
Abstract
In Sub-Saharan Africa, cereals are threatened by stress-inducing microbial pathogens and parasites such as fungi, bacteria, viruses and nematodes leading to significant losses. Mycotoxin-producing fungi like Fusarium graminearum induce head blight disease causing 30–70% of yield losses in wheat. In comparison, Fusarium verticillioides accounts for ear rot diseases that account for 13–70% of maize yield losses in Sub-Saharan Africa. Outbreaks of the devastating rice blast pathogen Magnaporthe oryzae have caused up to 48% yield losses of rice in Kenya. The grey leaf spot fungus, C. zeina, gradually becomes a threat to maize production, causing a 65% yield loss in South Africa. Maize smut and ergot-causing pathogens, Sporisorium ehrenbergii and Claviceps africana, under mildly humid and cold climatic conditions, impact grain quality and market value. The maize streak virus predominantly ravages maize fields all over Sub-Saharan Africa causing 30–100% yield losses while the diseases caused by the bacterium Xanthomonas oryzae pv. oryzae has led to 10 to 50% yield losses in rice production. In Nigeria, nematode pathogens like Pratylenchus spp have been reported to cause yield losses of up to 27% in maize production. This review fundamentally discusses these cereal pathogens and their current and future impact in the face of climate change in Africa. We seriously emphasize the need for more simulation studies on each of these pathogens across Sub-Saharan Africa and their potential impact under current and future climate change scenarios.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Africa is a continent known for its blend of tropical, semi-arid and arid vegetation with varying climate conditions covering an area of more than 7 million acres and is second in size only to Asia [1]. It is home to the world’s second-largest extent of continuous rainforest, the Congo Basin and the Namib desert. Its characteristic weather, diverse soil types and rich vegetation cover make it very suitable for cultivating and producing many stable crops like legumes, cereals, tubers, vegetables and fruits [2]. However, its unique climate creates a convenient environment for diverse pests and pathogens of crops to thrive. These pests and pathogens do not occur randomly and vary according to the specific climate zones within which they are present. As biotic stressors, they are considered a threat to plant health and might include various living organisms such as fungi, bacteria, viruses, nematodes, herbivores, parasitic plants, and weeds which affect the normal development and productivity of susceptible host plants [3]. For example, plant bacteria and viruses cause localized and systemic harm that results in chlorosis and stunting [4]. Fungal and bacterial pathogens invade plant cells through openings or invasively and quickly proliferate thereby inducing several symptoms including wilting, anthracnose, necrosis, defoliation and rots [5, 6]. Nematodes feed on plant tissue and, in synergy with other soil-inhabiting bacterial and fungal pathogens, induce nutritional deficiencies and stunted development in hosts. [7]. Herbivores like insects, mites, rodents, and other animals can also cause significant damage to crops, feeding on plant tissues and reducing plant growth [8]. Parasitic plants such as mistletoe, dodder, and witchweed attach themselves to host plants and tap into their vascular systems to obtain nutrients, further impacting the host plant's growth and productivity [9]. Weeds compete with crops for resources, reducing yield and increasing management costs [10]. Global agricultural production is severely impacted by these biotic stressors (pests and diseases) which cause estimated losses of up to $220 billion annually [11]. With an expanding world population, there is increased pressure to improve crop yield and production [12], and biotic stress management will become even more crucial.
Like all staple crops in Africa, cereals are affected by many plant pathogens. Africa, among other producing regions, plays host to 27% of the world's total cereal production [13]. The top 3 producers of cereals in Africa are Ethiopia, Nigeria and Egypt (Fig. 1). All cereal crops are rich sources of carbohydrates, dietary fibre, vitamins, and minerals [14] and are major components of local diets in African countries. Maize (Zea mays), Africa’s most widely grown cereal (Fig. 2), serves several purposes [15]. In East and Southern Africa, maize or corn is an essential staple food consumed as flour, meal, and porridge [16]. In West Africa, it is often eaten as a roasted snack or boiled maize cobs [17]. It also serves as a source of animal feed as well as a key ingredient for beer production [18]. Another cereal, sorghum (Sorghum bicolor), is widely grown in the arid and semi-arid regions of Africa [19] and used to produce traditional alcoholic beverages like "chang'aa," and "dolo" in East and West Africa, respectively. In the Sahel region, however, millet (Pennisetum glaucum) is often consumed as porridge or a side dish [20] and used to produce "tchapalo", a local alcoholic drink in West Africa. Rice (Oryza sativa) is cultivated in West Africa and is often consumed with a stew or sauce [21]. It can be cooked into a variety of traditional dishes, including jollof rice, biryani, and paella [22]. Wheat (Triticum spp), which is more often grown in northern and southern Africa, is a basic ingredient for making bread, cakes, and other baked products [23]. Barley (Hordeum vulgare) is mainly used in the production of beer and other alcoholic beverages [24].

Top 10 producing countries of total cereal in Africa as of 2021. Percentage values are derived from total cereal production by the ten countries (not total production in Africa). Ethiopia and Nigeria are leading countries. Source [13]

Percentage production of different cereal crops in Africa as of 2021. Maize is the most cultivated cereal followed by rice and wheat. Source [13]
The economic impact of plant pathogens and microbial parasites on cereal production in Africa is significant with massive losses incurred coupled with reduced marketability [25]. However, in the wake of climate change events, it appears drastic modifications to natural weather patterns might favour an increased incidence of diseases caused by these pathogens in cereal-producing regions or rather a decrease. Globally, alternating temperature and precipitation patterns, alongside with sharp changes in atmospheric pressure and humidity levels, characterize climate change. The long-lasting feature of climate change makes it difficult to accurately predict. This is in contrast to seasonal variations in weather patterns which are predictable on an annual basis using existing weather data [26, 27]. Africa is also faced with undulating daily temperatures, irregular rainfall patterns, and significant changes to microclimates, yet less attention is given to their potential impact on cereal production.
The rapid spread of diseases, events of climate change, and the lack of effective and affordable control measures plus little or no technical knowledge, have all complicated the severity and management of crop pests and diseases. This review provides a comprehensive overview of the current threats posed by biotic stressors, specifically fungi, bacteria, viruses and nematodes, on cereals in Africa. Through an analysis of the most recent scientific research, we have identified major diseases affecting cereals in this region and explored the underlying causes of their spread. By understanding the challenges posed by these biotic factors on cereals, we can prioritize areas in need of improved management strategies for increased agricultural production. Ultimately, this will help to ensure food security and enhance the livelihoods of farmers in Africa. Below, we highlight and give an overview of major pathogens affecting different cereal crops in Africa (Fig. 3).

Disease symptoms of some current pathogens affecting cereal production in Africa. From left to right: Magnaporthe oryzae (rice blast), Claviceps africana (sorghum ergot), Fusarium verticillioides (Gibberella rot), F. graminearum (wheat head blight), Cercospora zeina (grey spot), Maize streak virus, Ustilago maydis (maize smut), Maize lethal necrosis virus, Xanthomonas oryzae pv. oryzae and Rice yellow virus
2 Fungal pathogens threatening African cereal production
Several plant pathogenic fungi attack cereals and include the following examples: wheat rust fungi Puccinia spp., the rice blast fungus Magnaporthe oryzae, and mycotoxin-producing species like the Aspergillus flavus fungus¸the wheat head blight fungus Fusarium graminearum [28], and the maize ear rot-causing fungi Fusarium verticillioides. Globally, including in Africa, these fungal pathogens, and more, negatively impact yield quantity and quality, especially in cultivated fields characterized by conventional tillage, high nitrogen fertilization and monoculture cultivation [29]. In Table 1, we highlight fungal pathogens known to affect cereal crops cultivated in Africa. We then provide an overview of a few examples whose economic importance in Africa has been well documented and might become a more severe threat to cereal production in the face of climate change events within the region.
2.1 Mycotoxin-producing Fusarium species
The mycotoxin-producing Fusarium species are dominant fungal pathogens known to cause ear rot and head blight on grain crops including wheat, barley and maize [45]. Two species of Fusarium; F. graminearum Schwabe and F. verticillioides (Saccardo) Nirenberg are associated with mycotoxin synthesis [46]. In many cases, both Fusarium species are associated with the fusarium head blight disease in wheat as well as ear rot in maize [47]. However, more often, studies have shown that Fusarium graminearum is the most predominant of both species isolated from plant parts showing wheat head blight symptoms [48]. On the other hand, F. verticillioides appears to be the predominant causal pathogen of ear and stalk rots in maize [49]. In Africa, the economic presence of both fungal pathogens has been documented across South Africa, Ethiopia, Kenya, Nigeria, Botswana, and Mozambique [30, 50, 51].
2.2 Head blight-causing pathogen and Gibberella ear rot-Fusarium graminearum
F. graminearum, (teleomorph Gibberella zeae), the causal agent of head blight on wheat [52] and ear rot on maize [53], is ranked fourth among plant fungal pathogens based on its scientific and economic importance [28]. A homothallic saprophyte in crop residues, the fungus infects host plants through air-borne sexual (ascospores) and asexual spores (conidia) [54]. Characteristic symptoms of F. graminearum include yellowing to slight discolouration of infected spikelet (bleaching), late tillering and reduced vigour [55]. F. graminearum produces zearalenone (ZEA) and deoxynivalenol (DON), mycotoxins that are commonly detected in several wheat-producing parts of Africa [56]. In Nigeria, zearalenone levels of 115–779 μg/kg, which exceeded the legal allowable limit (LAL) of 100 μg/kg set by the European Commission (EU), have been detected in maize [57, 58]. In Tunisia, 83% of 65 wheat samples were contaminated with deoxynivalenol levels ranging from 12.8–30.5 μg/g which were above allowable limits, the maximum permitted limit of 1.75 μg/g set by the EC [59]. Severe outbreaks or epidemics of FHB caused by F. graminearum on wheat cultivated under centre-pivot irrigation have been recorded in the Northern Cape and KwaZulu Natal regions of South Africa with major losses [60]. In Ethiopia, the F. graminearum species complex is considered the second major contaminant of maize kernels [51]. In Tunisia, 30 to 70% of yield losses in wheat have been attributed to FHB [31] caused by F. graminearum.
2.3 Fusarium ear rot-causing pathogen-Fusarium verticillioides
F. verticillioides (teleomorph Gibberella fujikuroi) is mainly associated with maize by inducing ear and stalk rots [52]. F. verticillioides, as a heterothallic, endophytic pathogen, attacks maize kernels through airborne conidia which infect the silks or through conidia-infected seeds [61]. Disease infections and epidemics occur and are sustained by moderate rainfall, high humidity and temperature range between 24 and 29 °C—environmental conditions suitable to the natural climate of tropical Africa [55]. F. verticillioides induces the appearance of brown streaks which appear at the lower internodes and rotten whitish-pink pith tissue [62]. The Fusarium stalk/ear rot of maize has been associated with 30–50% losses in severely affected areas [49]. Specifically, the maize ear rot disease is widespread in major maize-producing regions of Africa including South Africa, Nigeria and Tanzania with major losses [60]. In Sub-Saharan Africa, F. verticillioides plays a vital role in 13–70% of maize yield losses caused by ear rot [32] and has been extensively detected and isolated from maize kernels in Ethiopia [51], Zambia [63] and South Africa [64]. F. verticillioides is renowned for the production of fumonisins in major maize-producing countries like South Africa, Algeria, Ethiopia and Nigeria [65]. High levels of fumonisins have been recorded in rural maize production areas of South Africa with levels up to 117 mg/kg, 50 times more than the maximum tolerable daily intake recommended by the Joint FAO/WHO Expert Committee on Food Additives [64]. Studies have detected fumonisin B1 levels of more than 1–3 mg/kg in high-quality maize kernels obtained from the Kisii district of western Kenya and other regions [66, 67]. In Tanzania, an extensive survey demonstrated the presence of fumonisin (FB2) levels between 178 and 38,217 μg kg−1 in 48% of maize kernels sampled [68] which were less than those reported in Malawi (1–7 mg kg−1) and Benin (8240–16,690 mg kg−1), respectively [32]. In Ethiopia, F. verticillioides is mainly responsible for the presence of fumonisin in maize kernels [51]. These and many more studies confirm the current and future devastating impact of the mycotoxins produced by these Fusarium species. Across Africa, head blight and ear rot disease conditions are widespread in cereal-producing regions.
2.4 The blast-causing fungi-Magnaporthe oryzae
Magnaporthe oryzae is a hemibiotrophic ascomycete fungus (anamorph Pyricularia oryzae Sacc.) infamous as the causative agent of the devastating rice blast disease [69]. With a wide host range of more than 50 species of Poaceae plants, Magnaporthe oryzae infects several cereals including rice, wheat, maize, barley and millet [34]. In Africa, blast symptoms induced by Magnaporthe oryzae were first identified in Uganda around 1922 followed by subsequent reports of its presence and economic impact in Ghana, Kenya, Congo, Egypt, Madagascar, Morocco, Senegal, South Africa and all rice-growing areas of Sub-Saharan Africa [35]. It’s no surprise given that many rice-cultivated fields in Africa are located in the upland ecologies characterised by high moisture that is 12 h or longer and warm temperatures [70]. An aggressive and dynamic pathogen, its conidiophores infect leaves, collars, panicles, and seeds under favourable conditions. Typical symptoms are characteristic elliptical grey-white lesions on any above-ground organ (leaves, neck or panicles). Severe infections of the neck have been known to cause 100% yield losses in sub-Saharan Africa [69]. Rice blast disease, Magnaporthe oryzae, has been highly devastating to small-scale producers of rice in Africa who cannot afford fungicides. Outbreaks of the disease have ravaged rice fields causing up to 48% yield losses in Kenya in 2009 [71]. The epidemic destroyed 13,840 acres of rice in the Central Province of Kenya, the major rice production area, hence making the country a nett importer of rice [71]. Even in new rice-growing areas within Tanzania, Burundi, and Rwanda, the pathogen has been detected through its symptoms [69]. In Zambia, blast symptoms caused by Magnaporthe oryzae have been observed on wheat crops grown in rainfed fields with a high disease incidence rate of about 84.8% [72]. Similarly in Kenya and Uganda, the fungal pathogen has led to a 100% incidence of blast disease symptoms in finger millet-growing areas with 10–80% yield losses [36] and 30–50% yield loss in rice-growing regions across Nigeria [73]. Certainly, the Magnaporthe oryzae and its existing pathotypes pose a huge threat to the regional production of rice in Africa.
2.5 Smut disease pathogens
The smut disease is known to affect major cereal crops such as maize, sorghum and pearl millet. The common smut fungus (Ustilago maydis), the head smut fungus (Sphacelotheca reiliana) and the sorghum long smut (Sporisorium ehrenbergii) cause severe damage to cereals globally [41, 74, 75]. The main characteristic symptom is the random and non-systemic development of deformed heads which transform into cylindrical, and vaguely curved sori covered with a thick creamy-brown membrane called peridium [42, 76]. In Africa, S. ehrenbergii is more distributed and aggressively devastating among the smut-causing fungi. The fungal pathogen is predominantly widespread in major sorghum-producing regions of Africa (Nigeria, Burkina Faso, Ethiopia, Senegal, Botswana, Malawi, Tanzania, Zimbabwe, Zambia and Egypt) and some parts of Asia (India, Turkey and Pakistan) causing 20–60% yield losses in sorghum-cultivated areas characterized by low rainfall, low soil moisture, and high temperatures. For example, the severity of the long smut disease of sorghum in Nigeria has been linked to persistent drought conditions [41]. A few studies in the past years have revealed the economic significance of S. ehrenbergii as the causal agent of the long smut disease of sorghum. In Nigeria, drought conditions influenced 20–60% of sorghum yield losses [42]. In Ethiopia, fields in South Tigray- a major sorghum cultivation area, were characterised by 88.6% field infection of long smut [77]. However, in recent times, despite the widespread incidence of the long smut disease in major sorghum-producing areas of Sub-Saharan Africa, little is known about its economic impact on sorghum production as well as its potential threat in the wake of climate change events.
2.6 Ergot-causing pathogens
Ergot, an important disease condition of grass inflorescences, is induced by certain species belonging to the genus Claviceps. Four species of economic relevance include C. purpurea, C. africana, C. fusiformis and C. sorghi. In Africa, C. africana is more predominant having originated from within the region and has a significant impact on all sorghum-producing areas of the region [39]. The first reference to the ergot disease of sorghum dates to a famine in 1903–1906 in Northern Cameroon, West Africa. Subsequent studies identified the economic presence of C. africana in Uganda, Kenya, Tanzania, South Africa, Nigeria, Zimbabwe and then Zambia [39, 78]. Infection is achieved by airborne macroconidia infecting the stigma of an unfertilized flower. Two to three weeks before flowering, cold night temperatures (less than 12 °C) favour ergot infection on sorghum by influencing the sterility of the host’s pollen [40]. Severe infections can result in substantial yield losses pre- and post-harvest. In Zimbabwe, a yield loss of 12–25% had been reported in hybrid seed production due to ergot disease. Similarly, in southern Africa, an 80% yield loss has been reported in hybrid sorghum seed and grain production [40]. Like some previously mentioned economically important fungal pathogens of cereals, recent extensive studies have not been done to estimate the economic impact of C. africana and its relatives on sorghum production as well as its potential threat in the wake of climate change events.
2.7 Grey leaf-spot-causing pathogens
Grey leaf spot (GLS) is one of the most important yield-reducing foliar diseases of maize in every region where the crop is cultivated. The disease is associated with two species: Cercospora zeae-maydis Tehon & E.Y. Daniels and C. zeina Crous & U. Braun [79]. Between both species, C. zeina has only been reported in Africa, China, Nepal and a few parts of Brazil and the United States [80]. Based on the scope of this study, we will focus on C. zeina which was first documented in KwaZulu-Natal Province, South Africa in 1988 causing 65% yield loss of maize and later confirmed in the later years to be genetically distinct from C. zeae-maydis [81]. C. zeina is now widespread in Sub-Saharan Africa inducing massive losses in Cameroon, DR Congo, Kenya, Rwanda, Uganda, South Africa, Zambia and Zimbabwe [37]. C. zeina is a polycyclic host-specific hemibiotroph ascomycete fungus that infects only maize with no current report of another host. Like other Cercospora species, the pathogen is disseminated, as conidia, by wind and rain splash while it overwinters on maize debris. On susceptible maize plants, symptoms appear as tan to grey rectangular lesions on leaves and can become very severe during flowering, extending to the stalks and husks. Although there is no robust account of how much losses are caused by C. zeina, a rough estimate puts it at 60 to 80% yield loss [80].
3 Viral pathogens threatening African cereal production
Viruses are primarily obligate unicellular pathogens with a seemingly simple structure composed of only one type of nucleic acid, either DNA or RNA and coated with a protein. Their shapes and sizes can vary from isometric or rod-shaped to gemini forms, [82]. Viruses make up 47% of the pathogens that cause emerging and re-emerging plant disease epidemics worldwide with an estimated global economic impact of > US$30 billion annually [83]. Some economically important viral diseases causing severe yield losses to cereals include rice yellow mottle virus (RYMV), African Cereal Streak Virus (ACSV), maize lethal necrosis disease (MLN), maize chlorotic mottle virus (MCMV), maize streak virus (MSV) etc., and these have been well documented in Africa [84]. The most common vectors of cereal viruses that have been implicated in their transmission include the more familiar leaf/plant hoppers, aphids, whiteflies, mites and others organisms such as fungi and nematodes [85]. Re-emerging virus disease epidemics are often associated with cereals, examples of which are cereal yellow dwarf, rice tungro and wheat streak mosaic [84]. For example, maize lethal necrosis disease emerged as a pandemic on maize crops across East and African countries such as Ethiopia, Kenya, Uganda and Tanzania covering 296 million acres [86]. The menace caused by viral diseases to cereal production worsens in tropical regions in Africa where arable lands are barely left uncultivated all year round. These crops are thus under recurrent stress due to virus particles spreading primarily by insect vectors from both cultivated and neighbouring wild plants [87]. Farmers, researchers, and policymakers have always taken great interest in plant viruses because of their tremendous yield losses to agriculturally important cereal crops like maize, rice, millet, sorghum, and wheat. As these crops constitute a major component of the world’s food supply, viral disease losses, therefore, take their toll on both farmers and consumers as they lead to reduced crop yields, which culminates in increased food prices and impact food security [88]. Intensive crop production and the commensurate increase in vector populations are vital players in the widespread distribution of cereal viruses. Losses induced by virus diseases rely on factors such as the incidence, severity, and duration of infection. Although the insect vectors can be controlled by insecticide seed treatments, these RNA and/or DNA viruses cannot be controlled directly by application of pesticides or agro-allied chemicals. Hence, healthy seeds and employing genetic resistance are central to ensuring high quality and abundant yields [89]. Table 2 gives a summary of plant viruses known to affect cereals in African countries.
Below, we provide an overview of a few cereal-infecting plant viruses and their corresponding economic importance in some regions of Africa.
4 Maize lethal necrosis (MLN) virus complex
Maize Lethal Necrosis (MLN) is considered the most devastating disease posing a serious threat to maize production globally. A synergistic infection between the maize chlorotic mottle virus (MCMV) and several maize-infecting viruses in the family Potyviridae is responsible for this viral disease [86]. It is widely distributed in East African countries including Ethiopia, Kenya, Tanzania, DR Congo, Uganda, and Rwanda with recurrent outbreaks recorded. Outbreaks in the Southern Rift Valley of Kenya caused an estimated 30–100% yield loss of almost all commercial maize varieties in 2011 [98]. Ever since, smallholder farmers across Ethiopia, Kenya, Rwanda, Tanzania, and Uganda have experienced an estimated combined loss of US$291–339 million [98]. Crop losses reach 73% of grain loss for susceptible varieties during high disease severity and incidence based on a recent simulation. Two recent robust reviews on maize lethal necrosis disease in Sub-Saharan Africa have been put together by Biswal et al. [96] and Redinbaugh and Stewart [86]; hence we choose not to extensively delve into many details.
4.1 Rice yellow mottle virus (RYMV)
RYMV, caused by Solemoviridae, is delineated as the most economically important viral pathogen of rice production in SSA [99]. First identified near Lake Victoria in East Africa in 1966, it has spread spatially towards the rice-growing regions of western Africa, becoming endemic and infecting different rice cultivars from Oryza sativa to Oryza glaberrima, and wild rice like Oryza logistiminata and Oryza barthi [100]. Infection occurs mainly through wounds and insect vectors. Common symptoms of the disease include yellow-green streaks, intense leaf mottling, stunted plants, decreased tillering, and sterile panicles [90]. Transmission is a result of plant injuries sustained during weeding and transplanting. Insects (chief vectors), rodents and farm animals also facilitate the spread of the virus. The primary vectors are chrysomelid beetles which transmit the virus semi-persistently [99]. Depending on the time of infection, the strain of the virus, rice genotype, and ecology, yield losses from RYMV can range between 10 to 100% [90]. In western Africa, recent studies and reports have revealed the massive economic impact of disease conditions induced by RYMV. In Burkina Faso, losses of 84% have been recorded on popular varieties such as FKR56N, FKR62N, and TS2 similar to over 90% yield losses reported in susceptible rice varieties cultivated in Nigeria and Côte d’Ivoire [101]. Sporadic infections of RYMV across rice fields have been observed in Niger with high incidences in other Sub-Saharan countries like Uganda (50–75%), Burundi, Ethiopia, Tanzania, Zimbabwe, the Central African Republic and the Democratic Republic of Congo [99]. With such reports, the RYMV stands out as a major biotic stressor threatening rice production in Africa.
4.2 Maize streak virus (MSV)
Maize streak virus (Genus Mastrevirus, Family Geminiviridae) is probably the most damaging viral crop disease in Africa infecting maize and a host of over 80 other plants belonging to the Poaceae family [102]. First recorded in southern Africa, the pathogen has spread to eastern and western Africa with reports of its presence in Nigeria, Kenya, Zaire, Zimbabwe and surrounding islands in the Indian Ocean [103]. While a single strain of the virus (MSV-A) causes streak disease in maize, other strains are known to infect barley, wheat, oats, rye, sugarcane, millet and many wild, mostly annual, grass species [104]. It is mainly transmitted by eight different species of the cicadellid leafhopper [93]. The complexity of the virus-vector biological interaction, in combination with meteorological and geographical factors, has made the epidemiology of the maize streak disease erratic in nature and poorly understood [105]. MSV is not disseminated mechanically by seed or pollen. Therefore, the epidemiology and ecology of MSV depend solely on the movements of its vector species [93]. Symptoms of infection of maize include the appearance of tiny, pale, round spots on the lowest open portion of young leaves which extend into white to yellow chlorotic streaks/lesions. Severe cases result in the death of the plant often at the seedling stage [47]. Drought conditions or irregular precipitation patterns are usually associated with outbreaks of the disease caused by MSV- hence, climate change events with such trend might make MSV a serious future threat to maize production. The virus remains uncontrolled in many parts of Africa and during epidemic periods, it influences widespread yield losses and famine [103]. Yield losses from 30 to 100% on maize crops have been reported by authors with a record US$120–480 million being lost annually in Sub-Saharan Africa [94]. The maize streak virus remains a huge threat to maize production and hence stands out as a major biotic factor affecting maize production.
5 Bacterial pathogens threatening African cereal production
With over 150 species widely distributed around the world, especially in subtropical and tropical regions, plant bacterial pathogens are responsible for various plant diseases characterised by typical symptoms including blights or deadening of tissue, spots on leaves or fruits and rotting or abnormal growths on roots [106]. In many cases, based on yield losses, bacterial infections are not as severe as viruses or fungi; however, the bacteria can enhance the susceptibility of the host plant to occurring abiotic stress like frosts leading to substantial damage [107]. Plant-pathogenic bacteria are explicitly dangerous in their ability to become endemic due to the ease in their intentional or unintentional dispersal through rain splash, infected seeds, water current within the soil matrix and propagation by infected farm equipment [108]. Among the wide range of crops affected by plant-pathogenic bacteria, cereals like maize, wheat, barley and rice play host to different bacterial pathogens, especially those belonging to the genera Erwinia, Pseudomonas, and Xanthomonas [109] (see Table 3 for a summary).
Below, we discuss a few bacterial pathogens that have been identified to cause damage to cereals cultivated within the African region.
5.1 Xanthomonas oryzae pv. oryzae (Xoo)
Xoo is a rod-shaped, non-spore-producing gram-negative bacterium known to cause the leaf blight disease of rice [115]. Distributed widely across Australia, Africa, Latin America, the Caribbean and the USA, it causes 10%–50% yield losses with epidemic reports recorded in the monsoon season in South-East Asia and India [116]. The pathogen is aerobic and thrives best between 27 ± 3 °C, between 6.5–7.5 pH [117]. Although Xoo primarily lives on infected seeds, roots of weeds and perennial plants like Leptochola chinensis, L. panacea and wild Oryza species. In rice plantations, Xoo is disseminated through irrigation water systems, splashing or wind-blown rain, as well as by contaminated rice stubble from the previous crop season, which is the most important source of primary inoculum [118]. A characteristic symptom of the pathogen is the release of exudates from the infected leaf tissues due to the clogging of xylem vessels with bacterial cells and extracellular polysaccharides [116]. Other symptoms could include the appearance of long, grey-to-white, opaque necrotic lesions [115]. The economic importance corresponds to the virulence of the pathogen which is greater in the irrigated and rain-fed lowland ecosystems. Xoo is responsible for severe yield loss and can survive the climate in temperate and tropical countries in Africa [119]. In Africa, bacterial blight was first reported in Mali over forty years ago and later spread across other countries including Senegal, Niger, Mauritania, Benin, Cameroon, Nigeria and Gabon [110]. The spread of the pathogen can be linked to the introduction of hybrid cultivars without strict quarantine measures in African borders. An increase in production using hybrid varieties can be costly in the case of using highly susceptible genotypes, for instance, in the case of Mali importing susceptible hybrid varieties from China [110]. About 10–50% yield losses in rice production have been recorded in some African countries as a result of leaf wilting caused by Xoo on rice plants [111]. Unique strains of Xoo have been isolated from rice plants cultivated in Burkina Faso, Mali, Niger and Cameroon showing the diversity of the bacterial pathogen across Western Africa [110].
5.2 Bacterial streak causing Xanthomonas
Several other species of Xanthomonas are known to cause bacterial streak symptoms on certain cereals such as X. translucens pv. undulosa (Xtu), another recognized Gram-negative bacterium that affects small grains such as wheat, barley and rye [114]. Infections caused by Xtu are favoured by warm and humid environmental conditions with high severities in fields irrigated with sprinklers [113]. After invading leaf tissues, Xtu induces water-soaked lesions which progress into greasy and necrotic longitudinal streaks. Reduced photosynthetic area and premature defoliation can lead to a reduction in grain fill and kernel weight. Predominantly considered a seed-borne pathogen, it overwinters in crop debris, winter cereals and some perennial weeds like smooth brome and quackgrass [120]. In Africa, this bacterium has been identified in Ethiopia, Kenya, Madagascar, Morocco, South Africa, Tanzania, Tunisia, and Zambia [114]. The Xanthomonas vasicola pv. vasculorum (Xvv) is another bacterial pathogen that exclusively infects maize and was previously restricted to South Africa. Xvv invades the leaves through stomata and colonizes the intercellular spaces. Typical symptoms are long thin yellow-to-brown lesions running parallel to the leaf vein and having wavy margins. However, there are no records of significant yield losses [112]. Diseases caused by plant pathogenic bacteria are not very prominent in Africa in terms of economic losses; however, they cannot be disregarded.
6 Nematode pathogens threatening African cereal production
Plant parasitic nematodes (PPN) are widely recognized as a major hindrance to crop production globally [121]. PPN pose a severe threat to cereal production worldwide, resulting in yield losses of up to 10% and causing economic losses estimated to be over USD 125 billion per year [122]. The yield loss caused by plant parasitic nematodes can vary greatly, depending on the species involved and other environmental conditions, and can reach up to 90% [123]. Abdulsalam et al. estimated that cereals production in Nigeria experienced an 18–67% yield reduction due to PPN, including 20–28.5% in maize, 20–30% in sorghum, and 25–67% in rice. Root lesion nematodes have been reported to cause a 27% decline in maize production in most areas of the South Western zone of Nigeria [125]. In Tunisia, PPN has resulted in yield reductions of 19–86% in barley and 26–96% in wheat [126]. With numerous genera of PPNs causing damage to yield or cereals, Meloidogyne spp., Heterodera spp., and Pratylenchus spp. have been identified as the most damaging to cereal production [122, 123]. While Heterodera spp. is less widely distributed in West Africa, Meloidogyne and Pratylenchus nematodes are frequently reported as causing significant damage to cereal production in this region [124]. Despite their importance as pathogens, there is limited data available regarding the loss in crop yield caused by these nematodes, as other factors such as drought, water stress, and nutrient deficiency can also contribute to yield loss [124]. Table 4 gives a summary of plant-parasitic nematodes known to affect cereals in African countries.
Below, we discuss a few nematode parasites that have been identified to attack some cereals cultivated within the African region.
6.1 Root-lesion nematodes (Pratylenchus spp)
The root-lesion nematodes (RLNs) are regarded as one of the most important plant-parasitic nematodes causing economic damage to cereals especially maize especially when existing in disease complexes with other aggressive maize-associated soil pathogens like Fusarium and Rhizoctonia species [131, 132]. The migratory habits of RLNs make it difficult to access their presence and damage, hence, despite being a major biotic stressor, RLNs have been relatively neglected in research [133]. As polyphagous and migratory endoparasites, Pratylenchus nematodes invade the endodermis, epidermis and cortex of the root and induce the formation of necrosis by their feeding activity which extends into darkened lesions in high populations [134]. Often, these symptoms are intensified by the rotting of roots due to secondary attacks from opportunistic soil fungi or bacteria. Consequently, a “water deficit” condition is induced which alters the plant’s physiological and biochemical processes. Cumulatively, these symptoms result in poor growth of maize plants and significant yield reductions [135]. On maize, the RLNs have been recorded to induce up to 28% grain yield losses making them a severe threat indeed [136]. RLNs have been reported as the most significant plant-parasitic nematodes in cereal-growing regions of Morocco [137]. In Nigeria, RLNs cause yield losses of up to 27% in maize production [125].
6.2 Root-knot nematodes (Meloidogyne spp)
Root-knot nematodes (RKNs) are the most damaging of all nematode genera with a vast host range that includes cereals with wide distribution in both tropical and temperate climates [138]. RKNs are typically characterized by galling symptoms on the roots of infected crops while above-ground symptoms do not depict their parasitic activity on the host crop [139]. Damages induced by their parasitic activities within the root tissues correlate to impaired water and nutrient uptake which influence loss of vigour and retarded development [140]. On maize, the effect of the RKN may not be very pronounced. Roots of the crop can look healthy with very tiny and almost unnoticeable galls and yet harbour a significant population of the nematode [141]. Heavy infection can drastically restrict the root system to shallow depths of only 8 or 10 cm below the surface succeeded by heavy losses if unchecked [142]. Despite limited information on the economic impact of RKNs on these crops, studies have shown that they can cause significant damage. M. incognita was found to reduce maize ear weight by 64% and grain yield by 56% in western Nigeria [124], while maize yield losses of up to 60% were reported in South Africa due to parasitism by either single or mixed populations of M. incognita and M. javanica [129]. In South Africa, Meloidogyne acronea is a reported parasite of sorghum, with studies indicating that it can cause yield reductions of 56–90% [124]. RKNs have also been shown to cause losses to sorghum in Malawi [143]. While RKNs have been associated with wheat in Nigeria, their exact role as parasites of wheat remains unclear [124].
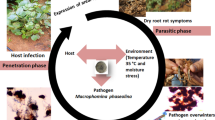
7 Climate change and the future challenges of cereal pathogens in Africa
Climate change is no new concept within the global space and has been characterised by a rise in global temperatures, changing precipitation patterns, elevated CO2 levels and extreme weather events. It differs from climate variability which refers to cyclical variations in temperature and/or precipitation within a year. Climate change is characterized by fluctuations in the climate that last for decades or longer yet seasonal climate variations only last longer than individual weather occurrences [27]. Although the impacts of climate change are not evenly distributed around the world, some regions are more vulnerable than others [144]. One such region is Africa which is warming at a faster rate than the rest of the globe with records of drought/flood disaster events, which already constitute 25% of disasters on the continent. By 2025, many parts of Africa are expected to face increased water stress and scarcity [145]. According to the Global Climate Risk Index 2021, seven of the ten countries most affected by climate change from 2000 to 2019 were in Africa. Over 80% of Africa's population depends on rain-fed agriculture for their livelihoods, which makes them particularly vulnerable to climate change.
In Africa, like other regions of the world, climate change events are expected to increase the spread of various plant pathogens leading up to increased incidence and severity of plant disease outbreaks which pose significant and growing risks to primary productivity and global food security. At the same time, the possibility of climate change events fostering reduced disease incidence and outbreaks is not unlikely. Both scenarios might be aided by altering pathogen evolution, changing host–pathogen interactions, the emergence of new strains of pathogens, and modifications of host-plant resistance [146]. On one hand, increased precipitation events will force microbial parasites like the nematodes as well as plant bacteria to proliferate and spread across various cereal-growing regions of Africa leading to more devastating losses [147]. In contrast, prolonged drought and warm temperatures will hamper the survival and pathogenic propensity of other pathogens that primarily rely on very humid conditions. Population dynamics of plant pathogens, their growth rates and the number of generations per certain period (for polycyclic species) are greatly influenced by weather parameters and hence will be impacted by climate change [146].
Temperature changes will play a predominant role in the future impact of various cereal pathogens. It is expected that warming temperatures will encourage the emergence of new pathogen strains that are more adaptable to new climatic conditions and at the same time, be more virulent [146, 148]. In wheat-producing regions of Sub-Saharan Africa that are getting warmer, the milder Fusarium culmorum might be replaced by the more aggressive F. graminearum, potentially leading to potential heavy losses. This is because F. graminearum thrives well in warm and humid conditions compared to F. culmorum which prefers cool and wet conditions [146]. However, in maize, it is speculated that warming temperatures and drought will foster an increased incidence of F. verticillioides at the expense of F. graminearum in the tropics [149]. Warming temperatures are also expected to increase rice susceptibility to rice blast disease through the induction of jasmonic acid biosynthesis and signalling genes by Magnaporthe oryzae [150]. A report has also shown that the severity of rice bacterial blight, caused by Xanthomonas oryzae pv. oryzae, increased under high temperature due to the breakdown of all tested resistance genes in the rice cultivar Oryza sativa [151]; although another study reports an opposite observation for a particular gene Xa7 in Oryza glaberrima [152].
Plant viruses and the activities of their insect vectors are greatly influenced by changes in climatic factors, especially temperature. At increasing temperatures, host plants defences, depending on the crop, might weaken and allow widespread viral infection, if the virus’ optimum temperature is not exceeded as well. In contrast, hardy crops like maize might be able to withstand high temperatures which do not favour its viral pathogens as well as combat any potential infection through anti-viral RNA silencing [153]. These dynamics in virus-plant interactions are unpredictable and can swing in any direction, whether positive or negative. Generally, elevated temperatures are also expected to promote the activities and populations of insect vectors which correlates with the severity of plant viruses like the maize lethal necrosis viruses and maize streak viruses [149]. For example, a field study showed a rapid increase in the vector population of the leafhopper Cicadulina mbila corresponding with a higher incidence of maize streak disease as temperature spiked above the 24 °C threshold [154]. In a different context, natural enemies of viral vectors might begin to migrate towards new areas; hence spreading the intensity of the disease conditions, especially in places where it was previously not established [96]. Although drought conditions accompanied by higher temperatures can affect the transmission of plant viruses by increasing their reproductive success and feeding rates [155], pathogens like the maize streak virus are well known to exhibit an unpredictable evolutionary nature in the face of climate change, making it a potentially serious threat to maize production in Africa within the next decade [156].
Plant parasitic nematodes, on the other hand, are speculated to reduce their reproductive capacity with increasing soil temperature (from 32 to 34 °C) but increase their virulence, leading to potential crop losses. This was based on a recent study that evaluated the effects of soil temperature on survival, reproduction, virulence, and disease severity of Rotylenchulus reniformis and Meloidogyne floridensis [157]. Although not a major pathogen of grain crops, nematodes like Radophilus similis, which are prevalent at lower altitudes, are expected to expand their geographical range, in response to temperature rise, towards higher altitudes of Sub-Saharan Africa and might become more threatening. This could be the same speculation for other plant-parasitic nematodes, especially those known to affect grain crops like upland rice [127].
Precipitation patterns play a significant role in the emergence and establishment of various cereal pathogens. Higher humidity levels are expected to encourage increasing cases of the mycotoxin deoxynivalenol by F. graminearum in almost every grain especially maize which accounts for nearly 50% of cereals produced within the African continent [146]. This is a similar expectation with aflatoxin-producing Aspergillus flavus and fumonisin-producing F. verticillioides if temperature rise, drought and insect infestations continue to spike upwards in subtropical and tropical parts of Africa [149, 158]. As a result, more economic losses and reduced food quality due to mycotoxin contaminations are predicted to occur more often if the current climate trends continue in this century [159]. Conditions of low precipitation (drought) were also found to increase the aggressiveness of Magnaporthe oryzae infections leading to greater pathogen populations and more obvious disease symptoms [160]. On the other hand, precipitation changes have the greatest influence on plant-parasitic nematodes. Increases in rainfall have been found to increase the abundance and diversity of M. minor, Heterodera spp. and Pratylenchus spp. in grasslands and cereal cropping systems of Northern Ireland. Current climate change scenarios might force a similar observation in major cereal-producing regions of Africa especially those already predisposed to high nematode infestations [123].
Levels of atmospheric CO2 are another important climatic factor influencing how much damage is done by plant pathogens. Elevated CO2 levels, independent of warming temperatures, have been found to increase the susceptibility of many cereals to attack from Fusarium head blight fungi which threaten wheat and barley crops in Africa [161, 162]. A weather-based model developed using data from the United Kingdom has suggested a slight increase in FHB severity as a direct effect of climate change [163]. Under elevated CO2, plant defence pathways of wheat (regardless of the resistant and susceptible genotypes tested) were drastically reduced leading to an enhanced susceptibility to F. graminearum which causes the Fusarium head blight. This might become the eventual scenario in wheat-producing regions of Africa such as Tunisia if increased industrialization facilitates high CO2 concentrations in the atmosphere [146]. On the other hand, under elevated CO2 conditions (700 ppm), the rice nematode M. graminicola induced greater damage in rice roots compared to ambient CO2 level (400 ppm) [147]. Likewise, elevated CO2 seem to aggravate root galling through M. incognita by repressing the jasmonic acid (JA)-mediated defence signalling pathway in infected tomato [164]. In general, however, concentrations of CO2 in the atmosphere don’t seem to impact the population and virulence of plant parasitic nematodes. Although most studies predict increased virulence of plant pathogens, it’s noteworthy to say that elevated levels of CO2 have been shown to improve the photosynthetic ability of C3 plants like rice, maize and wheat [165, 166]. There could likely be a cross-talk of increased pathogen virulence against more healthy crops leading to lesser losses or otherwise. More research should be done to investigate such interactions and predict potential outcomes of increased CO2 on pathogen virulence and plant physiology, specifically focused on cereal crops.
An important simulation study by spatial modelling has predicted an increasing incidence of bacterial leaf blight caused by Xanthomonas oryzae pv. oryzae, correlating with lesser losses. In contrast, under climate change scenarios, losses due to rice leaf blast are predicted to decline in Tanzania even though a global increase is expected. Fewer leaf blast epidemics across the entire country with a slight increase in temperature by 1.17 °C were predicted to occur in the future [167]. These observations correlated with a process-based model study in Italy which showed that an increase in precipitation (a growing reality in several African climates) is positively correlated with leaf blast severity, panicle blast incidence and hence a high disease impact [168]. At the same time, all smut-disease-causing pathogens of maize (Sporisorium holci-sorghi, Sphacelotheca reiliana and Ustilago maydis) thrive well under warm and dry conditions which is characteristic of many grain-producing regions in Sub-Saharan Africa. If their temperature thresholds are not exceeded, all smut diseases in maize are speculated to increase under current warming climates, independently of geographical locations [149].
While these simulations provide potential insight into the prediction of future climate trends, an accurate prediction of natural events is still glim. Extrapolations based on these simulations result in expectations that can be far from exactly what eventually happens. Current undulating and yet sharp changes in microclimate across the globe testify to unexpected changes in climatic conditions. However, the adoption of disease monitoring programs and alarm systems, which run based on accurate long-term weather data in combination with disease models, is gradually gaining prominence but still requires robust technological improvement, possibly with the inclusion of artificial intelligence. Unfortunately, disease projection and simulation studies that provide predictions on the futuristic impact of these current cereal pathogens have not been studied adequately in Africa. A limited focus has been given to envisaging the potential spread of these pathogens in correlation with currently occurring weather patterns across different climatic belts in Africa. It is imperative to carry out such studies in a bid to understand the trends of climatic change events in major producing regions of Africa and their corresponding influence on the pathogenicity and severity of each of the cereal pathogens considered in this review. Findings from these predictive studies will provide insights into the best-integrated disease management approach to uniquely adopt against the impact of these plant pathogens on cereal production within the African landscape.
8 Conclusion
Climate change events such as alternating temperatures, changing precipitation patterns and elevated CO2 levels can have direct or indirect positive or negative effects on the impacts of plant pathogens and the crops themselves well documented to attack cultivated cereal crops in Africa. While warmer and more humid weather with stronger winds will cause some of these pathogens such as Magnaporthe oryzae, Fusarium graminearum, and Xanthomonas oryzae pv. oryzae and Meloidogyne spp to become more widespread and more dangerous on host cereal crops, extremely hot and dry climates might reduce the impact of some diseases (such as rice blasts) while increasing that of others (like stalk rots on maize caused by heat and water stress). At the same, the host crops themselves will be affected positively or negatively by climate change events. In the event of more favourable weather conditions, most of these pathogens will ravage several cereal crops and have a devastating effect on the food security of various African countries. Like many developed countries, many cereal-producing African countries need to prioritize and establish, first, disease monitoring programs coupled with disease-warning signalling systems which use climate-based simulations and predictions. It is very important to encourage and emphasize the need for more simulation studies to predict the potential impact of these pathogens across all cereal-producing regions of Africa under current and future climate change scenarios.
Data availability
Not applicable.
Code availability
Not applicable.
References
Griffiths JF. Africa: climate of. In: Oliver JE, editor. Encyclopedia of world climatology. Springer: Netherlands; 2005. p. 6–14. https://doi.org/10.1007/1-4020-3266-8_4.
Sosef MSM, Dauby G, Blach-Overgaard A, Van Der Burgt X, Catarino L, Damen T, et al. Exploring the floristic diversity of tropical Africa. BMC Biol. 2017. https://doi.org/10.1186/s12915-017-0356-8.
Tak Y, Kumar M. Phenolics: a key defence secondary metabolite to counter biotic stress. In: Lone R, Shuab R, Kamili AN, editors. Plant phenolics in sustainable agriculture. Singapore: Springer Singapore; 2020. p. 309–29. https://doi.org/10.1007/978-981-15-4890-1_13.
Pallas V, García JA. How do plant viruses induce disease? Interactions and interference with host components. J Gen Virol. 2011;92:2691–705. https://doi.org/10.1099/vir.0.034603-0.
Pfeilmeier S, Caly DL, Malone JG. Bacterial pathogenesis of plants: future challenges from a microbial perspective: challenges in bacterial molecular plant pathology. Mol Plant Pathol. 2016;17:1298–313. https://doi.org/10.1111/mpp.12427.
Doehlemann G, Ökmen B, Zhu W, Sharon A. Plant pathogenic fungi. Microbiol Spectr. 2017. https://doi.org/10.1128/microbiolspec.FUNK-0023-2016.
Osman HA, Ameen HH, Mohamed M, Elkelany US. Efficacy of integrated microorganisms in controlling root-knot nematode Meloidogyne javanica infecting peanut plants under field conditions. Bull Natl Res Cent. 2020. https://doi.org/10.1186/s42269-020-00366-0.
Kumar S, Bhowmick MK, Ray P. Weeds as alternate and alternative hosts of crop pests. Indian J Weed Sci. 2021;53:14–29.
Lambers H, Oliveira RS. Biotic influences: parasitic associations. In: Plant physiological ecology. Cham: Springer International Publishing; 2019. p. 597–613. https://doi.org/10.1007/978-3-030-29639-1_15.
Sardana V, Mahajan G, Jabran K, Chauhan BS. Role of competition in managing weeds: an introduction to the special issue. Crop Prot. 2017;95:1–7.
FAO. FAO-News Article: Climate change fans spread of pests and threatens plants and crops, new FAO study. 2021. Available from: https://www.fao.org/news/story/en/item/1402920/icode/.
Liliane T, Charles M. Factors affecting yield of crops. In: Amanullah, editor. Agron-Clim Change Food Secur. IntechOpen; 2020. Available from: https://www.intechopen.com/books/agronomy-climate-change-food-security/factors-affecting-yield-of-crops.
FAOSTAT. Cereal production. 2021. Available from: https://www.fao.org/faostat/en/#data/QCL.
Fernandesa C, Sonawaneb S, Arya SS. Cereal based functional beverages: a review. J Microbiol Biotechnol Food Sci. 2018;8:914–9.
Erenstein O, Poole N, Donovan J. Role of staple cereals in human nutrition: separating the wheat from the chaff in the infodemics age. Trends Food Sci Technol. 2022;119:508–13.
Galani YJH, Orfila C, Gong YY. A review of micronutrient deficiencies and analysis of maize contribution to nutrient requirements of women and children in Eastern and Southern Africa. Crit Rev Food Sci Nutr. 2022;62:1568–91. https://doi.org/10.1080/10408398.2020.1844636.
Ekpa O, Palacios-Rojas N, Kruseman G, Fogliano V, Linnemann AR. Sub-Saharan African maize-based foods—processing practices, challenges and opportunities. Food Rev Int. 2019;35:609–39. https://doi.org/10.1080/87559129.2019.1588290.
Dabija A, Ciocan ME, Chetrariu A, Codină GG. Maize and sorghum as raw materials for brewing, a review. Appl Sci. 2021;11:3139.
Mwadalu R, Mwangi M. The potential role of sorghum in enhancing food security in semi-arid eastern Kenya: a review. J Appl Biosci. 2013;71:5786.
Poutanen KS, Kårlund AO, Gómez-Gallego C, Johansson DP, Scheers NM, Marklinder IM, et al. Grains—a major source of sustainable protein for health. Nutr Rev. 2022;80:1648–63.
Sawadogo-Lingani H, Owusu-Kwarteng J, Glover R, Diawara B, Jakobsen M, Jespersen L. Sustainable production of African traditional beers with focus on dolo, a west african sorghum-based alcoholic beverage. Front Sustain Food Syst. 2021;5: 672410. https://doi.org/10.3389/fsufs.2021.672410/full.
Osnato M. Evolution of flowering time genes in rice: from the paleolithic to the anthropocene. Plant Cell Environ. 2023;46:1046–59. https://doi.org/10.1111/pce.14495.
Tadesse W, Bishaw Z, Assefa S. Wheat production and breeding in Sub-Saharan Africa: challenges and opportunities in the face of climate change. Int J Clim Change Strateg Manag. 2019;11:696–715. https://doi.org/10.1108/IJCCSM-02-2018-0015/full/html.
Tricase C, Amicarelli V, Lamonaca E, Leonardo Rana R. Economic analysis of the barley market and related uses. In: Tadele Z, editor. Grasses food feed. IntechOpen; 2018. Available from: https://www.intechopen.com/books/grasses-as-food-and-feed/economic-analysis-of-the-barley-market-and-related-uses.
Mesterházy Á, Oláh J, Popp J. Losses in the grain supply chain: causes and solutions. Sustainability. 2020;12:2342.
Abbass K, Qasim MZ, Song H, Murshed M, Mahmood H, Younis I. A review of the global climate change impacts, adaptation, and sustainable mitigation measures. Environ Sci Pollut Res. 2022;29:42539–59.
Kwiecien O, Braun T, Brunello CF, Faulkner P, Hausmann N, Helle G, et al. What we talk about when we talk about seasonality—a transdisciplinary review. Earth-Sci Rev. 2022;225: 103843.
Dean R, Van Kan JAL, Pretorius ZA, Hammond-Kosack KE, Di Pietro A, Spanu PD, et al. The Top 10 fungal pathogens in molecular plant pathology: top 10 fungal pathogens. Mol Plant Pathol. 2012;13:414–30. https://doi.org/10.1111/j.1364-3703.2011.00783.x.
Różewicz M, Wyzińska M, Grabiński J. The most important fungal diseases of cereals—problems and possible solutions. Agronomy. 2021;11:714.
Kagot V, Okoth S, De Boevre M, De Saeger S. Biocontrol of Aspergillus and Fusarium mycotoxins in Africa: benefits and limitations. Toxins. 2019;11:109.
Abdallah-Nekache N, Laraba I, Ducos C, Barreau C, Bouznad Z, Boureghda H. Occurrence of Fusarium head blight and Fusarium crown rot in Algerian wheat: identification of associated species and assessment of aggressiveness. Eur J Plant Pathol. 2019;154:499–512. https://doi.org/10.1007/s10658-019-01673-7.
Ouko A, Okoth S, Netshifhefhe NEL, Viljoen A, Rose LJ. Tolerance to Fusarium verticillioides infection and fumonisin accumulation in maize F1 hybrids and subsequent F2 populations. Agron J. 2020;112:2432–44. https://doi.org/10.1002/agj2.20145.
Mitchell NJ, Bowers E, Hurburgh C, Wu F. Potential economic losses to the US corn industry from aflatoxin contamination. Food Addit Contam Part A. 2016;33:540–50. https://doi.org/10.1080/19440049.2016.1138545.
Chung H, Goh J, Han S-S, Roh J-H, Kim Y, Heu S, et al. Comparative pathogenicity and host ranges of Magnaporthe oryzae and related species. Plant Pathol J. 2020;36:305–13. https://doi.org/10.5423/PPJ.FT.04.2020.0068.
Onaga G, Suktrakul W, Wanjiku M, Quibod IL, Entfellner J-BD, Bigirimana J, et al. Magnaporthe oryzae populations in Sub-Saharan Africa are diverse and show signs of local adaptation. Genomics. 2020. https://doi.org/10.1101/2020.11.17.377325.
Margaret O, Wycliffe WL, Agnes K, Cecilia M, Matthew N, Fredah M, et al. Occurrence, distribution and severity of finger millet blast caused by Magnaporthe oryzae in Kenya. Afr J Plant Sci. 2020;14:139–49.
Meisel B, Korsman J, Kloppers FJ, Berger DK. Cercospora zeina is the causal agent of grey leaf spot disease of maize in southern Africa. Eur J Plant Pathol. 2009;124:577–83. https://doi.org/10.1007/s10658-009-9443-1.
Kibe M, Nair SK, Das B, Bright JM, Makumbi D, Kinyua J, et al. Genetic dissection of resistance to gray leaf spot by combining genome-wide association, linkage mapping, and genomic prediction in tropical maize germplasm. Front Plant Sci. 2020;11: 572027. https://doi.org/10.3389/fpls.2020.572027/full.
Pažoutová S, Frederickson DE. Genetic diversity of Claviceps africana on sorghum and Hyparrhenia. Plant Pathol. 2005;54:749–63. https://doi.org/10.1111/j.1365-3059.2005.01255.x.
Miedaner T, Geiger H. Biology, genetics, and management of Ergot (Claviceps spp.) in Rye, Sorghum, and Pearl Millet. Toxins. 2015;7:659–78.
Prom LK, Cisse N, Ndoye O. Assessing the vulnerability of selected sorghum lines from the United States of America to long smut (Sporisorium ehrenbergii Vánky) disease. Crop Prot. 2007;26:1771–6.
Prom LK, Perumal R, Cissé N, Little CR. Evaluation of selected Sorghum lines and hybrids for resistance to grain mold and long smut fungi in Senegal, West Africa. Plant Health Prog. 2014;15:74–7. https://doi.org/10.1094/PHP-RS-13-0128.
Prins R, Dreisigacker S, Pretorius Z, Van Schalkwyk H, Wessels E, Smit C, et al. Stem rust resistance in a geographically diverse collection of spring wheat lines collected from across Africa. Front Plant Sci. 2016;7. https://doi.org/10.3389/fpls.2016.00973/abstract.
Soko T, Bender CM, Prins R, Pretorius ZA. Yield loss associated with different levels of stem rust resistance in bread wheat. Plant Dis. 2018;102:2531–8. https://doi.org/10.1094/PDIS-02-18-0307-RE.
Kayim M, Nawaz H, Alsalmo A. Fungal diseases of wheat. In: Ansari M-R, editor. Wheat. IntechOpen; 2022. Available from: https://www.intechopen.com/chapters/80489.
Ji F, He D, Olaniran AO, Mokoena MP, Xu J, Shi J. Occurrence, toxicity, production and detection of Fusarium mycotoxin: a review. Food Prod Process Nutr. 2019;1:6. https://doi.org/10.1186/s43014-019-0007-2.
Muimba-Kankolongo A. Cereal production. In: Food crop production by smallholder farmers Southern Africa. Amsterdam: Elsevier; 2018. p. 73–121.
Zhu Z, Hao Y, Mergoum M, Bai G, Humphreys G, Cloutier S, et al. Breeding wheat for resistance to Fusarium head blight in the Global North: China, USA, and Canada. Crop J. 2019;7:730–8.
Gai X, Dong H, Wang S, Liu B, Zhang Z, Li X, et al. Infection cycle of maize stalk rot and ear rot caused by Fusarium verticillioides. PLoS ONE. 2018;13:e0201588. https://doi.org/10.1371/journal.pone.0201588.
Beukes I, Rose LJ, van Coller GJ, Viljoen A. Disease development and mycotoxin production by the Fusarium graminearum species complex associated with South African maize and wheat. Eur J Plant Pathol. 2018;150:893–910. https://doi.org/10.1007/s10658-017-1331-5.
Tsehaye H, Brurberg MB, Sundheim L, Assefa D, Tronsmo A, Tronsmo AM. Natural occurrence of Fusarium species and fumonisin on maize grains in Ethiopia. Eur J Plant Pathol. 2017;147:141–55. https://doi.org/10.1007/s10658-016-0987-6.
Baghbani F, Lotfi R, Moharramnejad S, Bandehagh A, Roostaei M, Rastogi A, et al. Impact of Fusarium verticillioides on chlorophyll fluorescence parameters of two maize lines. Eur J Plant Pathol. 2019;154:337–46. https://doi.org/10.1007/s10658-018-01659-x.
Reed H, Mueller B, Groves CL, Smith DL. Presence and correlation of Fusarium graminearum and deoxynivalenol accumulation in silage corn plant parts. Plant Dis. 2022;106:87–92. https://doi.org/10.1094/PDIS-03-21-0641-RE.
Beccari G, Colasante V, Tini F, Senatore MT, Prodi A, Sulyok M, et al. Causal agents of Fusarium head blight of durum wheat (Triticum durum Desf.) in central Italy and their in vitro biosynthesis of secondary metabolites. Food Microbiol. 2018;70:17–27.
Schmale DG, Bergstrom GC. Fusarium head blight (FHB) or scab. APSnet Feature Artic. 2003; Available from: https://www.apsnet.org/edcenter/disandpath/fungalasco/pdlessons/Pages/Fusarium.aspx.
Chen Y, Kistler HC, Ma Z. Fusarium graminearum trichothecene mycotoxins: biosynthesis, regulation, and management. Annu Rev Phytopathol. 2019;57:15–39. https://doi.org/10.1146/annurev-phyto-082718-100318.
Adejumo TO, Hettwer U, Karlovsky P. Survey of maize from south-western Nigeria for zearalenone, α- and β-zearalenols, fumonisin B1 and enniatins produced by Fusarium species. Food Addit Contaminant. 2007;24:993–1000. https://doi.org/10.1080/02652030701317285.
Olopade BK, Oranusi SU, Nwinyi OC, Gbashi S, Njobeh PB. Occurrences of Deoxynivalenol, Zearalenone and some of their masked forms in selected cereals from Southwest Nigeria. NFS J. 2021;23:24–9.
Bensassi F, Zaied C, Abid S, Hajlaoui MR, Bacha H. Occurrence of deoxynivalenol in durum wheat in Tunisia. Food Control. 2010;21:281–5.
Minnaar-Ontong A, Herselman L, Kriel W-M, Leslie JF. Morphological characterization and trichothecene genotype analysis of a Fusarium Head Blight population in South Africa. Eur J Plant Pathol. 2017;148:261–9. https://doi.org/10.1007/s10658-016-1085-5.
Oren L, Ezrati S, Cohen D, Sharon A. Early events in the Fusarium verticillioides—maize interaction characterized by using a green fluorescent protein-expressing transgenic isolate. Appl Environ Microbiol. 2003;69:1695–701. https://doi.org/10.1128/AEM.69.3.1695-1701.2003.
Tiru Z, Mandal P, Pratim Chakraborty A, Pal A, Sadhukhan S. Fusarium Disease of Maize and Its Management through Sustainable Approach. In: Mahyar Mirmajlessi S, editor. Fusarium—Overv Genus. IntechOpen; 2022. Available from: https://www.intechopen.com/chapters/79705.
Mukanga M, Derera J, Tongoona P, Laing MD. A survey of pre-harvest ear rot diseases of maize and associated mycotoxins in south and central Zambia. Int J Food Microbiol. 2010;141:213–21.
Schoeman A, Flett BC, Janse Van Rensburg B, Ncube E, Viljoen A. Pathogenicity and toxigenicity of Fusarium verticillioides isolates collected from maize roots, stems and ears in South Africa. Eur J Plant Pathol. 2018;152:677–89. https://doi.org/10.1007/s10658-018-1510-z.
Yli-Mattila T, Sundheim L. Fumonisins in African countries. Toxins. 2022;14:419.
Kedera CJ, Plattner RD, Desjardins AE. Incidence of Fusarium spp. and levels of Fumonisin B1 in maize in Western Kenya. Appl Environ Microbiol. 1999;65:41–4. https://doi.org/10.1128/AEM.65.1.41-44.1999.
Stasiewicz MJ, Falade TDO, Mutuma M, Mutiga SK, Harvey JJW, Fox G, et al. Multi-spectral kernel sorting to reduce aflatoxins and fumonisins in Kenyan maize. Food Control. 2017;78:203–14.
Kamala A, Ortiz J, Kimanya M, Haesaert G, Donoso S, Tiisekwa B, et al. Multiple mycotoxin co-occurrence in maize grown in three agro-ecological zones of Tanzania. Food Control. 2015;54:208–15.
Mutiga SK, Rotich F, Ganeshan VD, Mwongera DT, Mgonja EM, Wre VM, et al. Assessment of the virulence spectrum and its association with genetic diversity in Magnaporthe oryzae populations from Sub-Saharan Africa. Phytopathology. 2017;107:852–63. https://doi.org/10.1094/PHYTO-08-16-0319-R.
Te Beest, Guerber C, Ditmore M. Rice Blast. Plant Health Instr. 2007; Available from: https://www.apsnet.org/edcenter/disandpath/fungalasco/pdlessons/Pages/RiceBlast.aspx.
Kihoro J, Bosco NJ, Murage H, Ateka E, Makihara D. Investigating the impact of rice blast disease on the livelihood of the local farmers in greater Mwea region of Kenya. Springerplus. 2013;2:308. https://doi.org/10.1186/2193-1801-2-308.
Tembo B, Mulenga RM, Sichilima S, M’siska KK, Mwale M, Chikoti PC, et al. Detection and characterization of fungus (Magnaporthe oryzae pathotype triticum) causing wheat blast disease on rain-fed grown wheat (Triticum aestivum L.) in Zambia. PLoS ONE. 2020;15:e0238724. https://doi.org/10.1371/journal.pone.0238724.
Olufolaji DB. Incidence and preliminary control of blast disease of rice in Southwest Nigeria. In: Kharwar RN, Upadhyay RS, Dubey NK, Raghuwanshi R, editors. Microbial diversity and biotechnology in food security. Springer India: New Delhi; 2014. p. 235–42. https://doi.org/10.1007/978-81-322-1801-2_20.
Merkevičiūte-Venslovė L, Venslovas E, Mankevičienė A, Šlepetienė A, Cesevičienė J. Effect of Ustilago maydis on the nutritive value and aerobic deterioration of maize silage. Agronomy. 2022;13:111.
Zhao Y, Agrawal N, Ghareeb H, Habib MT, Dickmeis S, Schwachtje J, et al. The Sporisorium reilianum effector Vag2 promotes head smut disease via suppression of plant defense responses. J Fungi. 2022;8:498.
Nagaraja A, Chethana BS, Jain AK. Biotic stresses and their management. In: Millets and pseudo cereals. Amsterdam: Elsevier; 2021. p. 119–42.
Abebe Teferi T. Prevalence and intensity of economically important fungal diseases of Sorghum in South Tigray, Ethiopia. J Plant Sci Publ Group. 2015;3:92. https://doi.org/10.11648/j.jps.20150302.18.
Bhuiyan SA, Galea VJ, Ryley MJ, Tay D, Lisle AT. Survival of conidia of sorghum ergot (caused by Claviceps africana) on panicles, seed and soil in Australia. Australas Plant Pathol. 2002;31:137. https://doi.org/10.1071/AP01077.
Crous PW, Groenewald JZ, Groenewald M, Caldwell P, Braun U, Harrington TC. Species of Cercospora associated with grey leaf spot of maize. Stud Mycol. 2006;55:189–97. https://doi.org/10.3114/sim.55.1.189.
Duan C, Zhao L, Wang J, Liu Q, Yang Z, Wang X. Dispersal routes of Cercospora zeina causing maize gray leaf spot in China. J Integr Agric. 2022;21:2943–56.
Ward JMJ, Stromberg EL, Nowell DC, Nutter FW. Gray leaf Spot: a disease of global importance in maize production. Plant Dis. 1999;83:884–95. https://doi.org/10.1094/PDIS.1999.83.10.884.
Abdelkhalek A, Hafez E. Plant viral diseases in Egypt and their control. In: El-Wakeil N, Saleh M, Abu-hashim M, editors. Cottage industry of biocontrol agents and their applications. Cham: Springer International Publishing; 2020. p. 403–21. https://doi.org/10.1007/978-3-030-33161-0_13.
Manjunatha L, Rajashekara H, Uppala LS, Ambika DS, Patil B, Shankarappa KS, et al. Mechanisms of microbial plant protection and control of plant viruses. Plants. 2022;11:3449.
Jones AC. Global plant virus disease pandemics and epidemics. Plants. 2021;10:233.
Dietzgen R, Mann K, Johnson K. Plant virus-insect vector interactions: current and potential future research directions. Viruses. 2016;8:303.
Redinbaugh MG, Stewart LR. Maize lethal necrosis: an emerging, synergistic viral disease. Annu Rev Virol. 2018;5:301–22. https://doi.org/10.1146/annurev-virology-092917-043413.
Kreuze JF, Valkonen JP. Utilization of engineered resistance to viruses in crops of the developing world, with emphasis on sub-Saharan Africa. Curr Opin Virol. 2017;26:90–7.
Rao GP, Reddy MG. Overview of yield losses due to plant viruses. In: Applied plant virology. Amsterdam: Elsevier; 2020. p. 531–62.
Rakotondrafara AM, Byamukama E, Plumb RT. Virus diseases of cereals. 1st ed. USA: Wiley; 2017. p. 1–12. https://doi.org/10.1002/9780470015902.a0000762.pub3.
Sekiya N, Nakajima T, Oizumi N, Kurosawa C, Tibanyendela N, Peter MA, et al. Agronomic practices preventing local outbreaks of rice yellow mottle virus disease revealed by spatial autoregressive analysis. Agron Sustain Dev. 2022;42:15. https://doi.org/10.1007/s13593-022-00757-9.
Kija ED, Mziray MI. Resistant rice against rice yellow mottle virus: Rice yellow mottle virus & RYMV. PlantwisePlus Knowl Bank. 2013; 20147801019. https://doi.org/10.1079/pwkb.20147801019.
Magenya OEV, Mueke J, Omwega C. Significance and transmission of maize streak virus disease in Africa and options for management: a review. Afr J Biotechnol. 2008;7:4897–910.
Karavina C. Maize streak virus: a review of pathogen occurrence, biology and management options for smallholder farmers. Afr J Agric Res. 2014;9:2736–42.
Mbong GA, Suh C, Djomo Sime H, Anoumaa M, Fonkou T, Chimi Nkombo LL, et al. Phytosanitary situation of maize streak; in the main maize production zones of Cameroon. Agric Sci. 2021;12:339–53. https://doi.org/10.4236/as.2021.124022.
Signoret PA. Cereal viruses: maize/corn. In: Encyclopedia of virology. Amsterdam: Elsevier; 2008. p. 475–82.
Biswal AK, Alakonya AE, Mottaleb KA, Hearne SJ, Sonder K, Molnar TL, et al. Maize Lethal Necrosis Disease: review of molecular and genetic resistance mechanisms, socio-economic impacts, and mitigation strategies in sub-Saharan Africa. BMC Plant Biol. 2022;22:542. https://doi.org/10.1186/s12870-022-03932-y.
Abraha NW. Surveillance of Barley Yellow Dwarf Virus (BYDV) in all barley growing areas of Ethiopia. South Asian Res J Biol Appl Biosci. 2020;2:46–53.
Zhan B, Yang X, Lommel SA, Zhou X. Recent progress in maize lethal necrosis disease: from pathogens to integrated pest management. J Integr Agric. 2022;21:3445–55.
Odongo PJ, Onaga G, Ricardo O, Natsuaki KT, Alicai T, Geuten K. Insights into natural genetic resistance to rice yellow mottle virus and implications on breeding for durable resistance. Front Plant Sci. 2021;12:671355. https://doi.org/10.3389/fpls.2021.671355/full.
Allarangay MD, Traore O, Traore EVS, Millogo RJ, Guinko S, Konate G. Host Range of Rice yellow mottle virus in Sudano-Sahelian Savannahs. Pak J Biol Sci. 2007;10:1414–21.
Traoré V, Néya BJ, Camara M, Gracen V, Offei SK, Traoré O. Farmers’ perception and impact of rice yellow mottle disease on rice yields in Burkina Faso. Agric Sci. 2015;06:943–52. https://doi.org/10.4236/as.2015.69091.
Martin DP, Shepherd DN, Rybicki EP. Maize streak virus. In: Encyclopedia of virology. Amsterdam: Elsevier; 2008. p. 263–72.
Osei MK, Adjebeng-Danquah J, Bediako KA, Melomey LD, Agyare RY, Annor B, et al. Origin, evolution and bottlenecks of geminiviruses. In: Geminivirus: detection, diagnosis and management. Amsterdam: Elsevier; 2022. p. 79–93.
Monjane AL, Harkins GW, Martin DP, Lemey P, Lefeuvre P, Shepherd DN, et al. Reconstructing the history of maize streak virus strain—a dispersal to reveal diversification hot spots and its origin in Southern Africa. J Virol. 2011;85:9623–36. https://doi.org/10.1128/JVI.00640-11.
Martin DP, Monjane AL. Maize streak virus (Geminiviridae). In: Encyclopedia of virology. Amsterdam: Elsevier; 2021. p. 461–9.
Aguilar-Marcelino L, Mendoza-de-Gives P, Al-Ani LKT, López-Arellano ME, Gómez-Rodríguez O, Villar-Luna E, et al. Using molecular techniques applied to beneficial microorganisms as biotechnological tools for controlling agricultural plant pathogens and pest. In: Molecular aspects of plant beneficial microbes in agriculture. Amsterdam: Elsevier; 2020. p. 333–49.
Tampakaki AP, Hatziloukas E, Panopoulos NJ. Plant pathogens, bacterial. In: Encyclopedia of microbiology. Amsterdam: Elsevier; 2009. p. 655–77.
Pruvost O, Boyer K, Ravigné V, Richard D, Vernière C. Deciphering how plant pathogenic bacteria disperse and meet: molecular epidemiology of Xanthomonas citri pv. citri at microgeographic scales in a tropical area of Asiatic citrus canker endemicity. Evol Appl. 2019;12:1523–38. https://doi.org/10.1111/eva.12788.
Sharma A, Gautam S, Wadhawan S. Xanthomonas. In: Encyclopedia of food microbiology. Amsterdam: Elsevier; 2014. p. 811–7.
Verdier V, Vera Cruz C, Leach JE. Controlling rice bacterial blight in Africa: needs and prospects. J Biotechnol. 2012;159:320–8.
Health (PLH) EP on P, Jeger M, Candresse T, Chatzivassiliou E, Dehnen-Schmutz K, Gilioli G, et al. Pest categorisation of Xanthomonas oryzae pathovars oryzae and oryzicola. EFSA J. 2018;16:e05109. https://doi.org/10.2903/j.efsa.2018.5109.
Triplett L, Patel R. Xanthomonas vasicola pv. vasculorum (bacterial leaf streak of corn). CABI Compend. 2022; CABI Compendium:36777909. https://doi.org/10.1079/cabicompendium.36777909.
Osdaghi E. Xanthomonas translucens pv. undulosa (bacterial leaf streak of wheat and barley). CABI Compend. 2022;CABI Compendium:56979. https://doi.org/10.1079/cabicompendium.56979
T. Tambong J. Bacterial pathogens of wheat: symptoms, distribution, identification, and taxonomy. In: Ansari M-R, editor. Wheat. IntechOpen; 2022. Available from: https://www.intechopen.com/chapters/80665.
Ke Y, Hui S, Yuan M. Xanthomonas oryzae pv. oryzae inoculation and growth rate on rice by leaf clipping method. BIO-Protocol. 2017;7. Available from: https://bio-protocol.org/e2568.
Mansfield J, Genin S, Magori S, Citovsky V, Sriariyanum M, Ronald P, et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol Plant Pathol. 2012;13:614–29. https://doi.org/10.1111/j.1364-3703.2012.00804.x.
Saha S, Garg R, Biswas A, Rai A. Bacterial diseases of rice: an overview. J Pure Appl Microbiol. 2015;9:725–36.
Niño-Liu DO, Ronald PC, Bogdanove AJ. Xanthomonas oryzae pathovars: model pathogens of a model crop. Mol Plant Pathol. 2006;7:303–24. https://doi.org/10.1111/j.1364-3703.2006.00344.x.
Chu Z, Fu B, Yang H, Xu C, Li Z, Sanchez A, et al. Targeting xa13, a recessive gene for bacterial blight resistance in rice. Theor Appl Genet. 2006;112:455–61. https://doi.org/10.1007/s00122-005-0145-6.
Ledman KE, Curland RD, Ishimaru CA, Dill-Macky R. Xanthomonas translucens pv. undulosa identified on common weedy grasses in naturally infected wheat fields in Minnesota. Phytopathology. 2021;111:1114–21. https://doi.org/10.1094/PHYTO-08-20-0337-R.
Dababat AA, Imren M, Pridannikov MV, Özer G, et al. Plant-parasitic nematodes on cereals in northern Kazakhstan. J Plant Dis Prot. 2020;127:641–9.
Keçici Aİ, Bozbuğa R, Öcal A, Yüksel E, Özer G, Yildiz Ş, et al. Diversity and identification of plant–parasitic nematodes in wheat-growing ecosystems. Microorganisms. 2022;10:1534.
Fleming T, McGowan NE, Maule AG, Fleming CC. Prevalence and diversity of plant parasitic nematodes in Northern Ireland grassland and cereals, and the influence of soils and rainfall. Plant Pathol. 2016;65:1539–50.
Abdulsalam S, Shaibu AS, Ojonuba JM, Peng H, Peng D. Nematode pests of some major cereals in Nigeria: need for integration of morphological/morphometrical, biochemical, and molecular diagnostic approaches for accurate identification. Plant Pathol. 2021;70:1535–51. https://doi.org/10.1111/ppa.13412.
Aghale DN, Egbucha KC, Umeh OJ. Plant parasitic nematode threat to Nigerian agricultural food security. Int J Adv Res Bot. 2017;3. Available from: https://www.arcjournals.org/pdfs/ijarb/v3-i3/3.pdf.
Namouchi-Kachouri N, B’Chir MM, Hajji A, Riley IT, Nicol JM, Dababat AA. Global importance of the main nematodes associated with cereals in Tunisia. 2009;41–4.
Coyne DL, Cortada L, Dalzell JJ, Claudius-Cole AO, Haukeland S, Luambano N, et al. Plant–parasitic nematodes and food security in Sub-Saharan Africa. Annu Rev Phytopathol. 2018;56:381–403. https://doi.org/10.1146/annurev-phyto-080417-045833.
McDonald AH, Nicol JM. Nematode parasites of cereals. In: Luc M, Sikora RA, Bridge J, editors. Plant parasitic nematodes subtrop trop agric. 2nd ed. UK: CABI Publishing; 2005. p. 131–91. https://doi.org/10.1079/9780851997278.0131.
Mc Donald AH, Waele DD, Fourie H. Nematode pests of maize and other cereal crops. Nematol South Afr View 21st Century. 2017;183–99.
Bridge J, Plowright RA, Peng DeLiang PD. Nematode parasites of rice. In: Luc M, Sikora RA, Bridge J, editors. Plant Parasitic Nematodes Subtrop Trop Agric. 2nd ed. UK: CABI Publishing; 2005. p. 87–130. https://doi.org/10.1079/9780851997278.0087.
Bélair G, Mimee B, Duceppe MO, Miller S. First report of the root-lesion nematode, Pratylenchus alleni associated with Damage on Soybean in Quebec, Canada. Plant Dis. 2013;97:292–292. https://doi.org/10.1094/PDIS-03-12-0309-PDN.
Nguyen HT, Trinh QP, Couvreur M, Singh PR, Decraemer W, Bert W. Molecular and morphological characterisation of a new root-lesion nematode, Pratylenchus horti n. sp. (Tylenchomorpha: Pratylenchidae), from Ghent University Botanical Garden. Nematology. 2019;21:739–52.
Jones MP, Jones MGK, Fosu-Nyarko J. Molecular biology of root lesion nematodes (Pratylenchus spp.) and their interaction with host plants. Ann Appl Biol. 2014;164:163–81.
Jones JT. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol Plant Pathol. 2013;14:946–61.
da Silva MP, Tylka GL, Munkvold GP. Seed treatment effects on maize seedlings coinfected with Rhizoctonia solani and Pratylenchus penetrans. Plant Dis. 2017;101:957–63. https://doi.org/10.1094/PDIS-10-16-1417-RE.
Kagoda F, Derera J, Tongoona P, Coyne DL, Talwana HL. Grain yield and heterosis of maize hybrids under nematode infested and nematicide treated conditions. J Nematol 2011;209–19.
Mokrini F, Moens M, Viaene N. Root-lesion and cyst nematodes in cereal fields in Morocco: species identification, population diversity and crop resistance. 2016. https://doi.org/10.13140/RG.2.2.30262.29769.
Ibrahim HMM, Ahmad EM, Martínez-Medina A, Aly MAM. Effective approaches to study the plant-root knot nematode interaction. Plant Physiol Biochem. 2019;141:332–42.
Ribeiro Costa CDS, Silva Costa AE da, Dos Santos AMM, Lima Pereira JW de, E Carvalho RR da C, Carvalho Filho JLS de. Current status of the occurrence and reaction root-knot nematodes in the main botanical families of medicinal plants. J Exp Agric Int. 2019;1–21. Available from: http://www.journaljeai.com/index.php/JEAI/article/view/30096.
Abd-Elgawad MMM. Optimizing biological control agents for controlling nematodes of tomato in Egypt. Egypt J Biol Pest Control. 2020;30:58. https://doi.org/10.1186/s41938-020-00252-x.
Tiwari S, Eisenback JD, Youngman RR. Root-knot Nematode in Field Corn. United States of America: Virginia Cooperative Extension; 2009 p. 4. Report No.: Publication 444-107. Available from: www.ext.vt.edu.
Mathiyanga K, Manimegalai K, Mariselvi S. Efficacy of Meloidogyne incognita at different inoculum levels on the growth parameters of maize crop. Int J Hortic Crop Sci Res. 2016;6:21–7.
Talwana H, Sibanda Z, Wanjohi WJ, Kimenju W, Luambano-Nyoni N, Massawe C, et al. Agricultural nematology in East and Southern Africa: problems, management strategies and stakeholder linkages. Pest Manag Sci. 2016;72:226–45.
IPCC. Global Warming of 1.5°C: IPCC Special Report on Impacts of Global Warming of 1.5°C above Pre-industrial Levels in Context of Strengthening Response to Climate Change, Sustainable Development, and Efforts to Eradicate Poverty. 1st ed. Cambridge University Press; 2022. Available from: https://www.cambridge.org/core/product/identifier/9781009157940/type/book.
Cowan D, Lebre P, Amon C, Becker R, Boga H, Boulangé A, et al. Biogeographical survey of soil microbiomes across sub-Saharan Africa: structure, drivers, and predicted climate-driven changes. Microbiome. 2022;10:131. https://doi.org/10.1186/s40168-022-01297-w.
Singh BK, Delgado-Baquerizo M, Egidi E, Guirado E, Leach JE, Liu H, et al. Climate change impacts on plant pathogens, food security and paths forward. Nat Rev Microbiol. 2023; Available from: https://www.nature.com/articles/s41579-023-00900-7
Dutta TK, Phani V. The pervasive impact of global climate change on plant-nematode interaction continuum. Front Plant Sci. 2023;14:1143889. https://doi.org/10.3389/fpls.2023.1143889/full.
Velásquez AC, Castroverde CDM, He SY. Plant–pathogen warfare under changing climate conditions. Curr Biol. 2018;28:R619–34.
Juroszek P, Tiedemann AV. Climatic changes and the potential future importance of maize diseases: a short review. J Plant Dis Prot. 2013;120:49–56. https://doi.org/10.1007/BF03356454.
Qiu J, Xie J, Chen Y, Shen Z, Shi H, Naqvi NI, et al. Warm temperature compromises JA-regulated basal resistance to enhance Magnaporthe oryzae infection in rice. Mol Plant. 2022;15:723–39.
Webb KM, Oña I, Bai J, Garrett KA, Mew T, Vera Cruz CM, et al. A benefit of high temperature: increased effectiveness of a rice bacterial blight disease resistance gene. New Phytol. 2010;185:568–76. https://doi.org/10.1111/j.1469-8137.2009.03076.x.
Dossa GS, Oliva R, Maiss E, Vera Cruz C, Wydra K. High temperature enhances the resistance of cultivated African rice, Oryza glaberrima, to bacterial blight. Plant Dis. 2016;100:380–7. https://doi.org/10.1094/PDIS-05-15-0536-RE.
Tsai W-A, Brosnan CA, Mitter N, Dietzgen RG. Perspectives on plant virus diseases in a climate change scenario of elevated temperatures. Stress Biol. 2022;2:37. https://doi.org/10.1007/s44154-022-00058-x
Reynaud B, Delatte H, Peterschmitt M, Fargette D. Effects of temperature increase on the epidemiology of three major vector-borne viruses. Eur J Plant Pathol. 2009;123:269–80. https://doi.org/10.1007/s10658-008-9363-5.
Montes N, Pagán I. Challenges and opportunities for plant viruses under a climate change scenario. In: Adv Virus Res. Amsterdam: Elsevier; 2022. p. 1–66.
Emeraghi M, Achigan-Dako EG, Nwaoguala CNC, Oselebe H. Maize streak virus research in Africa: an end or a crossroad. Theor Appl Genet. 2021;134:3785–803. https://doi.org/10.1007/s00122-021-03914-y.
Khanal C, Land J. Study on two nematode species suggests climate change will inflict greater crop damage. Sci Rep. 2023;13:14185.
Shekhar M, Singh N. The Impact of Climate Change on Changing Pattern of Maize Diseases in Indian Subcontinent: A Review. In: Ahmed El-Esawi M, editor. Maize Genet Resour - Breed Strateg Recent Adv. IntechOpen; 2022. Available from: https://www.intechopen.com/chapters/79453.
Andersen KF, Madden LV, Paul PA. Fusarium head blight development and deoxynivalenol accumulation in wheat as influenced by post-anthesis moisture patterns. Phytopathology. 2015;105:210–9. https://doi.org/10.1094/PHYTO-04-14-0104-R.
Bidzinski P, Ballini E, Ducasse A, Michel C, Zuluaga P, Genga A, et al. Transcriptional basis of drought-induced susceptibility to the rice blast fungus Magnaporthe oryzae. Front Plant Sci. 2016;7. https://doi.org/10.3389/fpls.2016.01558/full.
Hay WT, Anderson JA, McCormick SP, Hojilla-Evangelista MP, Selling GW, Utt KD, et al. Fusarium head blight resistance exacerbates nutritional loss of wheat grain at elevated CO2. Sci Rep. 2022;12:15.
Martínez M, Biganzoli F, Arata A, Dinolfo MI, Rojas D, Cristos D, et al. Warm nights increase Fusarium Head Blight negative impact on barley and wheat grains. Agric For Meteorol. 2022;318:108909.
West JS, Holdgate S, Townsend JA, Edwards SG, Jennings P, Fitt BDL. Impacts of changing climate and agronomic factors on fusarium ear blight of wheat in the UK. Fungal Ecol. 2012;5:53–61.
Sun Y, Yin J, Cao H, Li C, Kang L, Ge F. Elevated CO2 influences nematode-induced defense responses of tomato genotypes differing in the JA pathway. PLoS ONE. 2011;6:e19751. https://doi.org/10.1371/journal.pone.0019751.
Shanker AK, Gunnapaneni D, Bhanu D, Vanaja M, Lakshmi NJ, Yadav SK, et al. Elevated CO2 and water stress in combination in plants: brothers in arms or partners in crime? Biology. 2022;11:1330.
Vanaja M, Sarkar B, Sathish P, Jyothi Lakshmi N, Yadav SK, Mohan Ch, et al. Elevated CO2 ameliorates the high temperature stress effects on physio-biochemical, growth, yield traits of maize hybrids. Sci Rep. 2024;14:2928.
Duku C, Sparks AH, Zwart SJ. Spatial modelling of rice yield losses in Tanzania due to bacterial leaf blight and leaf blast in a changing climate. Clim Change. 2016;135:569–83. https://doi.org/10.1007/s10584-015-1580-2.
Wang H, Mongiano G, Fanchini D, Titone P, Tamborini L, Bregaglio S. Varietal susceptibility overcomes climate change effects on the future trends of rice blast disease in Northern Italy. Agric Syst. 2021;193:103223.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
Benjamin Joshua conceived the design and structure of the review manuscript. All authors- Benjamin Joshua, Oyedokun David, Oziegbe Emmanuel, Oni Joshua, Ogundare Emmanuel, Ujah Godswill and Adebayo Aminat made substantial contributions to the drafting, critical revision, approval of the review manuscript and agree to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Benjamin, J., Oyedokun, D.O., Oziegbe, E.V. et al. Cereal production in Africa: the threat of current plant pathogens in changing climate-a review. Discov Agric 2, 33 (2024). https://doi.org/10.1007/s44279-024-00040-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s44279-024-00040-3