
Abstract
Anti-retroviral therapy (ART) can inhibit HIV proliferation but not achieve virus eradication from HIV-infected individuals. Under ART-based HIV control, virus-specific CD8+ T-cell responses are often reduced. Here, we investigated the impact of therapeutic vaccination inducing virus-specific CD8+ T-cell responses under ART on viral control in a macaque AIDS model. Twelve rhesus macaques received ART from week 12 to 32 after simian immunodeficiency virus (SIV) infection. Six of them were vaccinated with Sendai virus vectors expressing SIV Gag and Vif at weeks 26 and 32, and Gag/Vif-specific CD8+ T-cell responses were enhanced and became predominant. All macaques controlled viremia during ART but showed viremia rebound after ART cessation. Analysis of in vitro CD8+ cell ability to suppress replication of autologous lymphocytes-derived SIVs found augmentation of anti-SIV efficacy of CD8+ cells after vaccination. In the vaccinated animals, the anti-SIV efficacy of CD8+ cells at week 34 was correlated positively with Gag-specific CD8+ T-cell frequencies and inversely with rebound viral loads at week 34. These results indicate that Gag-specific CD8+ T-cell induction by therapeutic vaccination can augment anti-virus efficacy of CD8+ cells, which may be insufficient for functional cure but contribute to more stable viral control under ART.
Similar content being viewed by others

Introduction
HIV induces persistent infection leading to AIDS onset in humans. Anti-retroviral therapy (ART) can control HIV replication and prevent AIDS progression. However, ART is unable to eradicate viruses and HIV-infected individuals need to receive life-long therapy for AIDS prevention1,2,3,4. It has been suggested that ART does not always exhibit complete shutdown of viral replication, while blips of undetectable levels of viral replication under ART may cause chronic persistent inflammation, possibly leading to progression of aging-related diseases5,6,7,8,9.
Virus-specific cytotoxic CD8+ T-cell responses play a central role in the control of HIV and simian immunodeficiency virus (SIV) replication10,11,12,13,14. Because ART-based HIV control is often accompanied by reduced virus-specific CD8+ T-cell responses15,16, augmentation of effective CD8+ T-cell responses by therapeutic vaccination may contribute to more complete shutdown of viral replication under ART, although therapeutic vaccine trials under ART have failed to achieve functional HIV cure, sustained viral control after ART cessation17,18,19,20. Optimization of vaccine immunogens is important for induction of effective CD8+ T-cell responses. Cumulative studies have indicated strong anti-HIV efficacy of Gag-specific CD8+ T cells14,21,22,23,24,25,26. Furthermore, Vif-specific CD8+ T cells have recently been indicated to exert strong suppressive pressure against HIV replication27,28. However, the impact of therapeutic vaccination inducing these CD8+ T cells under ART have not fully been determined.
We have previously developed Sendai virus (SeV) vectors expressing SIV/HIV antigens and have shown the potential of these vectors to induce viral antigen-specific T-cell responses in macaques and humans29,30,31. In the present study, SIVmac239-infected rhesus macaques were immunized with SeV vectors expressing SIVmac239 Gag and Vif antigens under ART. This immunization enhanced Gag/Vif-specific CD8+ T-cell responses. Our analysis indicated association of Gag-specific CD8+ T-cell responses with anti-SIV efficacy of CD8+ cells post-vaccination.
Results
Analysis of plasma viral loads
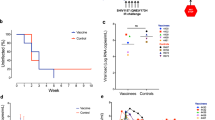
Twelve Burmese rhesus macaques consisting of six animals sharing the MHC-I haplotype 90–010-Ie (E), four sharing 89–075-Iw (W) and two sharing 91–010-Is (S) were used in the present study as shown in Table 1. These macaques were intravenously inoculated with SIVmac239 and received ART from week 12 to 32 post-infection. All the macaques showed persistent viremia after the SIV infection and reduced plasma viral loads after ART initiation at week 12 (Fig. 1). These macaques were divided into two groups, Group N (n = 6) receiving no vaccination and Group V (n = 6) receiving vaccination at weeks 26 and 32 post-infection. The Group V macaques consisting of three E-positive, two W-positive and one S-positive were intranasally immunized with Gag- and Vif-expressing SeV vectors at weeks 26 and 32 post-infection. Two of the Group N (NE3 and NW5) and all the six Group V animals were intravenously administered with polyclonal anti-SIV immunoglobulin G (anti-SIV IgG). After ART cessation at week 32, all macaques showed reappearance of plasma viremia. No clear difference was observed in viral loads post-ART between two anti-SIV IgG-treated and four untreated macaques in Group N. While two Group V macaques VE2 and VW4 exhibited relatively lower viral loads post-ART, no significant difference in viral loads post-ART was observed between Groups N and V.

Plasma viral loads after SIVmac239 infection. (a) Viral loads (SIV gag RNA copies/ml plasma) determined as described previously30. The lower limit of detection is approximately 4 × 102 copies/ml. Six Group N (left panel) and six Group V (right panel) macaques received ART from week 12 to 32 after SIVmac239 infection. Group V macaques received therapeutic SeV-Gag/Vif vaccination at weeks 26 and 32 post-infection. (b) Comparison of viral loads at weeks 12, 34 and 36 post-infection between Groups N and V. No significant difference was observed.
Analysis of antigen-specific CD8+ T-cell responses
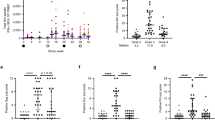
We examined CD8+ T-cell responses specific for SIV individual antigens in these macaques before ART initiation, during ART, and after ART cessation by detection of antigen-specific interferon-γ (IFN-γ) induction (Fig. 2). Before the ART initiation at week 12, macaques possessing the MHC-I haplotype E showed predominant induction of Nef-, Tat/Rev- and Env-specific CD8+ T-cell responses, whereas those possessing the haplotypes W/S predominantly induced Gag/Vif-specific CD8+ T-cell responses. At week 26 during ART, these antigen-specific CD8+ T-cell responses were reduced as expected. All the vaccinated Group V macaques showed induction and/or enhancement of Gag/Vif-specific CD8+ T-cell responses at week 27 post-infection, one week after the first SeV-Gag/Vif vaccination. After the second SeV-Gag/Vif vaccination and the ART cessation at week 32, SIV antigen-specific CD8+ T-cell responses were enhanced. Comparison revealed significantly higher Gag- and Vif-specific CD8+ T-cell responses in Group V than in Group N at weeks 27 and 34, whereas no significant difference was observed before vaccination (at week 26) (Fig. 3a,b). There was no significant difference in CD8+ T-cell responses targeting Nef that was not included in the vaccine antigens at week 26, 27 or 34 between Groups N and V (Fig. 3c).

SIV antigen-specific CD8+ T-cell responses after SIVmac239 infection. (a) Representative gating schema for detection of specific IFN-γ induction after peptide stimulation in flow cytometric analysis. Data on PBMCs of macaque VW4 at week 38 without stimulation (NC) and with stimulation using overlapping peptides spanning the N-terminal half of Gag proteins (Gag) are shown. (b) SIV antigen-specific CD8+ T-cell frequencies at indicated time points after SIVmac239 infection. CD8+ T-cell responses targeting SIV Gag, Vif, Nef, Pol, Vpx/Vpr, Tat/Rev, and Env were examined by detection of specific IFN-γ induction after stimulation using overlapping peptides spanning individual antigens. Log-transformed CD8+ T-cell frequencies are shown. PBMCs were obtained at weeks 2–12 (before ART initiation), 26 (just before the 1st vaccination), 27 (1 week after the 1st vaccination), 34 (2 weeks after ART cessation) and 38 post-infection and subjected to the analyses. ND not determined because of the limitation of available samples.

Comparison of Gag/Vif/Nef-specific CD8+ T-cell responses between Groups N and V. (a) Comparison of Gag-specific CD8+ T-cell frequencies between Groups N and V at weeks 26, 27, and 34 post-infection. Group V showed significantly higher frequencies than Group N at weeks 27 (P = 0.0087 by Mann–Whitney U test) and 34 (P = 0.0130). (b) Comparison of Vif-specific CD8+ T-cell frequencies between Groups N and V at weeks 26, 27, and 34 post-infection. Group V showed significantly higher frequencies than Group N at weeks 27 (P = 0.0022) and 34 (P = 0.0238). (c) Comparison of Nef-specific CD8+ T-cell frequencies between Groups N and V at weeks 26, 27, and 34 post-infection. No significant difference was observed.
The unvaccinated Group N animals showed similar patterns of immunodominance pre-ART and post-ART; i.e., Nef- and Env-specific CD8+ T-cell responses were predominant post-ART in the E-positive unvaccinated macaques (NE1, NE2 and NE3), while Gag/Vif-specific CD8+ T-cell responses were predominant in the W/S-positive (NW4, NW5 and NS6). In contrast, different patterns of immunodominance were shown pre-ART and post-ART in E-positive vaccinated animals; i.e., Gag/Vif-specific CD8+ T-cell responses were predominant post-ART (at week 34) in the E-positive vaccinated macaques (VE1, VE2 and VE3) as well as in the W/S-positive (VW4, VW5 and VS6) in Group V. These results indicate that the SeV-Gag/Vif vaccination under ART can enhance not only dominant Gag/Vif-specific CD8+ T-cell responses but also subdominant ones resulting in a change in the CD8+ T-cell immunodominance.
Analysis of in vitro anti-SIV efficacy of CD8+ cells
We performed in vitro viral suppression assay to examine the ability of CD8+ cells pre-ART, during ART and post-ART to suppress replication of SIVs derived from autologous peripheral blood mononuclear cells (PBMCs) pre-ART (referred to as pre-ART SIVs). The ability of CD8+ cells to suppress replication of SIV derived from pre-ART PBMCs (anti-pre-ART SIV CD8+ cell efficacy) was examined by using pre-ART CD8- cells obtained at week 10 post-infection from individual animals as the target cells. CD8+ cells derived from PBMCs at weeks 10, 27 (1 week after the first vaccination in Group V) and 34 were prepared as effector cells pre-ART, under ART and post-ART, respectively, and reductions in SIV production from the target cells by coculture with autologous CD8+ cells pre-ART, under ART and post-ART were assessed, respectively (Fig. 4a). The ability of CD8+ cells under ART to suppress replication of pre-ART SIVs was marginal and/or reduced compared to CD8+ cells pre-ART in Group N but enhanced in all the Group V except for one animal (VE1). The ratio of anti-pre-ART SIV efficacy of CD8+ cells under ART to pre-ART was significantly higher in Group V than Group N (P = 0.0173 by Mann–Whitney U test) (Fig. 4b), implying that the therapeutic SeV-Gag/Vif vaccination can augment in vitro anti-SIV efficacy of CD8+ cells. The ability of CD8+ cells post-ART to suppress replication of pre-ART SIVs was much higher than that under ART in all the Group N animals, and there was no significant difference in the ratio of anti-pre-ART SIV efficacy of CD8+ cells post-ART to pre-ART between Groups N and V.

In vitro efficacy of CD8+ cells against pre-ART PBMC-derived SIV replication. (a) In vitro ability of CD8+ cells to suppress replication of SIVs derived from autologous pre-ART PBMCs in Groups N and V. CD8− cells (target) derived from PBMCs at week 10 were cocultured with autologous CD8+ cells (effector) derived from PBMCs at week 10 (pre-ART), 27 (under ART), or 34 (post-ART), and the culture supernatants were subjected to analysis of SIV p27 concentrations by ELISA. The ratios of p27 concentrations in the supernatants from the coculture to those without CD8+ cells are shown as fold of reduction in replication of pre-ART PBMC-derived SIVs (in vitro anti-pre-ART SIV CD8+ cell efficacy). ND, not determined because of the limitation of sample availability. (b) Comparison of the ratios of anti-pre-ART SIV efficacy of CD8+ cells under ART (left panel) or post-ART (right panel) to pre-ART between Groups N and V. Group V showed significantly higher ratio of anti-pre-ART SIV efficacy of CD8+ cells under ART to pre-ART than Group N (P = 0.0173 by Mann–Whitney U test).
Next, correlation analyses were performed between in vitro anti-pre-ART SIV efficacy of CD8+ cells and antigen-specific CD8+ T-cell responses in the Group V animals. Anti-SIV efficacy of CD8+ cells pre-ART (at week 10) showed no correlation with Gag-specific nor Vif-specific CD8+ T-cell responses at week 10 post-infection (Fig. 5). Anti-pre-ART SIV efficacy of CD8+ cells under ART (at week 27) was significantly correlated with Gag-specific CD8+ T-cell responses (R = 0.9429, P = 0.0167 by Spearman test) but not with Vif-specific CD8+ T-cell responses at week 27 post-infection (1 week after the first vaccination) (Fig. 5). This indicates that induction of Gag-specific CD8+ T-cell responses by vaccination under ART can result in augmentation of the ability of CD8+ cells to suppress replication of pre-ART SIVs. However, the anti-pre-ART SIV efficacy of CD8+ cells post-ART showed no significant correlation with Gag-specific nor Vif-specific CD8+ T-cell responses at week 27 post-infection (Fig. 5).

Correlation analyses of anti-SIV CD8+ cell efficacy against pre-ART PBMC-derived SIVs with Gag/Vif-specific CD8+ T-cell responses in the vaccinated animals. Correlation analysis between anti-SIV efficacy of CD8+ cells pre-ART (left panels), under ART (middle panels) or post-ART (right panels) against pre-ART PBMC-derived SIVs and Gag-specific (upper panels) or Vif-specific (lower panels) CD8+ T-cell frequencies at week 12 (left panels), 27 (middle panels) or 34 (right panels) post-infection was performed.
Then, the ability of CD8+ cells to suppress replication of SIVs derived from autologous PBMCs post-ART (referred to as post-ART SIV) was examined by using post-ART CD8- cells obtained at week 34 post-infection from individual Group V animals as the target cells (Fig. 6a). The ability of CD8+ cells under ART to suppress replication of post-ART SIVs (anti-post-ART SIV efficacy of CD8+ cells under ART) showed no significant correlation with Gag-specific nor Vif-specific CD8+ T-cell responses at week 27 post-infection (Fig. 6b). Instead, anti-post-ART SIV efficacy of CD8+ cells post-ART exhibited a significant correlation with Gag-specific CD8+ T-cell responses (R = 0.8857, P = 0.0333) but not with Vif-specific CD8+ T-cell responses at week 34 post-infection (2 weeks after ART cessation) (Fig. 6b).

In vitro efficacy of CD8+ cells against post-ART PBMC-derived SIV replication. (a) In vitro ability of CD8+ cells to suppress replication of SIVs derived from autologous post-ART PBMCs in Group V. CD8- cells (target) derived from PBMCs at week 34 were cocultured with autologous CD8+ cells (effector) derived from PBMCs at week 27 (under ART) or 34 (post-ART), and the culture supernatants were subjected to analysis of SIV p27 concentrations by ELISA. The ratios of p27 concentrations in the supernatants from the coculture to those without CD8+ cells are shown as fold of reduction in replication of post-ART PBMC-derived SIV (in vitro anti-post-ART SIV CD8+ cell efficacy). (b) Correlation analysis between anti-post-ART SIV efficacy of CD8+ cells under ART (left panels) or post-ART (right panels) and Gag-specific (upper panels) or Vif-specific (lower panels) CD8+ T-cell frequencies at week 27 (left panels) or 34 (right panels) post-infection.
There were some discrepancies between anti-pre-ART SIV and anti-post-ART SIV efficacy of CD8+ cells as expected. Analysis found some difference in viral gag and vif sequences between individual pre-ART and post-ART SIVs obtained from the vaccinated macaque PBMCs for in vitro viral suppression assay (Fig. 7). Additional non-synonymous mutations in post-ART SIV gag were detected in macaques VW5 (GagM457V) and VS6 (GagK28R). Especially in the latter macaque VS6, Gag20–34 peptide-specific CD8+ T-cell responses were detected, and CD8+ cells under ART (at week 27) showed much lower anti-post-ART SIV efficacy compared to anti-pre-ART SIV efficacy, suggesting that the GagK28R mutation may be responsible for the difference between anti-pre-ART SIV and anti-post-ART SIV efficacy of CD8+ cells.

Non-synonymous gag and vif mutations in the vaccinated macaques. Viral gag and vif cDNAs were amplified from pre-ART and post-ART CD8- cells-derived SIVs obtained at in vitro viral suppression assay and subjected to sequence analyses. Viral gag and vif cDNAs were also amplified from plasma-derived RNAs at weeks 12 and 34 and subjected to sequence analyses. + indicates that the non-synonymous mutation resulting in amino acid substitution was detected predominantly or equivalently with the wild-type sequence. (+) indicates that the wild-type sequence was predominant while the non-synonymous mutation was subdominantly detected.
Correlation analyses between in vitro anti-SIV efficacy of CD8+ cells and plasma viral load in the vaccinated macaques
Finally, we examined correlation of in vitro anti-SIV CD8+ cell efficacy with plasma viral loads at weeks 34 and 36 (2 and 4 weeks after ART cessation) in Group V animals. There was no significant correlation between these plasma viral loads after ART and anti-pre-ART SIV efficacy of CD8+ cells under ART (Fig. 8). In contrast, anti-post-ART SIV efficacy of CD8+ cells post-ART was inversely correlated with plasma viral loads at weeks 34 (R = − 0.8857, P = 0.0333 by Spearman test) and 36 (R = − 0.9429, P = 0.0167), respectively (Fig. 8). Anti-post-ART SIV efficacy of CD8+ cells under ART exhibited a non-significant trend of association with lower viral load at week 34 (R = − 0.7714, P = 0.1028) but a significant inverse correlation with plasma viral load at week 36 (R = − 0.8857, P = 0.0333) (Fig. 8).

Correlation analyses between anti-SIV CD8+ cell efficacy and plasma viral loads in the vaccinated animals. Correlation analysis between anti-pre-ART SIV efficacy of CD8+ cells under ART (left panels) or anti-post-ART SIV efficacy of CD8+ cells under ART (middle panels) or post-ART (right panels) and viral loads at week 34 (upper panels) or 36 (lower panels) post-infection.
Discussion
Prophylactic vaccines which are administered to naive (unvaccinated) individuals can dominantly induce vaccine antigen-specific CD8+ T-cell responses. In contrast, therapeutic vaccines are supposed to be administered to individuals already primed by HIV infection. Some HIV-infected individuals have MHC/HLA (human leukocyte antigen) class I genotypes associated with predominant Gag/Vif-specific CD8+ T-cell response but others have those with predominant CD8+ T-cell responses targeting non-Gag/Vif antigens. It is important to define the patterns of CD8+ T-cell responses after therapeutic vaccination in the latter cases as well as the former. In the present study, we examined the impact of therapeutic SeV-Gag/Vif vaccination under ART on SIV antigen-specific CD8+ T-cell responses in SIVmac239-infected rhesus macaques. In the three Group V macaques possessing MHC-I haplotype W/S associated with predominant Gag/Vif-specific CD8+ T-cell responses, Gag/Vif-specific CD8+ T-cell responses were enhanced after SeV-Gag/Vif vaccination and remained predominant after ART cessation (at week 34). In the remaining three Group V macaques possessing MHC-I haplotype E associated with predominant non-Gag/Vif antigen-specific CD8+ T-cell responses, Gag/Vif-specific CD8+ T-cell responses were enhanced to become predominant after SeV-Gag/Vif vaccination and remained predominant even after ART cessation (at week 34), although Nef/Env-specific CD8+ T-cell responses were predominant post-ART in the E-positive Group N macaques. These results indicate that therapeutic SeV-Gag/Vif vaccination under ART can enhance even subdominant Gag/Vif-specific CD8+ T-cell responses and change the CD8+ T-cell immunodominance.
Early diagnosis of HIV infection is not easy, and most HIV-infected individuals start ART after the viremia setpoint. We then began ART in macaques at week 12 post-infection, after the viremia setpoint with induction of SIV-specific T-cell responses, in the present study. The first therapeutic vaccination was performed 3 months after the ART initiation, when SIV-specific CD8+ T-cell responses were largely reduced. This protocol is considered to be good for the analysis of changes in immunodominance by therapeutic vaccination described above. However, CD8+ T-cell escape SIV variants are expected to appear during the 12 weeks before the ART initiation. We thus examined efficacy of CD8+ cells to suppress replication of autologous PBMC-derived SIVs as described in the Materials and Methods, although it is difficult to exactly adjust effector-target ratios in the analysis. Analysis of the ability of CD8+ cells to suppress replication of autologous PBMC-derived SIVs revealed an increase in in vitro anti-SIV efficacy of CD8+ cells by the first SeV-Gag/Vif vaccination under ART, confirming that therapeutic SeV-Gag/Vif vaccination can augment anti-SIV efficacy of CD8+ cells. The anti-pre-ART SIV efficacy of CD8+ cells at week 27 (1 week after the first SeV-Gag/Vif vaccination) was significantly correlated with Gag-specific CD8+ T-cell responses at week 27, implying that induction of Gag-specific CD8+ T-cell responses by therapeutic vaccination can result in augmentation of anti-SIV efficacy of CD8+ cells. The vaccinated animals showed rebound viremia after ART cession, indicating that these augmented CD8+ cells are not able to contain SIV replication without ART. To increase the possibility of controlling viremia rebound after ART cession, anti-SIV IgGs were administered to help CD8+ T cell-based viremia control, but the administered anti-SIV IgGs were considered to be insufficient for showing any efficacy in our results. At week 34 (2 weeks after ART cessation), however, Gag-specific CD8+ T-cell responses were correlated with anti-post-ART SIV efficacy of CD8+ cells, which was inversely correlated with rebound plasma viral loads. Thus, augmentation of anti-SIV efficacy of CD8+ cells by vaccine-mediated Gag-specific CD8+ T-cell induction may contribute to more stable viral control under ART.
Current attempts of therapeutic vaccination inducing SIV-specific T-cell responses under ART in macaques have shown promising results, delay or control of viremia rebound after ART cession32,33. However, our experiments did not exhibit significant effect on viremia rebound after ART. One possible important difference between these previous studies and the present study may be the time point of ART initiation. ART was initiated in a week post-infection in the previous studies but at week 12 in the present study. It is speculated that CD8+ T-cell escape SIV variants appear during the 12 weeks post-infection14,34, which can disturb the efficacy of wild-type antigen-specific CD8+ T-cell responses induced by vaccination against viral replication. Indeed, several gag/vif mutations were detected before the ART initiation at week 12. Therefore, induction of CD8+ T-cell responses targeting Gag, the most conserved HIV/SIV antigen, by therapeutic vaccination could have greater impact on viral control as indicated by our results. In addition, SIV-specific T-cell responses induced before ART initiation post-infection can affect dominant/subdominant T-cell responses induced by therapeutic vaccination as described above. These results would provide insights into the development of therapeutic vaccination under ART initiated after the viremia setpoint.
In summary, the present study showed that therapeutic SeV-Gag/Vif vaccination under ART initiated after the viremia setpoint can enhance subdominant as well as predominant Gag/Vif-specific CD8+ T-cell responses in SIV-infected rhesus macaques. Our results indicate that induction of Gag-specific CD8+ T-cell responses by therapeutic vaccination can augment anti-virus efficacy of CD8+ cells, which may be insufficient for functional cure but result in more stable viral control under ART.
Materials and methods
Animal experiments
Animal experiments were carried out in the Institute for Frontier Life and Medical Sciences, Kyoto University after approval by the Committee on the Ethics of Animal Experiments in Kyoto University (permission number: R10-05 and R11-05) under the guidelines for animal experiments in accordance with the Guidelines for Proper Conduct of Animal Experiments established by Science Council of Japan (https://www.scj.go.jp/ja/info/kohyo/pdf/kohyo-20-k16-2e.pdf). The experiments were in accordance with Weatherall report for the use of non-human primates in research (https://royalsociety.org/topics-policy/publications/2006/weatherall-report/). Blood collection, virus inoculation, vaccination, and antibody administration were performed under ketamine anesthesia.
Two sets of similar experiments (Exp. 1 and Exp. 2) were performed using in total twelve Burmese rhesus macaques (Macaca mulatta) as shown in Table 1. We used three groups of rhesus macaques possessing the MHC-I haplotypes 90-010-Ie (E) (n = 6), 89-075-Iw (W) (n = 4), and 91-010-Is (S) (n = 2), respectively35,36,37. These twelve animals were intravenously inoculated with 1,000 50% tissue culture infective doses (TCID50) of SIVmac23938. Viral loads and T-cell responses pre-ART in macaques NE1, NE2, NW4, NS6, VE1, VE2, VW4, and VS6 were described before36. All of these macaques received ART from week 12 to 32 post-infection. During this period, food including Zidovudine/Lamivudine (AZT/3TC), Tenofovir disoproxil fumarate (TDF), and Lopinavir/Ritonavir (LPV/r) was given to the animals as described before39. The daily dosages were 300 mg of AZT and 150 mg of 3TC (Combivir, GlaxoSmithKline), 300 mg of TDF (Viread, Japan Tobacco) and 400 mg of Lopinavir and 100 mg of Ritonavir (Kaletra, Abbott Laboratories). Twelve animals were divided into Groups N (unvaccinated, n = 6) and V (vaccinated, n = 6). Group V macaques (three E+, two W+ and one S+) were intranasally immunized with 6 × 109 cell infectious units (CIU) of F-deleted replication-defective Sendai virus (SeV) vectors expressing SIVmac239 Gag (SeV-Gag) and Vif (SeV-Vif)28,29,30, respectively, at weeks 26 and 32 (2 days before ART cessation). Two Group N (NE3 and NW5) and all six Group V macaques were intravenously administered with 160 mg/4 ml of polyclonal anti-SIV IgGs at 2 days before and 4 days after ART cessation. The polyclonal anti-SIV IgG was purified from plasma of SIVmac239-infected macaques (2,520 mg IgG from approximately 380 ml of plasma) as described before40.
Analysis of antigen-specific CD8+ T-cell responses
We measured SIV antigen-specific CD8+ T-cell frequencies by flow cytometric analysis of IFN-γ induction after specific stimulation as described previously41. Peripheral blood mononuclear cells (PBMCs) were cocultured with autologous herpesvirus papio-immortalized B-lymphoblastoid cell lines (B-LCL) pulsed with peptide pools (at a final concentration of 1 μM for each peptides) using panels of overlapping peptides spanning the entire SIVmac239 Gag, Pol, Vif, Vpx, Vpr, Tat, Rev, Env and Nef amino acid sequences in the presence of GolgiStop (monensin; BD) for 6 h. Cells were harvested and intracellular IFN-γ staining was performed using a Cytofix-Cytoperm kit (Becton Dickinson) and fluorescein isothiocyanate-conjugated anti-human CD4 (BD), peridinin chlorophyll protein-conjugated anti-human CD8 (BD), allophycocyanin-conjugated anti-human CD3 (BD) and phycoerythrin-conjugated anti-human IFN-γ monoclonal antibodies (BioLegend) (Fig. 2a). Specific CD8+ T-cell frequencies were calculated by subtracting nonspecific IFN-γ+ CD8+ T-cell frequencies from those after peptide-specific stimulation. Specific CD8+ T-cell frequencies of less than 0.05% per CD8+ T cells were considered negative.
In vitro viral suppression assay
We examined the ability of CD8+ cells to suppress replication of SIVs derived from autologous CD8− cells. PBMCs were separated into CD8+ and CD8− cells by using Macs CD8 MicroBeads (Miltenyi Biotec). CD8− cells selected from PBMCs at weeks 10 (before ART initiation) and 34 (after ART cessation) were used for targets pre-ART and post-ART, respectively. These target cells (105 cells in cases with > 5 × 104 copies/ml of plasma viral load [in NE1, NE3, NW4, NW5, VE1, VE3, VW5, and VS6 for pre-ART and VW5 for post-ART] or 2 × 105 cells in cases with < 5 × 104 copies/ml of plasma viral load) [in others]) were cultured in the presence of 2 μg/ml of phytohemagglutinin L and 20 IU/ml of recombinant human interleukin-2 (Roche Diagnostics) for two days, and then, cocultured with 5 × 104 CD8+ cells (effector) selected from PBMCs at weeks 10 (pre-ART), 27 (under ART), and 34 (post-ART), respectively. After 6 days of culture, the culture supernatants were harvested and subjected to analysis of SIV p27 concentrations by enzyme-linked immunosorbent assay (ELISA) using SIV p27 Antigen Capture Assay (Advanced Bioscience Laboratories). If the non-CD8 culture (the culture without CD8+ cells) had lower than 100 ng/ml of p27, the culture supernatants on day 8 (VE2 pre-ART, VE3 post-ART, VW4 post-ART, and VS6 pre-ART) or 10 (VE2 post-ART) were used for the analysis. Because SIV replication kinetics are considered to be different among the cultures of target cells derived from individual SIV-infected macaque PBMCs, it is difficult to exactly adjust effector-target ratios in all the in vitro viral suppression assay by using limited available samples. Then, as described above, the viral suppression was analyzed under the condition that the individual negative control target cultures without CD8+ cells were adjusted to produce more than 100 ng/ml of p27 not on day 4 but on day 6 or later. The ratios of p27 concentrations in the supernatants from the coculture to those without CD8+ cells are shown as fold of reduction (in vitro anti-SIV efficacy)42.
Analysis of viral gag and vif sequences
Viral RNAs were extracted using the high pure viral RNA kit (Roche Diagnostics) from SIVs pre-ART and post-ART, the culture supernatants of CD8- cells derived from PBMCs at weeks 10 and 34 post-infection in individual animals. Viral RNAs were also extracted from plasma samples at weeks 12 and 34. Viral gag and vif cDNAs were amplified from these RNAs by reverse transcription and nested PCR using the PrimeScript one-step RT-PCR kit version 2 (Takara) and KOD-Plus version 2 (Toyobo), and subjected to direct sequencing by using dye terminator chemistry and an automated DNA sequencer (Applied Biosystems) as described before37.
Statistical analysis
Statistical analysis was performed using Prism software version 6.0d (GraphPad Software Inc.) with significance levels set at a P value of less than 0.05. Specific CD8+ T-cell frequencies, in vitro anti-SIV efficacy and plasma viral loads were log transformed. Comparison was performed by Mann–Whitney U test. Correlation analysis was performed by Spearman test.
References
Wong, J. K. et al. Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science 278, 1291–1295. https://doi.org/10.1126/science.278.5341.1291 (1997).
Finzi, D. et al. Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science 278, 1295–1300. https://doi.org/10.1126/science.278.5341.1295 (1997).
Palmer, S. et al. Low-level viremia persists for at least 7 years in patients on suppressive antiretroviral therapy. Proc. Natl. Acad. Sci. USA 105, 3879–3884. https://doi.org/10.1073/pnas.0800050105 (2008).
Hatano, H. et al. Evidence of persistent low-level viremia in long-term HAART-suppressed, HIV-infected individuals. AIDS 24, 2535–2539. https://doi.org/10.1097/QAD.0b013e32833dba03 (2010).
Neuhaus, J. et al. Markers of inflammation, coagulation, and renal function are elevated in adults with HIV infection. J. Infect. Dis. 201, 1788–1795. https://doi.org/10.1086/652749 (2010).
Lederman, M. M. et al. Immunologic failure despite suppressive antiretroviral therapy is related to activation and turnover of memory CD4 cells. J. Infect. Dis. 204, 1217–1226. https://doi.org/10.1093/infdis/jir507 (2011).
Kearney, M. F. et al. Lack of detectable HIV-1 molecular evolution during suppressive antiretroviral therapy. PLoS Pathog. 10, e1004010. https://doi.org/10.1371/journal.ppat.1004010 (2014).
Lorenzo-Redondo, R. et al. Persistent HIV-1 replication maintains the tissue reservoir during therapy. Nature 530, 51–56. https://doi.org/10.1038/nature16933 (2016).
Zicari, S. et al. Immune activation, inflammation, and non-AIDS co-morbidities in HIV-infected patients under long-term ART. Viruses 11, 200. https://doi.org/10.3390/v11030200 (2019).
Koup, R. A. et al. Temporal association of cellular immune responses with the initial control of viremia in primary human immunodeficiency virus type 1 syndrome. J. Virol. 68, 4650–4655 (1994).
Borrow, P., Lewicki, H., Hahn, B. H., Shaw, G. M. & Oldstone, M. B. Virus-specific CD8+ cytotoxic T-lymphocyte activity associated with control of viremia in primary human immunodeficiency virus type 1 infection. J. Virol. 68, 6103–6110 (1994).
Matano, T. et al. Administration of an anti-CD8 monoclonal antibody interferes with the clearance of chimeric simian/human immunodeficiency virus during primary infections of rhesus macaques. J. Virol. 72, 164–169 (1998).
Schmitz, J. E. et al. Control of viremia in simian immunodeficiency virus infection by CD8+ lymphocytes. Science 283, 857–860. https://doi.org/10.1126/science.283.5403.857 (1999).
Goulder, P. J. & Watkins, D. I. Impact of MHC class I diversity on immune control of immunodeficiency virus replication. Nat. Rev. Immunol. 8, 619–630. https://doi.org/10.1038/nri2357 (2008).
Dalod, M. et al. Broad, intense anti-human immunodeficiency virus (HIV) ex vivo CD8(+) responses in HIV type 1-infected patients: comparison with anti-Epstein–Barr virus responses and changes during antiretroviral therapy. J. Virol. 73, 7108–7116 (1999).
Addo, M. M. et al. Comprehensive epitope analysis of human immunodeficiency virus type 1 (HIV-1)-specific T-cell responses directed against the entire expressed HIV-1 genome demonstrate broadly directed responses, but no correlation to viral load. J. Virol. 77, 2081–2092. https://doi.org/10.1128/jvi.77.3.2081-2092.2003 (2003).
Kinloch-deLoes, S. et al. Impact of therapeutic immunization on HIV-1 viremia after discontinuation of antiretroviral therapy initiated during acute infection. J. Infect. Dis. 192, 607–617. https://doi.org/10.1086/432002 (2005).
Autran, B. et al. Greater viral rebound and reduced time to resume antiretroviral therapy after therapeutic immunization with the ALVAC-HIV vaccine (vCP1452). AIDS 22, 1313–1322. https://doi.org/10.1097/QAD.0b013e3282fdce94 (2008).
García, F. et al. A dendritic cell-based vaccine elicits T cell responses associated with control of HIV-1 replication. Sci. Transl. Med. 5, 166ra2. https://doi.org/10.1126/scitranslmed.3004682 (2013).
Pollard, R. B. et al. Safety and efficacy of the peptide-based therapeutic vaccine for HIV-1, Vacc-4x: a phase 2 randomised, double-blind, placebo-controlled trial. Lancet Infect. Dis. 14, 291–300. https://doi.org/10.1016/S1473-3099(13)70343-8 (2014).
Kiepiela, P. et al. CD8+ T-cell responses to different HIV proteins have discordant associations with viral load. Nat. Med. 13, 46–53. https://doi.org/10.1038/nm1520 (2007).
Streeck, H. et al. Recognition of a defined region within p24 gag by CD8+ T cells during primary human immunodeficiency virus type 1 infection in individuals expressing protective HLA class I alleles. J. Virol. 81, 7725–7731. https://doi.org/10.1128/JVI.00708-07 (2007).
Schneidewind, A. et al. Escape from the dominant HLA-B27-restricted cytotoxic T-lymphocyte response in Gag is associated with a dramatic reduction in human immunodeficiency virus type 1 replication. J. Virol. 81, 12382–12393. https://doi.org/10.1128/JVI.01543-07 (2007).
Kawada, M. et al. Gag-specific cytotoxic T-lymphocyte-based control of primary simian immunodeficiency virus replication in a vaccine trial. J. Virol. 82, 10199–10206. https://doi.org/10.1128/JVI.01103-08 (2008).
Miura, T. et al. HLA-B57/B*5801 human immunodeficiency virus type 1 elite controllers select for rare gag variants associated with reduced viral replication capacity and strong cytotoxic T-lymphocyte [corrected] recognition. J. Virol. 83, 2743–2755. https://doi.org/10.1128/JVI.02265-08 (2009).
Julg, B. et al. Enhanced anti-HIV functional activity associated with Gag-specific CD8 T-cell responses. J. Virol. 84, 5540–5549. https://doi.org/10.1128/JVI.02031-09 (2010).
Mudd, P. A. et al. Vaccine-induced CD8+ T cells control AIDS virus replication. Nature 491, 129–133. https://doi.org/10.1038/nature11443 (2012).
Iwamoto, N. et al. Control of simian immunodeficiency virus replication by vaccine-induced Gag- and Vif-specific CD8+ T cells. J. Virol. 88, 425–433. https://doi.org/10.1128/JVI.02634-13 (2014).
Takeda, A. et al. Protective efficacy of an AIDS vaccine, a single DNA priming followed by a single booster with a recombinant replication-defective Sendai virus vector, in a macaque AIDS model. J. Virol. 77, 9710–9715. https://doi.org/10.1128/jvi.77.17.9710-9715.2003 (2003).
Matano, T. et al. Cytotoxic T lymphocyte-based control of simian immunodeficiency virus replication in a preclinical AIDS vaccine trial. J. Exp. Med. 199, 1709–1718. https://doi.org/10.1084/jem.20040432 (2004).
Nyombayire, J. et al. First-in-human evaluation of the safety and immunogenicity of an intranasally administered replication-competent Sendai virus-vectored HIV type 1 Gag vaccine: induction of potent T-cell or antibody responses in prime-boost regimens. J. Infect. Dis. 215, 95–104. https://doi.org/10.1093/infdis/jiw500 (2017).
Borducchi, E. N. et al. Ad26/MVA therapeutic vaccination with TLR7 stimulation in SIV-infected rhesus monkeys. Nature 540, 284–287. https://doi.org/10.1038/nature20583 (2016).
Virnik, K. et al. Immunotherapy with DNA vaccine and live attenuated rubella/SIV gag vectors plus early ART can prevent SIVmac251 viral rebound in acutely infected rhesus macaques. PLoS ONE 15, e0228163. https://doi.org/10.1371/journal.pone.0228163 (2020).
O’Connor, D. H. et al. Acute phase cytotoxic T lymphocyte escape is a hallmark of simian immunodeficiency virus infection. Nat. Med. 8, 493–499. https://doi.org/10.1038/nm0502-493 (2002).
Naruse, T. K. et al. Diversity of MHC class I genes in Burmese-origin rhesus macaques. Immunogenetics 62, 601–611. https://doi.org/10.1007/s00251-010-0462-z (2010).
Nakamura, M. et al. Major histocompatibility complex class I-restricted cytotoxic T lymphocyte responses during primary simian immunodeficiency virus infection in Burmese rhesus macaques. Microbiol. Immunol. 55, 768–773. https://doi.org/10.1111/j.1348-0421.2011.00384.x (2011).
Nomura, T. et al. Association of major histocompatibility complex class I haplotypes with disease progression after simian immunodeficiency virus challenge in Burmese rhesus macaques. J. Virol. 86, 6481–6490. https://doi.org/10.1128/JVI.07077-11 (2012).
Kestler, H. W. 3rd. et al. Importance of the nef gene for maintenance of high virus loads and for development of AIDS. Cell 65, 651–662. https://doi.org/10.1016/0092-8674(91)90097-i (1991).
Horiike, M. et al. Lymph nodes harbor viral reservoirs that cause rebound of plasma viremia in SIV-infected macaques upon cessation of combined antiretroviral therapy. Virology 423, 107–118. https://doi.org/10.1016/j.virol.2011.11.024 (2012).
Yamamoto, H., Kawada, M., Takeda, A., Igarashi, H. & Matano, T. Post-infection immunodeficiency virus control by neutralizing antibodies. PLoS ONE 2, e540. https://doi.org/10.1371/journal.pone.0000540 (2007).
Ishii, H. et al. Association of lymph-node antigens with lower Gag-specific central-memory and higher Env-specific effector-memory CD8+ T-cell frequencies in a macaque AIDS model. Sci. Rep. 2, 30153. https://doi.org/10.1038/srep30153 (2016).
Ishii, H. et al. A novel immunogen selectively eliciting CD8+ T cells but not CD4+ T cells targeting immunodeficiency virus antigens. J. Virol. https://doi.org/10.1128/JVI.01876-19 (2020).
Acknowledgements
We thank Y. Koyanagi for his encouragement of animal experiments. This work was supported by the Ministry of Education, Culture, Sports, Science and Technology (MEXT) in Japan (JSPS Grants-in-Aid for Scientific Research [20390131, 23390115, 18H02666 to T.M.]), Japan Agency for Medical Research and Development (AMED [JP19fk0410011, JP18fk0410003, JP19fk0108049, JP19fk0108038, JP19jk0210002, and JP19kk0205024 to T.M.]).
Author information
Authors and Affiliations
Contributions
M.N.H., Y.T., and T.M. conceived and designed the experiments. T.T. and T.S. prepared SeV vector vaccines. H.S. and T.M. performed macaque experiments. M.N.H., Y.T., H.I., S.S., S.M., T.N., and H.Y. performed analytical experiments. M.N.H., Y.T., and T.M. analyzed the data. M.N.H. and T.M. wrote the paper. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
TS and TMatano are the inventors on Patent Cooperation Treaty (PCT) #8217019 concerning SeV vector HIV vaccines. Authors have no other competing interest to declare.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nakamura-Hoshi, M., Takahara, Y., Matsuoka, S. et al. Therapeutic vaccine-mediated Gag-specific CD8+ T-cell induction under anti-retroviral therapy augments anti-virus efficacy of CD8+ cells in simian immunodeficiency virus-infected macaques. Sci Rep 10, 11394 (2020). https://doi.org/10.1038/s41598-020-68267-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-68267-w
- Springer Nature Limited