
Abstract
The global demand for animal-derived foods has led to a substantial expansion in ruminant production, which has raised concerns regarding methane emissions. To address these challenges, microalgal species that are nutritionally-rich and contain bioactive compounds in their biomass have been explored as attractive feed additives for ruminant livestock production. In this review, we discuss the different microalgal species used for this purpose in recent studies, and review the effects of microalgal feed supplements on ruminant growth, performance, health, and product quality, as well as their potential contributions in reducing methane emissions. We also examine the potential complexities of adopting microalgae as feed additives in the ruminant industry.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
The livestock sector plays a vital role in terms of safeguarding global food security and nutrition, and animal-derived products provide a significant proportion of protein intake (one-third) and calories (17%) worldwide, with a substantial contribution coming from ruminants (FAO 2018). Unfortunately, the ongoing rise in our global population is predicted to increase total food demand by 35% to 56% between the years 2010 and 2050 (van Dijk et al. 2021). The resulting escalation in demand for animal-derived foods will almost certainly trigger the expansion of ruminant livestock production, which, in turn, poses several environmental challenges, such as the potential for a consequent increase in methane emissions (Alexandratos et al. 2012; Odegard et al. 2014; Tilman et al. 2011; Yan et al. 2024). Given the increasing demand for animal products and a growing awareness of their nutritional value, along with the importance of environmental sustainability in livestock industries and the need to address potential health risks associated with animal production (for a review, see (Gilbert et al. 2021)), there is a dire need for the development of strategies to resolve these issues. Ideally, these solutions would simultaneously improve the competitiveness of ruminant production and mitigate adverse environmental impacts from the livestock sector. In this context, exploring alternative feed additives, such as microalgae, offers promising avenues to meet market demands while at the same time promoting environmental sustainability in livestock industries (Madeira et al. 2017).
The inclusion of microalgae in animal diets has gained widespread attention for aquaculture, as well as the production of poultry, monogastric mammals, and ruminants (El-Ghany 2020; Ma et al. 2024; Madeira et al. 2017; Martins et al. 2021). Microalgae are abundant in aquatic environments, possessing favorable characteristics such as rapid proliferation, widespread habtitats, and environmental adaptability, which make them easily accessible as natural food and feed sources for various aquatic organisms (Olabi et al. 2023; Udayan et al. 2021). In addition, certain microalgal species can rapidly synthesize and accumulate large amounts of high-quality nutritional and bioactive compounds, including lipids, proteins, carbohydrates, vitamins, carotenoids, and omega-3 fatty acids, and are thus attractive as multifaceted feed additives for animal production and health (Dineshbabu et al. 2019).
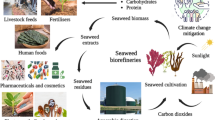
In this review, we discuss recent advances in the utilization of microalgal feed additives in ruminant livestock production (Fig. 1). We begin by considering the major microalgal species that have been assessed for their use as ruminant feed additives, along with their general nutritional compositions. We then discuss the influence of microalgal feed additives on the performance and health of ruminants, as well as their effects on meat and milk quality. Subsequently, we consider the influence of microalgal supplementation on ruminant methane emissions, and discuss the potential challenges and future perspectives of using microalgae as ruminant feed additives.

Graphical outline of effects of dietary microalgal supplementation on ruminant production and sustainability. The figure is generated in BioRender.
2 Representative microalga species used as ruminant feed additives
Both prokaryotic microalgae, such as Spirulina (also known as Arthrospira), and eukaryotic microalgae, including Chlorella (also known as Auxenochlorella), Scenedesmus, Schizochytrium (also known as Aurantiochytrium), Isochrysis, Nannochloropsis, Euglena, Micractinium, and Tetracystis, have been used as ruminant feed additives to date (Table 1). Due to the rapid growth and simple structure of microalgal cells, these species can accumulate extremely high amounts of one or more of the three major nutritional components (proteins, carbohydrates, and lipids) under optimized conditions (Table 1). Among the representative species, Spirulina spp. and Chlorella spp. are the most widely utilized as ruminant feed additives, owing to their remarkable nutritional composition and the relative ease with which they can be cultivated on large-scale, industrial levels. Typically, these species can accumulate 50-70% protein on a dry weight basis with an exceptional profile of essential amino acids for ruminant dietary requirements (Anvar et al. 2021; Bito et al. 2020; Gutiérrez-Salmeán et al. 2015).
The attractiveness of using microalgae as feed additives in ruminant diets also stems from their high contents of bioactive compounds in their biomass, including unsaturated fatty acids and essential vitamins, which play important roles in ruminant health and product quality. For example, Spirulina spp. and Chlorella spp. can produce high amounts of ω-6 unsaturated fatty acids (Gutiérrez-Salmeán et al. 2015) and essential vitamins (Bito et al. 2020), respectively, which can substantially enhance the health and well-being of ruminants by alleviating oxidative stress and reinforcing immunity (Ampofo et al. 2022). Nannochloropsis spp., on the other hand, can accumulate high levels of an omega-3 very long chain fatty acid termed eicosapentaenoic acid (EPA) under stressful conditions (Ma et al. 2014; Rebolloso-Fuentes et al. 2001), while Isochrysis galbana and Schizochytrium spp. can produce high amounts of a similar fatty acid termed docosahexaenoic acid (DHA) (Hien et al. 2022; Mohy El-Din 2019; Xu et al. 2021). Both EPA and DHA can enhance animal health and the nutritional value of animal products (Barta et al. 2021).
When using microalgae as ruminant feed supplements, it should be noted that microalgal biomass and the content of nutritional and bioactive components can be substantially affected by strains, culture conditions, and growth stages. Quality control of microalgal biomass is critical in this regard. For instance, the DHA content of the Aurantiochytrium sp. T66 strain can be increased to 52% of cell dry weight under optimized culture conditions, but it can be much lower under unfavorable conditions (Jakobsen et al. 2008). In addition to microalgal strain selection and cultural condition optimization, traditional mutagenesis and advanced metabolic engineering can effectively improve microalgal strains, such as increasing the contents of omega3-fatty acids or proteins, to meet various nutritional demands of ruminant growth and production (Grama et al. 2022; Trovão et al. 2022). Additionally, since only a very small proportion of microalgal species have been assessed for their value as ruminant feed additives to date, and the mechanisms driving their positive roles in ruminant production and nutrition are yet to be well-characterized, there is much room for further investment in this emerging field. For instance, other microalgal species with rapid growth rate and beneficial bioactive compounds could be evaluated, and omics technologies (e.g., genomics, transcriptomics, and metabolomics) with nutritional and physiological studies could help elucidate the specific mechanisms of how the microalgal supplementation influences rumen fermentation and nutrient metabolism of ruminants.
3 Influence of dietary microalgae supplementation on ruminant performance and health
3.1 Feed intake and body weight
The effects of microalgal feed additives on ruminant growth and performance have been reported previously (Table 2). Although the direct comparison of results across different studies is difficult due to variations in the microalgal species, amount of biomass, feeding frequency, and other factors used, the results, in general, demonstrated the promise of selected microalgal species as feed supplements. Among the tested species, Spirulina platensis, which contains a high amount of protein, has gained widespread recognition over the past decade. For example, the inclusion of S. platensis in daily steer diets increased feed intake and microbial protein production efficiency, but decreased digesta retention time (Panjaitan et al. 2015). Moreover, a linear correlation was observed between the supplementation of S. platensis-derived nitrogen and average daily gain (Costa et al. 2016). Furthermore, the incorporation of post-lipid-extraction S. platensis residue into feed diets showed a positive correlation with increased organic matter intake and digestibility (Drewery et al. 2014). Taken together, high-protein S. platensis appears to be a promising potential alternative supplement for cattle grazing low-protein forages, which could improve feed intake and the efficiency of nutrient utilization to promote animal growth and productivity (Costa et al. 2016; Drewery et al. 2014; Panjaitan et al. 2015).
Similarly, in the case of dairy cows, the inclusion of 200 g/day of S. platensis in the diet of Lithuanian black and white cows during their early lactation phase over a 90-day period led to improvements in body condition (8.5-11% fatter) (Kulpys et al. 2009). Moreover, the daily inclusion of S. platensis in the diets of Multiparous Finnish Ayrshire cows increased fiber and nitrogen digestibility (Lamminen et al. 2017); however, feed intake was not affected, which could be attributed to variations in the individual adaptability of cows to diets containing microalgal supplements and differences in the palatability of various doses of microalgae (Lamminen et al. 2017).
The benefits of Spirulina supplementation have also been reported in goats and lambs (Table 2). For instance, incorporating 10% (w/v) of S. platensis in daily diets over a six-week period led to an increase in the liveweight and body condition score of weaner lambs (Holman et al. 2012). Likewise, in simulated-drought basal diets, medium (100 mL) and high (200 mL) levels of 10% (w/v) of S. platensis supplementation per day over nine weeks resulted in an improvement in liveweight and average daily gain of White Suffolk- and Merino-sired lambs (Holman et al. 2014). In addition, the inclusion of 1 g S. platensis per 10 kg body weight in daily diets over 35 days not only improved live body weight and daily live weight gain, but also increased feed intake and feed conversion ratio in lambs (El-Sabagh et al. 2014).
Other microalgal species such as Chlorella spp. and Schizochytrium spp. have also been assessed in ruminant feed experiments (Table 2). While C. pyrenoidosa had positive effects in B. indicus steers (Costa et al. 2016), the incorporation of DHA-rich Aurantiochytrium limacinum did not show any significant influence on the body weight of ruminants (Carvalho et al. 2018; Meale et al. 2014). In addition to variations in the nutritional composition of microalgal species, this discrepancy could also be, at least in part, related to palatability and reduced feed intake (Carvalho et al. 2018). To ensure unbiased results when employing microalgae feed additives in ruminant production, it will likely be essential to evaluate the overall acceptability of the feed following the addition of microalgal species prior to feeding experiments.
3.2 Immunity and the reduction of oxidative stress
Ruminants with robust immune and antioxidant systems can reduce susceptibility to disease and enhance adaptability to environmental stressors (Ciliberti et al. 2022). This resilience is curial for maintaining growth and animal health. Microalgae are natural sources of antioxidants and bioactive compounds that help neutralize harmful free radicals in the body, reducing oxidative stress, and mitigating inflammatory effects (Mavrommatis et al. 2023), potentially improving ruminant health and well-being. Recent studies indicated that S. platensis supplementation may affect physiological parameters associated with immune and antioxidant systems of ruminants and improve their health and disease resistance. For instance, the supplementation of 1 g of S. platensis per 10 kg body weight into the daily diets of fattening lambs resulted in elevated levels of hemoglobin, total white blood cell count, serum globulin, and vitamin A (El-Sabagh et al. 2014). Similarly, diets including the addition of 2 g of S. platensis biomass per day led to an 8.9% increase in the amount of hemoglobin and a 13.1% rise in erythrocytes in milk cows (Šimkus et al. 2007). Additionally, the inclusion of 3% S. platensis on a dry matter basis in high-energy diets over 74 days increased the activity of superoxide dismutase, total antioxidant capacity and IgG concentration in the serum of Hu lambs (Liang et al. 2020). As of yet, however, very few studies have assessed the impact of microalgal species other than S. platensis on ruminant immune and antioxidant systems. While one study indicated that the inclusion of 170 g and 255 g of Schizochytrium sp. in the diet per day over a 60-day period had no significant effect on the growth or health of dairy cows (Liu et al. 2020), another suggested that the supplementation with 100 g or 200 g of Schizochytrium sp. per day over 49 days enhanced the total antioxidant capacity and concentration of glutathione peroxidase in the meat of beef cattle (Xu et al. 2021), indicating that individual differences in ruminants might affect the efficacy of microalgal feed additives.
Taken together, it is clear that at least certain microalgal feed additives can improve ruminant immunity and resistance to oxidative stress, and consequently ruminant health, but further studies with different microalgal species and various ruminants are required to expand our knowledge in this area. In addition, there is also a paucity of information regarding the associated mechanisms underlying the effects of microalgal supplementation on ruminants. Recent studies have indicated that a balanced and diverse gut microbiome contributes to a well-functioning immune and antioxidant system in ruminants (Cholewinska et al. 2020; Newbold et al. 2020). Since microalgal additives in feed may provide probiotic effects on rumen microbes, it is possible that they might function to restore microbiome composition, thus alleviating inflammation and enhancing oxidative resistance (Patel et al. 2021). An extensive analysis of the dynamic interactions between microalgal feed additives and rumen microbes could help address these questions and increase our understanding of how microalgal feed additives can enhance the performance and health of ruminants.
4 Influence of dietary microalgae supplementation on ruminant meat quality
Superior meat quality is characterized by taste, texture, and nutritional attributes, often warranting premium prices due to consumer preferences (Hathwar et al. 2012). As such, improving meat quality, and particularly its nutritional content, can substantially advance the profitability of ruminant production (Delgado-Pertíñez et al. 2021). Some microalgal species contain high levels of polyunsaturated fatty acids (PUFAs), vitamins, and minerals, and can be used to improve meat quality, especially PUFA contents (Table 3). Schizochytrium spp. stand out as a particularly intriguing microalgal species for improving meat nutritional attributes because of its high DHA content and rapid growth under heterotrophic culture in regular fermenters. Many studies have reported that feed diets supplemented with Schizochytrium spp. biomass led to enhanced nutritional quality in lamb muscle, characterized by a more favorable fatty acid profile and increased DHA and EPA levels (Carvalho et al. 2018; Díaz et al. 2017; Fan et al. 2019; Hopkins et al. 2014; Meale et al. 2014; Ponnampalam et al. 2016; Urrutia et al. 2016; Xu et al. 2021). In terms of other algal species, the inclusion of a dinoflagellate (Dinophyceae) in the diet of lambs also led to an increase in EPA and DHA levels in meats (Cooper et al. 2004; Elmore et al. 2005), and dietary supplementation with 4% Isochrysis sp. on a dry matter basis in daily diets increased the content of α-linolenic acid in lamb meat (De la Fuente-Vazquez et al. 2014).
Although the inclusion of certain microalgal species in diets could increase PUFA content in meat, this dietary supplementation can also lead to reduced ratings in terms of odor and flavor (Ponnampalam et al. 2016; Urrutia et al. 2016). One possible reason for this is that high amounts of unsaturated fatty acids in ruminant meats make them more susceptible to oxidation when exposed to air, impacting overall color and flavor (Lindahl et al. 2001; Wood et al. 2008). Hence, in practical applications, careful consideration will be required to prevent potential negative impacts on ruminant meat quality, such as adopting the use of microalgae species with favorable nutritional profiles, avoiding species with pronounced and undesirable odors, and implementing strict quality control for the flavor and taste of microalgae-based additives.
5 Influence of dietary microalgae supplementation on the yield and nutrition of ruminant milk
Ruminant milk is one of the most widely consumed beverages worldwide (Graulet 2014), and there is a growing interest in studying the influence of microalgal supplementation on dairy production (Table 4). Lithuanian Black-and-White cows fed forage supplemented with 2 g S. platensis per day over a 60-day period led to a noteworthy 7.6% or 136 kg increase in average milk production compared to a standard forage diet (Šimkus et al. 2007). Likewise, the inclusion of S. platensis additives in daily rations led to improved body condition and milk production in individual cows (Kulpys et al. 2009), and the supplementation with C. vulgaris increased the milk yield of lactating Damascus goats (Kholif et al. 2016).
Conversely, there have been instances where the dietary inclusion of algae did not have a significant impact on milk production. For example, basal diets supplemented with S. platensis or Schizochytrium spp. had no discernible effect on dry matter intake or milk yield in various studies (Bichi et al. 2013; Franklin et al. 1999; Klop et al. 2016; Lamminen et al. 2017; Liu et al. 2020; Moate et al. 2013; Papadopoulos et al. 2002; Vahmani et al. 2013). In contrast, diets supplemented with DHA-enriched Schizochytrium sp. resulted in a substantial 45% reduction in both dry matter intake and milk yield in dairy cows (Boeckaert et al. 2008). The disparities among these studies primarily stem from variations in the intake efficiency of feeds containing unpalatable microalgae. However, despite the negative impact on milk yield, Schizochytrium supplementation significantly increased the proportion of omega-3 fatty acids in ruminant milk (Bichi et al. 2013; Boeckaert et al. 2008; Franklin et al. 1999; Klop et al. 2016; Liu et al. 2020; Moate et al. 2013; Papadopoulos et al. 2002; Póti et al. 2015; Vahmani et al. 2013).
High levels of omega-3 fatty acids, such as DHA and EPA, in ruminant milk may provide potential benefits to improve human cardiovascular health, brain development, and immune function (Barta et al. 2021). Nevertheless, PUFAs are more susceptible to oxidation than saturated and monounsaturated fatty acids (Ponnampalam et al. 2024), and therefore increased PUFA content in milk may lead to a higher risk of lipid oxidation during storage and processing, which can negatively impact the flavor and aroma of dairy products (Ponnampalam et al. 2024). In this regard, when certain microalgal species are included in ruminant diets, it is essential to carry out extensive research in comprehensively evaluating the pros and cons, such as nutritional value, flavor, palatability, shelf life, and market acceptance of the milk and milk products.
Unlike milk yield and fatty acid composition, protein and lactose levels in ruminant milk are not significantly influenced by dietary microalgae in general (Bichi et al. 2013; Boeckaert et al. 2008; Klop et al. 2016; Lamminen et al. 2017; Liu et al. 2020; Póti et al. 2015; Vahmani et al. 2013), although several studies have reported an increase in protein (Kulpys et al. 2009; Papadopoulos et al. 2002; Šimkus et al. 2007) or lactose content (Kholif et al. 2016; Kulpys et al. 2009; Šimkus et al. 2007). Taken together, these findings suggest that the inclusion of protein-rich microalgae can enhance milk production without affecting feed intake in general, while the addition of DHA-rich microalgal biomass to feed can elevate the content of omega-3 fatty acids in milk, albeit with a negative effect on feed intake due to decreased palatability. Since palatability is an important issue, it may be necessary to refine the sensory properties of at least certain microalgae feed additives to ensure their effective application.
It has been suggested that the effect of microalgal feed additives on milk production may be determined predominantly by alterations in feed intake, digestion efficiency, and rumen fermentation, which are critical for ruminant growth (Kholif et al. 2016; Kulpys et al. 2009). In terms of milk composition, modifications might possibly stem from the influence of microalgal supplementation on rumen biohydrogenation and milk synthesis. During rumen biohydrogenation, microbes convert unsaturated fatty acids such as linoleic acid and linolenic acid to saturated fatty acids, which are then incorporated into milk production (Harvatine et al. 2009; Kholif et al. 2016). Supplementation with PUFA-rich microalgal biomass has been proposed to potentially impede the hydrogenation of unsaturated fatty acids in the rumen, but at the same time also provide more conjugated linoleic acid, EPA and DHA into milk production (Kholif et al. 2016; Lourenco et al. 2010). However, further research is needed to expand our understanding of rumen biohydrogenation, as well as the influence of microalgal supplementation on this intricate process, to unravel these mechanisms in full.
6 Influence of dietary microalgae supplementation on ruminant methane emissions
The livestock industry contributes a substantial proportion of greenhouse gas emissions, especially methane released during the digestive processes of ruminants (Yan et al. 2024). The potential value of dietary microalgae supplementation on the reduction of methane emissions have been studied using different microalgal species, mainly through in vitro assays (Table 5), and at least in certain cases the results have indicated a positive effect. For instance, adding 100 g of Euglena gracilis per kg of feed dry matter led to a noteworthy 9.1% reduction in methane production from non-lactating Holstein cows’ rumen fluid without adverse effects on the in vitro fermentation profile (Aemiro et al. 2016). Similarly, the incorporation of 5% Nannochloropsis oculate in Barki sheeps’ rumen fluid also reduced methane production (Gomaa et al. 2018). Moreover, diets containing post-lipid-extraction Scenedesmus sp. biomass significantly reduced methane production, while whole cells did not have the same effect (Tibbetts et al. 2017). Additionally, the inclusion of Chlorella spp. at 25% of the total incubated dry matter in the feed also resulted in a slight decrease in methane production (Sucu 2020).
Nevertheless, the effects of microalgal feed additives on the production of rumen-derived methane tend to vary with different microalgal strains. When the effects of Scenedesmus sp., C. vulgaris, Nannochloris bacillaris, Tetracystis sp., Micractinium reisseri, and Nannochloropsis granulate on methane production were assessed using in vitro assays, total methane production varied across the different microalgal species, with only Tetracystis sp. displaying a potential trend towards reducing the generation of methane (Anele et al. 2016). In other in vitro studies, the inclusion of C. vulgaris, Nannochloropsis limnetica, and Schizochytrium sp. did not reduce, and in some cases even increased, methane emissions (Kholif et al. 2016; Klop et al. 2016; Marrez et al. 2017; Ruiz-Gonzalez et al. 2018). Since most of these studies were carried out using in vitro experiments, it will be of interest to examine the value of microalgae in terms of decreasing methane emissions in vivo in ruminants.
It should be noted that, to date, the most effective dietary algae supplementation found to mitigate methane emissions is not microalgae, but the red macroalga Asparagopsis, primarily due to its natural synthesis of bromoform (Zhu et al. 2021). Bromoform can interact with vitamin B12 to influence the cobamide-dependent methyltransferase reaction, which is a pivotal step in methane production (Glasson et al. 2022; Thapa et al. 2020). Extensive research has confirmed the significant contribution of Asparagopsis to methane reduction in ruminants (Camer-Pesci et al. 2023). While the current microalgal studies have not yet identified species as promising as Asparagopsis, only a very limited number of the over 72,500 microalgae species have been investigated thus far (Grama et al. 2022). Given the shared evolution and growth habitats of microalgae and red macroalgae, it is certainly possible that microalgae with similar bromoform synthetic capabilities might be identified in the future. In addition, since the genetic engineering of certain microalgal species has been well-developed, the genetic reconstruction of bromoform synthesis in microalgae could also provide a potential means of contributing to methane reductions downstream. However, bromoform might be a suspected carcinogen (www.epa.gov) and further research is needed in this regard. Finally, the screening of microalgae-derived bioactive compounds for their ability to inhibit the activities of specific methanogenic archaea or methanogenic processes in the rumen could also open alternative avenues for decreasing ruminant-derived methane emissions through the use of microalgae-based supplements.
7 Challenges for adopting microalgae as feed additives in ruminant production
While microalgae as ruminant feed additives have garnered attention due to their unique nutritional value and growth characteristics, certain challenges still need to be addressed. For example, compared with other feed sources, microalgal biomass is very expensive. This is, at least in part, due to the fact that prevailing culture methods, especially autotrophic culture, have relatively low productivity (Dębowski et al. 2020), and harvesting and dewatering processes are energy-intensive and costly (Deepa et al. 2023). Method optimization of large-scale cultivation, harvesting and dewatering, and contamination control are needed to improve commercial viability while ensuring the quantity and quality of microalgal biomass. Moreover, some microalgal species, such as Schizochytrium and Chlorella, can grow well heterotrophically using well-developed large-scale fermentation processes (for a review, see (Chen et al. 2006)), which makes them promising species for the production of microalgal feed additives. Another challenge comprises obtaining approvals for the use of microalgae in food and feed. Currently, only limited microalgal species have been approved for food use, such as Chlorella, Spirulina, and Schizochytrium (Niccolai et al. 2019). Obtaining regulatory approvals for microalgal feed additives can be lengthy and costly, and requires strong support from research results. As such, it is currently more attractive to develop and test microalgal species with regulatory approvals in ruminant diets to mitigate this issue. Since the supplementation of microalgae in ruminant diets may result in reduced intake due to the unfavorable flavor of feed (Carvalho et al. 2018; Meale et al. 2014), further research will also be needed to improve the palatability of microalgal feed additives to ensure effective adoption. Finally, it will also be essential to comprehensively understand the effect of various microalgal species on ruminant nutrition and health, the potential interaction between microalgae and rumen microbes, and the underlying mechanisms driving their effects.
8 Conclusions and perspectives
The integration of microalgae-based feed additives holds great promise to promote the sustainability of livestock industries and improve ruminant production. The increasing recognition of the nutritional value from microalgae has sparked significant interest regarding their effects as dietary supplements for ruminants. Cumulative studies have demonstrated the unique value of microalgae as nutritional supplements, effectively meeting the dietary requisites of ruminants, improving their physiological performance, and enhancing the nutritional profile of ruminant-derived products. However, further research is needed to improve microalgal production, expand our understanding of the molecular mechanisms driving the beneficial effects of microalgal feed additives, and assess the function of microalgal feed supplementation on methane emission reductions from the livestock industry, which will facilitate the use of microalgae as ruminant feed supplements in the future.
Availability of data and materials
Not applicable.
References
Aemiro A, Watanabe S, Suzuki K, Hanada M, Umetsu K, Nishida T. Effects of Euglena (Euglena gracilis) supplemented to diet (forage: concentrate ratios of 60:40) on the basic ruminal fermentation and methane emissions in in vitro condition. Anim Feed Sci Technol. 2016;212:129–35.
Alexandratos N, Bruinsma J. World agriculture towards 2030/2050. ESA Working Paper. 2012. https://www.fao.org/3/ap106e/ap106e.pdf. Accessed 1 May 2024
Amorim ML, Soares J, Vieira BB, Batista-Silva W, Martins MA. Extraction of proteins from the microalga Scenedesmus obliquus BR003 followed by lipid extraction of the wet deproteinized biomass using hexane and ethyl acetate. Bioresour Technol. 2020;307: 123190.
Ampofo J, Abbey L. Microalgae: bioactive composition, health benefits, safety and prospects as potential high-value ingredients for the functional food industry. Foods. 2022;11(12):1744.
Anele UY, Yang WZ, McGinn PJ, Tibbetts SM, McAllister TA. Ruminal in vitro gas production, dry matter digestibility, methane abatement potential, and fatty acid biohydrogenation of six species of microalgae. Can J Anim Sci. 2016;96(3):354–63.
Anvar AA, Nowruzi B. Bioactive properties of Spirulina: a review. Microb Bioact. 2021;4(1):134–42.
Barta DG, Coman V, Vodnar DC. Microalgae as sources of omega-3 polyunsaturated fatty acids: biotechnological aspects. Algal Res. 2021;58:102410.
Bichi E, Hervas G, Toral PG, Loor JJ, Frutos P. Milk fat depression induced by dietary marine algae in dairy ewes: persistency of milk fatty acid composition and animal performance responses. J Dairy Sci. 2013;96(1):524–32.
Bito T, Okumura E, Fujishima M, Watanabe F. Potential of Chlorella as a dietary supplement to promote human health. Nutrients. 2020;12(9):2524.
Boeckaert C, Vlaeminck B, Dijkstra J, Issa-Zacharia A, Van Nespen T, Van Straalen W, Fievez V. Effect of dietary starch or microalgae supplementation on rumen fermentation and milk fatty acid composition of dairy cows. J Dairy Sci. 2008;91(12):4714–27.
Camer-Pesci B, Laird DW, van Keulen M, Vadiveloo A, Chalmers M, Moheimani NR. Opportunities of Asparagopsis sp. cultivation to reduce methanogenesis in ruminants: a critical review. Algal Res. 2023;76:103308.
Carvalho JRR, Brennan KM, Ladeira MM, Schoonmaker JP. Performance, insulin sensitivity, carcass characteristics, and fatty acid profile of beef from steers fed microalgae. J Anim Sci. 2018;96(8):3433–45.
Chen G, Chen F. Growing phototrophic cells without light. Biotechnol Lett. 2006;28(9):607–16.
Cholewinska P, Czyz K, Nowakowski P, Wyrostek A. The microbiome of the digestive system of ruminants - a review. Anim Health Res Rev. 2020;21(1):3–14.
Ciliberti MG, Caroprese M, Albenzio M. Climate resilience in small ruminant and immune system: an old alliance in the new sustainability context. Small Rumin Res. 2022;210:106662.
Cooper S, Sinclair L, Wilkinson R, Hallett K, Enser M, Wood J. Manipulation of the n-3 polyunsaturated fatty acid content of muscle and adipose tissue in lambs. J Anim Sci. 2004;82(5):1461.
Costa D, Quigley S, Isherwood P, McLennan S, Poppi D. Supplementation of cattle fed tropical grasses with microalgae increases microbial protein production and average daily gain. J Anim Sci. 2016;94(5):2047–58.
De la Fuente-Vazquez J, Díaz-Díaz-Chirón MT, Pérez-Marcos C, Cañeque V, Sánchez-González CI, Alvarez-Acero I, Fernández-Bermejo C, Rivas-Cañedo A, Lauzurica S. Linseed, microalgae or fish oil dietary supplementation affects performance and quality characteristics of light lambs. Span J Agric Res. 2014;12(2):436.
Dębowski M, Zieliński M, Kazimierowicz J, Kujawska N, Talbierz S. Microalgae cultivation technologies as an opportunity for bioenergetic system development-advantages and limitations. Sustainability. 2020;12(23):9980.
Deepa P, Sowndhararajan K, Kim S. A review of the harvesting techniques of microalgae. Water. 2023;15(17):3074.
Delgado-Pertíñez M, Horcada A. Better animal feeding for improving the quality of ruminant meat and dairy. Foods. 2021;10(5):1076.
Díaz MT, Pérez C, Sánchez CI, Lauzurica S, Cañeque V, González C, De La Fuente J. Feeding microalgae increases omega 3 fatty acids of fat deposits and muscles in light lambs. J Food Compost Anal. 2017;56:115–23.
Dineshbabu G, Goswami G, Kumar R, Sinha A, Das D. Microalgae-nutritious, sustainable aqua- and animal feed source. J Funct Foods. 2019;62:103545.
Drewery M, Sawyer J, Pinchak W, Wickersham T. Effect of increasing amounts of postextraction algal residue on straw utilization in steers. J Anim Sci. 2014;92(10):4642–9.
El-Ghany WAA. Microalgae in poultry field: a comprehensive perspectives. Adv Anim Vet Sci. 2020;8(9):888–97.
Elmore JS, Cooper SL, Enser M, Mottram DS, Sinclair LA, Wilkinson RG, Wood JD. Dietary manipulation of fatty acid composition in lamb meat and its effect on the volatile aroma compounds of grilled lamb. Meat Sci. 2005;69(2):233–42.
El-Sabagh MR, Eldaim MA, Mahboub D, Abdel-Daim M. Effects of Spirulina platensis algae on growth performance, antioxidative status and blood metabolites in fattening lambs. J Agric Sci. 2014;6(3):92–8.
Fan Y, Ren C, Meng F, Deng K, Zhang G, Wang F. Effects of algae supplementation in high-energy dietary on fatty acid composition and the expression of genes involved in lipid metabolism in Hu sheep managed under intensive finishing system. Meat Sci. 2019;157:107872.
FAO. World livestock: transforming the livestock sector through the sustainable development goals. 2018. https://openknowledge.fao.org/items/c1f3e566-9c9a-4314-a1d9-621b0adb3589. Accessed 1 May 2024.
Franklin S, Martin K, Baer R, Schingoethe D, Hippen A. Dietary marine algae (Schizochytrium sp.) increases concentrations of conjugated linoleic, docosahexaenoic and transvaccenic acids in milk of dairy cows. J Nutr. 1999;129(11):2048–54.
Gilbert W, Thomas LF, Coyne L, Rushton J. Review: mitigating the risks posed by intensification in livestock production: the examples of antimicrobial resistance and zoonoses. Animal. 2021;15(2):100123.
Glasson CRK, Kinley RD, de Nys R, King N, Adams SL, Packer MA, Svenson J, Eason CT, Magnusson M. Benefits and risks of including the bromoform containing seaweed Asparagopsis in feed for the reduction of methane production from ruminants. Algal Res. 2022;64:102673.
Gomaa AS, Kholif AE, Kholif AM, Salama R, El-Alamy HA, Olafadehan OA. Sunflower oil and Nannochloropsis oculata microalgae as sources of unsaturated fatty acids for mitigation of methane production and enhancing diets’ nutritive value. J Agric Food Chem. 2018;66(8):1751–9.
Grama SB, Liu Z, Li J. Emerging trends in genetic engineering of microalgae for commercial applications. Mar Drugs. 2022;20(5):285.
Graulet B. Ruminant milk: a source of vitamins in human nutrition. Anim Front. 2014;4(2):24–30.
Gutiérrez-Salmeán G, Fabila-Castillo L, Chamorro-Cevallos G. Nutritional and toxicological aspects of Spirulina (Arthrospira). Nutr Hosp. 2015;32(1):34–40.
Harvatine KJ, Boisclair YR, Bauman DE. Recent advances in the regulation of milk fat synthesis. Animal. 2009;3(1):40–54.
Hathwar SC, Rai AK, Modi VK, Narayan B. Characteristics and consumer acceptance of healthier meat and meat product formulations-a review. J Food Sci Technol. 2012;49:653–64.
Hien HTM, Thom LT, Ha NC, Tam LT, Thu NTH, Nguyen TV, Loan VT, Dan NT, Hong DD. Characterization and optimization of culture conditions for Aurantiochytrium sp. SC145 isolated from Sand Cay (Son Ca) Island, Vietnam, and antioxidative and neuroprotective activities of its polyunsaturated fatty acid mixture. Mar Drugs. 2022;20(12):780.
Holman BWB, Kashani A, Malau-Aduli AEO. Growth and body conformation responses of genetically divergent Australian sheep to Spirulina (Arthrospira platensis) supplementation. J Exp Agric Int. 2012;2:160–73.
Holman BWB, Kashani A, Malau-Aduli AEO. Effects of Spirulina (Arthrospira platensis) supplementation level and basal diet on liveweight, body conformation and growth traits in genetically divergent Australian dual-purpose lambs during simulated drought and typical pasture grazing. Small Rumin Res. 2014;120(1):6–14.
Hopkins DL, Clayton EH, Lamb TA, van de Ven RJ, Refshauge G, Kerr MJ, Bailes K, Lewandowski P, Ponnampalam EN. The impact of supplementing lambs with algae on growth, meat traits and oxidative status. Meat Sci. 2014;98(2):135–41.
Jakobsen AN, Aasen IM, Josefsen KD, Strm AR. Accumulation of docosahexaenoic acid-rich lipid in thraustochytrid Aurantiochytrium sp. strain T66: effects of N and P starvation and O2 limitation. Appl Microbiol Biotechnol. 2008;80(2):297–306.
Ji MK, Yun HS, Hwang JH, Salama ES, Jeon BH, Choi J. Effect of flue gas CO2 on the growth, carbohydrate and fatty acid composition of a green microalga Scenedesmus obliquus for biofuel production. Environ Technol. 2017;38(16):2085–92.
Kholif AE, Morsy TA, Matloup OH, Anele UY, Mohamed AG, El-Sayed AB. Dietary Chlorella vulgaris microalgae improves feed utilization, milk production and concentrations of conjugated linoleic acids in the milk of Damascus goats. J Agric Sci. 2016;155(3):508–18.
Kholif AE, Elghandour MMY, Salem AZM, Barbabosa A, MÁRquez O, Odongo NE. The effects of three total mixed rations with different concentrate to maize silage ratios and different levels of microalgae Chlorella vulgaris on in vitro total gas, methane and carbon dioxide production. J Agric Sci. 2016;155(3):494–507.
Klop G, Hatew B, Bannink A, Dijkstra J. Feeding nitrate and docosahexaenoic acid affects enteric methane production and milk fatty acid composition in lactating dairy cows. J Dairy Sci. 2016;99(2):1161–72.
Kulpys J, Paulauskas E, Pilipavicius V, Stankevicius R. Influence of cyanobacteria Arthrospira (Spirulina) platensis biomass additive towards the body condition of lactation cows and biochemical milk indexes. Agron Res. 2009;7(2):823–35.
Lamminen M, Halmemies-Beauchet-Filleau A, Kokkonen T, Simpura I, Jaakkola S, Vanhatalo A. Comparison of microalgae and rapeseed meal as supplementary protein in the grass silage based nutrition of dairy cows. Anim Feed Sci Technol. 2017;234:295–311.
Liang Y, Bao Y, Gao X, Deng K, An S, Wang Z, Huang X, Liu D, Liu Z, Wang F, Fan Y. Effects of Spirulina supplementation on lipid metabolism disorder, oxidative stress caused by high-energy dietary in Hu sheep. Meat Sci. 2020;164:108094.
Lindahl G, Lundstrom K, Tornberg E. Contribution of pigment content, myoglobin forms and internal reflectance to the colour of pork loin and ham from pure breed pigs. Meat Sci. 2001;59(2):141–51.
Liu G, Yu X, Li S, Shao W, Zhang N. Effects of dietary microalgae (Schizochytrium spp.) supplement on milk performance, blood parameters, and milk fatty acid composition in dairy cows. Czech J Anim Sci. 2020;65(5):162–71.
Lourenco M, Ramos-Morales E, Wallace RJ. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal. 2010;4(7):1008–23.
Ma Y, Wang Z, Yu C, Yin Y, Zhou G. Evaluation of the potential of 9 Nannochloropsis strains for biodiesel production. Bioresour Technol. 2014;167:503–9.
Ma M, Hu Q. Microalgae as feed sources and feed additives for sustainable aquaculture: prospects and challenges. Rev Aquac. 2024;6(2):818–35.
Madeira MS, Cardoso C, Lopes PA, Coelho D, Afonso C, Bandarra NM, Prates JA. Microalgae as feed ingredients for livestock production and meat quality: a review. Livest Sci. 2017;205:111–21.
Marrez D, Cieślak A, Gawad R, Ebeid H, Chrenková M, Gao M, Yanza Y, El-Sherbiny M, Szumacher-Strabel M. Effect of freshwater microalgae Nannochloropsis limnetica on the rumen fermentation in vitro. J Anim Feed Sci. 2017;26(4):359–64.
Martins CF, Ribeiro DM, Costa M, Coelho D, Alfaia CM, Lordelo M, Almeida AM, Freire JPB, Prates JAM. Using microalgae as a sustainable feed resource to enhance quality and nutritional value of pork and poultry meat. Foods. 2021;10(12):2933.
Mavrommatis A, Tsiplakou E, Zerva A, Pantiora PD, Georgakis ND, Tsintzou GP, Madesis P, Labrou NE. Microalgae as a sustainable source of antioxidants in animal nutrition, health and livestock development. Antioxidants (Basel). 2023;12(10):1882.
Meale S, Chaves A, He M, McAllister T. Dose-response of supplementing marine algae (Schizochytrium spp.) on production performance, fatty acid profiles, and wool parameters of growing lambs. J Anim Sci. 2014;92(5):2202–13.
Moate PJ, Williams SR, Hannah MC, Eckard RJ, Auldist MJ, Ribaux BE, Jacobs JL, Wales WJ. Effects of feeding algal meal high in docosahexaenoic acid on feed intake, milk production, and methane emissions in dairy cows. J Dairy Sci. 2013;96(5):3177–88.
Mohy El-Din SM. Accumulation of lipids and triglycerides in Isochrysis galbana under nutrient stress. Appl Biochem Biotechnol. 2019;189(2):359–73.
Nalder TD, Miller MR, Packer MA. Changes in lipid class content and composition of Isochrysis sp. (T-Iso) grown in batch culture. Aquac Int. 2015;23(5):1293–312.
Newbold CJ, Ramos-Morales E. Review: ruminal microbiome and microbial metabolome: effects of diet and ruminant host. Animal. 2020;14(S1):78–86.
Niccolai A, Chini Zittelli G, Rodolfi L, Biondi N, Tredici MR. Microalgae of interest as food source: biochemical composition and digestibility. Algal Res. 2019;42:101617.
Nur F, Erfianti T, Andeska DP, Egistha Putri PA, Nurafifah I, Sadewo BR, Suyono EA. Enhancement of microalgal metabolite production through Euglena sp. Local Strain and Glagah Strain Consortia. Biosaintifika. J Biol Biol Educ. 2023;15(1):36–47.
Odegard IYR, van der Voet E. The future of food - scenarios and the effect on natural resource use in agriculture in 2050. Ecol Econ. 2014;97:51–9.
Olabi AG, Shehata N, Sayed ET, Rodriguez C, Anyanwu RC, Russell C, Abdelkareem MA. Role of microalgae in achieving sustainable development goals and circular economy. Sci Total Environ. 2023;854:158689.
Panjaitan T, Quigley SP, McLennan SR, Swain AJ, Poppi DP. Spirulina (Spirulina platensis) algae supplementation increases microbial protein production and feed intake and decreases retention time of digesta in the rumen of cattle. Anim Prod Sci. 2015;55(4):535–43.
Papadopoulos G, Goulas C, Apostolaki E, Abril R. Effects of dietary supplements of algae, containing polyunsaturated fatty acids, on milk yield and the composition of milk products in dairy ewes. J Dairy Res. 2002;69(3):357–65.
Patel AK, Singhania RR, Awasthi MK, Varjani S, Bhatia SK, Tsai ML, Hsieh SL, Chen CW, Dong CD. Emerging prospects of macro- and microalgae as prebiotic. Microb Cell Fact. 2021;20(1):112.
Ponnampalam EN, Burnett VF, Norng S, Hopkins DL, Plozza T, Jacobs JL. Muscle antioxidant (vitamin E) and major fatty acid groups, lipid oxidation and retail colour of meat from lambs fed a roughage based diet with flaxseed or algae. Meat Sci. 2016;111:154–60.
Ponnampalam EN, Priyashantha H, Vidanarachchi JK, Kiani A, Holman BW. Effects of nutritional factors on fat content, fatty acid composition, and sensorial properties of meat and milk from domesticated ruminants: an overview. Animals. 2024;14(6):840.
Póti P, Pajor F, Bodnár Á, Penksza K, Köles P. Effect of micro-alga supplementation on goat and cow milk fatty acid composition. Chilean J Agric Res. 2015;75(2):259–63.
Rebolloso-Fuentes MM, Navarro-Pe´rez A, Garcı´a-Camacho F, Ramos-Miras JJ, Guil-Guerrero JL. Biomass nutrient profiles of the microalga Nannochloropsis. J Agric Food Chem. 2001;49(6):2966–72.
Ruiz-Gonzalez A, Debruyne S, Dewanckele L, Escobar M, Vandaele L, Van Den Broeck W, Fievez V. Supplementation of DHA-Gold pre and/or postnatally to goat kids modifies in vitro methane production and rumen morphology until 6 mo old. J Anim Sci. 2018;96(11):4845–58.
Šimkus A, Oberauskas V, Laugalis J, Želvytė R, Monkevičienė I, Sederevičius A, Šimkienė A, Pauliukas K. The effect of weed Spirulina platensis on the milk production in cows. Vet Zootech. 2007;38(60):74–7.
Sucu E. Effects of microalgae species on in vitro rumen fermentation pattern and methane production. Ann Anim Sci. 2020;20(1):207–18.
Thapa HR, Lin Z, Yi D, Smith JE, Schmidt EW, Agarwal V. Genetic and biochemical reconstitution of bromoform biosynthesis in Asparagopsis lends insights into seaweed reactive oxygen species enzymology. ACS Chem Biol. 2020;15(6):1662–70.
Tibbetts SM, Fredeen AH. Nutritional evaluation of whole and lipid-extracted biomass of the microalga Scenedesmus sp. AMDD for animal feeds: simulated ruminal fermentation and in vitro monogastric digestibilit. Curr Biotechnol. 2017;6(3):264–72.
Tilman D, Balzer C, Hill J, Befort BL. Global food demand and the sustainable intensification of agriculture [Sustainability Science]. Proc Natl Acad Sci USA. 2011;108(50):20260–4.
Trovão M, Schüler LM, Machado A, Bombo G, Navalho S, Barros A, Pereira H, Silva J, Freitas F, Varela J. Random mutagenesis as a promising tool for microalgal strain improvement towards industrial production. Mar Drugs. 2022;20(7):440.
Udayan A, Pandey AK, Sharma P, Sreekumar N, Kumar S. Emerging industrial applications of microalgae: challenges and future perspectives. Syst Microbiol and Biomanuf. 2021;1(4):411–31.
Urrutia O, Mendizabal JA, Insausti K, Soret B, Purroy A, Arana A. Effects of addition of linseed and marine algae to the diet on adipose tissue development, fatty acid profile, lipogenic gene expression, and meat quality in lambs. PLoS One. 2016;11(6):e0156765.
Vahmani P, Fredeen AH, Glover KE. Effect of supplementation with fish oil or microalgae on fatty acid composition of milk from cows managed in confinement or pasture systems. J Dairy Sci. 2013;96(10):6660–70.
Valenzuela-Espinoza E, Millán-Núñez R, Núñez-Cebrero F. Protein, carbohydrate, lipid and chlorophyll a content in Isochrysis aff. galbana (clone T-Iso) cultured with a low cost alternative to the f/2 medium. Aquac Eng. 2002;25(4):207–16.
van Dijk M, Morley T, Rau ML, Saghai Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat Food. 2021;2(7):494–501.
Wood J, Enser M, Fisher A, Nute G, Sheard P, Richardson R, Hughes S, Whittington F. Fat deposition, fatty acid composition and meat quality: a review. Meat Sci. 2008;78(4):343–58.
Xu C, Zhang S, Sun B, Xie P, Liu X, Chang L, Lu F, Zhang S. Dietary supplementation with microalgae (Schizochytrium sp.) improves the antioxidant status, fatty acids profiles and volatile compounds of beef. Animals. 2021;11(12):3517.
Yan KT, Hie IS, Samaranayake EA, Chang JL, Wang AZ. Medium and process optimizations for Euglena gracilis with high biomass production enriched with protein. Algal Res. 2023;75:103265.
Yan X, Ying Y, Li K, Zhang Q, Wang K. A review of mitigation technologies and management strategies for greenhouse gas and air pollutant emissions in livestock production. J Environ Manage. 2024;352:120028.
Zhu P, Li D, Yang Q, Su P, Wang H, Heimann K, Zhang W. Commercial cultivation, industrial application, and potential halocarbon biosynthesis pathway of Asparagopsis sp. Algal Res. 2021;56: 102319.
Acknowledgements
This work was supported by Natural Sciences and Engineering Research Council of Canada Discovery Grants (L.L.G. and G.C.), the Canada Research Chairs Program (L.L.G. and G.C.), China Scholarship Council (M.Z.), and Agriculture Agri-Food Canada (S.D.S).
Author information
Authors and Affiliations
Contributions
L.L.G. and G.C. conceptualized the review. M.Z., L.L.G., and G.C. generated the outline. M.Z. wrote the first version of the manuscript. All authors edited the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
The work has not been published or under consideration for publication elsewhere. The authors declare that they agreed with the content and that all gave explicit consent to submit and that they obtained consent from the responsible authorities at the institute where the work has been carried out.
Competing interests
The authors declare no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhu, M., Singer, S.D., Guan, L.L. et al. Emerging microalgal feed additives for ruminant production and sustainability. Adv. Biotechnol. 2, 17 (2024). https://doi.org/10.1007/s44307-024-00024-w
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s44307-024-00024-w