
Abstract
Myocardial infarction (MI) is a leading cause of death worldwide. Reperfusion is considered as an optimal therapy following cardiac ischemia. However, the promotion of a rapid elevation of O2 levels in ischemic cells produces high amounts of reactive oxygen species (ROS) leading to myocardial tissue injury. This phenomenon is called ischemia reperfusion injury (IRI). We aimed at identifying new and effective compounds to treat MI and minimize IRI. We previously studied heart regeneration following myocardial injury in zebrafish and described each step of the regeneration process, from the day of injury until complete recovery, in terms of transcriptional responses. Here, we mined the data and performed a deep in silico analysis to identify drugs highly likely to induce cardiac regeneration. Fisetin was identified as the top candidate. We validated its effects in an in vitro model of MI/IRI in mammalian cardiac cells. Fisetin enhances viability of rat cardiomyocytes following hypoxia/starvation – reoxygenation. It inhibits apoptosis, decreases ROS generation and caspase activation and protects from DNA damage. Interestingly, fisetin also activates genes involved in cell proliferation. Fisetin is thus a highly promising candidate drug with clinical potential to protect from ischemic damage following MI and to overcome IRI.
Similar content being viewed by others

Introduction
Myocardial infarction (MI), the most common ischemic heart disease, is a leading cause of death worldwide1. It is a multifactorial disease: besides genetic predispositions, other parameters, e.g., smoking and obesity, are considered as prominent risk factors2. MI is most often caused by the blockade of a coronary artery after the rupture of an atherosclerotic plaque. This obstruction prevents blood supply (ischemia) to a part of the heart. Consequently, heart cells normally supplied with oxygen and nutrients by the impacted coronary artery die and are replaced by a fibrotic scar. While formation of this scar is crucial to prevent the rupture of the ventricular wall, excessive fibrotic response and reactive fibrosis in the uninjured myocardium can ultimately lead to left ventricular dysfunction, heart failure and death3.
During MI, myocardial cells are deprived from oxygen (hypoxia) and nutrients (starvation), leading to cell death. Apoptosis appears to play a predominant role in cardiomyocyte loss after MI, both in the infarcted and peri-infarcted myocardium. It determines infarct size, degree of left ventricular remodeling and onset of heart failure4,5. Two main apoptotic pathways can be observed: the intrinsic or mitochondrial pathway, and the extrinsic or death receptor pathway. Both are linked, can influence each other and converge on the same terminal signalling resulting in DNA fragmentation, protein degradation, formation of apoptotic bodies and ingestion by phagocytic cells6. The extrinsic apoptotic pathway is activated by extracellular signals via ligands interacting with transmembrane death receptors, members of the tumor necrosis factor (TNF) superfamily7,8,9. The intrinsic apoptotic pathway is induced by different stimuli, such as hypoxia, deprivation of growth factors or oxidative stress. This pathway is activated when the mitochondrial membrane, whose integrity is regulated by BCL-2 family members, is permeabilized7,10,11. Caspases, a family of cytoplasmic endoproteases, are key regulators of apoptosis7. Following MI, cells suffer both from hypoxia and deprivation of growth factors, but also from oxidative stress due to the direct decrease in O2 level and the generation of reactive oxygen species (ROS)12,13. Mitochondria are the main consumers of O2 and the major producers of ROS in the cell. At physiological levels, ROS modulate various cellular processes, including: hypoxic response, growth factor signaling and inflammation. However, at higher concentration, ROS promote damages in DNA, proteins and membrane lipids, i.e., oxidative stress. Consequently, the cellular level of ROS must be highly regulated by the antioxidative capacity of the cell. In response to hypoxia, mitochondria modify their metabolism through modification of their respiratory chain and activation of hypoxia inducible factors (HIF) to keep ROS at relatively low levels13,14,15.
MI therapies aim at quickly restoring the blood flow to the heart, using either medications to dissolve the thrombotic clot or surgery, e.g., percutaneous coronary intervention (PCI). However, although reoxygenation is crucial for patient survival, reperfusion promotes a rapid elevation in O2 levels in ischemic cells, which induces a high production of ROS in mitochondria resulting in cardiomyocytes injury. This phenomenon, called ischemia reperfusion injury (IRI), can lead to cardiac remodeling and heart failure16,17. Because of these challenges and a relatively slow progress in the development of new drugs, there is a crucial need for new effective therapies to treat MI and overcome IRI.
To meet this clinically-relevant need, we implemented here a systematic approach for drug repositioning, which tapped into an extensive collection of expression signatures obtained from treated cancer cell lines. Our hypothesis was that a drug predicted to induce, in vitro, expression signatures observed at different key stages of heart regeneration in the zebrafish could also represent a promising compound for cardiac damage healing and IRI reduction. We selected the top candidate compound: fisetin, a flavonoid. Flavonoids, phytonutrients present in almost all kinds of fruits and vegetables, are polyphenolic compounds that fulfill many functions in plants18. They have recently emerged as powerful antioxidants providing health benefits in humans, protecting from degenerative diseases linked to oxidative stress such as cancer, diabetes and cardiovascular diseases19,20.
This study follows up our previous findings, where we studied the different steps of heart regeneration following myocardial injury in the zebrafish21. We were able to elucidate the dynamic gene co-expression network associated with key stages of heart regeneration following injury. Here, based on these data and the above-stated hypothesis, we developed a new computational method to find drugs capable to induce the transcriptional patterns required for heart regeneration as observed in the zebrafish. We predicted and ranked candidate drugs, and performed experiments on mammalian cardiac cells subjected to an in vitro MI/IRI model to validate the effect of the top candidate drug: fisetin. Results indicate that fisetin is able to enhance cell viability of rat cardiomyocytes following hypoxia/starvation – reoxygenation, to protect from apoptotic cell death, by decreasing ROS generation as well as caspases activation, and to reduce DNA damage. These findings put forward fisetin as an excellent candidate for limiting cardiac damage due to ischemia following MI and for overcoming IRI.
Results
Identification of candidate compounds for repositioning through a systematic computational prediction strategy
We generated an integrated, statistically-ranked list of compounds positively (or negatively) matched to the expression signatures identified in our heart regeneration gene expression dataset (Fig. 1, Methods). Our strategy is based on the exact estimation of probability distributions of rank product statistics. In our project, such signatures represented the dynamic transcriptional changes observed at each time point during the regeneration process as compared to 4 hours after injury: 5 time points, from 1 day to 90 days post-injury. We analyzed hundreds of gene expression signatures potentially relevant and sufficiently robust to mirror salient molecular states associated with the regeneration process. Based on a combination of computational processing and human expert analysis, we selected different sets of most relevant dynamic signatures for different post-injury (and reference) time points. The size of these signatures ranged from a few dozen to hundreds of genes, and they were significantly enriched in diverse biological processes relevant to cardiac regeneration. Next, we focused on a set of 7 signatures that accurately and meaningfully represented key stages of the regeneration process. They included, for example, a signature representing expression changes at 1 day post-injury (in relation to 4 hrs post-injury).

Systems-level prediction of candidate drugs with potential to induce transcriptional responses as observed in vivo during heart regeneration in the zebrafish. We predicted candidate compounds for repositioning based on a new computational strategy that matches multiple heart regeneration expression signatures with drug-induced expression profiles in the Connectivity Map (CMap).
We identified, ranked and curated a list of candidate compounds for repositioning. Such compounds were predicted to either induce or inhibit transcriptional programs that closely resemble those observed during the zebrafish regeneration process. This resulted in the identification of fisetin as the top repositioning candidate (integrated p-value = 2.17E-11). We also identified withaferin A, a compound with diverse clinical applications including in cancer22, as negatively connected to cardiac cell regeneration in the zebrafish (integrated p-value = 1.80E-09). Thus, withaferin was predicted as a compound that is detrimental to cardiac cell regeneration by inducing, unlike fisetin, transcriptional signatures negatively correlated with the expression signatures observed during heart regeneration. Full list of the predicted positive and negative candidate drugs are available on https://lih-biomod.lu/infused/.
Fisetin enhances viability of rat H9c2 cardiomyocytes following hypoxia/starvation - reoxygenation
In order to study the effect of fisetin on cell survival, we tested ten different concentrations of the drug (0, 5, 10, 15, 20, 30, 40, 60, 80 and 100 μM) on H9c2 cardiomyocytes. Cells were cultured under normoxia, in hypoxia/starvation (HS) or subjected to hypoxia/starvation – reoxygenation (HS/R)23,24. In each experiment, fisetin was applied during 24 h before cell analysis (see Methods). Cell viability was assessed by CyQuant Assay (Fig. 2A). Fisetin showed no sign of cell toxicity, even at the highest concentration (100 μM) and didn’t affect the viability of cardiomyocytes cultured in normoxia. Compared to cells cultured in normoxia, HS/R and HS caused a decrease of 75% and 40% in cell survival, respectively. Fisetin significantly increased cell survival, with a peak at 15 μM where the drug restored cell viability by up to 70% for cells subjected to HS/R. For cells cultured in HS, the peak in viability was at 20 μM with 90% of cell survival. On the contrary, withaferin A, decreased cell survival in a concentration-dependent manner, with an IC50 of 2.6 μM (Data not shown). DMSO, used as drug vehicle, didn’t affect cell viability. These results indicate that fisetin markedly enhanced survival of H9c2 cardiomyocytes following HS or HS/R. We focused our work on HS/R, our in vitro MI/IRI model. We therefore used the most effective concentration of 15 μM of fisetin in the subsequent experiments. We first validated the effect of fisetin on cell viability by FACS. H9c2 cardiomyocytes were cultured in serum free medium at 0.5%02 during 24 h to mimic hypoxia/starvation, then during 24 h in normoxic conditions in complete medium to mimic reoxygenation. When indicated (HS/R + F) fisetin was applied during the whole reoxygenation phase. As expected, 15 μM of fisetin significantly reduced the cytotoxic effect of HS/R treatment (Fig. 2B).

Fisetin enhanced the viability of H9c2 cardiomyocytes. (A) Survival tests were performed in normoxia, HS or HS/R experiments. Different concentrations of fisetin (from 0 to 100 μM) were tested. In normoxia experiments cells were cultured during 24 h in normal conditions, while in HS experiments cells were cultured in serum free DMEM at 0.5% O2, both in presence of the drug. In the HS/R experiments, cells were treated with fisetin following HS, during the whole reoxygenation phase. Cell survival was monitored by CyQuant assay. Results are expressed as the mean of three independent experiments performed in triplicate (error bars: SEM). (B) Effect of fisetin on cardiomyocytes viability upon HS/R treatment. Cell viability was measured in FACS by annexin V/PI staining: annexin V−/PI− cells were considered as viable. Normoxia: cells cultured under normoxia without fisetin (control group). HS/R: cells subjected to HS/R, treated with DMSO as vehicle control. HS/R + F: cells subjected to HS/R, treated with 15 μM fisetin. Results are expressed as the mean of three independent experiments. One-way ANOVA. Post-hoc analysis by Tukey. *P ≤ 0.05, ****P ≤ 0.0001.
Fisetin protects H9c2 cardiomyocytes from apoptotic cell death
In our experiments, survival/viability was measured by CyQuant Direct assay, based on estimation of DNA content of healthy cells which can be directly correlated with cell number, or by annexin V/PI staining which labels apoptotic cells and allows to determine the number of remaining healthy cells in a given population. Since fisetin affected cell survival/viability, i.e. cell numbers, we studied more precisely the effects of the drug on H9c2 cardiomyocytes proliferation and apoptosis following HS/R. Fisetin markedly increased proliferation of neonatal rat cardiomyocytes (2 fold) measured by Ki67 staining (Fig. 3A), a result confirmed by cell cycle analysis showing a higher number of cells in S phase following fisetin treatment (Data not shown). The effect of the drug - increase in proliferation of 0.6% - was however not statistically significant and too low to impact the global number of cells and explain the results obtained in the survival/viability experiments. In order to investigate whether fisetin was able to protect cardiomyocytes from HS/R induced cell death, we performed an annexin V/PI staining. Annexin V−/PI− cells were considered as viable cells, while annexin V+/PI− cells were considered as early apoptotic and annexin V+/PI+ cells as late apoptotic (Fig. 3B). HS/R significantly increased apoptotic cell death compared to cells cultured in normoxia (from 6.9% to 15.5% for early apoptosis, and from 1.9% to 19.5% for late apoptosis, P ≤ 0.001). In the presence of fisetin, cell apoptosis in HS/R was markedly reduced: from 15.5% to 11% for early apoptosis and from 19.5% to 1.7% for late apoptosis (P ≤ 0.001). These results clearly show the protective effect of fisetin against HS/R-induced cardiomyocyte death.

Effect of fisetin on cardiomyocytes proliferation and apoptosis. Flow cytometry experiments were carried out to measure the amount of proliferating and apoptotic cells in each experimental group. (A) Proliferation of H9c2 cells assessed by Ki67 staining. Ki67+ cells are proliferating. Results are expressed as the mean of three independent experiments performed in triplicate. One-way ANOVA. Post-hoc analysis by Tukey. No significant difference: P > 0.05. (B) Dot plot representation of H9c2 apoptotic ratio measured by annexin V/PI staining. Annexin V−/PI− cells were considered as viable, annexin V+/PI− cells were considered as early apoptotic and annexin V+/PI+ cells were considered as late apoptotic. Normoxia: control group, cells cultured under normoxia. HS/R: cells subjected to HS/R, treated with DMSO as vehicle control. HS/R + F: cells subjected to HS/R, treated with 15 μM fisetin. Results are expressed as the mean of three independent experiments performed in triplicate. Two-way ANOVA. Post-hoc analysis by Tukey. ***P ≤ 0.001.
Fisetin regulates expression of genes related to cardioprotection and to cardiomyocytes proliferation and maturation
We studied the effect of fisetin on cardioprotection, cardiomyocyte proliferation and cardiomyocyte maturation by qRT-PCR (Fig. 4). Of note, while we couldn’t detect any noticeable increase in sirt1 (sirtuin 1, a known fisetin target25) expression in our experimental model, we observed a significant up-regulation of four cardioprotective genes following fisetin treatment: hmox1 (heme oxygenase 1), il6 (interleukin 6, another known fisetin target26,27), fgf228,29 (fibroblast growth factor 2) and igf1r (insulin-like growth factor receptor 1). Moreover, while inhibition of the TGFβ1 pathway is known to protect H9c2 cardiomyocytes from IRI30,31,32,33,34,35, fisetin markedly decreased tgfβ1 expression. Interestingly, while the observed increase in Ki67+ cardiomyocytes did not reach statistical significance (Fig. 3A), we observed an increase in the expression of genes involved in cell proliferation including foxm1 (Forkhead Box M1), ccnd2 (cyclin D2), cdk6 (cyclin-dependent kinase 6), ccne1 and 2 (cyclin E), as well as cdk1 (cyclin-dependent kinase 1), whose activity is required for the cell cycle G1/S transition. This could explain the increased number of cardiomyocytes during the S phase of the cell cycle following fisetin treatment. Intriguingly, cdkn1a (p21) was highly up-regulated by fisetin in our experiments. While this result may seem in contradiction with the apparent activation of the above mentioned G1/S transition, it is possibly related to the protective role of p21 against apoptosis and DNA damage36,37. Indeed, foxm1 was also increased, and was recently shown to be a key regulator in DNA repair38. Finally, fisetin seemed to promote cardiomyocytes maturation, as indicated by the up-regulation of the cardiac muscle markers actc1 (α-actin) and actn2 (α-actinin), while elevation of cardiac progenitor markers like gata4 (GATA Binding Protein 4) and nkx2–5 (NK2 Homeobox 5) were either not detectable or not significant, respectively.

Fisetin regulated a panel of genes related to cardioprotection, proliferation and maturation. Gene expression was assessed by qRT-PCR in cells cultured under normoxia, subjected to HS/R and treated with DMSO as vehicle control (HS/R) or subjected to HS/R and treated with 15 μM fisetin (HS/R + F) during 24 h. Results are expressed as the mean of four independent experiments. Statistical significance was determined using a one-way ANOVA corrected for multiple testing with a Tukey-Kramer as post-test (corrected p-value < 0.05). Statistically significant results are indicated in bold with a star. Fold change < 1: genes down-regulated compared to normoxia (yellow). Fold change > 1: genes up-regulated compared to normoxia (blue).
Fisetin decreases ROS production
Hypoxia and reoxygenation are known to trigger an increase in ROS generation, mainly from mitochondria, leading to cell death. In our experiments fisetin protected against cell death. We therefore investigated its effect on ROS generation in H9c2 cardiomyocytes subjected to HS/R, using dihydrorhodamine 123 as ROS detection probe. ROS level was significantly elevated (1.5 fold, P ≤ 0.01) when cells were cultured in HS/R compared to cells cultured in normoxia (Fig. 5). In the presence of fisetin, ROS level in cells subjected to HS/R decreased to reach the level found in cells cultured in normoxia (P ≤ 0.01). Of note, fisetin also decreased ROS production in cardiomyocytes in HS experiments (data not shown). Altogether, fisetin prevents ROS elevation and may thus inhibit cell apoptosis by inhibiting ROS generation in cardiomyocytes subjected to HS/R.

Fisetin decreased ROS expression level in cardiomyocytes. The cell permeable, nonfluorescent dihydrorhodamine 123 probe enters the cell where it is oxidized by ROS to fluorescent rhodamine 123. Fluorescence intensity, proportional to ROS expression level, was measured by flow cytometry in each experimental group. Normoxia: control group; HS/R: cells subjected to HS/R, treated with DMSO as vehicle control; HS/R + F: cells subjected to HS/R, treated with 15 μM fisetin. Results are expressed as the mean of three independent experiments. One-way ANOVA. Post-hoc analysis by Tukey. **P ≤ 0.01.
Fisetin inhibits caspases 8, 9 and 3 activation
Caspases are key regulator enzymes of both extrinsic and intrinsic pathways of apoptosis. In order to study the putative inhibitory effect of fisetin on caspases activation, we performed caspases activity assays to measure the activity of 3 different caspases in cardiomyocytes following HS/R: caspase 8 (specific of the extrinsic apoptosis pathway), caspase 9 (activated by the mitochondrial apoptosis pathway) and caspase 3 (a final executioner caspase activated through both apoptosis pathways). The number of cells expressing activated caspases 8, 9 and 3 was highly increased when H9c2 cells were cultured in HS/R compared to cardiomyocytes cultured in normoxia (Fig. 6): 2.2 fold of increase for caspase 8 (P ≤ 0.05); 2,8 fold for caspase 9 (P ≤ 0.01) and 5 fold for caspase 3. Interestingly, fisetin was able to reverse this phenomenon, decreasing the number of caspase 8, 9 and 3 positive cells in cardiomyocytes cultured in HS/R down to the level found in cardiomyocytes cultured in normoxia. This indicates that fisetin is able to inhibit activation of caspases in both intrinsic and extrinsic apoptosis pathways in H9c2 cardiomyocytes subjected to HS/R.

Fisetin inhibited activation of caspases 8, 9 and 3 in H9c2 cells. Caspases activity was assessed by flow cytometry using active caspase staining kits specific to caspase 8, 9 and 3, respectively. Dot plots represent the percentage of cardiomyocytes expressing activated caspase 8, 9 or 3. Normoxia: control group. HS/R: cells subjected to HS/R, treated with DMSO as vehicle control. HS/R + F: cells subjected to HS/R, treated with 15 μM fisetin. Results are expressed as the mean of three independent experiments. Two-way ANOVA. Post-hoc analysis by Tukey. **P ≤ 0.01, *P ≤ 0.05.
Fisetin protects from DNA damage
Hypoxia, starvation, ROS generation and caspases activation in cells lead to DNA damage. We thus investigated whether fisetin was capable of decreasing DNA damage in cardiomyocytes subjected to HS/R, measuring 8-Hydroxyguanosine level as DNA damage marker. Intriguingly, we were not able to detect any DNA damage in cells following HS/R (data not shown). We hypothesized that cardiomyocytes may be able to repair DNA damage during the reoxygenation phase. Therefore, we aimed to detect DNA damage directly at the end of the HS period: the drug was applied during the whole hypoxia/starvation period after which cells were immediately collected and analyzed (Fig. 7). Results indicate that HS generated DNA damage in cardiomyocytes (2.9% of 8-Hydroxyguanosine positive cells when cardiomyocytes were cultured in normoxia compared to 34.6% following HS, P ≤ 0.001), while fisetin was able to reduce the proportion of damaged cells from 34.6% to 25.1% (P ≤ 0.01). Thus, fisetin shows a protective effect against DNA damage.

Fisetin protected cardiomyocytes cultured under HS from DNA damage. The proportion of damaged cells in each experimental group was measured by flow cytometry using the anti-8 Hydroxyguanosine antibody as DNA damage marker. Normoxia: control group. HS: cells in HS, treated with DMSO as vehicle control. HS + F: cells in HS, treated with 15 μM fisetin. Results are expressed as the mean of three independent experiments. ANOVA two factors (1: biological replicates/random; 2: Treatments). Post-hoc analysis by Tukey. ***P ≤ 0.001, **P ≤ 0.01.
Discussion
In a previous article, we studied cardiac regeneration in zebrafish using cryoinjury as MI model21. Unlike mammals, zebrafish has the ability to fully regenerate its heart after severe damage. The whole regeneration is tightly regulated by the dynamic interplay of multiple genes. We coupled molecular biology techniques and in silico modelling to analyze the dynamic transcriptional responses associated with each step of the regeneration process. In the current work, we built on such data to predict drugs likely to activate cardiac regeneration following MI.
Here, we aimed to identify novel therapeutic associations between the transcriptional profiles observed during heart regeneration in the zebrafish and those induced by drugs in several cancer cell lines. This was done through the systematic statistical integration of such datasets. Our hypothesis was that potentially relevant candidate drugs are those capable to induce gene expression profiles in vitro that closely resemble gene expression profiles underlying heart regeneration in vivo. Our multiple-signature matching pipeline predicted fisetin, a flavonoid, as the top candidate.
Flavonoids recently appeared to be powerful antioxidants. This antioxidant effect is mediated through different mechanisms such as inhibition of enzymes involved in ROS generation, ROS scavenging, chelation of metal ions implicated in generation of free radicals or activation of antioxidant defenses18,19. Previous studies indicate that flavonoids offer protective effects against both the initiation and the progression of atherosclerosis, which is one of the leading causes of MI39,40. They inhibit a key mechanism in the development of atherosclerosis: the oxidation of low density lipoproteins (LDL). Moreover, they protect nitric oxide (NO) from oxidation and increase its bioavailability, which helps restoring vascular endothelial function. Flavonoids also reduce inflammation by modulating pro-inflammatory cytokines like TNFα, decrease the expression of vascular cell adhesion molecules limiting thereby the recruitment of inflammatory cells to the arterial wall, inhibit platelets aggregation and enhance vaso-relaxation19,41,42,43,44. Based on their chemical structure, flavonoids can be divided in different subclasses45,46. Fisetin, a member of the flavonols family43,47, provides different beneficial effects through its anti-inflammatory48, antioxidant49, hypoglycemic50 or anti-obesity51 action. It is also known to have strong anti-cancer activity by inhibiting angiogenesis52, promoting cell cycle arrest in G2/M phase and by inducing apoptosis in cancer cells53,54. Moreover, it protects from coronary artery diseases via inhibition of atherosclerosis55.
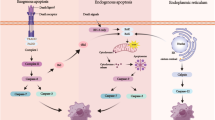
Fisetin has the advantage of being non-toxic and cheap due to its wide availability as a natural product in a huge variety of fruits and vegetables. Since IRI is a major hurdle to cardiac recovery following MI, we also investigated fisetin’s protective properties against IRI for mediating cardiac recovery. We consequently validated its cardioprotective effects in mammalian cardiac cells subjected to an in vitro model of MI/IRI. In our model, cells were cultured under hypoxia and starvation to mimic MI, then exposed to normal conditions to mimic reoxygenation before their analysis. Fisetin was supplied to the cells during the whole reoxygenation phase to simulate its application in the clinical setting. Results indicated that fisetin clearly enhanced survival of cardiomyocytes subjected to MI/IRI. The drug was able to protect cells from apoptosis, decreased ROS generation, inhibited caspases activation and protected from DNA damage (Fig. 8).

Schematic representation of fisetin cardioprotective effects on neonatal rat cardiomyocytes. Fisetin protects cells from apoptosis by targeting both intrinsic and extrinsic apoptotic pathways. Fisetin also enhances proliferation by activating expression of cell cycle activators.
The increased viability of cardiomyocytes in the presence of fisetin was mediated by the antiapoptotic effect of the drug. Fisetin inhibited expression of caspase 9, the initiator caspase specific of the intrinsic apoptotic pathway typically induced by hypoxia, starvation or oxidative stress, which are major consequences of HS/R. In view of the effect of fisetin on caspase 9 and ROS production, a deeper study of mitochondrial function following fisetin treatment is relevant to dissect the molecular pathways leading to fisetin’s anti-apoptotic effect. In addition, MI and IRI are also associated with elevated plasma level of TNFα56, one of the main activators of the extrinsic apoptotic pathway. Since fisetin inhibited expression of caspase 8, mediator of the extrinsic apoptotic pathway, we measured expression of TNFα in H9c2 cardiomyocytes culture medium after HS (6 h and over-night) and HS/R (4 h of reoxygenation) by ELISA. Intriguingly, we found that the expression of TNFα was too low to be detected in our experimental conditions (data not shown): expression of TNFα might be visible only after a very short reoxygenation period. Alternatively, fisetin treatment may instead affect regulation of other death receptors, e.g., FAS.
Furthermore, we investigated the effect of fisetin on cardioprotection. In agreement with our findings, Shanmugam et.al. recently showed the cardioprotective effect of fisetin using a Langendorff isolated heart perfusion system57. In their paper, mice were pretreated with fisetin before heart excision and induction of ischemia on the perfused heart. They showed that fisetin pretreatment attenuated myocardial injury and blunted the oxidative stress via inhibition of glycogen synthase kinase 3β (GSK3β). Despite the differences between our two experimental models - ex vivo experiments and pretreatment with fisetin prior to hypoxia vs in vitro experiments and fisetin treatment during the reoxygenation phase to mimic medication following surgery or dissolution of the thrombotic clot – our results are concordant. Our data also suggest that fisetin may have roles beyond cardiac protection, promoting cardiomyocyte proliferation and stimulating cardiomyocyte maturation. Further studies of cyclin - CDK complexes expression, degradation, phosphorylation activity and cellular localization would allow to better understand the effect of fisetin on cell cycle and DNA repair.
Summing up, we showed that fisetin protects cardiomyocytes from oxidative damage, and that further research could enable the establishment of protocols for myocardial regeneration. These findings demonstrate the value of fisetin as a candidate drug for the repositioning in MI treatment, by inhibiting ischemic damage following MI and overcoming IRI.
Methods
Brief protocol descriptions can be found hereunder. Detailed methods are described in the Supplementary Material Supplementary Methods.
Prediction of candidate drugs for repositioning
Candidate compounds were identified through the integrated matching of zebrafish heart regeneration expression signatures against expression signatures obtained from drug-treated cell lines in the Connectivity Map database (CMap, build 2), which contains more than 7 K expression profiles representing more than 1.3 K compounds58. Before implementing that procedure, we mapped gene sequences from zebrafish to humans59 and we developed an algorithm for matching multiple regeneration signatures to CMap drug signatures. Our prediction pipeline provided a statistically-ranked, integrated list of compounds predicted to have positive, pro-regeneration potential (Supplementary Methods).
Chemicals
Fisetin (3,3′,4′,7-Tetrahydroxyflavone, F4043) was purchased from Sigma-Aldrich (St Louis, MO, USA). Drug was dissolved in DMSO to a stock concentration of 100 mM. The final DMSO concentration in cell culture medium never exceeded 0.1%60.
Cell culture
Neonatal rat H9c2 cardiac cells (ATCC® CRL-1446™) were purchased from ATCC (Rockville, MD, USA), routinely cultured at 37 °C 5%CO2 in Dulbecco’s Modified Eagle’s Medium DMEM (ATCC® 30–2002™) supplemented with 10% Fetal Bovine Serum FBS (ATCC® 30–2020™) and 4 mM UltraGlutamine I (LOBE17–605E/U1, Westburg), and regularly tested for mycoplasma contamination. When sub-confluent, cells were detached with 1X Trypsin-EDTA Solution (ATCC® 30–2101™) to be subcultured.
Hypoxia/starvation – reoxygenation and drug treatment
Cells were cultured in normal conditions (20%O2/5%CO2 at 37 °C in DMEM 10%FBS) during 24 h. Hypoxia/starvation was then simulated by culturing the cells in serum free DMEM in a hypoxia incubator (0.5%O2, 5%CO2) for 24 h23. To mimic reoxygenation, cells were subsequently exposed to normal conditions (20%O2 and DMEM 10%FBS) for 24 h23,24. Fisetin, at 15 μM, was applied during the whole reoxygenation period. Control cells were treated with DMSO alone (final concentration of 0.025%) as vehicle control. Cells were harvested and analyzed at the end of the reoxygenation phase.
In the cell survival assay, a range of 10 different drug concentrations was tested (from 0 to 100 μM). For the DNA damage experiment, the drug was applied during the whole hypoxia/starvation period after which cells were immediately collected and analyzed. Normoxia control was performed by culturing the cells in normal conditions during the whole experiment.
Cell survival assay
Cell survival was assessed using the CyQuant Direct Cell Proliferation Assay Kit (C35011) from Molecular Probes (OR, USA) according to the manufacturer’s protocol (Supplementary Methods).
Cell proliferation assay
Experiments were performed by flow cytometry. Cells were harvested following drug treatment, washed, then stained with Ki67 antibody as described in Supplementary Methods.
Gene expression measurements
From 1 to 5 × 106 H9c2 cells were harvested following drug treatment and washed in 1x PBS before RNA extraction using TRI Reagent® (Sigma-Aldrich). Subsequent RNA isolation, qRT-PCR techniques and data analysis are described in Supplementary Methods.
Cell death assay
Cell death was assessed by annexin V/PI staining followed by flow cytometry as described in Supplementary Methods.
Measurement of reactive oxygen species
Cells were harvested, washed in pre-warmed to 37 °C 1x PBS, and incubated for 30 min at 37 °C in the dark in a solution of pre-warmed 1x PBS - 10 μM Dihydrorhodamine 123 (D23806, Invitrogen) at a cell density of 106 cells/mL. Cells were further washed in pre-warmed 1x PBS, resuspended in pre-warmed 1x PBS containing 1 μg/mL propidium iodide (P3566, Invitrogen) and analyzed by FACS on a BDFACS Canto™ Flow cytometer (BD Biosciences). We reported the mean intensity of three independent experiments acquired and analyzed using DIVA (BD Biosciences) and GraphPad Prism 7 (GraphPad).
Caspases activity assay
Caspases activity assays were performed using the CaspGLOW™ Fluorescein Active Caspase Staining Kits (88–7004, 88–7005 and 88–7006 for caspase-3, 8 and 9 respectively, eBioscience), following the manufacturer’s protocol. Data were acquired and analyzed by FACS on a BDFACS Canto™ Flow cytometer (BD Biosciences) using DIVA (BD Biosciences) and GraphPad Prism 7 (GraphPad). We reported the mean intensity of three independent experiments.
Measurement of DNA damage
Drugs were applied during the whole hypoxia/starvation period. Cells were harvested immediately after and subjected to flow cytometry analysis. Cells were washed in HBSS, 2% FBS, 10 mM HEPES pH7.4, stained with 1 µg/ml of LIVE/DEAD® Fixable Near-IR Dead Cell Stain Kit (L34975, Invitrogen) then fixed and permeabilized in BD Cytofix/Cytoperm™ solution (554722, BD Biosciences). Cells were then washed in BD Perm/Wash™ buffer (554723, BD Biosciences) and stained for 30 min in the dark with the FITC- Mouse Anti-8 Hydroxyguanosine antibody [15A3] (ab183393, Abcam) used at 1 µL/106 cells or the FITC- Mouse IGg2b antibody (21275533, Immunotools) as isotype control. We reported the mean intensity of three independent experiments. Data acquisition and analysis were performed using DIVA (BD Biosciences) and GraphPad Prism 7 (GraphPad).
References
Finegold, J. A., Asaria, P. & Francis, D. P. Mortality from ischaemic heart disease by country, region, and age: statistics from World Health Organisation and United Nations. Int. J. Cardiol. 168, 934–945, https://doi.org/10.1016/j.ijcard.2012.10.046 (2013).
Anand, S. S. et al. Risk factors for myocardial infarction in women and men: insights from the INTERHEART study. Eur. Heart J. 29, 932–940, https://doi.org/10.1093/eurheartj/ehn018 (2008).
Talman, V. & Ruskoaho, H. Cardiac fibrosis in myocardial infarction-from repair and remodeling to regeneration. Cell Tissue Res. 365, 563–581, https://doi.org/10.1007/s00441-016-2431-9 (2016).
Abbate, A. et al. Increased myocardial apoptosis in patients with unfavorable left ventricular remodeling and early symptomatic post-infarction heart failure. J. Am. Coll. Cardiology 41, 753–760 (2003).
Teringova, E. & Tousek, P. Apoptosis in ischemic heart disease. J. Transl. Med. 15, 87, https://doi.org/10.1186/s12967-017-1191-y (2017).
Elmore, S. Apoptosis: a review of programmed cell death. Toxicol. Pathol. 35, 495–516, https://doi.org/10.1080/01926230701320337 (2007).
McIlwain, D. R., Berger, T. & Mak, T. W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 5, a008656, https://doi.org/10.1101/cshperspect.a008656 (2013).
Samraj, A. K., Keil, E., Ueffing, N., Schulze-Osthoff, K. & Schmitz, I. Loss of caspase-9 provides genetic evidence for the type I/II concept of CD95-mediated apoptosis. J. Biol. Chem. 281, 29652–29659, https://doi.org/10.1074/jbc.M603487200 (2006).
Jost, P. J. et al. XIAP discriminates between type I and type II FAS-induced apoptosis. Nat. 460, 1035–1039, https://doi.org/10.1038/nature08229 (2009).
Cain, K., Bratton, S. B. & Cohen, G. M. The Apaf-1 apoptosome: a large caspase-activating complex. Biochim. 84, 203–214 (2002).
Li, M., Gao, P. & Zhang, J. Crosstalk between Autophagy and Apoptosis: Potential and Emerging Therapeutic Targets for Cardiac Diseases. Int. J. Mol. Sci. 17, 332, https://doi.org/10.3390/ijms17030332 (2016).
Hernansanz-Agustin, P. et al. Acute hypoxia produces a superoxide burst in cells. Free. Radic. Biol. Med. 71, 146–156, https://doi.org/10.1016/j.freeradbiomed.2014.03.011 (2014).
Fuhrmann, D. C. & Brune, B. Mitochondrial composition and function under the control of hypoxia. Redox Biol. 12, 208–215, https://doi.org/10.1016/j.redox.2017.02.012 (2017).
Gorlach, A. et al. Reactive oxygen species, nutrition, hypoxia and diseases: Problems solved? Redox Biol. 6, 372–385, https://doi.org/10.1016/j.redox.2015.08.016 (2015).
Santos, C. X., Anilkumar, N., Zhang, M., Brewer, A. C. & Shah, A. M. Redox signaling in cardiac myocytes. Free. Radic. Biol. Med. 50, 777–793, https://doi.org/10.1016/j.freeradbiomed.2011.01.003 (2011).
Lee, S. et al. Preventive role of propofol in hypoxia/reoxygenation-induced apoptotic H9c2 rat cardiac myoblast cell death. Mol. Med. Rep. 4, 351–356, https://doi.org/10.3892/mmr.2011.432 (2011).
Neri, M., Riezzo, I., Pascale, N., Pomara, C. & Turillazzi, E. Ischemia/Reperfusion Injury following Acute Myocardial Infarction: A Critical Issue for Clinicians and Forensic Pathologists. Mediators Inflamm. 2017, 7018393, https://doi.org/10.1155/2017/7018393 (2017).
Kumar, S. & Pandey, A. K. Chemistry and biological activities of flavonoids: an overview. ScientificWorldJournal 2013, 162750, https://doi.org/10.1155/2013/162750 (2013).
Scalbert, A., Manach, C., Morand, C., Remesy, C. & Jimenez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 45, 287–306, https://doi.org/10.1080/1040869059096 (2005).
Manach, C., Mazur, A. & Scalbert, A. Polyphenols and prevention of cardiovascular diseases. Curr. Opin. Lipidol. 16, 77–84 (2005).
Rodius, S. et al. Analysis of the dynamic co-expression network of heart regeneration in the zebrafish. Sci. Rep. 6, 26822, https://doi.org/10.1038/srep26822 (2016).
Lee, I. C. & Choi, B. Y. Withaferin-A–A Natural Anticancer Agent with Pleitropic Mechanisms of Action. Int. J. Mol. Sci. 17, 290, https://doi.org/10.3390/ijms17030290 (2016).
Keeley, T. P. & Mann, G. E. Defining Physiological Normoxia for Improved Translation of Cell Physiology to Animal Models and Humans. Physiological Rev. 99, 161–234, https://doi.org/10.1152/physrev.00041.2017 (2019).
Kuznetsov, A. V., Javadov, S., Sickinger, S., Frotschnig, S. & Grimm, M. H9c2 and HL-1 cells demonstrate distinct features of energy metabolism, mitochondrial function and sensitivity to hypoxia-reoxygenation. Biochim. Biophys. Acta 1853, 276–284, https://doi.org/10.1016/j.bbamcr.2014.11.015 (2015).
Kim, S. C. et al. Fisetin induces Sirt1 expression while inhibiting early adipogenesis in 3T3-L1 cells. Biochemical biophysical Res. Commun. 467, 638–644, https://doi.org/10.1016/j.bbrc.2015.10.094 (2015).
Kim, H. J., Kim, S. H. & Yun, J. M. Fisetin inhibits hyperglycemia-induced proinflammatory cytokine production by epigenetic mechanisms. Evid. Based Complement. Altern. Med. 2012, 639469, https://doi.org/10.1155/2012/639469 (2012).
Fontes, J. A., Rose, N. R. & Cihakova, D. The varying faces of IL-6: From cardiac protection to cardiac failure. Cytokine 74, 62–68, https://doi.org/10.1016/j.cyto.2014.12.024 (2015).
Manning, J. R. et al. Low molecular weight fibroblast growth factor-2 signals via protein kinase C and myofibrillar proteins to protect against postischemic cardiac dysfunction. Am. J. Physiol. Heart circulatory physiology 304, H1382–1396, https://doi.org/10.1152/ajpheart.00613.2012 (2013).
Chen, Q. et al. FGF-2 Transcriptionally Down-Regulates the Expression of BNIP3L via PI3K/Akt/FoxO3a Signaling and Inhibits Necrosis and Mitochondrial Dysfunction Induced by High Concentrations of Hydrogen Peroxide in H9c2 Cells. Cell Physiol. Biochem. 40, 1678–1691, https://doi.org/10.1159/000453217 (2016).
Zhang, H. et al. Glutamine protects cardiomyocytes from hypoxia/reoxygenation injury under high glucose conditions through inhibition of the transforming growth factor-beta1-Smad3 pathway. Arch. Biochem. Biophys. 596, 43–50, https://doi.org/10.1016/j.abb.2016.03.003 (2016).
Leask, A. & Abraham, D. J. TGF-beta signaling and the fibrotic response. FASEB journal: Off. Publ. Federation Am. Societies Exp. Biol. 18, 816–827, https://doi.org/10.1096/fj.03-1273rev (2004).
Hanna, A. & Frangogiannis, N. G. The Role of the TGF-beta Superfamily in Myocardial Infarction. Front. Cardiovasc. Med. 6, 140, https://doi.org/10.3389/fcvm.2019.00140 (2019).
Baxter, G. F., Mocanu, M. M., Brar, B. K., Latchman, D. S. & Yellon, D. M. Cardioprotective effects of transforming growth factor-beta1 during early reoxygenation or reperfusion are mediated by p42/p44 MAPK. J. Cardiovasc. Pharmacol. 38, 930–939, https://doi.org/10.1097/00005344-200112000-00015 (2001).
Schroder, D., Heger, J., Piper, H. M. & Euler, G. Angiotensin II stimulates apoptosis via TGF-beta1 signaling in ventricular cardiomyocytes of rat. J. Mol. Med. 84, 975–983, https://doi.org/10.1007/s00109-006-0090-0 (2006).
Rainer, P. P. et al. Cardiomyocyte-specific transforming growth factor beta suppression blocks neutrophil infiltration, augments multiple cytoprotective cascades, and reduces early mortality after myocardial infarction. Circulation Res. 114, 1246–1257, https://doi.org/10.1161/CIRCRESAHA.114.302653 (2014).
Cazzalini, O., Scovassi, A. I., Savio, M., Stivala, L. A. & Prosperi, E. Multiple roles of the cell cycle inhibitor p21(CDKN1A) in the DNA damage response. Mutat. Res. 704, 12–20, https://doi.org/10.1016/j.mrrev.2010.01.009 (2010).
Karimian, A., Ahmadi, Y. & Yousefi, B. Multiple functions of p21 in cell cycle, apoptosis and transcriptional regulation after DNA damage. DNA Repair. 42, 63–71, https://doi.org/10.1016/j.dnarep.2016.04.008 (2016).
Zona, S., Bella, L., Burton, M. J., Nestal de Moraes, G. & Lam, E. W. FOXM1: an emerging master regulator of DNA damage response and genotoxic agent resistance. Biochim. Biophys. Acta 1839, 1316–1322, https://doi.org/10.1016/j.bbagrm.2014.09.016 (2014).
Siasos, G. et al. Flavonoids in atherosclerosis: an overview of their mechanisms of action. Curr. Med. Chem. 20, 2641–2660 (2013).
Ambrose, J. A. & Singh, M. Pathophysiology of coronary artery disease leading to acute coronary syndromes. F1000Prime Rep. 7, 08, https://doi.org/10.12703/P7-08 (2015).
Leyva-Lopez, N., Gutierrez-Grijalva, E. P., Ambriz-Perez, D. L. & Heredia, J. B. Flavonoids as Cytokine Modulators: A Possible Therapy for Inflammation-Related Diseases. Int J Mol Sci 17, https://doi.org/10.3390/ijms17060921 (2016).
Adegbola, P., Aderibigbe, I., Hammed, W. & Omotayo, T. Antioxidant and anti-inflammatory medicinal plants have potential role in the treatment of cardiovascular disease: a review. Am. J. Cardiovasc. Dis. 7, 19–32 (2017).
Salvamani, S., Gunasekaran, B., Shaharuddin, N. A., Ahmad, S. A. & Shukor, M. Y. Antiartherosclerotic effects of plant flavonoids. Biomed. Res. Int. 2014, 480258, https://doi.org/10.1155/2014/480258 (2014).
Li, A. N. et al. Resources and biological activities of natural polyphenols. Nutrients 6, 6020–6047, https://doi.org/10.3390/nu6126020 (2014).
Manach, C., Scalbert, A., Morand, C., Remesy, C. & Jimenez, L. Polyphenols: food sources and bioavailability. Am. J. Clin. Nutr. 79, 727–747, https://doi.org/10.1093/ajcn/79.5.727 (2004).
Petrussa, E. et al. Plant flavonoids–biosynthesis, transport and involvement in stress responses. Int. J. Mol. Sci. 14, 14950–14973, https://doi.org/10.3390/ijms140714950 (2013).
Koosha, S., Alshawsh, M. A., Looi, C. Y., Seyedan, A. & Mohamed, Z. An Association Map on the Effect of Flavonoids on the Signaling Pathways in Colorectal Cancer. Int. J. Med. Sci. 13, 374–385, https://doi.org/10.7150/ijms.14485 (2016).
Seo, S. H. & Jeong, G. S. Fisetin inhibits TNF-alpha-induced inflammatory action and hydrogen peroxide-induced oxidative damage in human keratinocyte HaCaT cells through PI3K/AKT/Nrf-2-mediated heme oxygenase-1 expression. Int. Immunopharmacol. 29, 246–253, https://doi.org/10.1016/j.intimp.2015.11.014 (2015).
Kang, K. A. et al. Fisetin attenuates hydrogen peroxide-induced cell damage by scavenging reactive oxygen species and activating protective functions of cellular glutathione system. Vitro Cell Dev. Biol. Anim. 50, 66–74, https://doi.org/10.1007/s11626-013-9681-6 (2014).
Prasath, G. S. & Subramanian, S. P. Modulatory effects of fisetin, a bioflavonoid, on hyperglycemia by attenuating the key enzymes of carbohydrate metabolism in hepatic and renal tissues in streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 668, 492–496, https://doi.org/10.1016/j.ejphar.2011.07.021 (2011).
Lee, Y. & Bae, E. J. Inhibition of mitotic clonal expansion mediates fisetin-exerted prevention of adipocyte differentiation in 3T3-L1 cells. Arch. Pharm. Res. 36, 1377–1384, https://doi.org/10.1007/s12272-013-0226-z (2013).
Bhat, T. A., Nambiar, D., Pal, A., Agarwal, R. & Singh, R. P. Fisetin inhibits various attributes of angiogenesis in vitro and in vivo–implications for angioprevention. Carcinogenesis 33, 385–393, https://doi.org/10.1093/carcin/bgr282 (2012).
Haddad, A. Q. et al. Novel antiproliferative flavonoids induce cell cycle arrest in human prostate cancer cell lines. Prostate Cancer Prostatic Dis. 9, 68–76, https://doi.org/10.1038/sj.pcan.4500845 (2006).
Lu, X. et al. Fisetin inhibits the activities of cyclin-dependent kinases leading to cell cycle arrest in HT-29 human colon cancer cells. J. Nutr. 135, 2884–2890, https://doi.org/10.1093/jn/135.12.2884 (2005).
Lian, T. W., Wang, L., Lo, Y. H., Huang, I. J. & Wu, M. J. Fisetin, morin and myricetin attenuate CD36 expression and oxLDL uptake in U937-derived macrophages. Biochim. Biophys. Acta 1781, 601–609, https://doi.org/10.1016/j.bbalip.2008.06.009 (2008).
Kleinbongard, P., Schulz, R. & Heusch, G. TNFalpha in myocardial ischemia/reperfusion, remodeling and heart failure. Heart Fail. Rev. 16, 49–69, https://doi.org/10.1007/s10741-010-9180-8 (2011).
Shanmugam, K., Ravindran, S., Kurian, G. A. & Rajesh, M. Fisetin Confers Cardioprotection against Myocardial Ischemia Reperfusion Injury by Suppressing Mitochondrial Oxidative Stress and Mitochondrial Dysfunction and Inhibiting Glycogen Synthase Kinase 3beta Activity. Oxid. Med. Cell Longev. 2018, 9173436, https://doi.org/10.1155/2018/9173436 (2018).
Lamb, J. et al. The Connectivity Map: using gene-expression signatures to connect small molecules, genes, and disease. Sci. 313, 1929–1935, https://doi.org/10.1126/science.1132939 (2006).
de Klein, N., Ibberson, M., Crespo, I., Rodius, S. & Azuaje, F. A gene mapping bottleneck in the translational route from zebrafish to human. Front. Genet. 5, 470, https://doi.org/10.3389/fgene.2014.00470 (2014).
Timm, M., Saaby, L., Moesby, L. & Hansen, E. W. Considerations regarding use of solvents in in vitro cell based assays. Cytotechnology 65, 887–894, https://doi.org/10.1007/s10616-012-9530-6 (2013).
Acknowledgements
We thank C. Guerin and L. Guyonnet for technical support in flow cytometry; PV. Nazarov, A. Muller, T. Kaoma and SY. Kim for statistical and informatics support. This work was supported by FNR, the Luxembourg National Research Fund, FNR-CORE INFUSED project. At the NorLux Laboratory and the Proteome and Genome Research Unit of LIH, it was also supported by funding from Luxembourg’s Ministry of Higher Education and Research (MESR).
Author information
Authors and Affiliations
Contributions
S.R. designed and performed the experiments, analysed the data, and wrote the paper. N.D.K. generated the list of ranked candidate compounds for repositioning, and contributed to the writing of the paper. C.J. performed the qPCR experiments, analysed the qPCR data and contributed to writing the paper. I.C., M.I. and I.X. contributed to the analysis of the bioinformatics data. H.S. and N.M. participated in the analysis of the experimental data and in the writing of the paper. G.D. and S.N. provided lab resources and contributed to the writing of the paper. F.A. co-ordinated the study, analysed the bioinformatics data, and contributed to the writing of the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rodius, S., de Klein, N., Jeanty, C. et al. Fisetin protects against cardiac cell death through reduction of ROS production and caspases activity. Sci Rep 10, 2896 (2020). https://doi.org/10.1038/s41598-020-59894-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-59894-4
- Springer Nature Limited
This article is cited by
-
Gestational Fisetin Exerts Neuroprotection by Regulating Mitochondria-Directed Canonical Wnt Signaling, BBB Integrity, and Apoptosis in Prenatal VPA-Induced Rodent Model of Autism
Molecular Neurobiology (2024)
-
Mini-encyclopedia of mitochondria-relevant nutraceuticals protecting health in primary and secondary care—clinically relevant 3PM innovation
EPMA Journal (2024)
-
The toxicity of 4-tert-butylphenol in early development of zebrafish: morphological abnormality, cardiotoxicity, and hypopigmentation
Environmental Science and Pollution Research (2023)
-
Role of polyphenolic compounds and their nanoformulations: a comprehensive review on cross-talk between chronic kidney and cardiovascular diseases
Naunyn-Schmiedeberg's Archives of Pharmacology (2023)
-
Hesperidin Attenuates Oxidative Stress, Inflammation, Apoptosis, and Cardiac Dysfunction in Sodium Fluoride‐Induced Cardiotoxicity in Rats
Cardiovascular Toxicology (2022)