
Abstract
The ability to absorb water from the soil is likely to be a primary determinant of plant growth in water-limited ecosystems. Vertical root biomass distributions have long been used to understand how plants absorb water. However, a growing body of evidence suggests that root water uptake is more complicated. Some roots absorb water quickly, others slowly (i.e., root activity). Some species move their active roots through the soil more quickly than others (i.e., root plasticity). Here, a framework for understanding how vertical root distributions, root activity, and root plasticity affect plant growth and coexistence is developed. It is suggested that plants with a large transpiration demand use a deep and dynamic rooting strategy to provide a large, stable soil water supply. Plants with a smaller transpiration demand use a shallow and pre-emptive strategy to intercept enough water to produce herbaceous aboveground growth. The root strategy that best matches water availability at a site will have a competitive advantage. Tracer experiments can describe these rooting strategies and ecohydrologic models can predict their effects on water uptake over time. This framework provides a new way of understanding and measuring plant water uptake that can be expected to improve predictions of plant growth on the landscape and in response to changing climate conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Root biomass is not the same as water uptake
Vertical root distributions are widely thought to determine how much water a species can absorb, which species can coexist, and how much water reaches the atmosphere and aquifer in water-limited systems [1,2,3]. For example, Walter’s two-layer hypothesis suggests that shallow roots provide grasses pre-emptive and superior access to water as it enters the soil while deep roots allow woody plants to avoid competition with shallow-rooted grasses [4, 5]. This hypothesis has been popular for over 100 years, but a growing body of evidence suggests that by focusing on vertical root distributions and ignoring variation in root activity (i.e., the rate of water uptake per root length), root plasticity (i.e., the rate of change in active root distributions), and soil water flow, the two-layer hypothesis has oversimplified and potentially misguided our understanding of water uptake (Text box) [6,7,8,9].
Vertical root distributions Root biomass as a function of soil depth is most often measured by sieving and weighing or by image analyses in rhizotrons. These biomass measurements provide a direct measure of root biomass but an indirect measure of water uptake. In contrast, natural abundance stable isotope techniques and tracer techniques provide more direct measures of vertical water uptake, but more indirect measures of root biomass. |
Fine roots It has long been recognized that large, suberized roots do not actively absorb water from the soil and so measurements of non-suberized roots or roots < 2mm diameter are used to infer the distribution of active roots. |
Active roots Roots that actively absorb water or nutrients into cell membranes. Thin roots, roots with low water potentials, and roots with more or more active membrane transport proteins (e.g., aquaporins) will be more active (i.e., transport water more quickly into root cells). |
Root activity The rate of resource (i.e., water) uptake. Active roots must be located in ‘wet’ soils to absorb water. Soil water will generally flow into roots when the water potentials in the roots are lower than the water potentials in the soil. |
Root plasticity The rate of change in active root distributions. Species can change their active root distributions by growing new roots, thin roots, highly active roots, or by changing root-fungal interactions. Some species can make these changes quickly over large areas (plastic root responses) while other species maintain consistent vertical active root distributions (static root responses). |
A central problem for the two-layer hypothesis, and our understanding of water uptake in general, is that root biomass is not the same as water uptake [6, 10, 11]. Some roots can absorb water more quickly than others (i.e., are more active) [12, 13]. Root hairs, root thickness, aquaporin abundance and activity and mycorrhizal colonization all affect the rate of water uptake [14,15,16]. Further, some species can quickly change their active root profiles in response to water availability (i.e., root plasticity), while other species have a more static rooting strategy [7, 17, 18]. Water uptake, therefore, is a function not just of root location, but also of root activity and root plasticity [19,20,21].
In addition to root location, activity, and plasticity, it is also important to know when and where water is available in the soil because even active roots cannot absorb water from dry soils [6, 22]. Fortunately, computational models based on physical processes are quite good at predicting water flow through soils and into plant roots [22, 23]. For example, soil water flow models can predict exactly how shallow water will be when precipitation events are small, soil texture is fine, evapotranspiration is fast, and roots are shallow [18, 22, 23]. These models are used extensively by hydrologists and environmental engineers to simulate runoff, aquifer recharge, salinity, and pollution plume dynamics [24,25,26,27].
Validation of soil water flow model predictions has revealed that root plasticity is important at the ecosystem level. For example, models that assume that plant roots ‘follow’ water as it percolates through the soil outperform models that assume that roots maintain a static distribution in the soil [18, 28]. It is likely that this root plasticity varies phylogenetically or as a suite of plant traits [8, 29]. Unfortunately, little is known about variation in root plasticity among species [20, 30, 31].
To summarize, active roots must be present in wet soils to absorb water. This seemingly simple concept has been difficult to demonstrate because it is difficult to measure water uptake across the root systems of different species in the field [2, 9, 32]. Addressing this knowledge gap can be expected to improve predictions of plant growth, coexistence, and response to climate change [31, 33, 34]. In short, it should not be surprising that the amount of water a root system can absorb could predict plant abundance in water-limited systems, but until recently, this process has been difficult to measure and understand [2, 9, 32].
2 An emerging experimental approach
Tracer-informed ecohydrologic modeling (i.e., tracer/modeling) approaches are providing new insight into root location, activity, and plasticity [9, 35,36,37]. Some of the most detailed of these approaches injects hydrologic tracers to different soil depths in different replicate plots [38,39,40]. By measuring tracer concentration in plant tissues or transpired water, it is possible to describe the distribution of active roots of different species [41, 42]. Injections can be repeated over time or space to describe root plasticity [17]. Though more difficult to perform than natural abundance stable isotope techniques, tracer techniques provide more detailed, deeper, and robust descriptions of species water uptake [32, 43, 44]. The level of detail provided by tracer experiments may be necessary because small differences in root water uptake can have biologically important effects on plant growth [9, 21, 45].
Both a strength and weakness of tracer approaches is that they measure water absorbed from a certain soil depth regardless of how that water was absorbed. Plants can manipulate water uptake by producing more roots, finer roots, roots with mycorrhizal symbionts, roots with more or more active aquaporins, by producing roots that outcompete roots of adjacent species, or by hydraulically redistributing water. Tracer uptake describes the amount of water absorbed from different depths, but does not distinguish these different pathways.
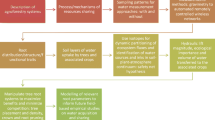
Tracer injections describe water uptake at a point in time. The amount of water that flows into different root systems between tracer injections can be estimated using soil water flow models [46, 47]. These models simulate how water moves over land, through the soil profile, or to the atmosphere via evaporation or root uptake. These processes are driven by precipitation, available energy for evaporation, resistance to evaporation (stomatal and aerodynamic), matric and gravitational forces on water in soil and the presence of plant roots [27, 48, 49]. These models typically use energy-balance approaches such as the Penman–Monteith or Hargreaves equations to simulate heat and water vapor transfers and they solve Richard’s equation to explain water flow through soils [23, 27, 48, 50]. These models can be used to simulate water flow into roots between injection dates, or for different environmental conditions (e.g., across the landscape or over time; Fig. 1) [51]. Root water uptake can then be converted to biomass production to estimate the growth of different plant species as a function of their root location, activity, and plasticity and their water use efficiency [22, 52, 53].

Water matching predicts grass and tree cover across a range of conditions in savannas of South Africa [51]. The response surfaces show the amount of water that tree or grass root systems were estimated to absorb as a function of soil water availability and mean annual precipitation. Tracer experiments were used to describe active root distributions in four sites with different soil (clay or sand) and climate [~ 350 mm or 700 mm mean annual precipitation (MAP)] conditions. Water uptake by active tree and grass root systems between sampling events was estimated using the soil water flow model Hydrus 1D. The mean depth of active tree roots (~ 30 cm) was slightly deeper than the mean depth of active grass roots (~ 20 cm). Slightly deeper roots were estimated to absorb more water than shallow roots under most conditions. There were two somewhat counterintuitive hydrologic ‘windows’ where shallow grass roots were expected to absorb more water than deeper tree roots. In dry conditions (MAP < 350 mm) with deep percolation, shallow roots had a water uptake advantage that predicted the observed increase in grass abundance (lower left corner). In wet conditions (MAP > 500 mm) with shallow water percolation (< 250 mm) grasses had an advantage (right edge). Water uptake predictions made using this tracer/modeling approach predicted tree and grass abundance on the landscape (see inserts) [51].
In the handful of sites where it has been tested, the tracer/modeling approach has predicted species water uptake and abundance on the landscape. In a shrub-steppe site in North America, the tracer/modeling approach predicted landscape abundances (R2 = 0.90) of five species that represented 84% of plant cover [54]. In that site, larger plants demonstrated deeper and more dynamic rooting patterns that absorbed more water, and these plants were more abundant on the landscape. In sub-tropical savannas of South Africa, the tracer/modeling approach similarly found that deep roots extracted more water than shallow roots across a range of abiotic conditions [51]. The extent of this ‘deep root advantage’ predicted tree cover on the landscape (Fig. 1). In both studies, larger plants were found to have deeper and more dynamic rooting distributions, for example absorbing the most water at 20 cm one month and at 60 cm the next month [51, 54]. The fact that plant landscape abundance could be predicted from estimates of water uptake alone suggests that water uptake is a primary determinant of plant growth upon which other factors such as fire, herbivory, life history, water use efficiency, etc. act. An important aspect of the tracer/modeling approach is that model estimates of soil water content can be validated using observed soil moisture.
2.1 Water matching: a new framework for understanding water uptake
Here a framework for understanding and measuring water uptake that incorporates vertical root distributions, root activity, root plasticity, and soil water flow is proposed. In this framework, total transpiration demand is the ultimate cause of root strategies in water-limited ecosystems [55]. Niche partitioning resulting from these different strategies is the proximate cause of species growth and coexistence [56]. Species with large transpiration demands must access large soil water pools [21, 57]. Deep roots provide access to a wide range of soil depths and, therefore, a potentially large soil water pool (Fig. 2). Further, because they are large, deep roots are likely to have stored carbon reserves that allow the production of ‘flushes’ of fine roots that forage for water pulses [58,59,60]. These flushes can occur vertically or horizontally [61, 62]. These deep and dynamic root systems come with large construction and maintenance costs [63]. For example, larger plants are likely to be taller with large water potential gradients across long, costly transpiration systems. These systems are more likely to realize large repair and replacement costs due to cavitation [58,59,60,61,62]Large transpiration demand, therefore, induces a large investment/large return strategy [8, 64, 65]. This strategy can be described as deep and dynamic.

Larger and dominant plants have deeper roots than smaller plants and sub-dominant plants. The mean depth of tracer uptake by 28 tree, grass, and forb species (points) sampled in savannas of Kruger Park, South Africa versus plant height (TRY database) [51]. The positive slopes of the curves indicate that taller plants absorb deeper water than shorter plants. The higher grey curve indicates that dominant plants absorb deeper water than sub-dominant plants (blue points and curve)
Plants with small transpiration demands tend to be small and produce herbaceous tissues that can be produced quickly from relatively small amounts of water [4, 34]. These plants can rapidly cover the ground and reproduce using smaller or more ephemeral pools of water [34, 66]. Shallow roots allow these species to intercept water as it enters the soil [4, 66]. This interception strategy often does not provide the most water (Fig. 2), but it provides enough water to allow growth and reproduction of small, fast-growing plants [51].
These strategies can be self-reinforcing. For deep-rooted species, shallow roots may result in a low return on investment due to competition from dense root mats of shallow-rooted species [67]. For shallow-rooted species, deep roots will come at the cost of aboveground growth and competitive ability with other shallow-rooted plants.
These competing strategies suggest that root activity and plasticity are especially important for larger and woodier species. This perspective explains that root biomass measurements may have misled our understanding of root function because some roots absorb water more quickly than others [11, 14]. It also suggests that measures of root activity and plasticity will be necessary to understand water uptake over time and space, particularly for woody species [7, 18, 19].
Large transpiration demands for an individual plant can be caused by several factors [48, 68]. Large size, leaf area, stomatal density and poor stomatal control will increase water demand [22, 69]. Taller plants will have less aerodynamic resistance and greater heat loading, both increasing water demand [68,69,70].
Site conditions will determine whether shallow or deep roots have a water uptake advantage [71, 72]. Shallow-rooted species benefit from conditions preventing deep percolation, such as frequent small precipitation events, clay soils, and precipitation during high evapotranspiration demand (i.e., summer) [73,74,75]. In contrast, deep-rooted species benefit from conditions allowing deep percolation, such as large infrequent precipitation events, sandy soils, and winter precipitation. In other words, plants with root systems that best ‘match’ water availability in a system will be able to absorb the most water and grow more than other species (Fig. 2).
Interestingly, there is a feedback between root distributions and availability that can affect the shallow or deep root advantage [51]. For example, dense, shallow roots will result in shallow percolation and water availability which will benefit shallow-rooted species. Deep roots will allow deeper percolation and water availability, which will benefit deeper-rooted species. These positive ecohydrologic feedbacks should act as a mechanism of positive frequency dependence that results in patchy vegetation distributions – a common, but poorly-understood vegetation pattern [76, 77].
The water matching hypothesis differs from the long-standing two-layer hypothesis in several ways. The water matching hypothesis predicts that in contrast to shallow-rooted plants, deep-rooted plants (1) typically absorb more water, (2) will dominate in arid and semiarid conditions (Fig. 3), and (3) rely more on variation in root activity and plasticity. Further, the water matching hypothesis explains that pre-emption allows grass coexistence in arid areas and that an ecohydrologic ‘window’ allows grass dominance where water availability is shallow. In contrast, it is difficult to understand how woody plants would co-exist with grasses if grasses are more water use efficient and have pre-emptive and superior access to soil water, as suggested by the two-layer hypothesis [4].

The shrub to grass cover ratio reveals greater shrub cover in more arid conditions in the western US. Greater shrub cover in arid conditions in the south and west of the US is consistent with the ‘water matching’ hypothesis and inconsistent with the two-layer hypothesis. Red values indicate greater shrub than grass cover. Blue values indicate greater grass than shrub cover. Analysis of data from the Bureau of Land Management’s Assessment, Inventory and Monitoring Program (https://www.blm.gov.aim)
The water matching hyhpothesis is consistent with many existing hypotheses but provides important advantages [78]. The ‘two-pools’ model suggests deep roots provide maintenance water [79]. A complementary idea is that small resource pulses benefit smaller, fast-growing plants [80, 81]. Classic ecohydrologic theory suggests that water flow will vary with soil texture [5, 82] and precipitation event size [75, 83]. Several studies have described ‘optimal rooting’ strategies as essentially ‘as shallow as possible’ [84,85,86]. Others still have emphasized the role of phenology [87, 88]. The water matching hypothesis integrates these ecohydrologic concepts but provides (1) a new explanation for how and why root water uptake varies among different plant species and functional groups, and (2) provides a means of measuring these differences.
3 Concluding remarks
Many factors determine plant growth and landscape abundance. Fire, herbivory, disease, intrinsic growth rates, life history traits, plant soil feedbacks, human distrubance etc. can all affect plant growth. However, it is reasonable to expect that the ability of a plant to absorb water in water-limited ecosystems will play a foundational role on which these other factors will act. Despite its potential importance, water uptake by different species remains difficult to measure in the field. Better estimates of root water uptake over time can be expected to improve predictions of plant growth, coexistence, and response to climate change [3, 17]. The water matching hypothesis provides a framework for understanding how different plants absorb water and tracer/modeling approaches provide a means of testing this understanding. Where species abundances vary from predictions from the water matching hypothesis, it is likely that other factors such as herbivory, water use efficiency, and human disturbance override the effects of water uptake on plant abundance. Continued research in this direction can be expected to transform our understanding of water uptake from qualitative to quantitative and allow a better understanding of the various factors that determine plant growth and coexistence under both current and anticipated climate conditions.
Data availability
No datasets were generated or analysed during the current study.
Code availability
Not applicable.
References
Smithwick EAH, Lucash MS, McCormack ML, Sivandran G. Improving the representation of roots in terrestrial models. Ecol Modell. 2014;291:193–204.
Sprenger M, Allen ST. What ecohydrologic separation is and where we can go with it. Water Resour Res. 2020. https://doi.org/10.1029/2020WR027238.
Schreiner-McGraw AP, et al. Woody plant encroachment has a larger impact than climate change on dryland water budgets. Sci Rep. 2020;10:8112.
Ward D, Wiegand K, Getzin S. Walter’s two-layer hypothesis revisited: back to the roots! Oecologia. 2013;172:617–30.
Case MF, Nippert JB, Holdo RM, Staver AC. Root-niche separation between savanna trees and grasses is greater on sandier soils. J Ecol. 2020;108:2298–308.
Zhu W, et al. Matching root water uptake patterns to fine root and soil water distributions. Plant Soil. 2024;495:499–516.
Zhao Y, Wang L, Chun KP, Ziegler AD, Evaristo J. Dynamic hydrological niche segregation: How plants compete for water in a semi-arid ecosystem. J Hydrol (Amst). 2024;630:130677.
Illuminati A, Querejeta JI, Pías B, Escudero A, Matesanz S. Coordination between water uptake depth and the leaf economic spectrum in a Mediterranean shrubland. J Ecol. 2022;110:1844–56.
Orlowski N, et al. Challenges in studying water fluxes within the soil-plant-atmosphere continuum: a tracer-based perspective on pathways to progress. Sci Total Environ. 2023;881: 163510.
Benettin P, et al. Tracing and closing the water balance in a vegetated lysimeter. Water Resour Res. 2021. https://doi.org/10.1029/2020WR029049.
Freschet GT, et al. Root traits as drivers of plant and ecosystem functioning: current understanding, pitfalls and future research needs. New Phytol. 2021;232:1123–58.
Klimešová J, Herben T. The hidden half of the fine root differentiation in herbs: nonacquisitive belowground organs determine fine-root traits. Oikos. 2023. https://doi.org/10.1111/oik.08794.
Ahmed MA, et al. Root type matters: measurement of water uptake by seminal, crown, and lateral roots in maize. J Exp Bot. 2018;69:1199–206.
Gambetta GA, Knipfer T, Fricke W, McElrone AJ. Aquaporins and Root Water Uptake. In: Chaumont F, Tyerman SD, editors. Plant Aquaporins. Cham: Springer International Publishing; 2017. p. 133–53.
Quiroga G, et al. The arbuscular mycorrhizal symbiosis regulates aquaporins activity and improves root cell water permeability in maize plants subjected to water stress. Plant Cell Environ. 2019;42:2274–90.
Kakouridis A, et al. Routes to roots: direct evidence of water transport by arbuscular mycorrhizal fungi to host plants. New Phytol. 2022;236:210–21.
Kulmatiski A, Beard KH. Woody plant encroachment facilitated by increased precipitation intensity. Nat Clim Chang. 2013;3:833–7.
Cai G, Vanderborght J, Couvreur V, Mboh CM, Vereecken H. Parameterization of root water uptake models considering dynamic root distributions and water uptake compensation. Vadose Zone J. 2018;17: 160125.
Nehemy MF, et al. Tree water deficit and dynamic source water partitioning. Hydrol Process. 2021;35: e14004.
Kühnhammer K, et al. Investigating the root plasticity response of Centaurea jacea to soil water availability changes from isotopic analysis. New Phytol. 2020;226:98–110.
Guderle M, et al. Dynamic niche partitioning in root water uptake facilitates efficient water use in more diverse grassland plant communities. Funct Ecol. 2018;32:214–27.
Palmquist KA, Bradford JB, Martyn TE, Schlaepfer DR, Lauenroth WK. STEPWAT 2: an individual-based model for exploring the impact of climate and disturbance on dryland plant communities. Ecosphere. 2018;9: e02394.
Šimůnek J, van Genuchten MTh, Šejna M. Recent developments and applications of the HYDRUS computer software packages. Vadose Zone J. 2016. https://doi.org/10.2136/vzj2016.04.0033.
Miguez-Macho G, Fan Y. Spatiotemporal origin of soil water taken up by vegetation. Nature. 2021;598:624–8.
Karandish F, Šimůnek J. A comparison of the HYDRUS (2D/3D) and SALTMED models to investigate the influence of various water-saving irrigation strategies on the maize water footprint. Agric Water Manag. 2019;213:809–20.
Jarvis NJ. A review of non-equilibrium water flow and solute transport in soil macropores: Principles, controlling factors and consequences for water quality. Eur J Soil Sci. 2020;71:279–302.
Radcliffe, D. E. & Simunek, J. Soil Physics with HYDRUS: Modeling and Applications. (CRC Press, 2010).
Šimůnek J, Hopmans JW. Modeling compensated root water and nutrient uptake. Ecol Modell. 2009;220:505–21.
Zhou Y, Wigley BJ, Case MF, Coetsee C, Staver AC. Rooting depth as a key woody functional trait in savannas. New Phytol. 2020;227:1350–61.
Grellier S, et al. Water uptake plasticity of savanna trees in encroached grassland: small trees match the mature trees. Afr J Range Forage Sci. 2021;38:231–43.
Kulmatiski A, Holdrege MC, Chirvasa C, Beard KH. Root distributions predict shrub-steppe responses to precipitation intensity. Biogeosci Dis. 2023;2023:1–29.
Rasmussen CR, Kulmatiski A. Improving inferences from hydrological isotope techniques. Trends Plant Sci. 2021;26:206–9.
Berry RS, Kulmatiski A. A savanna response to precipitation intensity. PLoS ONE. 2017;12: e0175402.
Wigley BJ, Coetsee C, February EC, Dobelmann S, Higgins SI. Will trees or grasses profit from changing rainfall regimes in savannas. New Phytol. 2024. https://doi.org/10.1111/nph.19538.
Penna D, Geris J, Hopp L, Scandellari F. Water sources for root water uptake: Using stable isotopes of hydrogen and oxygen as a research tool in agricultural and agroforestry systems. Agric Ecosyst Environ. 2020;291: 106790.
Dawson TE, Pausch RC, Parker HM. The role of hydrogen and oxygen stable isotopes in understanding water movement along the soil-plant-atmospheric continuum. In: Griffiths H, Griffiths H, editors. stable isotopes. New York: Garland Science; 2020. p. 169–83.
Smith A, Tetzlaff D, Landgraf J, Dubbert M, Soulsby C. Modelling temporal variability of in situ soil water and vegetation isotopes reveals ecohydrological couplings in a riparian willow plot. Biogeosciences. 2022;19:2465–85.
Guo H, Zhao Y. Using isotopic labeling to investigate root water uptake in an alley cropping system within Taklimakan desert oasis. China Agroforestry Systems. 2021;95:907–18.
Uugulu S, Wanke H, Koeniger P. Determining the source water and active root depth of woody plants using a deuterium tracer at a Savannah site in northern Stampriet Basin Namibia. Hydrogeol J. 2023. https://doi.org/10.1007/s10040-023-02747-x.
Plamboeck AH, Grip H, Nygren U. A hydrological tracer study of water uptake depth in a Scots pine forest under two different water regimes. Oecologia. 1999;119:452–60.
Magh R-K, et al. Competition for water rather than facilitation in mixed beech-fir forests after drying-wetting cycle. J Hydrol (Amst). 2020;587:124944.
Beyer M, et al. A deuterium-based labeling technique for the investigation of rooting depths, water uptake dynamics and unsaturated zone water transport in semiarid environments. J Hydrol (Amst). 2016;533:627–43.
von Freyberg J, Allen ST, Grossiord C, Dawson TE. Plant and root-zone water isotopes are difficult to measure, explain, and predict: some practical recommendations for determining plant water sources. Methods Ecol Evol. 2020;11:1352–67.
Wang S, et al. The natural abundance of stable water isotopes method may overestimate deep-layer soil water use by trees. Hydrol Earth Syst Sci. 2023;27:123–37.
Kulmatiski A, Beard KH, Holdrege MC, February EC. Small differences in root distributions allow resource niche partitioning. Ecol Evol. 2020;10:9776–87.
Clément C, et al. Comparing the deep root growth and water uptake of intermediate wheatgrass (Kernza®) to alfalfa. Plant Soil. 2022;472:369–90.
Dubbert M, Werner C. Water fluxes mediated by vegetation: emerging isotopic insights at the soil and atmosphere interfaces. New Phytol. 2019;221:1754–63.
Parton WJ. Abiotic Section of ELM. In: I GS, editor. Grassland Simulation Model. New York: Springer, New York; 1978. p. 31–53.
Ranatunga K, Nation ER, Barratt DG. Review of soil water models and their applications in Australia. Environ Model Softw. 2008;23:1182–206.
Schlaepfer DR, Lauenroth WK, Bradford JB. Ecohydrological niche of sagebrush ecosystems. Ecohydrology. 2012;5:453–66.
Kulmatiski A, Beard KH. A modern two-layer hypothesis helps resolve the ‘savanna problem.’ Ecol Lett. 2022;25:1952–60.
Peters DPC. Plant species dominance at a grassland–shrubland ecotone: an individual-based gap dynamics model of herbaceous and woody species. Ecol Modell. 2002;152:5–32.
Reichmann LG, Sala OE, Peters DPC. Precipitation legacies in desert grassland primary production occur through previous-year tiller density. Ecology. 2013;94:435–43.
Kulmatiski A, Adler PB, Foley KM. Hydrologic niches explain species coexistence and abundance in a shrub–steppe system. J Ecol. 2020;108:998–1008.
Laughlin DC, et al. Root traits explain plant species distributions along climatic gradients yet challenge the nature of ecological trade-offs. Nat Ecol Evol. 2021;5:1123–34.
Herben T, et al. The shape of root systems in a mountain meadow: plastic responses or species-specific architectural blueprints? New Phytol. 2022;235:2223–36.
Zhu W, et al. Effects of stand age and structure on root distribution and root water uptake in fast-growing poplar plantations. J Hydrol (Amst). 2023;616:128831.
Kleine L, Tetzlaff D, Smith A, Wang H, Soulsby C. Using water stable isotopes to understand evaporation, moisture stress, and re-wetting in catchment forest and grassland soils of the summer drought of 2018. 2020. Hydrol Earth Syst Sci. https://doi.org/10.5194/hess-24-3737-2020.
Zwetsloot MJ, Bauerle TL. Repetitive seasonal drought causes substantial species-specific shifts in fine-root longevity and spatio-temporal production patterns in mature temperate forest trees. New Phytol. 2021;231:974–86.
Chen X, Eamus D, Hutley LB. Seasonal patterns of fine-root productivity and turnover in a tropical savanna of northern Australia. J Trop Ecol. 2004;20:221–4.
Holdo RM, Nippert JB. Linking resource- and disturbance-based models to explain tree-grass coexistence in savannas. New Phytol. 2022. https://doi.org/10.1111/nph.18648.
Kulmatiski A, Beard KH, Verweij RJT, February EC. A depth-controlled tracer technique measures vertical, horizontal and temporal patterns of water use by trees and grasses in a subtropical savanna. New Phytol. 2010;188:199–209.
Anderegg WRL, et al. Tree mortality predicted from drought-induced vascular damage. Nat Geosci. 2015;8:367–71.
Knighton J, Fricke E, Evaristo J, de Boer HJ, Wassen MJ. Phylogenetic underpinning of groundwater use by trees. Geophys Res Lett. 2021. https://doi.org/10.1029/2021GL093858.
Tumber-Dávila SJ, Schenk HJ, Du E, Jackson RB. Plant sizes and shapes above and belowground and their interactions with climate. New Phytol. 2022;235:1032–56.
February EC, Higgins SI, Bond WJ, Swemmer L. Influence of competition and rainfall manipulation on the growth responses of savanna trees and grasses. Ecology. 2013;94:1155–64.
Cabal C, Martínez-García R, de Castro Aguilar A, Valladares F, Pacala SW. The exploitative segregation of plant roots. Science. 2020;370:1197–9.
Campbell GS, Norman JM. An introduction to environmental biophysics. Germany: Springer Science Business Media; 2000.
Allen RG, Pereira LS, Raes D, Smith M. Crop evapotranspiration-Guidelines for computing crop water requirements-FAO Irrigation and drainage paper 56. Fao, Rome. 1998;300:D05109.
Kool D, et al. A review of approaches for evapotranspiration partitioning. Agric For Meteorol. 2014;184:56–70.
Langley JA, et al. Do trade-offs govern plant species’ responses to different global change treatments? Ecology. 2022. https://doi.org/10.1002/ecy.3626.
Renne RR, Bradford JB, Burke IC, Lauenroth WK. Soil texture and precipitation seasonality influence plant community structure in North American temperate shrub steppe. Ecology. 2019;100: e02824.
Jones SK, Collins SL, Blair JM, Smith MD, Knapp AK. Altered rainfall patterns increase forb abundance and richness in native tallgrass prairie. Sci Rep. 2016;6:20120.
Luo W, et al. Responses of a semiarid grassland to recurrent drought are linked to community functional composition. Ecology. 2022. https://doi.org/10.1002/ecy.3920.
Knapp AK, et al. Consequences of more extreme precipitation regimes for terrestrial ecosystems. Bioscience. 2008;58:811–21.
Staver AC, Asner GP, Rodriguez-Iturbe I, Levin SA, Smit IPJ. Spatial patterning among savanna trees in high-resolution, spatially extensive data. Proc Natl Acad Sci. 2019;116:10681–5.
Rodriguez-Iturbe I, Chen Z, Staver AC, Levin SA. Tree clusters in savannas result from islands of soil moisture. Proc Natl Acad Sci. 2019;116:6679–83.
Schenk HJ, Jackson RB. Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems. J Ecol. 2002. https://doi.org/10.1046/j.1365-2745.2002.00682.x.
Ryel RJ, Ivans CY, Peek MS, Leffler AJ. Functional differences in soil water pools: a new perspective on plant water use in water-limited ecosystems. In: L U, B W, M J, editors. Progress in botany. Berlin: Springer, Berlin Heidelberg; 2008. p. 397–422.
Schwinning S, Sala OE. Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia. 2004;141:211–20.
Sala OE, Golluscio RA, Lauenroth WK, Soriano A. Resource partitioning between shrubs and grasses in the Patagonian steppe. Oecologia. 1989;81:501–5.
Noy-Meir I. Desert ecosystems: environment and producers. Annu Rev Ecol Syst. 1973;4:25–51.
Holdrege MC, Kulmatiski A, Beard KH, Palmquist KA. Precipitation Intensification Increases Shrub Dominance in Arid, Not Mesic Ecosystems. Ecosystems. 2022. https://doi.org/10.1007/s10021-022-00778-1.
Schenk HJ. The shallowest possible water extraction profile: a null model for global root distributions. Vadose Zone Journal. 2008;7:1119–24.
Laio F, D’Odorico P, Ridolfi L. An analytical model to relate the vertical root distribution to climate and soil properties. Geophys Res Lett. 2006. https://doi.org/10.1029/2006GL027331.
Collins DBG, Bras RL. Plant rooting strategies in water-limited ecosystems. Water Resour Res. 2007. https://doi.org/10.1029/2006WR005541.
Xu X, Medvigy D, Rodriguez-Iturbe I. Relation between rainfall intensity and savanna tree abundance explained by water use strategies. Proc Natl Acad Sci. 2015;112:12992–6.
Scholes RJ, Walker BH. An African savanna: synthesis of the nylsvley study. Cambridge: Cambridge University Press; 1993.
Funding
This research was supported by the Utah Agricultural Experiment Station, Utah State University, and approved as journal paper number 9756.
Author information
Authors and Affiliations
Contributions
AK conceived and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Kulmatiski, A. Water matching: an explanation for plant growth and coexistence in water-limited systems. Discov. Soil 1, 2 (2024). https://doi.org/10.1007/s44378-024-00002-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s44378-024-00002-7