for children under five years of age and adults older than 20 years of age. The complications including pneumonia, mouth ulcer, sinus and ear infections, diarrhea, malnutrition, blindness and brain damage may occur due to measles3. There is no specific medicine for the treatment of infected people with measles. According to the complications of the patients, a specific treatment may be suggested. Patients may need complete bed rest, fluids, control of fever and pains, and antibiotics9. Now, the measles vaccine is available, which is effective and inexpensive, and it has been possible to remarkably reduce the number of people dying from measles through vaccination5. The measles, mumps, and rubella (MMR) vaccines efficacy is 95% for preventing measles if the first dose is given to children at 12 months of age, and the efficacy increase to 99% after the second dose is given to children at greater than 12 months of age10.
Many countries and various public health organizations such as WHO, UNICEF, American Red Cross, Centers for Disease Control and Prevention (CDC), and United Nations Foundation have conducted tremendous efforts worldwide to fight against measles. These organizations launched the Measles and Rubella Initiative (MRI), a global partnership among these organizations to stop measles and rubella, in 2001 to reduce measles deaths globally by 90% by 2010 compared to 2000 estimates11. Presently, this partnership has taken the Measles and Rubella Strategic Framework 2021–2030 (MRSF 2021–2030) for a world free from measles and rubella12. However, despite being vaccine-preventable, measles is still a public health problem in many developing countries globally, especially in parts of Asia and Africa, because of low awareness, civil strife, vaccine hesitancy, lower immunization system and poor health infrastructures13.
In Bangladesh, one of the South East Asia Region (SEAR) countries, measles outbreaks occurred several times in different areas during 2000–2016. There were about 70,273 reported measles cases and 33,213 confirmed measles cases in Bangladesh during this period14. Although the Expanded Program of Immunization (EPI) was started in Bangladesh in 1979 to control and prevent measles15, the government adopted more initiatives like strengthening the surveillance system and introduction of the second dose of measles-containing vaccine (MCV2) in 2014 to eliminate measles from the country by 201816. Consequently, measles cases were reduced up to 84% over the last decades. However, the number of estimated measles cases has been increasing since 2016 nationwide. Despite some existing challenges, Rohingya refugees is another challenge for the removal of measles from Bangladesh.
In recent decades, the research relating to measles in epidemiology has been one of the most important research fields to researchers. Many researchers have already proposed their ideas and accomplished their research mathematically, theoretically or experimentally, using different deterministic or compartmental models, to find the comparatively best ways for measles control and prevention, focusing on different areas of the world, for example, London17, Afghanistan18, Kenya19, Madagascar20, Ontario21, Cape Coast22, Italy23, Senegal23, Taiwan24 and China25,26. Moreover, Momoh et al.27 studied an SEIR deterministic epidemic model to investigate the impact of asymptomatic individuals at the latent period on measles dynamics. Adewale et al.28 developed a mathematical model to ascertain the effect of distance between infected and non-infected persons in controlling the measles virus transmission. They observed that the number of infected individuals decreases due to increases in distance between infected and susceptible persons. Also, two studies highlighted the efficiency of vaccination in controlling and prevention of measles transmission29,30. Garba et al.31 also studied a compartmental mathematical model to examine the effect of vaccination and treatment on measles dynamics. Beay4 proposed a SIQR epidemic model and accomplished the numerical analysis of the model to explore the effect of treatment and quarantine on measles dynamics. The study demonstrated that the combined application of quarantine and treatment is more effective to control and prevent measles. It also observed that the measles spread reduces due to the treatment and quarantine of infected individuals.
In this study, we develop a novel compartmental measles model to simulate the prevalence of measles estimation in Bangladesh. We use the next-generation matrix method to determine the basic reproduction number of the system and found that this is an essential determinant for disease dynamics. To supplement and validate the analytic process, we use numerical techniques to solve the model equations and explore the epidemic trajectory for a range of possible parameters values and initial conditions. The local stability analyses of the disease-free and endemic equilibria are examined using the Routh-Hurwitz criteria. Following this, we perform a sensitivity analysis to investigate the model parameters that greatly influence measles prevalence. Finally, we investigate the impact of progression rate, transmission rate and double dose vaccinations on the dynamics of the measles outbreak.
Methods and materials
Model description
We developed a compartmental transmission dynamics measles model between the following mutually exclusive compartments: susceptible individuals, \(\mathrm{S}(\mathrm{t})\); those who have not yet infected with the disease but might become infected; first dose vaccinated individuals, \({\mathrm{V}}_{1}(\mathrm{t})\); those who have received the first dose of vaccine; second dose vaccinated individuals, \({\mathrm{V}}_{2}(\mathrm{t})\); those who have received the second dose of vaccine; Exposed individuals, \(\mathrm{E}(\mathrm{t})\); representing those that are infected and have not yet developed active measles disease; Infected individuals, \(\mathrm{I}(\mathrm{t})\); those who are infected and infectious; and Recovered individuals, \(\mathrm{R}(\mathrm{t})\); those who were previously infected and successfully recovered. Individuals in the recovery class are neither infectious nor susceptible, including people in treatment, isolation, no longer contacting others or dead.
The total population size \(\mathrm{N}(\mathrm{t})\) is assumed to be constant and well mixed:
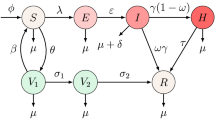
To ensure the population size constant, we replace all deaths as newborns in the susceptible compartment. It includes death through natural causes, which occurs in all states at the constant per-capita rate \(\upmu\), and measles-related deaths, which occur at the constant per capita rate \(\updelta\). Susceptible population \((\mathrm{S})\) who receive the first dose of vaccine move to the vaccinated compartment at a rate \(\upeta\). The first dose of vaccinated population \({\mathrm{V}}_{1}\) moves to the susceptible compartment at a rate \(\uprho\), and the rest of the population moves to the second dose of vaccinated population \({\mathrm{V}}_{2}\) at a per-capita rate \(\upsigma\). The second dose of the vaccinated population also moves to the recovery compartment at a rate \(\upomega\). Individuals in the \(\mathrm{S}\) compartment may be infected with the measles virus at a rate \(\uplambda =\mathrm{\upbeta SI}\), where \(\upbeta\) is the transmission rate between infected and susceptible population. Then infected individuals move to the exposed compartment \(\mathrm{E}\). A proportion of the exposed population progress to the infected compartment at a per-capita rate \(\mathrm{\upalpha }\). A proportion of the infected individuals move to the recovery compartment due to the treatment and natural recovery rate \(\upgamma\). The model flow diagram is presented in Fig. 1.

Schematic diagram of measles model for Bangladesh measles setting.
From those as mentioned above, the transmission dynamics of measles is given by the following compartmental system of nonlinear ordinary differential equations that describe the model:
The initial conditions of the system (2)–(7) are of the form.
It can be easily shown that the solution of the system (2)–(7) subject to the initial conditions (8) exists and is nonnegative for all \(\mathrm{t}\ge 0\).
Given the non-negative initial conditions of the system (2)–(7), it is direct to show that each state variable remains non-negative. Summing Eqs. (2)–(7), we find that the total population, \(\mathrm{N}(\mathrm{t})\) satisfies in the absence of death due to measles or if there are no infected individuals (i.e. I = 0)32, then we have
Integrating this equation, we find
Given the constant population size and positivity of solutions, it naturally follows that each states \(\mathrm{S}, {\mathrm{V}}_{1}, {\mathrm{V}}_{2},\mathrm{ E},\mathrm{ I},\mathrm{ R}\) are bounded.
Note that Eqs. (2)–(6) are independent of the recovery population; hence if we only wish to track measles incidence and prevalence, we can focus our attention on the following system:
Given the positivity and boundedness of the system solutions, we find that the feasible region for Eqs. (9)–(13) given by.
where D is the positively invariant region for the system (9)–(13). Therefore, in this study, we consider Eqs. (9)–(13) in the set \(\mathrm{D}\).
Ethical approval
This study is based on aggregated measles surveillance data in Bangladesh provided by the World Health Organization. No confidential information was included because mathematical analyses were performed at the aggregate level.
Results
Existence of equilibria
Two types of equilibrium solutions appear in this system: the disease-free equilibrium, which is reached when the basic reproduction number is less than one, i.e. \({\mathrm{R}}_{0}<1\) and the endemic equilibrium, which is reached when the basic reproduction number is greater than one, i.e. \({\mathrm{R}}_{0}>1\). We discuss these in order below.
Disease-free equilibrium point \(({\mathbf{X}}^{0})\)
In this section, we obtain the disease-free equilibrium point of the system (9)–(13) at which the epidemic is eliminated by applying \(\mathrm{E}=\mathrm{I}=0\). Hence, the disease-free equilibrium point is given by
That describes the state in which there is no infection in the community, and the total population \(\mathrm{N}\) is constant at time \(\mathrm{t}=0\).
Basic reproduction number \(({\mathbf{R}}_{0})\)
The basic reproduction number can be determined using the method of next-generation matrix33. The next-generation matrix is the production of matrices \(\mathrm{T}\) and \(- {\Sigma }^{-1}\) where the matrix \(\mathrm{T}\) represents the rate of infection transmission in \(\mathrm{E}\) and \(\mathrm{I}\) compartments and the matrix \(\Sigma\) describes all other transfer across the compartments. The matrices \(\mathrm{T}\) and \(\Sigma\) are given as
The next-generation matrix is
The basic reproduction number is the Eigen-value of the largest magnitude of the next-generation matrix \((\mathrm{K})\). Hence the basic reproduction number is obtained as
Endemic equilibrium point \(({\mathbf{X}}^{*})\)
The endemic equilibrium point of the system (9)–(13) is discovered by applying \(\mathrm{S}\ne {\mathrm{V}}_{1}\ne {\mathrm{V}}_{2}\ne \mathrm{E}\ne \mathrm{I}\ne 0\). Hence, the endemic equilibrium point is given by.
\({\mathrm{X}}^{*}=\left({\mathrm{S}}^{*}, {\mathrm{V}}_{1}^{*}, {\mathrm{V}}_{2}^{*}, {\mathrm{E}}^{*}, {\mathrm{I}}^{*}\right)\) where
Equation (15) shows that if \({\mathrm{R}}_{0}>1\) then the endemic equilibrium \({\mathrm{X}}^{*}({\mathrm{S}}^{*}, {\mathrm{V}}_{1}^{*}, {\mathrm{V}}_{2}^{*}, {\mathrm{E}}^{*}, {\mathrm{I}}^{*})\in \mathrm{D}\).
Stability analysis
To examining the stability of the equilibria of system (9)–(13), the following outcomes are proven:
Lemma 1
The disease-free equilibrium of the model is locally asymptotically stable if \({\mathrm{R}}_{0}<1\) and unstable if \({\mathrm{R}}_{0}>1\).
Proof
We consider the Jacobian of the system (9)–(13) which is given by
which, at the infection-free equilibrium point, \({\mathrm{X}}^{0}\), reduces to
Now we have to provide that all the eigenvalues of \(\mathrm{J}\left({\mathrm{X}}^{0}\right)\) are negative. As the 3rd column indicates only the diagonal terms which form the one negative eigenvalue, \(-(\upomega +\upmu )\) the other eigenvalues can be derived from the sub-matrix, \({\mathrm{J}}_{1}({\mathrm{X}}^{0})\) formed by excluding the 3rd row and column of \(\mathrm{J}\left({\mathrm{X}}^{0}\right)\). Which gives
This matrix can be written in block form as
where, \({\mathrm{A}}_{1}=\left(\begin{array}{cc}-(\upeta +\upmu )&\uprho \\\upeta & -(\uprho +\upsigma +\upmu )\end{array}\right)\), \({\mathrm{A}}_{2}=\left(\begin{array}{cc}0& -\upbeta {\mathrm{S}}^{0}\\ 0& 0\end{array}\right)\), \({\mathrm{A}}_{3}=\left(\begin{array}{cc}0& 0\\ 0& 0\end{array}\right)\) and \({\mathrm{A}}_{4}=\left(\begin{array}{cc}-(\mathrm{\upalpha }+\upmu )&\upbeta {\mathrm{S}}^{0}\\ \mathrm{\upalpha }& -(\upgamma +\updelta +\upmu )\end{array}\right).\)
The characteristic equation of the two-by-two block matrix \({\mathrm{J}}_{1}({\mathrm{X}}^{0})\) is
Since \({\mathrm{A}}_{3}=\left(\begin{array}{cc}0& 0\\ 0& 0\end{array}\right)\) this reduces to
Now we can apply the Routh-Hurwitz criteria for stability to matrices \({\mathrm{A}}_{1}\) and \({\mathrm{A}}_{4}\) directly and independently. We require that the trace is negative and the determinant is positive for each matrix.
Now for \({\mathrm{A}}_{1}\) matrix
and
Again for \({\mathrm{A}}_{4}\) matrix
and
which we can rewrite as
Hence, the disease-free equilibrium \({\mathrm{X}}^{0}\) is locally asymptotically stable for \({\mathrm{R}}_{0}<1\). If either \({\mathrm{R}}_{0}>1\), at least one of the roots of the characteristic equation has a positive real part and \({\mathrm{X}}^{0}\) is unstable.
Lemma 2
The endemic equilibrium \({\mathrm{X}}^{*}\) is locally asymptotically stable if \({\mathrm{R}}_{0}>1\).
Proof
We consider the Jacobian of the system (9)–(13) at \({\mathrm{X}}^{*}=({\mathrm{S}}^{*}, {\mathrm{V}}_{1}^{*}, {\mathrm{V}}_{2}^{*}, {\mathrm{E}}^{*}, {\mathrm{I}}^{*})\) which is given by
The 3rd column indicates only the diagonal terms which form the one negative eigenvalues,\(-(\upomega +\upmu )\), the other eigenvalues can be derived from the sub-matrix, \({\mathrm{J}}_{1}({\mathrm{X}}^{*})\) formed by excluding the 3rd rows and columns of \(\mathrm{J}({\mathrm{X}}^{*})\). Which gives
The characteristics equation of \({\mathrm{J}}_{1}\left({\mathrm{X}}^{*}\right)\) is defined as,
where
On simplify
and
From (17) it is easy to verify that
\({\mathrm{a}}_{3}>0, \, {\mathrm{a}}_{2}>0, \, {\mathrm{a}}_{1}>0\) and \({\mathrm{a}}_{0}>0\) if \({\mathrm{I}}^{*}>0\). From (15) it is also clear that \({\mathrm{I}}^{*}\) is positive if \({\mathrm{R}}_{0}>1\).
Hence, by the Routh–Hurwitz stability criterion, the endemic equilibrium point \({\mathrm{X}}^{*}\) is locally asymptotically stable for \({\mathrm{R}}_{0}>1\).
In order to verify the nature of the disease-free and endemic equilibrium analysis, we used Monte Carlo simulation34 to prove the conditions by calculating the real part of the eigenvalues of the Jacobian matrix of disease-free and endemic equilibriums. The simulation outcomes are presented in Figs. 2 and 3.

Numerical simulation for the infection-free equilibrium stability conditions and the real part distribution of the eigenvalues \(({\uplambda }_{1}, \, {\uplambda }_{2}, \, {\uplambda }_{3}, \, {\uplambda }_{4}, \, {\uplambda }_{5})\). (A1) depict that \({\mathrm{R}}_{0}<1\) always hold, and (A2) represents the related distribution of the real part of the eigenvalues for the disease-free conditions.

Numerical simulation for the endemic equilibrium stability conditions and the real part distribution of the eigenvalues \(({\uplambda }_{1}, {\uplambda }_{2}, {\uplambda }_{3}, {\uplambda }_{4}, {\uplambda }_{5})\). (B1) depicts that \({\mathrm{R}}_{0}>1\) always hold, and (B2) represents the related distribution of the real part of the eigenvalues for the endemic conditions.
Figure 2 represented that the disease-free equilibrium is locally stable as all the five eigenvalues (real part) are negative \((\mathrm{i}.\mathrm{e}. \, {\uplambda }_{1}, \, {\uplambda }_{2}, \, {\uplambda }_{3}, \, {\uplambda }_{4}, \, \mathrm{ and } \, {\uplambda }_{5}<0)\). Whilst Fig. 3 represented that the endemic equilibrium is unstable as two eigenvalues (real part) is positive \((\mathrm{i}.\mathrm{e}. \, {\uplambda }_{4}, \, {\uplambda }_{5}>0)\).
Parameters estimation
We estimated the measles model parameters from fitting different combinations of parameters in Eqs. (2)–(7) to the actual number of measles cases in Bangladesh from 2000 to 201935. In order to parameterise measles model (2)–(7), we obtained some of the parameter values from the literature (see Table 1), rest of the parameters were estimated from data fitting (see Fig. 4). The estimation of parameters was carried out using the least-squared method, which minimises summation of the square errors given by \(\sum {\left(\mathrm{M}\left(\mathrm{t},\mathrm{p}\right)-{\mathrm{N}}_{\mathrm{actual}}\right)}^{2}\) subject to the measles model (2)–(7), where \({\mathrm{N}}_{\mathrm{actual}}\) is the actual reported measles data, and \(\mathrm{M}\left(\mathrm{t}, \, \mathrm{p}\right)\) denotes the solution of the model corresponding to the number of measles cases over time \(\mathrm{t}\) with the set of estimated parameters, denoted by \(\mathrm{p}\).

Cumulative number of confirmed measles cases from 2000 to 2019 (red dot) and the corresponding model best fit (solid blue curve) in Bangladesh.
Estimation of basic reproduction number \(({\mathbf{R}}_{0})\)
The basic reproduction number is well-defined as the estimated number of secondary cases produced by a single infectious case presented into an exclusively susceptible population. The disease can spread in a population only if the basic reproduction number is greater than one. Our main objective here is to calculate the basic reproduction number \(({\mathrm{R}}_{0})\) of the measles model (2)–(7). Using the values \(\mathrm{\upalpha }=0.018,\upbeta =7.45\times {10}^{-7},\mathrm{ N}=\mathrm{163,046,161},\upmu =\frac{1}{70},\uprho =0.6,\) \(\upsigma =0.93\),
\(\upgamma =0.6, \, \updelta =0.125, \, \upeta =0.94\), and substituting it into the basic reproduction number \(({\mathrm{R}}_{0})\) expression result in:
Hence, the basic reproduction number, \({\mathrm{R}}_{0}\) is approximately 1.44. It indicates that a single infected individual can spread the measles disease to 1 or 2 susceptible individuals.
Sensitivity analysis
We perform the sensitivity of the model basic reproduction number \(({\mathrm{R}}_{0})\) and measles prevalence \(({\mathrm{I}}^{*})\) to the model parameters using the Latin Hypercube Sampling (LHS) method with 10,000 runs per simulation. The LHS is a Monte Carlo stratified sampling technique that permits us to concurrently achieve an unbiased assessment of the model output for a particular set of input parameter values. We estimated the Partial Rank Correlation Coefficients (PRCCs): a global sensitivity analysis method using LHS of crucial output variables. We allowed a uniform distribution from 0 to 4 times the baseline value for each input parameter to explore the relationship between model output variable and parameters. The PRCCs for the basic reproduction number and measles prevalence in Figs. 5 and 6 have been produced using the expressions \({\mathrm{R}}_{0}\) and \({\mathrm{I}}^{*}\). Results show that parameters transmission rate \((\upbeta )\) and progression rates \((\mathrm{\upalpha and \uprho })\) have a positive correlation with the model outcomes \({\mathrm{R}}_{0}\) and \({\mathrm{I}}^{*}\), which means that decreasing these parameters values will reduce the prevalence of measles. On the other hand, parameters \(\upeta , \, \upsigma , \, \upgamma\) and \(\updelta\) have a negative correlation with the model outcomes \({\mathrm{R}}_{0}\) and \({\mathrm{I}}^{*}\), which indicates that increasing these parameters will decrease the prevalence of measles.

Correlation between basic reproduction number \(({\mathrm{R}}_{0})\) and the model parameters \(\upbeta , \, \mathrm{ \upalpha }, \, \uprho , \, \upsigma , \, \upgamma , \, \mathrm{ \upeta and \delta }\).

Correlation between measles prevalence \(({\mathrm{I}}^{*})\) and the model parameters \(\upbeta , \, \mathrm{ \upalpha }, \, \uprho , \, \upsigma , \, \upgamma , \, \mathrm{ \upeta and \delta }\).
From the explicit formula for basic reproduction number \({\mathrm{R}}_{0}\), the analytical expression for the sensitivity indices can be derived applying the method in36,37,38,39 to each of the parameters, e.g.
Using the parameter values in Table 1, we have the following values and the nature of their signs in Table 2.
In the sensitivity indices of \({\mathrm{R}}_{0}\) the most sensitive parameter is the transmission rate \((\upbeta )\) of measles. Another significant parameter is the first dose of vaccination rate \((\upeta )\). The least sensitive parameter is the measles related death rate \((\updelta )\). Hence, increasing (or decreasing) the transmission rate \((\upbeta )\) of measles by 100% increases (or decreases) the basic reproduction number \({\mathrm{R}}_{0}\) by 100%. Similarly, increasing (or decreasing) the measles-related death rate \((\updelta )\) by 100% decreases (or increases) \({\mathrm{R}}_{0}\) by 16.9%.
Numerical simulation
In this section, we carry out detailed numerical simulations to support the analytic results and to assess the impact of model parameters, including progression rate, transmission rate and double dose vaccination. For illustration, we have chosen baseline parameter values consistent with measles infection and transmission (see Table 1). We found two equilibrium points following the analytical results: the disease-free equilibrium \(({\mathrm{X}}^{0})\) and an endemic equilibrium \({(\mathrm{X}}^{*})\). We used different initial conditions for the exposed and infected population and found that if the basic reproduction number less than one \((\mathrm{i}.\mathrm{e}.\, {\mathrm{R}}_{0}<1)\), then the disease-free equilibrium is locally asymptotically stable. Furthermore, if \({\mathrm{R}}_{0}>1\) then measles persists in the population.
Figure 7 illustrates the stability of the disease-free equilibrium \((\mathrm{i}.\mathrm{e}.\,\mathrm{ when } {\mathrm{R}}_{0}<1)\) by depicting system trajectories through the \(\mathrm{E vs} \mathrm{I}\) plane originating from different initial conditions. In this case, measles disease dies out. Figure 8 shows the stability of the endemic equilibrium \((\mathrm{i}.\mathrm{e}.\mathrm{ when } {\mathrm{R}}_{0}>1)\), and in this case, measles disease persists in the population.

Infection-free equilibrium: \({\mathrm{R}}_{0}<1\). In this case measles disease dies out (black dot). All parameter values assume their baseline values given in Table 1.

Endemic equilibrium: \({\mathrm{R}}_{0}>1\). In this case measles disease persist in the population (black dot). All parameter values assume their baseline values given in Table 1.
Figures 9 and 10 show that the effect of progression rate and transmission rate on measles prevalence. From these figures, we observed that the burden of the measles prevalence increase if the progression and transmission rates increase, which means that those have a positive correlation with a measles outbreak. Figures 11 and 12 show that the increases in first and second dose vaccination rates reduce the measles prevalence and reduce the risk of an outbreak.

Impact of progression rate \((\mathrm{\upalpha })\) on measles prevalence.

Impact of transmission rate \((\upbeta )\) on measles prevalence.

Impact of first dose vaccine \((\upeta )\) on measles prevalence.

Impact of second dose vaccine \((\upsigma )\) on measles prevalence.
Summary and conclusion
This paper has developed and analyzed a compartmental transmission dynamics measles model with double dose vaccination in Bangladesh. We have determined an analytic expression for the basic reproduction number using the next-generation matrix and found that the disease-free equilibrium is locally asymptotically stable if the basic reproduction number is less than one. We have also found that measles disease persists in the community if the basic reproduction number is greater than one. Sensitivity analysis has also been performed to explore the impact of model parameters and findings showed that the spread of the measles disease largely depends on the transmission rate. Therefore, effort should be made to minimize unnecessary transmission with measles infected individuals. However, if we treat early measles infected individuals, it will also reduce transmission from infected person to uninfected person. This study has also highlighted the significance of vaccination in controlling and preventing the spread of measles in the community of Bangladesh. Vaccination in a population is the best way to control an outbreak of measles. Numerical analysis has revealed that vaccination has a negative impact on the prevalence of measles. This finding indicates that the improvement in vaccination dose rate decreases the spread of measles. Therefore, to attain a high level of herd immunity for the disease, mass vaccination exercise should be encouraged to cover most of the population to prevent an outbreak of measles in Bangladesh.
Data availability
The datasets produced during the study are available from the corresponding author on reasonable request.
References
Yanagi, Y., Takeda, M. & Ohno, S. Measles virus: Cellular receptors, tropism and pathogenesis. J. Gen. Virol. 87, 2767–2779 (2006).
Griffin, D. E. The immune response in measles: Virus control, clearance and protective immunity. Viruses 8, 282 (2016).
Perry, R. T. & Halsey, N. A. The clinical significance of measles: A review. J. Infect. Dis. 189, S4–S16 (2004).
Beay, L. K. AIP Conference Proceedings (AIP Publishing LLC, 2004).
Abad, C. & Safdar, N. The reemergence of measles. Curr. Infect. Dis. Rep. 17, 1–8 (2015).
WHO. More than 140,000 Die from Measles as Cases Surge Worldwide (WHO, 2019).
Healio. Measles Killed 207K People in 2019 as Cases Hit 23-Year High. (2020).
WHO. New Measles Data August 2019 (WHO, 2019).
CDC. Measles Cases and Outbreaks. (2021).
Bester, J. C. Measles and measles vaccination: A review. JAMA Pediatr. 170, 1209–1215 (2016).
WHO. Global Measles and Rubella Strategic Plan (WHO, 2012).
WHO. Measles and Rubella Strategic Framework 2021–2030 (WHO, 2020).
WHO. Status Report on Progress Towards Measles and Rubella Elimination SAGA Working Group on Measles and Rubella (WHO, 2012).
Khanal, S. et al. Progress toward measles elimination: Bangladesh, 2000–2016. MMWR Morb. Mortal. Wkly. Rep. 66, 753 (2017).
Wiesen, E. et al. Stability of the age distribution of measles cases over time during outbreaks in Bangladesh, 2004–2006. J. Infect. Dis. 204, S414–S420 (2011).
Sultana, S. Elimination of measles from Bangladesh: Progression and challenges ahead. J. Microbiol. Exp. 5, 00174 (2017).
Magpantay, F., King, A. & Rohani, P. Age-structure and transient dynamics in epidemiological systems. J. R. Soc. Interface 16, 20190151 (2019).
Cilli, A., Ergen, K. & Akat, E. Some mathematical models and applications used in epidemic. Sigma (2019).
Fred, M. O., Sigey, J. K., Okello, J. A., Okwoyo, J. M. & Kangethe, G. J. Mathematical modeling on the control of measles by vaccination: Case study of KISII County, Kenya. SIJ Trans. Comput. Sci. Eng. Appl. (CSEA) 2, 38–46 (2014).
Alhamami, H. A Susceptible-Exposed-Infected-Recovered-Vaccinated (SEIRV) Mathematical Model of Measles in Madagascar (Springer, 2019).
Hooker, G., Ellner, S. P., Roditi, L. D. V. & Earn, D. J. Parameterizing state–space models for infectious disease dynamics by generalized profiling: Measles in Ontario. J. R. Soc. Interface 8, 961–974 (2011).
Okyere-Siabouh, S. & Adetunde, I. Mathematical model for the study of measles in Cape Coast Metropolis. Int. J. Modern Biol. Med. 4, 110–113 (2013).
Manfredi, P. & Williams, J. R. Realistic population dynamics in epidemiological models: The impact of population decline on the dynamics of childhood infectious diseases: Measles in Italy as an example. Math. Biosci. 192, 153–175 (2004).
Chen, S., Chang, C., Jou, L. & Liao, C. Modelling vaccination programmes against measles in Taiwan. Epidemiol. Infect. 135, 775–786 (2007).
Huang, J., Ruan, S., Wu, X. & Zhou, X. Seasonal transmission dynamics of measles in China. Theory Biosci. 137, 185–195 (2018).
Yang, W., Li, J. & Shaman, J. Characteristics of measles epidemics in China (1951–2004) and implications for elimination: A case study of three key locations. PLoS Comput. Biol. 15, e1006806 (2019).
Momoh, A., Ibrahim, M., Uwanta, I. & Manga, S. Mathematical model for control of measles epidemiology. Int. J. Pure Appl. Math. 87, 707–717 (2013).
Adewale, S., Mohammed, I. & Olopade, I. Mathematical analysis of effect of area on the dynamical spread of measles. IOSR J. Eng. 4, 43–57 (2014).
Smith, R., Archibald, A., MacCarthy, E., Liu, L. & Luke, N. S. A mathematical investigation of vaccination strategies to prevent a measles epidemic. NCJ Math. Stat. 2, 29–44 (2016).
Peter, O., Afolabi, O., Victor, A., Akpan, C. & Oguntolu, F. Mathematical model for the control of measles. J. Appl. Sci. Environ. Manag. 22, 571–576 (2018).
Garba, S., Safi, M. & Usaini, S. Mathematical model for assessing the impact of vaccination and treatment on measles transmission dynamics. Math. Methods Appl. Sci. 40, 6371–6388 (2017).
Tilahun, G. T., Demie, S. & Eyob, A. Stochastic model of measles transmission dynamics with double dose vaccination. Infect. Dis. Model. 5, 478–494 (2020).
Diekmann, O., Heesterbeek, J. & Roberts, M. G. The construction of next-generation matrices for compartmental epidemic models. J. R. Soc. Interface 7, 873–885 (2010).
Campo-Duarte, D. E., Vasilieva, O., Cardona-Salgado, D. & Svinin, M. Optimal control approach for establishing wMelPop Wolbachia infection among wild Aedes aegypti populations. J. Math. Biol. 76, 1907–1950 (2018).
WHO. Measles: Number of Reported Cases in Bangladesh (WHO, 2020).
Asamoah, J. K. K. et al. Global stability and cost-effectiveness analysis of COVID-19 considering the impact of the environment: using data from Ghana. Chaos Solitons Fractals 140, 110103 (2020).
Asamoah, J. K. K., Oduro, F. T., Bonyah, E. & Seidu, B. Modelling of rabies transmission dynamics using optimal control analysis. J. Appl. Math. 2017, 1–23 (2017).
Kuddus, M. A., Meehan, M. T., White, L. J., McBryde, E. S. & Adekunle, A. I. Modeling drug-resistant tuberculosis amplification rates and intervention strategies in Bangladesh. PLoS ONE 15, e0236112 (2020).
Kuddus, M. A., Meehan, M. T., Sayem, M. A. & McBryde, E. S. Scenario analysis for programmatic tuberculosis control in Bangladesh: a mathematical modelling study. Sci. Rep. 11, 1–17 (2021).
Economy, C. Bangladesh Population in 2019. https://countryeconomy.com/demography/population/bangladesh (2020).
Yang, Y., Li, J., Ma, Z. & Liu, L. Global stability of two models with incomplete treatment for tuberculosis. Chaos Solitons Fractals 43, 79–85 (2010).
Acknowledgements
The authors would like to sincerely thank the reviewers and the editor for their thoughtful comments and efforts towards improving our manuscript and shape the final version.
Author information
Authors and Affiliations
Contributions
M.A.K., M.M. and A.R. conceived the project concept; M.A.K. cleaned the data and wrote the initial code; M.A.K. and M.M. performed the data analysis, model development and interpretation under the supervision of A.R.; A.R. refined the analyses results further; M.A.K. and M.M. wrote the initial draft of this manuscript; A.R. provided critical feedback to improve the manuscript; and all authors provided input into revisions and approved the final manuscript and submission for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kuddus, M.A., Mohiuddin, M. & Rahman, A. Mathematical analysis of a measles transmission dynamics model in Bangladesh with double dose vaccination. Sci Rep 11, 16571 (2021). https://doi.org/10.1038/s41598-021-95913-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-95913-8
- Springer Nature Limited
This article is cited by
-
Cost-effectiveness analysis of COVID-19 intervention policies using a mathematical model: an optimal control approach
Scientific Reports (2024)
-
Analyzing the stochastic dynamics of COVID-19 waves in Turkey using real data and piecewise sinusoidal functions
International Journal of Dynamics and Control (2024)
-
Analysis and dynamics of measles with control strategies: a mathematical modeling approach
International Journal of Dynamics and Control (2023)