
Abstract
Zinc (Zn) is an important microelement for rice and plays a key role in many physiological processes. This study assessed the physio-biochemical responses involved in biosynthesis of 2-acety-1-pyrroline (2-AP), which is a key compound in the aroma of fragrant rice, in four different fragrant rice varieties, i.e., Meixiangzhan-2, Xiangyaxiangzhan, Ruanhuayou-134, and Yunjingyou. Four concentrations (0, 0.50, 1.00 and 2.00 g L−1) of zinc chloride were applied to fragrant rice foliage at the heading stage and named CK, Zn1, Zn2 and Zn3, respectively. Our results showed that compared with CK, the Zn1, Zn2 and Zn3 treatments all significantly increased the 2-AP concentration in mature grains of the four fragrant rice genotypes. Furthermore, exogenous application of Zn not only enhanced the activities of enzymes, including proline dehydrogenase (PDH), △1-pyrroline-5-carboxylic acid synthetase (P5CS), and diamine oxidase (DAO), which are involved in 2-AP biosynthesis, but also improved the contents of the related precursors, such as Δ1-pyrroline, proline and pyrroline-5-carboxylic acid (P5C). In addition, compared to the CK treatment, the Zn2 treatment markedly increased the net photosynthetic rate of fragrant rice during the grain filling stage and increased the seed-setting rate, 1000-grain weight and grain yield in all fragrant rice genotypes. Foliar application of Zn also markedly increased the grain Zn content. In general, 1.00 g L−1 seemed to be the most suitable application concentration because the highest 2-AP content and grain weight were recorded with this treatment.
Similar content being viewed by others

Introduction
Fragrant rice is a special type of rice that carries a special aroma. In recent years, the market price of fragrant rice has greatly increased1. Therefore, many researchers and farmers have begun to pay attention to fragrant rice production and quality regulation. As the most special characteristic of fragrant rice, the aroma is a determining factor in fragrant rice quality and significantly affects the price of fragrant rice2. It is generally believed that 2-acetyl-1-pyrroline (2-AP) is the primary component responsible for the fragrant rice aroma3, and a few studies have shown that the exogenous application of some metallic and non-metallic elements could markedly improve the 2-AP content in fragrant rice. For example, the study by Li et al.4 illustrated that application of manganese (Mn) induced regulation of 2-AP biosynthesis in fragrant rice. A previous study demonstrated that the application of silicon (Si) fertilizer could improve both the photosynthetic rate and 2-AP content of fragrant rice5. Thus, exogenous application of microelements could be an effective way to regulate 2-AP formation in fragrant rice.
Zinc (Zn) is an essential micronutrient for the normal growth and development of crops, and it plays an important physiological and biochemical role6. Zn is also an essential micronutrient in human health. Zn deficiency ranks 11th among more than 20 factors causing disease worldwide and 5th among the top 10 factors causing disease in developing countries7. Zn deficiency often leads to physical decline, vision loss or blindness, mental retardation, reduced immune function and disease resistance, and a high mortality and disease rate8. The study by Fu et al.9 also revealed that the reduction in photosynthesis under Zn deficiency was primarily due to non-stomatal limitations, in particular the changes in photosystem II (PS II). Therefore, in recent years, the nutritional status of plant trace elements and their relationship with human health, especially the biological fortification of iron (Fe) and Zn in cereal crops, has attracted increasing attention from researchers. Han et al.10 indicated that foliar application of Zn increased the leaf Zn concentration, thereby maintaining superoxide dismutase activity and membrane stability and protecting the photosynthetic apparatus against heat damage. Mo et al.11 revealed that supplementation with Zn could improve the 2-AP content in detached fragrant rice panicles in vitro. However, there have been no reports about the effect of foliar application of Zn on fragrant rice performance in field crop production, and the mechanism by which Zn application affects 2-AP biosynthesis in fragrant rice is still unclear.
Hence, the objective of this study was to evaluate 2-AP biosynthesis under foliar application of Zn at different concentrations using four fragrant rice varieties. The activities of five enzymes and contents of three precursors and two related compounds involved in 2-AP biosynthesis in fragrant rice were investigated. The grain yield and related trials were also estimated. We hope that this study will be useful for increasing the aroma in cultivated fragrant rice.
Results
2-AP concentration in mature grains
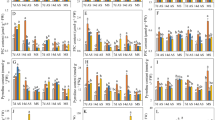
As shown in Fig. 1, different Zn applications affected the 2-AP content in grains differently. For Meixiangzhan-2, 14.83%, 21.93% and 19.03% higher 2-AP concentrations were recorded in Zn1, Zn2 and Zn3, respectively, than in CK. For Xiangyaxiangzhan, compared to CK, Zn1, Zn2 and Zn3 increased the 2-AP content by 9.47%, 14.46% and 14.02%, respectively. For Ruanhuayou-134, the 2-AP contents in grain under the Zn1, Zn2 and Zn3 treatments were 0.20-, 0.33- and 0.30-fold higher than those under CK. For Yunjingyou, compared to CK, Zn1, Zn2 and Zn3 increased the 2-AP content by 10.77%, 24.07% and 26.43%, respectively.

Effect of exogenous Zn on 2-AP content mature grains. Means sharing a common letter don’t differ significantly at (P ≤ 0.05) according to least significant difference (LSD) test.
Enzymes involved in 2-AP biosynthesis in grains
As shown in Table 1, the application of Zn significantly improved the activities of some enzymes involved in 2-AP biosynthesis in grains. Compared with CK, the Zn1, Zn2 and Zn3 treatments significantly enhanced the activities of proline dehydrogenase (PDH), △1-pyrroline-5-carboxylic acid synthetase (P5CS) and diamine oxidase (DAO), and the enzyme activity trends were recorded as CK < Zn1 < Zn2 = Zn3. However, there was no significant difference among all treatments in ornithine aminotransferase (OAT) activity, and a similar result was also observed for betaine aldehyde dehydrogenase (BADH) activity. The enzyme activity trends were similar among the four fragrant rice genotypes.
Contents of precursors and related compounds involved in 2-AP biosynthesis in grains
As shown in Table 2, exogenous application of Zn regulated the contents of some compounds involved in 2-AP biosynthesis. Compared with CK, Zn application significantly increased the contents of methylglyoxal, P5C, proline and Δ1-pyrroline. The trends of methylglyoxal, P5C, proline and Δ1-pyrroline were recorded as CK < Zn1 < Zn2 = Zn3 in all fragrant rice genotypes. There were no significant differences in the GABA content among all treatments in both Xiangyaxiangzhan and Yunjingyou. However, for Meixiangzhan-2 and Ruanhuayou-134, the GABA content of Zn3-treated plants was significantly higher than that of CK-treated plants, while there was no notable difference among the CK, Zn1 and Zn2 treatment groups.
Net photosynthetic rate at filling stage
As shown in Fig. 2, the different concentrations of Zn affected the fragrant rice differently. For Meixiangzhan-2, 7.43% and 17.16% higher net photosynthetic rates were recorded under Zn1 and Zn2, respectively, than under CK. For Xiangyaxiangzhan, compared to CK, Zn1 and Zn2 significantly increased the net photosynthetic rate by 7.26% and 20.28%, respectively. For Ruanhuayou-134, the net photosynthetic rates under Zn1 and Zn2 were 0.06 and 0.13-fold higher, respectively, than those under CK. For Yunjingyou, compared to CK, Zn1 and Zn2 increased the net photosynthetic rate at the filling stage by 7.14% and 20.57%, respectively.

Effect of exogenous Zn on the net photosynthetic rate of fragrant rice at the filling stage. Means sharing a common letter don’t differ significantly at (P ≤ 0.05) according to least significant difference (LSD) test.
Zn content in grains
As shown in Fig. 3, exogenous Zn application significantly improved the Zn content in the grains of fragrant rice. For Meixiangzhan-2, 11.16%, 25.53% and 26.78% higher Zn concentrations were recorded under Zn1, Zn2 and Zn3, respectively, than under CK. For Xiangyaxiangzhan, compared to CK, Zn1, Zn2 and Zn3 increased the 2-AP content by 10.10%, 23.81% and 22.08%, respectively. For Ruanhuayou-134, the 2-AP contents in grain under Zn1, Zn2 and Zn3 were 0.10-, 0.27- and 0.28-fold higher, respectively, than those under CK. For Yunjingyou, compared to CK, Zn1, Zn2 and Zn3 increased the 2-AP content by 7.33%, 26.04% and 23.77%, respectively.

Effect of exogenous Zn on the Zn content of mature grains. Means sharing a common letter don’t differ significantly at (P ≤ 0.05) according to least significant difference (LSD) test.
Grain yield and yield-related trials
As shown in Table 3, Zn application modulated fragrant rice yield formation to a certain degree. Compared with CK, all Zn treatments (except Zn3 in Meixiangzhan-2 and Ruanhuayou-134) significantly increased the grain yield, and the highest yield was recorded with the Zn2 treatment for all rice genotypes. Furthermore, the Zn2 treatment also significantly improved the seed-setting rate and 1000-grain weight for all rice varieties. There was no notable difference among the effects of CK, Zn1, Zn2 and Zn3 in both the effective panicle number and grain number.
Correlation analysis
As shown in Table 4, the PDH, P5CS and DAO activities were all significantly and positively correlated with the 2-AP content in mature grains. Similar correlations were also observed between the 2-AP content and the contents of methylglyoxal, P5C, proline, and Δ1-pyrroline.
As shown in Table 5, there was no significant correlation between panicle number and grain yield for all rice genotypes. Similar conditions were also found between grain number and grain yield. However, both the seed-setting rate and 1000-grain weight had a significant and positive correlation with the grain yield of the four fragrant rice genotypes.
Discussion
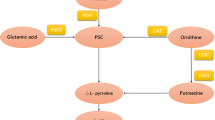
2-AP biosynthesis in fragrant rice is a complicated phenomenon. Many researchers have conducted studies on this process. For example, Yoshihashi12 showed that 2-AP was not produced at all by the cooking or postharvest processes according to a stable isotope dilution method. Moreover, Yoshihashi also discovered that adding proline, ornithine and glutamic acid to the solution increased the content of 2-AP in fragrant rice seedlings and calli13. In 2008, Chen14 demonstrated that the presence of a dominant Badh2 allele encoding betaine aldehyde dehydrogenase (BADH) inhibits the biosynthesis of 2-AP in rice because BADH decreased 2-AP formation by catalysing 4-aminobutyraldehyde (GABald) into GABA, while GABald may be one of the precursors of 2-AP biosynthesis. The investigation of Mo15 also indicated that there was a correlation between the GABA content and the 2-AP content in fragrant rice grains. Furthermore, the study of Poonlaphdecha16 proved that 1-pyrroline is an important precursor in the biosynthesis of 2-AP in fragrant rice by using rice callus cultures. In recent years, it has generally been believed that five enzymes play a key role in the process of 2-AP biosynthesis in fragrant rice: PDH, P5CS, OAT, DAO and BADH17. Therefore, one possible 2-AP synthesis pathway in fragrant rice is shown in Fig. 4.

Possible mechanism of 2-AP formation in fragrant rice.
The present study revealed the modulation of 2-AP biosynthesis and yield formation by Zn in different fragrant rice genotypes. The results showed that three concentrations of Zn all improved the 2-AP content in mature grains, and the highest improvement was recorded with Zn2 for all fragrant rice genotypes. The increase in 2-AP due to Zn application could be attributed to the enhanced activities of enzymes, including PDH, P5CS and DAO. This study showed that exogenous Zn application at the heading stage promoted the activities of PDH and P5CS during the filling stage, and the activities of these enzymes all had a significant positive correlation with the 2-AP content, which was corroborated by the early research of Deng18. PDH is a key enzyme in mitochondria that catalyses the degradation of proline to P5C, while regulatory feedback of P5CS plays an important role in controlling the plant proline level because P5CS has glutamyl-gamma-semialdehyde deaminase (GSADH) and glutamate kinase (gamma-GK) activities, and it can catalyse the phosphorylation of glutamate to glutamic semialdehyde (GSA) and reduce GSA19. Li (Li et al.4) indicated that the 2-AP concentration in fragrant rice grains was positively correlated with the activities of PDH and P5CS, along with the contents of proline, P5C, methylglyoxal and △1-pyrroline in fragrant rice, which revealed that a higher 2-AP concentration is related to increased activities of the enzymes involved in its biosynthesis. Higher contents of proline, P5C, methylglyoxal and △1-pyrroline, which were accepted as precursors and intermediates for 2-AP biosynthesis, also appeared in the present study following the Zn treatments. On the other hand, DAO transforms putrescine into GABald, which further cyclizes spontaneously to Δ1-pyrroline, or synthesizes GABA, depending on the absence or presence of functional BADH2 enzyme20. Considering the changes in enzyme activities and contents of precursors and intermediates, the modulation of 2-AP biosynthesis by exogenous Zn application in fragrant rice may occur by the mechanisms shown in Fig. 5.

Regulation of 2-AP biosynthesis by Zn application in fragrant rice.
In addition, we observed that exogenous application of Zn at a 1.00 g L−1 concentration greatly increased the grain yield for the four fragrant rice genotypes. This increase could be explained by the increase in the seed-setting rate and 1000-grain weight. The increase may further be attributed to the improvement in the net photosynthetic rate during the grain filling stage due to exogenous Zn application at the heading stage. Photosynthesis is one of the most important physiological processes in rice and significantly influences the grain yield21. An early study illustrated the importance of photosynthesis during the grain filling phase to rice yield by showing that higher net photosynthesis at the filling stage could increase grain yield22. Our result was consistent with those of Tavallali23, who demonstrated that the application of Zn could increase the net photosynthetic rate in pistachio. Previous studies also revealed that carbonic anhydrase (CA), a zinc-containing metalloenzyme that is widely distributed in plants and mainly exists in chloroplasts, could catalyse the hydration of carbon dioxide, promote the fixation of carbon dioxide in photosynthesis, and reduce the activity of CA due to zinc deficiency24,25.
Conclusion
In conclusion, foliar application of Zn at the heading stage may have a great effect on 2-AP biosynthesis in fragrant rice. Among the studied fragrant rice varieties, it was shown in Meixiangzhan-2, Xiangyaxiangzhan, Ruanhuayou-134 and Yunjingyou that exogenous application of Zn regulated the enzymes involved in 2-AP biosynthesis, including PDH, P5CS and DAO, and increased the contents of precursors, such as proline and P5C. Moreover, Zn application upregulated the net photosynthetic rate during the grain filling stage and increased the grain yield of fragrant rice. The Zn content in grains also increased due to foliar application of Zn. Considering that the highest yield and 2-AP content were recorded with the Zn2 treatment for all fragrant rice genotypes, 1.00 g L−1 can be recommended as the best foliar application concentration. To delve more deeply into the mechanism of 2-AP formation under the influence of Zn application, further work should be done at the molecular level.
Materials and Methods
Plant materials and growing conditions
Seeds of four fragrant rice varieties, ‘Meixiangzhan-2’, ‘Xiangyaxiangzhan’, ‘Ruanhuayou-134’, and ‘Yunjingyou’, which are widely grown fragrant rice varieties in Guangdong Province, China, were provided by the College of Agriculture, South China Agricultural University, Guangzhou China, and used in the present study. A field experiment during 2018 was conducted at the Experimental Research Farm, College of Agriculture, South China Agricultural University, Zengcheng (23°16′N, 113°22′E and 11 m above mean sea level), China. The experimental soil was sandy loam consisting of 15.91 g/kg organic matter, 1.34 g/kg total nitrogen, 55.16 mg/kg available nitrogen, 1.40 g/kg total phosphorus, 18.11 mg/kg available phosphorus, 13.22 mg/kg total potassium, and 128.06 mg/kg available potassium, with a soil pH of 6.70. The experimental site has a subtropical monsoon climate. The temperatures during the experiment are shown in Fig. 6. Before sowing, the seeds were soaked in water for 24 h, germinated in manually climate-controlled boxes for another 12 h, and then shade-dried, followed by sowing in polyvinyl chloride trays for nursery cultivation. Then, 20-day-old seedlings were transplanted to fields at a planting distance of 30 × 12 cm2.

Daily maximum and minimum temperatures during the experiment
Treatments and plant sampling
The four Zn treatments were set as follows: overhead sprinkling with 0, 0.50, 1.00 and 2.00 g·L−1 zinc chloride at the heading stage; these treatments were named CK, Zn1, Zn2 and Zn3, respectively. The treatments were arranged in a randomized complete block design (RCBD) in triplicate with a net plot size of 16 m2 (4 m·4 m). A total of 1.20 litres of the corresponding liquid was applied to each plot. Fresh grains were separated and collected from the rice plants after 15 days at the heading stage, washed with double-distilled water and stored at −80 °C for physio-biochemical analysis. At maturity, fresh grains were separated and collected from the rice plants and stored at −80 °C for 2-AP determination.
Determination of the 2-acetyl-1-pyrroline (2-AP) content in grains
Approximately 0.5 g of fresh grains were homogenized in 5 mL of 60% ethanol and treated for 4 h in an oscillator (HZS-H, China) at 200 oscillations per minute. The absorbance after the reaction was read at 645 nm, and the 2-AP amount was expressed as µg g−1. The 2-AP concentration was determined by the synchronization distillation and extraction (SDE) method combined with a GCMS-QP 2010 Plus (Shimadzu Corporation, Japan) according to Mo26, and the 2-AP contents were expressed as ug kg−1.
Determination of the Δ1-pyrroline, proline, pyrroline-5-carboxylic acid (P5C), γ-aminobutyric acid (GABA) and methylglyoxal contents in grains
The Δ1-pyrroline content in grains was estimated by the method of Hill27. The amount of △1-pyrroline in reaction mixtures containing 1, 4-diaminobutane was determined immediately and after 30 min at room temperature. Proline contents in grains were estimated according to the method of Bates28 using ninhydrin, and the absorbance was read at 520 nm. The P5C concentration was estimated following the method of Wu29. The mixture contained 0.2 ml of enzyme extraction supernatant, 0.5 ml of 10% trichloroacetic acid (TCA) and 0.2 ml of 40 mM 2-aminobenzaldehyde. After the reaction, the absorbance was read at 440 nm. The GABA content was measured according to the methods described by Zhao30. Grains of approximately 0.5 g were homogenized in 5 ml of 60% ethanol and treated for 4 h in an oscillator (HZS-H, China) at 200 oscillations per minute. The absorbance after the reaction was read at 645 nm. The content of methylglyoxal was estimated by the methods described by Banu31. The reaction system was 1 ml, with 250 L 7.2 mM 1, 2-diaminobenzene, 100 L 5 M perchloric acid, and 650 μL extract. The absorbance was determined at 336 nm.
Determination of proline dehydrogenase (PDH), △1-pyrroline-5-carboxylic acid synthetase (P5CS), ornithine aminotransferase (OAT), diamine oxidase (DAO) and betaine aldehyde dehydrogenase (BADH) activities in grains
The PDH activity was assayed by following the methods of Ncube32. Following the reaction, the absorbance was read at 440 nm, and the activity was calculated using a molar extinction coefficient. The activity of P5CS was estimated according to the methods described by Zhang33. The reaction mixture included 50 mM Tris-HCL buffer, 20.0 mM MgCl2, 50 mM sodium glutamate, 10 mM ATP, 100 mM hydroxamate-HCL and 0.5 ml of enzyme extract. OAT activity was measured according to the methods of Chen34. The absorbance of the supernatant fraction was read at 440 nm, and the activity was calculated by the extinction coefficient 2.68 mM−1 cm−1. The DAO activity was assayed using the methods described by Su35. Reaction solutions (2.9 mL) contained 2.0 mL of 70 mmol/L sodium phosphate buffer (pH 6.5), 0.5 mL of crude enzyme extracts, 0.1 mL of horseradish peroxidase (250 U/mL), and 0.2 mL of 4-aminoantipyrine/N,N-dimethylaniline. The betaine aldehyde dehydrogenase 2 (BADH2) enzyme activity was measured using a Plant PRODH ELISA Kit (Mlbio, Shanghai, China) according to the manufacturer’s protocol. The absorbance was measured using a BioTek Epoch spectrophotometer (BioTek, Vermont, USA) at 450 nm.
Determination of photosynthesis
At 15 days after the heading stage, a portable photosynthesis system (LI-6400, LI-COR, USA) was used to determine the net photosynthetic rate from 09:00–10:30 a.m. with the following adjustments: photosynthetically active radiation at the leaf surface was 1100–1200 μmol m−2 s−1, the ambient CO2 concentration was 385.0–400.0 μmol mol−1, and the air temperature was 31.5 ± 0.5 °C with 60–80% RH.
Determination of yield and yield-related traits
At the maturity stage, the rice grains were harvested from the five-unit sampling areas (1.75 m2) in each plot and then threshed by machine. The harvested grains were sun-dried and weighed to determine the grain yield. Twenty hills of rice from different locations in each plot were sampled to estimate the average effective panicle number per hill. Then, six representative hills of plants were taken to estimate the yield-related traits.
Determination of Zn content in grains
Approximately 2.000 g of the crushed sample was weighed and placed into a digestive tube with a concentrated nitric acid and perchloric acid mixture in a 4:1 ratio. The mixture was shaken well and left in a fume hood for cold digestion overnight. The next day, the sample was heated and digested in a digestive furnace. The procedure of heat digestion was as follows: 60 min digestion at 60°C, 60 min digestion at 100°C, 100 min digestion at 180°C, 180 min digestion at 195°C. If the liquid became brown-black during digestion, a small amount of nitric acid was added after cooling, and then the digestive liquid was heated until it was colourless or slightly transparent after cooling. The digestive tube was washed with ultra-pure water and filtered through medium-speed qualitative filter paper. The volume of the liquid was set to 25 ml, and the blank was made at the same time. The filtrate was determined by atomic absorption spectrophotometry, and then the zinc content in the sample was calculated based on a standard curve.
Statistical analysis
Data were analysed with Statistix 8.1 (Analytical Software, Tallahassee, FL, USA), while differences among means were separated by using the least significant difference (LSD) test at the 5% probability level. Graphs were drawn with Sigma Plot 14.0 (Systat Software Inc., California, USA).
References
Giraud, G. The World Market of Fragrant Rice, Main Issues and Perspectives. International Food & Agribusiness Management Review 16, 1–20 (2013).
Ren, Y. et al. Irrigation and Nitrogen Management Practices Affect Grain Yield and 2-Acetyl-1-Pyrroline Content in Aromatic Rice. Appl Ecol Env Res 15, 1447–1460 (2017).
Bradbury, L. M. T., Gillies, S. A., Brushett, D. J., Waters, D. L. E. & Henry, R. J. Inactivation of an aminoaldehyde dehydrogenase is responsible for fragrance in rice. (2008).
Li, M. et al. Manganese-induced regulations in growth, yield formation, quality characters, rice aroma and enzyme involved in 2-acetyl-1-pyrroline biosynthesis in fragrant rice. Plant Physiol BiocH 103, 167–175 (2016).
Mo, Z. et al. Silicon fertilization modulates 2-acetyl-1-pyrroline content, yield formation and grain quality of aromatic rice. J Cereal Sci 75, 17–24 (2017).
Silva, D. O. M. D. & Santos, C. A. F. Adaptability and Stability Parameters of Iron and Zinc Concentrations and Grain Yield in Cowpea Lines in the Brazilian Semiarid Region. Crop Sci 57, 2922 (2017).
Prasad, A. S. Impact of the discovery of human zinc deficiency on health. J Trace Elem Med Bio 28, 357–363 (2014).
Adamo, A.M. et al. Early Developmental Marginal Zinc Deficiency Affects Neurogenesis Decreasing Neuronal Number and Altering Neuronal Specification in the Adult Rat Brain. Front Cell Neurosci 13 (2019).
Fu, C. et al. Morphology, photosynthesis, and internal structure alterations in field apple leaves under hidden and acute zinc deficiency. SCI Hortic-Amsterdam 193, 47–54 (2015).
Han, W., Huang, L. & Owojori, O. J. Foliar application of zinc alleviates the heat stress of pakchoi (Brassica chinensis L.). J Plant NutR (2019).
Mo, Z. et al. Supplementation of 2-Ap, Zn and La Improves 2-Acetyl-1-Pyrroline Concentrations in Detached Aromatic Rice Panicles In Vitro. Plos One 11, e149523 (2016).
Yoshihashi, T. Quantitative analysis on 2-acetyl-1-pyrroline of an aromatic rice by stable isotope dilution method and model studies on its formation during cooking = Analyse quantitative de 2-acétyl-1-pyrroline de riz aromatique par une méthode de dilution d’isotope. J Food Sci (2002).
Yoshihashi, T., Huong, N. & Inatomi, H. Precursors of 2-acetyl-1-pyrroline, a potent flavor compound of an aromatic rice variety. J Agr Food Chem 50, 2001–2004 (2002).
Chen, S. et al. Badh2, Encoding Betaine Aldehyde Dehydrogenase, Inhibits the Biosynthesis of 2-Acetyl-1-Pyrroline, a Major Component in Rice Fragrance. The Plant Cell 20, 1850–1861 (2008).
Mo, Z. et al. Shading during the grain filling period increases 2-acetyl-1-pyrroline content in fragrant rice. Rice 8 (2015).
Poonlaphdecha, J. et al. Biosynthesis of 2-acetyl-1-pyrroline in rice calli cultures: Demonstration of 1-pyrroline as a limiting substrate. Food Chem 197, 965–971 (2016).
Bao, G. et al. Molecular basis for increased 2-acetyl-1-pyrroline contents under alternate wetting and drying (AWD) conditions in fragrant rice. Plant Physiol Bioch 133, 149–157 (2018).
Deng, Q. Q. et al. Mild Drought in Interaction with Additional Nitrogen Dose at Grain Filling Stage Modulates 2-Acetyl-1-Pyrroline Biosynthesis and Grain Yield in Fragrant Rice. Appl Ecol Env Res 16, 7741–7758 (2018).
Szabados, L. & Savoure, A. Proline: a multifunctional amino acid. Trends Plant Sci 15, 89–97 (2010).
Wakte, K. et al. Thirty-three years of 2-acetyl-1-pyrroline, a principal basmati aroma compound in scented rice (Oryza sativa L.): a status review. J Sci Food Agr 97, 384–395 (2017).
Cao, X. et al. Mixed-nitrogen nutrition-mediated enhancement of drought tolerance of rice seedlings associated with photosynthesis, hormone balance and carbohydrate partitioning. Plant Growth Regul 84, 451–465 (2018).
Tang, S. et al. Exogenous spermidine enhances the photosynthetic and antioxidant capacity of rice under heat stress during early grain-filling period. Funct Plant Biol 45, 911–921 (2018).
Tavallali, V. Interactive effects of zinc and boron on growth, photosynthesis, and water relations in pistachio. J Plant Nutr 40, 0–0 (2017).
Tavallali, V. et al. Zinc influence and salt stress on photosynthesis, water relations, and carbonic anhydrase activity in pistachio. Sci Hortic-Amsterdam 123, 272–279 (2009).
Ohki, K. Effect of Zinc Nutrition on Photosynthesis and Carbonic Anhydrase Activity in Cotton. Physiol Plantarum 38, 300–304 (2010).
Mo, Z. et al. Nitrogen application at the booting stage affects 2-acetyl-1-pyrroline, proline, and total nitrogen contents in aromatic rice. Chil J Agr Res 78, 165–172 (2018).
Hill, J. M. The inactivation of pea-seedling diamine oxidase by peroxidase and 1, 5-diaminopentane. Biochem J 104, 1048 (1967).
Bates, L. S., Waldren, R. P. & Teare, I. D. Rapid determination of free proline for water-stress studies. Plant & Soil 39, 205–207 (1973).
Wu, M. L., Chou, K. L., Wu, C. R., Chen, J. K. & Huang, T. C. Characterization and the possible formation mechanism of 2-acetyl-1-pyrroline in aromatic vegetable soybean (Glycine max L.). J Food Sci 74, S192–S197 (2009).
Zhao, D. W. et al. Determination of The γ-aminobutyric Acid in Barley. Journal of Triticeae Crops (2009).
Banu, M. N. A. et al. Proline and glycinebetaine ameliorated NaCl stress via scavenging of hydrogen peroxide and methylglyoxal but not superoxide or nitric oxide in tobacco cultured cells. Biosci Biotechnol Biochem 74, 2043–2049 (2010).
Ncube, B., Finnie, J. F. & Van, S. J. Dissecting the stress metabolic alterations in in vitro Cyrtanthus regenerants. Plant Physiology & Biochemistry 65, 102–110 (2013).
Zhang, C. S., Lu, Q. & Verma, D. P. Removal of feedback inhibition of delta 1-pyrroline-5-carboxylate synthetase, a bifunctional enzyme catalyzing the first two steps of proline biosynthesis in plants. J Biol Chem 270, 20491–20496 (1995).
Chen, C. T., Chen, L., Lin, C. C. & Kao, C. H. Regulation of proline accumulation in detached rice leaves exposed to excess copper. Plant Sci 70, 43–48 (1990).
Su, G., An, Z., Zhang, W. & Liu, Y. Light promotes the synthesis of lignin through the production of H 2 O 2 mediated by diamine oxidases in soybean hypocotyls. J Plant Physiol 162, 1297–1303 (2005).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (31971843), National Key R&D Program of China (2016YFD0700301), the Technology System of Modern Agricultural Industry in Guangdong (2017 LM1098) and the World Bank Loan Agricultural Pollution Control Project in Guangdong (0724-1510A08N3684).
Author information
Authors and Affiliations
Contributions
Haowen Luo and Bin Du designed the research. Longxin He, Lian Hu and Jing He performed the experiments and collected the data. Haowen Luo analysed the data and wrote the manuscript. Xiangru Tang and Shenggang Pan provided guidance during the experiment. All authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Luo, H., Du, B., He, L. et al. Exogenous application of zinc (Zn) at the heading stage regulates 2-acetyl-1-pyrroline (2-AP) biosynthesis in different fragrant rice genotypes. Sci Rep 9, 19513 (2019). https://doi.org/10.1038/s41598-019-56159-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-56159-7
- Springer Nature Limited