
Abstract
Soil, as a primary repository of plastic debris, faces an escalating influx of microplastics. Microplastics have the potential to decrease soil bulk density and pH, as well as alter soil pore structure and aggregation. These changes in soil physicochemical properties subsequently lead to habitat degradation for microbes and environmental shifts that impact plant growth. Masquerading as soil carbon storage, microplastics can distort assessments of the soil carbon pool by introducing plastic-carbon and associated leachates, influencing soil organic matter (SOM) turnover through priming effects (e.g., dilution, substrate switching, and co-metabolisms). Additionally, microplastics can influence the distribution of soil carbon in particulate and mineral-associated organic matter, consequently affecting the accumulation and stability of soil carbon. Furthermore, microplastics can also influence the chemodiversity of dissolved organic matter (DOM) in soils by increasing DOM aromaticity and molecular weight while deepening its humification degree. The changes observed in soil DOM may be attributed to inputs from microplastic-derived DOM along with organo-organic and organo-mineral interactions coupled with microbial degradation processes. Acting as an inert source of carbon, microplastics create a distinct ecological niche for microbial growth and contribute to necromass formation pathways. Conventional microplastics can reduce microbial necromass carbon contribution to the stable pool of soil carbon, whereas bio-microplastics tend to increase it. Furthermore, microplastics exert a wide range of effects on plant performance through both internal and external factors, influencing seed germination, vegetative and reproductive growth, as well as inducing ecotoxicity and genotoxicity. These impacts may arise from alterations in the growth environment or the uptake of microplastics by plants. Future research should aim to elucidate the impact of microplastics on microbial necromass accumulation and carbon storage within mineral-associated fractions, while also paying closer attention to rhizosphere dynamics such as the microbial stabilization and mineral protection for rhizodeposits within soils.
Graphical Abstract

Highlights
• Microplastics (MPs) have either positive or negative effects on SOM mineralization.
• MPs affect soil carbon distribution in particulate and mineral-associated fraction.
• MPs increase the aromaticity, molecular weight and humification degree of soil DOM.
• Conventional MPs can reduce microbial necromass, whereas bio-MPs cannot.
• MPs influence plant performance through both internal and external factors.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
The terrestrial environment serves as a significant reservoir for microplastics, receiving 4–23 times more plastic waste annually compared to the marine environment (de Souza Machado et al. 2018a; Nizzetto et al. 2016). However, due to the challenges associated with separating microplastics from soil organic matter (SOM) and minerals, it was not until Rillig’s call for research in 2012 that scientific attention began to focus on surveying microplastics in soil (Rillig 2012). Subsequently, the scientific community discovered the presence of microplastics in various soils worldwide, including home gardens, greenhouses, agricultural lands, coastal areas, industrial sites, and floodplain soils (Table S1). Based on our global inventory of microplastics in soil environments, recovered microplastic concentrations range widely from baseline levels up to 20 mg kg–1 in inhabited areas and reach as high as 67,500 mg kg–1 in industrial soil (Figs. 1 and S1), while some studies have suggested even more severe levels of microplastic contamination that may occur in specific soils (Huerta Lwanga et al. 2017). The sources of soil microplastics primarily encompass sewage sludge amendment, irrigation, composting, plastic mulching, dry and wet deposition (i.e., rain and snow) from the atmosphere, fragmentation and degradation processes, the mismanaged runoff by sewer systems from roads or littering activities, landfills, as well as plastics-processing plants (Table S1) (Chen et al. 2022c; Chen et al. 2020; Chen et al. 2023a; Feng et al. 2020; van den Berg et al. 2020). Microplastics exhibit remarkable resistance to environmental degradation (de Souza Machado et al. 2018a; He et al. 2023) and can accumulate persistently in the soil environment over an extended time period while exerting continuous ecosystem effects (Gao et al. 2023; Geyer et al. 2017; Li et al. 2024a; Pan et al. 2023). The pervasive presence, high abundance, and non-natural and persistent characteristics of microplastics in soil underscore their non-negligible role in soil ecology and emphasize the urgency to address their adverse impacts.

Global microplastics abundance in different soils. Here, the data expressed as numerical concentration (items per kilogram soil) were ranked and mapped, whereas global comparison of data expressed as mass concentration (mg per kilogram soil) were provided in Fig. S1. Details information about the sampling and processing methods were provided in Table S1
In recent years, there has been a gradual shift in research focus towards investigating the ecosystem impacts of microplastics in soil (Rillig and Lehmann 2020). Upon entering the soil environment, microplastics have the potential to modify physicochemical properties, reshape microbial habitats, and alter resident microbial communities and activities, consequently influencing soil function and nutrient cycling. Regarding the carbon cycle in soils, microplastics can directly influence the composition of soil carbon pools by introducing both recalcitrant and labile carbon fractions such as soluble additives. Additionally, microplastics may induce positive or negative priming effects on SOM mineralization thereby impacting soil carbon stability (Zhang et al. 2023a). Notably, soil microbes play dual roles in mediating the soil carbon cycle by promoting canbon release through catabolic activities while also preventing canbon release via canbon stabilization mechanisms (Schimel and Schaeffer 2012). Microplastics could potentially affect microbial functions associated with SOM decomposition. Furthermore, microplastic interactions with microbial death pathways may regulate the accumulation and stability of microbial necromass (Camenzind et al. 2023; Chen et al. 2023b). Other mechanisms of direct microplastic interaction with microbes such as electrochemistry-based “electron shuttling” and “microbial frustration” might also impact the soil carbon cycle dynamics (Rillig et al. 2021). However, the impact of microplastics on soil carbon pool composition and stability remains largely unexplored, and the underlying mechanisms have yet to be comprehensively examined.
Soil plants play a crucial role in soil carbon and soil health by fixing carbon through photosynthesis. The rhizosphere of plant is a hotspot of microbial activity and significantly influences the soil carbon cycle. Existing evidence suggests that microplastics impact the rhizosphere environment, including aggregates and microbial communities (Rong et al. 2023; Song et al. 2023), thereby affecting the input of rhizodeposits into soil carbon pools and the stability of soil organic carbon (SOC). Moreover, plastic debris could accumulate pesticides and other toxins present in the soil and leach out additives with potential carcinogenicity and mutagenicity (Ramos et al. 2015), which may affect crop quality, thus posing a threat to human health through the food chain (Fu and Du 2011; Ge et al. 2021; Khalid et al. 2020; Li et al. 2024b; Wang et al. 2022, 2015a). In the context of the near-permanent increase in microplastic contamination, a holistic understanding of ecosystem feedbacks of soil to microplastic contamination is needed.
In the present review, we conducted an exhaustive literature review to address the following two topics: (1) the effects of microplastics on the content, composition, and turnover of SOM, especially the more sensitive dissolved organic matter (DOM) and the more stable microbial necromass carbon, and (2) the effects of microplastics on plant performance and potential mechanisms, as well as plant responses to microplastic stressors. For each topic, knowledge gaps and future prospects are proposed.
2 Effects of microplastics on soil physicochemical properties
Ample evidence from the scientific literature suggests that microplastics can modify soil physicochemical properties in many ways, including soil density, porosity, aggregation, pH, and fertility (Fig. 2). These property changes will subsequently shape the habitat for microbial growth, regulate microbial activity, mediate organic carbon bioaccessibility and turnover rate, and affect plant performance. The effects of microplastics on soil properties depend on polymer type, size class, shape, dose, and soil type.

The conceptual diagram depicting the impact of microplastics on soil structure, soil microbes, and turnover of SOM. Microplastics infiltrate the soil environment through various pathways, including mulching, composting, littering, atmospheric deposition, irrigation, and sludge application. These microplastics can alter physicochemical properties of the soil such as density, porosity, aggregation, pH levels, and nutrient availability. Consequently, these changes in properties shape the microbial habitat by influencing growth patterns and regulating activity while also mediating bioaccessibility and turnover rate of SOM. Soil microbes play a dual role in controlling SOM turnover; they promote carbon release through catabolic activities but also prevent its release via mechanisms like necromass production and stabilization. Microplastics may influence SOM mineralization through both positive and negative priming effects with mechanisms primarily involving co-metabolism, dilution effects, and substrate switching
2.1 Soil bulk density
Soil bulk density is generally associated with soil physical quality and rootability, and high soil bulk density usually means strong penetration resistance and poor root growth (Dexter 2004). The density of microplastics ranges from 0.88 to 1.70 g cm–3 (Table S2), which is generally less dense than the predominant minerals in different soils (Lincmaierová et al. 2023). Therefore, the involvement of microplastics often decreases the bulk density of soils. For example, de Souza Machado et al. (2018b) found that polyacrylic fibers, polyamide (PA) beads, polyester (PES) fibers, and polyethylene (PE) fragments all decreased the bulk density of loamy sand soils, with only PES fibers possessing a significant effect at an application rate of 0.4% (w/w) and having a concentration-dependent response. However, the decrease in soil bulk density by microplastic addition does not necessarily indicate positive effects on root growth, as it might not increase soil porosity. Moreover, microplastics could increase soil density in the rhizosphere despite decreasing bulk density, thus exerting complex effects on plant growth (de Souza Machado et al. 2019).
Although the majority of studies suggested a decrease in soil bulk density by microplastic addition, some also showed divergent results (Li et al. 2023). For example, Zhang et al. (2019) suggested that no detectable changes in soil bulk density were observed for clay loam soils at an application rate of 0.3% (w/w). Therefore, it is plausible to speculate that soil texture is a crucial factor regulating the influence of microplastics on soil properties. However, more evidence is needed to verify this speculation.
2.2 Soil porosity
Soil porosity plays a key role in regulating the infiltration rate of water into soil and the rate of nutrient loss from soil through water evaporation, as well as soil microbial respiration. In addition, small pores could reserve soil water and reduce leaching through capillary action (Major et al. 2009). Zhang et al. (2019) observed that PES fibers could alter the pore structure in clay loam soil. de Souza Machado et al. (2018b) demonstrated that PES fibers affected the soil water-holding capacity, which could further affect soil moisture and evapotranspiration (de Souza Machado et al. 2019). Wan et al. (2019) found that PE film could increase water evaporation from clay soil. Jiang et al. (2017) suggested that plastic-film residues (∼15 kg ha−1) could significantly alter the initial gravimetric water content, bulk density, total porosity, and soil water distribution. In addition, Wang et al. (2015c) demonstrated that due to their waterproofness, plastic-film residues could block soil pores and decrease soil pore size, thus affecting water infiltration and resulting in the loss of soil water. Moreover, changes in porosity could alter the sorption capacity of soils and affect the ability of microbes to reach and degrade potential organic substrates (Baldock and Skjemstad 2000). Such deviations from a natural state suggest the potential threat of microplastics to soil ecosystems (Guo et al. 2022).
2.3 Soil aggregates
Soil aggregates are the basis of soil function and play a crucial role in regulating soil structure, shaping the habitat for soil organisms, and determining SOM turnover (Bronick and Lal 2005; Tisdall and Oades 1982). In addition, soil aggregates also significantly affect soil porosity, which in turn influences the movement of gases and water and the activities of associated microbial communities (Rillig and Lehmann 2020; Rillig et al. 2017). The size and stability of soil aggregates regulate soil erodibility (López et al. 2000; Planchon et al. 2000; Somaratne and Smettem 1993). A decrease in soil aggregates might decrease the diversity of soil microenvironments, thereby impoverishing the soil structure (Six et al. 2006; Zheng et al. 2016). PA beads were more prone to be incorporated into the soil matrix (de Souza Machado et al. 2018b) and soil minerals (Chen et al. 2022d) than other polymer types, and PA beads are unlikely to be isolated from soils after incubations. In addition, fragment-shaped microplastics were loosely integrated into soil aggregates, while fiber-shaped microplastics were more tightly integrated (Guo et al. 2020). Thus, microplastics could integrate with soil to varying degrees, which further influences the formation of aggregates (Liu et al. 2023). Zhang and Liu (2018) reported that 72% of the recovered microplastics in soil were associated with soil aggregates and that fibrous microplastics were most commonly found in microaggregates. Boots et al. (2019) observed that the addition of high density polyethylene (HDPE), polylactic acid (PLA), and synthetic fiber all significantly decreased the mean size of water-stable soil aggregates irrespective of the presence of Lolium perenne. Similarly, the addition of polyacrylic fibers (0.05%, 0.10%, 0.20%, and 0.40%, w/w) and PES fibers (0.20% and 0.40%, w/w) all significantly lowered soil water-stable aggregates, and the effects were more significant at higher concentrations (de Souza Machado et al. 2018b). The decrease in soil water-stable aggregates thus increased the water-holding capacity and evapotranspiration. However, the effects of other types of microplastics, such as PA beads and PE fragments, on soil aggregates were much weaker, and low concentrations of PA and high concentrations of PE even exerted divergent results (de Souza Machado et al. 2018b). Moreover, Zhang et al. (2019) observed an increase in soil water-stable aggregates after the addition of 0.1% and 0.3% (w/w) PES fibers. The significant disparity among various studies can be attributed to the use of different polymer types and microplastic concentrations, as well as the influence of distinct soil textures and time duration (Chen et al. 2023b).
A meta-analysis demonstrated that soil microbes contributed to the formation of soil aggregation, although the positive correlation displayed considerable variability regarding the species investigated (Lehmann et al. 2017). A recent study showed that exposure to PE fragments increased the correlation between microbial activity and soil aggregation (de Souza Machado et al. 2018b). The shifts in the correlation between microbial activity and soil aggregation might suggest either a shift in the microbial community or an alteration in the decay of SOM (de Souza Machado et al. 2018b). However, further efforts are required to disentangle the possible underlying mechanisms among the changes in soil physiochemical properties, soil biota and SOM turnover.
2.4 Soil pH
Soil pH is one of the most important chemical attributes in soil and plays a central role in regulating soil properties and microbial succession (Aciego Pietri and Brookes 2008; Rousk et al. 2009; Wang et al. 2024). Moreover, it can also affect plant performance by directly regulating physiological activities in seed germination and root growth, as well as indirectly affecting microbial activity, nutrient availability, and precipitation and dissolution equilibrium through its effects on ion mobility (Bloom 2000). Bandow et al. (2017) reported a decrease in soil pH in the presence of HDPE pellets after 6 and 12 weeks of exposure to photo- and thermo-oxidative conditions. Boots et al. (2019) also observed that the pH of the soil was 0.62 units lower than that of the control after exposure to HDPE in soil planted with Lolium perenne for 30 days. Similarly, a decrease in pH (0.16‒0.21 units) was observed in soil planted with Oryza sativa L. after the first supplementary fertilization (Chen et al. 2022b). Given the weak possibility of the photo-oxidation of microplastics in soil, it is plausible to speculate that the decreased pH was associated with the changed status of cation exchange in the soil and free exchange of protons in the soil water. Further work is required to verify this speculation.
2.5 Soil fertility and nutrient cycling
Microplastics can also affect the fertility and nutrient cycling of soil. A significant increase in the concentrations of DOM, inorganic nitrogen and total phosphorus was observed after 28% (w/w) microplastic treatment in a sandy loam soil, whereas no obvious changes were observed for 28% (w/w) microplastic treatment over 30 days (Liu et al. 2017). Similarly, bioplastic addition significantly increased the DOM and dissolved organic nitrogen content in soil, whereas no significant effects were observed for LDPE (Meng et al. 2022). Notably, the aforementioned treatments are far beyond the environmentally related microplastic concentrations, which may show divergent results from natural conditions. Thus, future studies should underline these phenomena regarding soils of different textures over a long time span. A field study on the contamination of plastic-mulching residues showed that SOM and alkali-hydrolysable N were greatly reduced after a 500 kg ha–1 microplastic application, and an increasing microplastic application rate of 2,000 kg ha–1 further decreased the available P (Hegan et al. 2015). These results confirmed the role of plastic debris in diminishing soil fertility (Xiang et al. 2023), although further work under environmentally related conditions should be conducted to better understand this impact. N-containing microplastics may serve as a nitrogen source and enrich the soil nitrogen content (Palmer 2001). This speculation can be partly supported by high N2O emissions and N-acquiring enzyme activity (β-N-acetyl-glucosaminidase) in artificial soils using PA as the sole carbon source (Chen et al. 2022d).
3 Effects of microplastics on soil carbon pool
As microplastics are mainly composed of carbon, they may disguise as soil carbon storage and affect the content composition and turnover of SOM (Rillig and Lehmann 2020). However, the rate of microplastic carbon entering the soil and its ecosystem effects on SOM are still open questions.
3.1 Disguising as carbon storage and serving as an inert carbon source in soil
Most microplastics are composed of more than 90% carbon, among other elements (Rillig and Lehmann 2020). They are incorporated into soil aggregates in consort with minerals and organic matter. Microplastics can be disguised as part of soil carbon storage independent of photosynthesis and net primary production (Rillig and Bonkowski 2018). Due to its resistance to decomposition, microplastic carbon tends to accumulate in the soil and eventually be immobilized by microbes after a long time span, which may somehow contribute to the carbon cycle and affect SOM turnover (Chen et al. 2022d; Liu et al. 2017).
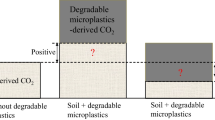
Many previous records have shown the microbial mineralization of microplastic carbon based on direct and indirect evidence, such as changes in surface roughness and functional groups, the diminishment of particle size, the loss of plastic weight, and the colonization of biofilms (McCormick et al. 2014; Yang et al. 2018). In the soil environment, the degradation of microplastics was proven to be very slow (Rillig and Bonkowski 2018). For example, only 0.1 to 0.4% weight loss was observed for PE after 800 days of burial in soil (Albertsson 1980). In addition, only 0.4% weight loss was observed for polypropylene (PP) after one year of incubation (Arkatkar et al. 2009). Moreover, no degradation was found for polyvinyl chloride (PVC) even after 10 to 35 years of burial in soil (Ali et al. 2014; Otake et al. 1995; Santana et al. 2012). However, Chen et al. (2022d) further monitored the CO2 emissions derived from polymer carbon to investigate the accurate degradation pattern of microplastic carbon in artificial soil. They showed that the mineralization ratio of microplastic carbon ranged from 0.007% to 0.876% after 56 days, the upper limit of which was in close proximity to that of SOM (~ 1%) and even exceeded that of biochar-derived carbon (> 0.1%) (Yang et al. 2022). Despite the inert nature of polymer carbon, the results of this fundamental research indicated that the mineralization of polymer carbon is a non-negligible source of CO2 emissions (Chen et al. 2022d) and should be taken into careful consideration in the assessment of carbon storage and carbon emissions (Rillig and Lehmann 2020). In particular, such effects are supposed to be more significant for emerging bioplastics with short lifespans (Zhang et al. 2023a).
In the real soil environment, however, it is difficult to distinguish whether CO2 emissions are derived from microplastic carbon or native SOM. One possible solution is to use carbon isotope labeling to track the carbon source. Zumstein Michael et al. (2018) tracked the carbon from biodegradable polymers into CO2 and microbial biomass in soil based on 13C-labeled polymers and isotope-specific analytical methods. The polymer used in this study was a self-synthesized poly(butylene adipate-co-terephthalate) using 13C-labeled monomers. Zhang et al. (2023a) distinguished the carbon source of CO2 based on the distinct δ13C values of poly-hydroxyalkanoates (PHA), polybutylene succinate (PBS), and PLA from the soils. The observed priming effects across farmland soils were 552% − 1744% for PHA, 44% − 179% for PBS, and − 29% − 43% for PLA. The study underscores the influence of microplastic degradability on the amplitude of priming effects, highlighting the potential significance of microbial nitrogen mining in driving long-term priming effects, given the nitrogen deficiency associated with microplastics. For other commonly used plastic products, such as PE, the 13C isotope labeling of the monomers seems not easy to accomplish. Thus far, it remains largely unknown how much microplastics contribute to carbon emissions.
3.2 Input of DOM leachates in the soil active carbon pool
Various additives, such as plasticizers, stabilizers, antioxidants, flame retardants, and pigments, are usually used to improve plastic quality. The most commonly used additives detected in the environment include phthalates, nonylphenols, bisphenol-A, and brominated flame retardants (Hermabessiere et al. 2017), many of which have been classified as endocrine disruptors and carcinogens (Cherif Lahimer et al. 2017, Wang et al. 2015a). In addition to plastic additives, DOM leachates also include the weathering products of plastics, which mainly comprise long-chain alkanes, high-molecular-weight acids, and benzoic ether (Zhang et al. 2022), which are associated with monomers and oligomers that initially form the polymer chain (Lee and Hur 2020).
These microplastic leachates can be released from microplastics and enter the soil system. On the one hand, leachates serve as an active carbon pool and are directly involved in the soil carbon cycle (Cristina et al. 2022). Romera-Castillo et al. (2018) estimated that global plastic leachates amounted to 23,600 metric tons in the marine environment annually, approximately 60% of which are accessible to microbial utilization in less than 5 days. A 56-day incubation suggested that microplastic leachates had higher degradability than riverine humic acid (Zhang et al. 2022). Thus, it is plausible to speculate that microplastic leachates may have a vital contribution to soil carbon emissions. On the other hand, leachates can be integrated into minerals and SOM (Lee and Hur 2020), exert extreme toxicity to soil microbes (Gaylor et al. 2013; Wei et al. 2019b), and alter the activity, diversity, community structure and metabolism of soil microbes, possibly by destroying the fluidity of the cell membrane (Xie et al. 2010; Zhou et al. 2005). The selection effects on soil biota suggested shifts in the SOM utilization pathway by microbes. Alternatively, leachates are somehow a more active carbon source than plastic solids and certain organic matter and may affect SOM turnover through priming effects (Chen et al. 2022d; Zhang et al. 2022). The priming effects of leached DOM from plastics may, in turn, instigate alterations in DOM across diverse soil types (Sun et al. 2022).
3.3 Changes in SOM composition and mineralization
As mentioned above, microplastics and their leachates could contribute to the inert and active soil carbon pool. Microplastics could be detected as carbon through the current methods applied to quantify SOM. Correspondingly, the SOM contents in soil are prone to increase after microplastic addition (Kim et al. 2021; Li et al. 2022a). However, some studies have also showed that the SOM content was not measurably affected (Boots et al. 2019; Meng et al. 2022) or even decreased (Meng et al. 2022) after microplastic treatments, which was potentially attributed to the low microplastic application rate and the mineralization of polymer carbon or SOC.
Several recent studies have noted shifts in the composition and turnover rate of SOM after microplastic application. For example, PES and bioplastics showed adverse effects on the abiotic characteristics of SOM, such as physical stability (indicated by water molecule bridges), water binding (indicated by decreased desorption enthalpy or faster desorption), and the stability of SOM aliphatic crystallites (Fojt et al. 2022). Chen et al. (2024) observed that polybutylene adipate terephthalate (PBAT) microplastics can masquerade as soil carbon to promote the formation of particulate organic carbon, and facilitate the generation of mineral-associated organic carbon and dissolved organic carbon through microbial transformation of PBAT-derived C and selective consumption in dissolved organic nitrogen. To comprehend the impact of microplastics on soil carbon fluxes, further investigation is warranted to explore how both conventional and biodegradable microplastics influence the accumulation and stabilization mechanisms of different carbon fractions. Current studies have suggested divergent effects of microplastics on soil CO2 emissions, such as increasing (Awet et al. 2018; Shi et al. 2022a; Xiao et al. 2022; Yang et al. 2018), decreasing (Chen et al. 2023b; Wang et al. 2016; Yu et al. 2021), or having no significant effects on CO2 emissions (Blöcker et al. 2020; Yang et al. 2018). However, whether microplastics directly contribute to CO2 emissions is still unclear. Liu et al. (2017) reported an accelerated degradation of SOM with high concentrations of PP addition, which led to distinct metabolite profiles after 7 and 30 days. Xiao et al. (2021) carried out a three-source-partitioning study and showed that low microplastic application can more significantly accelerate SOM decomposition than high microplastic application. However, Yu et al. (2021) showed that microplastic addition reduced the decomposition of SOM by decreasing microbially available SOM but increasing mineral-associated organic matter content.
DOM is a more sensitive indicator of soil quality changes in the SOM pool (Gong et al. 2009) and has a central role in numerous physical, chemical, and biological processes in soil (Kalbitz et al. 2000). Although only accounting for < 0.25% of the total SOM, DOM is crucial in regulating the turnover of SOM, the transport of nutrients, the solubility and mobility of heavy metals and organic pollutants, and the activity of microbes (Kalbitz et al. 1997, 2003; Temminghoff et al. 1997). Microplastic input could alter the quantity of DOM in soil, which depends on the imbalance between the production and mineralization of DOM. For example, a majority of studies have observed increased DOM content after the addition of different microplastics (Liu et al. 2017; Meng et al. 2022; Shi et al. 2022a; Zhou et al. 2020a). This is understandable, as the increased activity of enzymes involved in the degradation of recalcitrant (phenolic) compounds may lead to the decomposition of the poorly dissolved large compounds in SOM into easily dissolved small compounds (Keuskamp et al. 2015). However, some studies have also suggested a decrease in DOM content in microplastic-introduced soils (Liu et al. 2019), which was potentially due to the sorption of DOM by microplastics and the degradation of DOM (Chen et al. 2018). The divergence may also be associated with polymer type, microplastic concentration, and incubation time (Ren et al. 2020). Moreover, microplastics may also influence the chemodiversity of soil DOM, such as increasing DOM aromaticity and molecular weight and deepening the DOM humification degree. (Chen et al. 2022a, 2023b; Feng et al. 2022; Li et al. 2022b; Liu et al. 2017). For instance, the introduction of biodegradable microplastics could enhance the relative abundance of labile compounds in soil DOM, such as lipid-like, protein/amino sugar-like, and carbohydrate-like compounds, while conventional polystyrene (PS) microplastics may decrease the relative abundance of stable compounds like lignin-like and more condensed aromatic-like compounds (Sun et al. 2022). As suggested by Qiu et al. (2024), labile components underwent degradation and transformation after microplastic addition, leading to increased aromaticity and oxidation degree, reduced molecular diversity, as well as altered nitrogen and sulfur contents within soil DOM. The changes in soil DOM might be the combined results of microplastic-derived DOM input, organo-organic and organo-mineral interactions, and microbial degradation. These factors warrant further investigation. In fact, chemodiversity is an important indicator to evaluate the environmental reactivity and destination of soil DOM. In general, DOM of lower molecular weight and aromaticity has higher bioavailability (Fouché et al. 2020; Ye et al. 2020). Additionally, DOM contains polar groups and phenolic structures, which have a stronger binding force with heavy metals (Dong et al. 2020; Wang et al. 2018). Therefore, the shifts in DOM compositions after microplastic input are an important topic.
There are several hypothesized pathways by which microplastics affect the turnover of native SOM (Fig. 2). (1) Microplastics could change soil physicochemical properties such as soil aggregates (habitats for SOM stabilization) and therefore affect microbe growth and activity, thus changing the turnover of native SOM. (2) Microplastics could serve as an inert carbon source and establish a unique ecological niche for microbial settlement and growth, thus affecting SOM turnover by directly altering the microbial community. (3) Microplastics can adsorb SOM, interact with soil minerals, and affect the interaction between minerals and SOM, thus affecting SOM accessibility to microbes. (4) Microplastics may impact SOM mineralization through negative priming effects stemming from dilution and substrate switching or positive priming effects arising from co-metabolism, etc. The latter is likely to have marginal importance given the inert nature of microplastic carbon. Alternatively, these effects might be initiated by the more easily metabolizable bioplastics or microplastic leachates (Zhang et al. 2023a). (5) Conventional PE microplastics can influence CO2 emissions solely by altering DOM electron transfer capabilities. In contrast, the application of biodegradable microplastics (e.g., PLA) can impact gas emissions by increasing both the quantities and transfer capabilities of soil DOM (Shi et al. 2023).
3.4 Microbial response and contribution
Microorganisms have two critical, contrasting roles in controlling terrestrial canbon fluxes: promoting release of canbon to the atmosphere through their catabolic activities, but also preventing release by stabilizing canbon into a form that is not easily decomposed (e.g., the accumulation and stabilization of microbial necromass) (Liang et al. 2017). It is of significant importance to understand the adaptive and evolutionary responses of soil microorganisms to microplastics.
3.4.1 Microbial community structure
The soil microbial community is an important player in regulating nutrient cycling, maintaining soil structure, and detoxifying noxious chemicals. In addition, microbial diversity serves as a sensitive indicator of soil quality, which can reflect subtle changes and soil function (He et al. 2015; Sebiomo et al. 2011). Microplastics exhibit certain filtration effects on soil biota (Guo et al. 2020), and recent studies have reported shifts in community structure, diversity, and evolutionary consequences of microbes in soil in the presence of microplastics (Han et al. 2024; Lu et al. 2023; Rillig et al. 2019a). Microplastics have been proven to serve as substrates for microbial colonization and assemblage in soil environments, leading to the formation of a unique environment termed the plastisphere (Rillig et al. 2023). Distinct differences were noted for the microbial communities between microplastic surface and ambient soils. For example, the PE microplastic surface was found to carry more plastic-degrading bacteria and pathogens than the surrounding soils (Huang et al. 2019). A similar colonization of plastic-degrading bacteria on microplastic surfaces was also observed in an e-waste dismantling field (Chai et al. 2020). The selection phenomena of microplastics on soil biota shed light on screening microbes for microplastic biodegradation.
Many studies have also reported alterations in bulk microbial diversity and composition in soils after microplastic addition. For example, the bacterial composition in microplastic-amended soils showed significant variations from the control after 90 days of incubation (Huang et al. 2019). Membranous PE and fibrous PP could raise the alpha diversities of the soil microbial community (Yi et al. 2021). In contrast, plastic film residues sharply decreased the soil microbial community and its diversity (Wang et al. 2016), which was attributed to the negative effects of lipophilic phthalate ester additives on the soil biota by destroying cell membrane fluidity (Xie et al. 2010; Zhou et al. 2005). Additionally, the soil microbial diversity was lowered in the presence of PE and PVC microplastics (Fei et al. 2020). Until now, the effects of microplastics on microbial community structures have remained unclear. The contradictory results may be attributed to different microplastic types, concentrations, and additive contents in soil. In general, high microplastic concentrations may induce a quick response in the soil microbiota, whereas lower microplastic concentrations may have insignificant effects on the microbial communities. For example, the addition of PE, polyethylene terephthalate (PET), and PVC microplastics at rather low concentrations (< 1%, w/w) after 9 months barely altered the microbial community structure in soil (Judy et al. 2019).
To ease traditional microplastic contamination, biodegradable microplastics have received wide attention and application. As biodegradable microplastics have short lifespans, they are supposed to have more significant effects on soil ecosystems regarding changes in soil biota, greenhouse gas emissions, plant performance, etc. For example, more significant effects on wheat growth were observed for biodegradable microplastics compared to traditional PE microplastics, potentially attributed to microbial immobilization (Qi et al. 2018). More significant variations in bacterial communities, such as the increase in the relative abundance of the genera Bacillus and Variovorax, were noted for biodegradable microplastics than for traditional PE microplastics (Qi et al. 2020b). In addition, PES and PP were observed to increase root symbiosis by arbuscular mycorrhizal fungi, whereas PET had the opposite effect (de Souza Machado et al. 2019). Notably, microplastics are N-limited carbon materials. Thereby, Zhang et al. (2023a) proposed that bioplastics may promote the proliferation of fast-growing r-strategists through co-metabolism, thereby fostering a positive priming effect in the short term. Conversely, under conditions of severe nitrogen deficiency and labile carbon exhaustion, this may lead to the active growth of K-strategists alongside microbial necromass from r-strategists.
In general, microplastics possess a range of effects on soil microbial community, which are still largely unknown. To better underline the ecosystem effects of microplastics, the adaptive and evolutionary responses of soil biota to microplastic stresses should be further addressed in future studies (Rillig 2018; Rillig et al. 2019a).
3.4.2 Enzyme activity
Soil enzymes are useful for monitoring soil health because of their sensitivity to soil stress, energy flow, and nutrient availability (Wang et al. 2015b). Microplastics could affect the excretion of various soil enzymes by microbes, and extracellular enzymes could attach to microplastic surfaces or affect other soil substrates, thus regulating microplastic degradation and turnover of SOM. de Souza Machado et al. (2018b) first evaluated the effects of microplastics on the hydrolysis of fluorescein diacetate enzyme activity and observed a significant correlation between microplastic concentration and microbial activity irrespective of polymer type. This is consistent with the increased hydrolysis of fluorescein diacetate enzyme activity after PE addition (Liu et al. 2017). Similarly, the addition of membranous PE, fibrous PP, and microsphere PP all increased the urease, dehydrogenase, and alkaline phosphatase enzyme activities in soil (Yi et al. 2021). Additionally, the addition of PP microplastics at a rather high application rate (28%, w/w) could significantly increase the activities of fluorescein diacetate hydrolase and phenol oxidase in sandy loam soils (Yang et al. 2018), thus affecting soil C, N, and P cycling and increasing nutrient availability to plants by enhancing microbial hydrolytic activity on SOM (Liu et al. 2017; Yang et al. 2018). However, a lower application rate (7%, w/w) of PP only had marginal effects on the enzyme activities of fluorescein diacetate hydrolase, urease, and phenol oxidase (Yang et al. 2018). In addition, PE and PVC microplastics could increase the activity of urease and acid phosphatase, whereas the activity of fluorescein diacetate hydrolase activity was inhibited (Fei et al. 2020). Decreases in dehydrogenase activity and enzyme activities involved in the C-(β-glucosidase and cellobiohydrolase), N-(leucine-aminopeptidase), and P-(alkaline-phosphatase) cycles were also observed in soils after 28 days of incubation under 100 and 1000 ng g–1 PS nanoplastic treatments, indicating a broad and detrimental impact of PS nanoplastics on soil microbial activity (Awet et al. 2018). Moreover, the residues of plastic film (67.5 kg ha–1) significantly lowered the activity of fluorescein diacetate hydrolase and dehydrogenase by 10% and 20%, respectively (Wang et al. 2016), which may be attributed to the negative effects of concomitant plastic additives. The contradictory results of enzyme activities after microplastic application are potentially attributed to microplastic concentrations and constitutions, plastic additives, and soil properties, the effects of which should be further addressed to better understand the mechanisms. Song et al. (2023) elucidated that PVC elevated β-glucosidase, leucine aminopeptidase, and acid phosphatase activities in both hot- and coldspots within the rice rhizosphere. In contrast, PLA influenced enzyme activities exclusively in the hotspot soil, showing no impact in the coldspot soil. These variations arose from changes in microbial enzyme systems favoring nutrient mining, potentially mitigating some of the adverse effects of microplastics on soil nutrient processes. The close association between soil enzyme activities and soil carbon dynamics necessitates greater emphasis on investigating the response of enzyme activity to microplastic addition, as well as its correlation with other biotic and abiotic processes in soil.
3.4.3 Microbes and enzymes for microplastic degradation
Microplastics can establish a unique ecological niche for certain microbes by providing habitat for microbial settlement and growth. In return, these microbes may contribute to the degradation of microplastics by utilizing polymer carbon as an inert carbon source, consequently impacting the mineralization process of native SOC. As summarized in Table 1, both bacteria and fungi have been proven to have the potential to promote the degradation of microplastics (Russell Jonathan et al. 2011; Shah et al. 2008; Zafar et al. 2013). Some studies have also shown that several bacteria and fungi can use plastic as the sole carbon source, including either in solid or liquid matrices, such as soil (Mohan et al. 2016), compost (Jeon and Kim 2013), and sea water (Harshvardhan and Jha 2013), thus highlighting the potential of such microbes for plastic remediation. Other microbes involved in plastic degradation include Alcaligenes faecalis, Comamonas acidovorans TB-35, Pseudomonas putida, Pseudomonas stutzeri, Saccharomycopsis, Streptomyces sp., and Staphylococcus sp. (Akutsu et al. 1998; Benedict et al. 1983; Caruso 2015; Ghosh et al. 2013).
During the degradation process, it is also very important to identify the enzymes involved (Auta et al. 2018; Jaiswal et al. 2020). Extracellular enzymes secreted by microbes are prone to depolymerize microplastics through hydrolysis reactions (Shah et al. 2008). Lipases, cutinases, carboxylesterases, and laccases have been proven to efficiently degrade microplastics (Lucas et al. 2008). The polymer chain can be cleaved into micromolecular water-soluble intermediates, which may be absorbed by the cells and undergo a special metabolism (Gewert et al. 2015). The final degradation products may end up as CO2, H2O and CH4 and are released into the ambient environment (Tokiwa et al. 2009).
3.4.4 Contribution of microbial biomass and necromass to soil carbon pool
Microbial biomass represents the living or actively growing microorganisms, which is associated with the decomposition efficiency of SOM. To date, data on the effects of microplastics on the accumulation of soil microbial biomass are far from sufficient. A 28-day laboratory incubation showed that the addition of 100 and 1000 ng g–1 PS nanoplastics both significantly decreased soil microbial biomass (Awet et al. 2018), suggesting potentially broad antimicrobial activity of PS nanoplastics on soil microbiota. Interestingly, a gradual increase in soil microbial biomass with time was observed for 10 ng g–1 PS nanoplastic-amended soil throughout 28 days of incubation (Awet et al. 2018). At day 1, the soil microbial biomass remained almost unchanged, whereas the enzyme activities, basal respiration rate, and metabolic quotient decreased, suggesting a sublethal effect with 10 ng g–1 PS nanoplastic amendment. At day 28, the soil microbial biomass was significantly higher, potentially due to the increased antimicrobial activity of PS nanoplastics against some microbial genera over time. Thus, dead cells might provide substrates for resistant microorganisms and thus result in cryptic growth (PostGate 1967). Similarly, several studies also observed a decreased microbial biomass in 1% PP-, PE-, or PLA-amended soils (Blöcker et al. 2020; Shi et al. 2022a). In this case, the decrease in microbial biomass was unlikely caused by toxicity effects, as the used plastics were free of antimicrobial additives. However, Zhang et al. (2023a) noted that bioplastics, such as PHA, PBS, and PLA, elevated microbial biomass carbon and dissolved organic carbon levels. This implies that biodegradable microplastics can concurrently expedite microbial assimilation and the transformation of SOM into dissolved organic substrates.
Microbial necromass refers to the non-living remnants of microorganisms, constituting over 50% of SOC pools and approximately 40 times the amount of live microbial biomass carbon (Liang et al. 2017). Chen et al. (2023b) discovered that the addition of PE microplastics could decrease fungal necromass carbon but had minimal impact on bacterial necromass carbon. Zhang et al. (2024) observed distinct effects on microbial necromass accumulation between conventional PE microplastics and biodegradable microplastics (polypropylene carbonate and polybutylene adipate terephthalate synthetic material), with these effects varying across different land-use types.
We propose several hypothesized mechanisms through which microplastics impact the accumulation of microbial necromass carbon. (1) The soluble fraction of microplastics (e.g., plastic additives, long-chain alkanes, high-molecular-weight acids, and benzoic ether) can act as a carbon source for soil microbes, thereby influencing microbial population and necromass accumulation. (2) The toxic compounds present in microplastics may pose a threat to soil microbes, affecting their growth and death pathways. (3) Microbial necromass could serve as a carbon source and enhance the availability of soil carbon (Caldwell 2005). Microplastic-derived carbon may have positive or negative priming effects on native SOC mineralization, including the mineralization of microbial necromass carbon. (4) As soil aggregates are fundamental units for SOC protection, microplastics can also mediate microbial necromass accumulation and decomposition by impacting soil aggregation. Changes in soil aggregation also shape microbial habitats and influence microbial population dynamics and mortality rates. Moreover, binding agents secreted by soil microbes could reciprocally alter soil aggregation processes that subsequently affect microbial metabolism and SOM turnover. Currently, there is limited research on the accumulation of microbial necromass caused by microplastics; thus, further investigation is required to verify the underlying mechanisms.
4 Effects of microplastics on terrestrial plants
Plants play a vital role in maintaining the health of soil ecosystems and are widely applied to detect and evaluate the toxicity of different environmental stressors. Additionally, plants significantly contribute to soil carbon sequestration through photosynthetic carbon fixation. Moreover, the plant rhizosphere serves as a hotspot for microbial activity and exerts substantial influence on the soil carbon cycle. The presence of microplastics can impact both rhizodeposit secretion by plants and the decomposition and stabilization of rhizodeposits within the soil carbon pool, thereby bearing significant implications for the soil carbon cycle.
4.1 Effects of microplastics on plant performance
As summarized in Fig. 3 and Table 2, microplastics have been observed to affect plant performance regarding seed germination (Bosker et al. 2019; Sforzini et al. 2016; Shi et al. 2022b), vegetative and reproductive growth (Miao et al. 2023; Qi et al. 2018), root and leaf traits (de Souza Machado et al. 2019), crop production (Chen et al. 2022b; Hegan et al. 2015), and ecotoxicity and genotoxicity on plants (Jiang et al. 2019).

Concept graphic about the effects of microplastics on plant performance. Microplastics can impede seed germination by obstructing testa pores and infiltrating seed tissues. Furthermore, they can influence aboveground plants’ vegetative or reproductive growth. Additionally, microplastics have the potential to alter root traits and rhizosphere, thereby affecting root elongation
The incubation time could affect the apparent impact of microplastics. For example, PS microplastics within a size range of 0.05–4.80 μm were observed to delay seed germination of Lepidium sativum during 8 h of incubation, whereas the inhibition effects were no longer observed after 24 h of incubation (Bosker et al. 2019). Additionally, the promotion or inhibition effects on root growth were no longer found as the incubation time was prolonged from 24 to 48 h (Bosker et al. 2019). Moreover, different impacts of microplastics were noted in regard to the physiological responses of Lepidium sativum after 6 days of acute exposure and 21 days of chronic exposure experiments (Pignattelli et al. 2020). Therefore, the design of the experimental span could affect the assessment of the ecological risks of microplastics.
Microplastic concentration is also a crucial factor governing plant performance. For instance, microplastics at higher concentrations exerted more significant delaying effects on seed germination (Bosker et al. 2019). Additionally, PS microplastics at higher concentrations exerted more significant inhibitory effects on the root length and fresh and dry weight of Vicia faba (Jiang et al. 2019). In addition, some research data indicated that crop yields could decrease when mulch film residues amounted to 58.5 kg ha‒1 (Dong et al. 2015; Li et al. 2014).
Different polymer types may exert different effects on the exposed plants. For example, PE is widely applied as mulch film in agriculture, and residues have been observed to decrease crop production (Jiang et al. 2017; Zhang et al. 2016, 2023b). However, bioplastic film, a seemingly more environmentally friendly alternative to traditional mulch film, was observed to exert a stronger negative impact on the vegetative and reproductive growth of Triticum aestivum (Qi et al. 2018), which aroused debates about its wide application (Campanale et al. 2024; Tan et al. 2016). In addition, the impact on the root-leaf ratio, root tissue density, root colonization by arbuscular mycorrhizal fungi (AMF), and leaf traits of Allium fistulosum varied with polymer type (de Souza Machado et al. 2019).
Microplastic size and shape also have very different effects on plant performance. For example, PS microplastics were prone to inhibit the growth of Vicia faba, whereas stronger inhibiting effects were observed for PS nanoplastics; PS microplastics of larger size exerted stronger delaying effects on seed germination (Bosker et al. 2019). Additionally, exposure to 50 nm PS particles resulted in a significant increase in root growth of Lepidium sativum after 24 h of incubation, while exposure to 500 nm PS particles led to a significant decrease in root growth, and exposure to 4800 nm had no significant influence (Bosker et al. 2019). Rillig et al. (2019b) proposed the major hypothesized effect pathway for different shapes and sizes and suggested that fibers of large size may exert positive effects on plant growth by altering soil structure and bulk density, films of intermediate size may exert negative effects on plant growth by increasing soil water evaporation, and beads or fragments of minor size could also possess certain effects similar to minor changes in soil texture.
In the aforementioned studies, the effects of shape or size cannot be completely separated from those of polymer type. Considering the various impacts of microplastics owing to their dazzling range of chemical makeup, additives, persistence, surface properties, sizes and shapes, future research should investigate the role and correlation of different factors on root growth parameters, nutrient uptake, and root-colonizing microbes over different time spans (Rillig et al. 2019b).
4.2 Mechanisms underlying the impact of microplastics on plant performance
Different mechanisms were noted for the impact of microplastics on plant performance, such as serving as root barriers, changing soil microbial structure and metabolism, accumulating in plants, regulating the response of plants to microplastic contamination, etc. Such effects could occur separately or concurrently but are discussed separately as follows (Fig. 3).
4.2.1 External effects of altering the environment in regard to plant growth
First, microplastics could alter soil physicochemical parameters, which may alter water transport, nutrient availability, microbial activities, and penetration resistance for plant roots, thus affecting plant performance. For instance, PES exerted stronger effects on soil structure and interaction with water than other polymer types, which potentially drove the response of plant traits (de Souza Machado et al. 2019). The substantially enhanced water-holding capacity in soil and the maintenance of high water levels in plant tissues by PES addition could alter plant physiological status (e.g., physiological proxies of photosynthesis) and thus result in changes in specific plant traits (Faucon et al. 2017). Song et al. (2023) also suggested that PVC could suppress shoot growth by influencing nutrient and water absorption abilities (Qi et al. 2020a), as well as integrating into soil aggregates to inhibit the activities of soil enzymes and change the uptake of nutrients and water (Gao et al. 2021; Hou et al. 2021; Ng et al. 2021). Alternatively, shifts in water dynamics may affect nutrient availability by altering chemical speciation processes within soils or changing soil microbial activities (de Souza Machado et al. 2019). As plant performance is also highly dependent on soil biota (Wagg et al. 2014), and particularly on root-colonizing microbes such as N-fixers (Liu et al. 2024), pathogens and mycorrhizal fungi (Powell and Rillig 2018), the altered microbial community either by the direct impact of microplastics or the indirect impact of changed soil physicochemical properties could also play a significant role. For example, the changed habitat due to PES addition resulted in the root colonization of AMF, which conferred positive effects to plant growth by increasing nutrient availability (de Souza Machado et al. 2019).
Second, microplastics can accumulate at plant roots and affect root growth and nutrient uptake. For example, PS microplastics could affect root growth of Lepidium sativum by accumulating on the root hairs (Bosker et al. 2019). Similarly, PS microplastics 5 μm in size, due to their large size, were prone to accumulate at the root surface and thus affected the root growth and fresh and dry weight of Vicia faba, potentially by blocking the absorption or uptake of essential nutrients and water through root tips (Jiang et al. 2019).
Third, microplastics contain some biogeochemically active elements that might exert selective pressure on soil microbes and plants. For nitrogen-free microplastics like PLA, they can be hydrolyzed to form water-soluble low-molecular-weight oligomers, which induce microbial immobilization and assimilation of essential nutrients and increase stress in plants (Song et al. 2023). For N-containing microplastics (i.e., PA, polyacrylonitrile, and polyaramide), their effects on soil structure and plant performance could be partly attributed to the enrichment of soil nitrogen. PA is generally manufactured through the polymerization of amines and carboxylic acids, and its components loosely interact (Palmer 2001). Thus, the remaining monomers and compounds from the production process could easily leach into the soil and lead to effects analogous to fertilization. This can partly explain the nearly twofold increase in leaf N content in Allium fistulosum by PA application (de Souza Machado et al. 2019). For polymers containing chloride, such as PVC, the leaching of certain components could lead to biogeochemical changes, as very strong filtration effects on microbes were observed for PVC-derived DOM (Zhang et al. 2022). The phenomenon in which plants are unable to counteract PVC toxicity and produce high concentrations of H2O2 during the time period of chronic exposure is also potentially related to the chemical toxicity of chloride (Pignattelli et al. 2020). Moreover, it is plausible to speculate that in the long run, even the inert carbon in plastic polymers may constitute a relevant carbon pool in the soil (Rillig et al. 2019a) and a selection pressure for soil microbes (Chen et al. 2022d). Particularly for bioplastics with weaker persistence, the biodegradation and microbial immobilization of polymer carbon could be more profound, which may lead to more pronounced effects on plant performance (Qi et al. 2018).
Fourth, microplastics may carry toxic additives or adsorb pollutants such as the concurrently used pesticides (Ramos et al. 2015), which may either ease or exacerbate the environmental stress from microplastics (Zhou et al. 2020b). Microplastics can increase the exposure of soil organisms to adsorbed pollutants (Hodson et al. 2017), and the adsorbed toxic substances can exert adverse effects on plant roots and their symbionts (Gaylor et al. 2013; Wei et al. 2019b), thus negatively affecting plant growth. In addition, microplastics may have stimulated opportunistic plant pathogens and induce phytotoxicity due to acidification of the soil during PLA degradation (Song et al. 2023). On the other hand, the adsorption of contaminants by microplastics could lower their availability to soil biota and plants, thus leading to positive effects on plant growth (Kleinteich et al. 2018; Rehse et al. 2018). Thus far, the combined effects of microplastics and their coexisting pollutants are still largely unknown and merit further investigation (Wang et al. 2019).
4.2.2 Internal effects through uptake of microplastics by plants
The root rhizodermis is a barrier to microplastic uptake. As microsized particles are not expected to be transported into the root, the following discussion mainly concerns nanoplastics, which belong to a broad range of microplastics. Nanoplastics are likely to be transported to and accumulate in terrestrial plants, which may cause damage to plants (e.g., changes in the cell membrane, intracellular molecules, and generation of oxidative stress) (Navarro et al. 2008) and could be a threat to the ecological environment and human health (Maity and Pramanick 2020). For example, Jiang et al. (2019) showed that 100 mg L–1 PS nanoplastics (100 nm) exerted a profound inhibitory effect on Vicia faba growth potentially by blocking cell connections or cell wall pores thus inhibiting nutrient transport. Additionally, PS nanoplastics could accumulate in the testa pores and delay seed germination of Lepidium sativum by physically blocking the pores in the seed capsule (Bosker et al. 2019). However, the realization of nanoplastic toxicity is not limited to entering the plant tissue. Alternatively, plastic leachates, such as additives, plasticizers and flame retardants, may serve as contributors to the toxic effects on plants. An exposure experiment of PS nanoplastics showed morpho- and cytogenotoxic effects in Allium cepa by inducing reactive oxygen species production and chromosomal abnormalities and lowering the expression of the cdc2 gene, whereas no deposition of PS beads inside root tissue was observed (Maity et al. 2020). Microplastics of 4.8 μm in size clogged pores in seed capsules and their adherence to seedlings rather than entering the plant tissue also indicated clogging (Bosker et al. 2019). Microplastics are good vectors for other pollutants, such as heavy metals, dioxins, and persistent organic pollutants (Kumar et al. 2020; Wang et al. 2020). Additionally, microplastics usually contain large varieties of additives of great ecological risks, such as flame retardants, plasticizers, heat stabilizers, and antioxidants (Rillig and Lehmann 2020; Zhou et al. 2020b). Such characteristics could contribute to the negative effects of microplastics on plants.
4.3 Response of plants to microplastic threats
In contaminated fields, plants have developed several strategies to deal with xenobiotics, such as avoiding the intake of such compounds, expanding the root system (Boots et al. 2019), and developing detoxification mechanisms through enzymatic reactions such as oxidation, reduction, and conjugation reactions (Wang et al. 2019). As mentioned above, microplastics can affect the water and nutrient supply to plant roots. Correspondingly, several detoxification mechanisms are involved in alleviating such adverse effects. Long-term stressed plants are prone to secrete extracellular enzymes to degrade microplastics and coexisting pollutants and additives. For example, exposure to PS microplastics leads to shifts in the antioxidant defenses of Vicia faba by secreting antioxidant enzymes such as superoxide dismutase and peroxidase enzymes (Jiang et al. 2019). The increased production of H2O2 in plants, as well as the triggered production of low molecular weight compounds with antioxidant action, also supported the antioxidant defenses of plants to microplastic stressors (Pignattelli et al. 2020). Moreover, the microorganisms and microanimals in plant roots could alleviate the toxic effects by degrading and utilizing microplastics and coexisting intermediates. Unfortunately, the influence of microplastics on plant health and microplastic biodegradation in the rhizosphere of plants is still largely unknown and merits further study.
5 Future research directions
Due to the irresistible increase in plastic production and the lack of effective waste disposal countermeasures, the current pressure of microplastic contamination in soil is expected to continue for many years to come. To better understand the ecosystem effects and risks of microplastics in the soil environment, there are still some critical unknowns that need to be addressed in future work: (1) elucidating the correlation between alterations induced by microplastics in physicochemical properties and soil carbon cycling; (2) clarifying the influence of microplastics on microbial stabilization and mineral protection of soil carbon, with particular attention given to microbial necromass accumulation and carbon storage within mineral-associated fractions; (3) determining the source of carbon emissions from native SOC and microplastics through utilization of 13C isotope technology; (4) investigating the effects of microplastics on rhizosphere dynamics, particularly microbial activity and function, as well as microbial stabilization and mineral protection mechanisms for rhizodeposits within soils; (5) exploring ecosystem-level consequences associated with so-called "eco-friendly" bioplastics, since microbioplastics may exert more pronounced effects on soil biophysical properties, which should be considered during their safe management within agricultural contexts.
Availability of data and materials
The datasets used or analyzed during the current study are available from the corresponding author on reasonable request. The supporting information contains figure of global microplastics abundance in different soils expressed as mass concentration, and tables of the density information of commonly used microplastics and basic information about density solution used for microplastic extraction.
Abbreviations
- AMF:
-
Arbuscular mycorrhizal fungi
- DOM:
-
Dissolved organic matter
- HDPE:
-
High density polyethylene
- PA:
-
Polyamide
- PBAT:
-
Polybutylene adipate terephthalate
- PBS:
-
Polybutylene succinate
- PE:
-
Polyethylene
- PES:
-
Polyester
- PET:
-
Polyethylene terephthalate
- PHA:
-
Poly-hydroxyalkanoates
- PLA:
-
Polylactic acid
- PP:
-
Polypropylene
- PS:
-
Polystyrene
- PVC:
-
Polyvinyl chloride
- SOC:
-
Soil organic carbon
- SOM:
-
Soil organic matter
References
Aciego Pietri JC, Brookes PC (2008) Relationships between soil pH and microbial properties in a UK arable soil. Soil Biol Biochem 40(7):1856–1861
Akutsu Y, Nakajima-Kambe T, Nomura N, Nakahara T (1998) Purification and Properties of a Polyester Polyurethane-Degrading Enzyme from Comamonas acidovorans TB-35. Appl Environ Microbiol 64(1):62–67
Albertsson AC (1980) The shape of the biodegradation curve for low and high density polyethenes in prolonged series of experiments. Eur Polymer J 16(7):623–630
Ali MI, Ahmed S, Robson G, Javed I, Ali N, Atiq N, Hameed A (2014) Isolation and molecular characterization of polyvinyl chloride (PVC) plastic degrading fungal isolates. J Basic Microbiol 54(1):18–27
Arkatkar A, Arutchelvi J, Bhaduri S, Uppara PV, Doble M (2009) Degradation of unpretreated and thermally pretreated polypropylene by soil consortia. Int Biodeterior Biodegradation 63(1):106–111
Auta HS, Emenike CU, Fauziah SH (2017) Screening of Bacillus strains isolated from mangrove ecosystems in Peninsular Malaysia for microplastic degradation. Environ Pollut 231:1552–1559
Auta HS, Emenike CU, Jayanthi B, Fauziah SH (2018) Growth kinetics and biodeterioration of polypropylene microplastics by Bacillus sp. and Rhodococcus sp. isolated from mangrove sediment. Mar Pollut Bull 127:15–21
Awet TT, Kohl Y, Meier F, Straskraba S, Grün AL, Ruf T, Jost C, Drexel R, Tunc E, Emmerling C (2018) Effects of polystyrene nanoparticles on the microbiota and functional diversity of enzymes in soil. Environ Sci Eur 30(1):11
Baldock JA, Skjemstad JO (2000) Role of the soil matrix and minerals in protecting natural organic materials against biological attack. Org Geochem 31(7):697–710
Bandow N, Will V, Wachtendorf V, Simon F-G (2017) Contaminant release from aged microplastic. Environ Chem 14(6):394–405
Benedict CV, Cameron JA, Huang SJ (1983) Polycaprolactone degradation by mixed and pure cultures of bacteria and a yeast. J Appl Polym Sci 28(1):335–342
Blöcker L, Watson C, Wichern F (2020) Living in the plastic age - Different short-term microbial response to microplastics addition to arable soils with contrasting soil organic matter content and farm management legacy. Environ Pollut 267:115468
Bloom, P.R. (2000) Soil pH and pH buffering. E. M. Sumner (Ed.), Handbook of Soil Science, CRC Press, Roca Raton, London, New York, Washington DC, USA
Boots B, Russell CW, Green DS (2019) Effects of Microplastics in Soil Ecosystems: Above and Below Ground. Environ Sci Technol 53(19):11496–11506
Bosker T, Bouwman LJ, Brun NR, Behrens P, Vijver MG (2019) Microplastics accumulate on pores in seed capsule and delay germination and root growth of the terrestrial vascular plant Lepidium sativum. Chemosphere 226:774–781
Bronick CJ, Lal R (2005) Soil structure and management: a review. Geoderma 124(1):3–22
Caldwell BA (2005) Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 49(6):637–644
Camenzind T, Mason-Jones K, Mansour I, Rillig MC, Lehmann J (2023) Formation of necromass-derived soil organic carbon determined by microbial death pathways. Nat Geosci 16(2):115–122
Campanale C, Galafassi S, Di Pippo F, Pojar I, Massarelli C, Uricchio VF (2024) A critical review of biodegradable plastic mulch films in agriculture: Definitions, scientific background and potential impacts. TrAC, Trends Anal Chem 170:117391
Caruso G (2015) Plastic Degrading Microorganisms as a Tool for Bioremediation of Plastic Contamination in Aquatic Environments. Journal of Pollution Effects and Control 3:3
Chai B, Li X, Liu H, Lu G, Dang Z, Yin H (2020) Bacterial communities on soil microplastic at Guiyu, an E-Waste dismantling zone of China. Ecotoxicol Environ Saf 195:110521
Chen M, Cao M, Zhang W, Chen X, Liu H, Ning Z, Peng L, Fan C, Wu D, Zhang M, Li Q (2024) Effect of biodegradable PBAT microplastics on the C and N accumulation of functional organic pools in tropical latosol. Environ Int 183:108393
Chen M, Zhao X, Wu D, Peng L, Fan C, Zhang W, Li Q, Ge C (2022a) Addition of biodegradable microplastics alters the quantity and chemodiversity of dissolved organic matter in latosol. Sci Total Environ 816:151960
Chen S, Feng Y, Han L, Li D, Feng Y, Jeyakumar P, Sun H, Shi W, Wang H (2022b) Responses of rice (Oryza sativa L.) plant growth grain yield and quality, and soil properties to the microplastic occurrence in paddy soil. J Soils and Sediments 22(8):2174–2183
Chen W, Ouyang Z-Y, Qian C, Yu H-Q (2018) Induced structural changes of humic acid by exposure of polystyrene microplastics: A spectroscopic insight. Environ Pollut 233:1–7
Chen Y, Gao B, Xu D, Sun K, Li Y (2022c) Catchment-wide flooding significantly altered microplastics organization in the hydro-fluctuation belt of the reservoir. iScience 25(6):104401
Chen Y, Gao B, Yang Y, Pan Z, Liu J, Sun K, Xing B (2022d) Tracking microplastics biodegradation through CO2 emission: Role of photoaging and mineral addition. J Hazard Mater 439:129615
Chen Y, Leng Y, Liu X, Wang J (2020) Microplastic pollution in vegetable farmlands of suburb Wuhan, central China. Environ Pollut 257:113449
Chen Y, Niu J, Xu D, Zhang M, Sun K, Gao B (2023a) Wet Deposition of Globally Transportable Microplastics (<25 μm) Hovering over the Megacity of Beijing. Environ Sci Technol 57(30):11152–11162
Chen Y, Wang Z, Sun K, Ren J, Xiao Y, Li Y, Gao B, Gunina A, Aloufi AS, Kuzyakov Y (2023b) Biochar and microplastics affect microbial necromass accumulation and CO2 and N2O emissions from soil. ACS ES&T Engineering 4(3):603–614
Cherif Lahimer M, Ayed N, Horriche J, Belgaied S (2017) Characterization of plastic packaging additives: Food contact, stability and toxicity. Arab J Chem 10:S1938–S1954
Cristina R-C, Rebeca M-F, Marola S-Y, Xosé Antón Á-S (2022) Leaching and bioavailability of dissolved organic matter from petrol-based and biodegradable plastics. Mar Environ Res 176:105607
de Souza Machado AA, Kloas W, Zarfl C, Hempel S, Rillig MC (2018a) Microplastics as an emerging threat to terrestrial ecosystems. Glob Change Biol 24(4):1405–1416
de Souza Machado AA, Lau CW, Kloas W, Bergmann J, Bachelier JB, Faltin E, Becker R, Görlich AS, Rillig MC (2019) Microplastics Can Change Soil Properties and Affect Plant Performance. Environ Sci Technol 53(10):6044–6052
de Souza Machado AA, Lau CW, Till J, Kloas W, Lehmann A, Becker R, Rillig MC (2018b) Impacts of Microplastics on the Soil Biophysical Environment. Environ Sci Technol 52(17):9656–9665
Dexter A.R (2004) Soil physical quality: Part I. Theory effects of soil texture density and organic matter and effects on root growth. Geoderma 120(3):201–214
Dong H, Liu T, Han Z, Sun Q, Li R (2015) Determining time limits of continuous film mulching and examining residual effects on cotton yield and soil properties. J Environ Biol 36(3):677–684
Dong Y, Li Y, Kong F, Zhang J, Xi M (2020) Source, structural characteristics and ecological indication of dissolved organic matter extracted from sediments in the primary tributaries of the Dagu River. Ecol Ind 109:105776
Faucon M-P, Houben D, Lambers H (2017) Plant Functional Traits: Soil and Ecosystem Services. Trends Plant Sci 22(5):385–394
Fei Y, Huang S, Zhang H, Tong Y, Wen D, Xia X, Wang H, Luo Y, Barceló D (2020) Response of soil enzyme activities and bacterial communities to the accumulation of microplastics in an acid cropped soil. Sci Total Environ 707:135634
Feng S, Lu H, Tian P, Xue Y, Lu J, Tang M, Feng W (2020) Analysis of microplastics in a remote region of the Tibetan Plateau: Implications for natural environmental response to human activities. Sci Total Environ 739:140087
Feng Y, Chen S, Han L, Wang B, Sun H, Xie W, Lu Q, Feng Y, Poinern GEJ, Xue L (2022) Hydrochar and microplastics disturb soil dissolved organic matter and prominently mitigate ammonia volatilization from wheat growing soil. Appl Soil Ecol 178:104552
Fojt J, Denková P, Brtnický M, Holátko J, Řezáčová V, Pecina V, Kučerík J (2022) Influence of Poly-3-hydroxybutyrate Micro-Bioplastics and Polyethylene Terephthalate Microplastics on the Soil Organic Matter Structure and Soil Water Properties. Environ Sci Technol 56(15):10732–10742
Fouché J, Christiansen CT, Lafrenière MJ, Grogan P, Lamoureux SF (2020) Canadian permafrost stores large pools of ammonium and optically distinct dissolved organic matter. Nat Commun 11(1):4500
Fu X, Du Q (2011) Uptake of Di-(2-ethylhexyl) Phthalate of Vegetables from Plastic Film Greenhouses. J Agric Food Chem 59(21):11585–11588
Gao B, Chen Y, Xu D, Sun K, Xing B (2023) Substantial burial of terrestrial microplastics in the Three Gorges Reservoir. China Communications Earth & Environment 4(1):32
Gao B, Yao H, Li Y, Zhu Y (2021) Microplastic addition alters the microbial community structure and stimulates soil carbon dioxide emissions in vegetable-growing soil. Environ Toxicol Chem 40(2):352–365
Gaylor MO, Harvey E, Hale RC (2013) Polybrominated diphenyl ether (PBDE) accumulation by earthworms (eisenia fetida) exposed to biosolids-, polyurethane foam microparticle-, and penta-bde-amended soils. Environ Sci Technol 47(23):13831–13839
Ge J, Li H, Liu P, Zhang Z, Ouyang Z, Guo X (2021) Review of the toxic effect of microplastics on terrestrial and aquatic plants. Sci Total Environ 791:148333
Gewert B, Plassmann MM, MacLeod M (2015) Pathways for degradation of plastic polymers floating in the marine environment. Environ Sci Process Impacts 17(9):1513–1521
Geyer R, Jambeck JR, Law KL (2017) Production, use, and fate of all plastics ever made. Sci Adv 3(7):e1700782
Ghosh SK, Pal S, Ray S (2013) Study of microbes having potentiality for biodegradation of plastics. Environ Sci Pollut Res 20(7):4339–4355
Gong W, Yan X-Y, Wang J-Y, Hu T-X, Gong Y-B (2009) Long-term manuring and fertilization effects on soil organic carbon pools under a wheat–maize cropping system in North China Plain. Plant Soil 314(1):67–76
Guo J-J, Huang X-P, Xiang L, Wang Y-Z, Li Y-W, Li H, Cai Q-Y, Mo C-H, Wong M-H (2020) Source, migration and toxicology of microplastics in soil. Environ Int 137:105263
Guo Z, Li P, Yang X, Wang Z, Lu B, Chen W, Wu Y, Li G, Zhao Z, Liu G, Ritsema C, Geissen V, Xue S (2022) Soil texture is an important factor determining how microplastics affect soil hydraulic characteristics. Environ Int 165:107293
Gupta KK, Devi D (2020) Characteristics investigation on biofilm formation and biodegradation activities of Pseudomonas aeruginosa strain ISJ14 colonizing low density polyethylene (LDPE) surface. Heliyon 6(7):e04398
Hadad D, Geresh S, Sivan A (2005) Biodegradation of polyethylene by the thermophilic bacterium Brevibacillus borstelensis. J Appl Microbiol 98(5):1093–1100
Han L, Chen L, Feng Y, Kuzyakov Y, Chen, Q.a., Zhang, S., Chao, L., Cai, Y., Ma, C., Sun, K. and Rillig, M.C. (2024) Microplastics alter soil structure and microbial community composition. Environ Int 185:108508
Harshvardhan K, Jha B (2013) Biodegradation of low-density polyethylene by marine bacteria from pelagic waters, Arabian Sea. India Marine Pollution Bulletin 77(1):100–106
He J, Ma W, Han L, Chen L, Xu EG, Xing B, Yang Z (2023) Unraveling the role of natural and pyrogenic dissolved organic matter in photodegradation of biodegradable microplastics in freshwater. Carbon Research 2(1):18
He L, Gielen G, Bolan NS, Zhang X, Qin H, Huang H, Wang H (2015) Contamination and remediation of phthalic acid esters in agricultural soils in China: a review. Agron Sustain Dev 35(2):519–534
Hegan D, Tong L, Zhiquan H, Qinming S, Ru L (2015) Determining time limits of continuous film mulching and examining residual effects on cotton yield and soil properties. J Environ Biol 36(3):677–684
Hermabessiere L, Dehaut A, Paul-Pont I, Lacroix C, Jezequel R, Soudant P, Duflos G (2017) Occurrence and effects of plastic additives on marine environments and organisms: A review. Chemosphere 182:781–793
Hodson ME, Duffus-Hodson CA, Clark A, Prendergast-Miller MT, Thorpe KL (2017) Plastic Bag Derived-Microplastics as a Vector for Metal Exposure in Terrestrial Invertebrates. Environ Sci Technol 51(8):4714–4721
Hou J, Xu X, Yu H, Xi B, Tan W (2021) Comparing the long-term responses of soil microbial structures and diversities to polyethylene microplastics in different aggregate fractions. Environ Int 149:106398
Huang Y, Zhao Y, Wang J, Zhang M, Jia W, Qin X (2019) LDPE microplastic films alter microbial community composition and enzymatic activities in soil. Environ Pollut 254:112983
Huerta Lwanga E, Gertsen H, Gooren H, Peters P, Salánki T, van der Ploeg M, Besseling E, Koelmans AA, Geissen V (2017) Incorporation of microplastics from litter into burrows of Lumbricus terrestris. Environ Pollut 220:523–531
Jaiswal S, Sharma B, Shukla P (2020) Integrated approaches in microbial degradation of plastics. Environ Technol Innov 17:100567
Jeon HJ, Kim MN (2013) Isolation of a thermophilic bacterium capable of low-molecular-weight polyethylene degradation. Biodegradation 24(1):89–98
Jiang X, Chen H, Liao Y, Ye Z, Li M, Klobučar G (2019) Ecotoxicity and genotoxicity of polystyrene microplastics on higher plant Vicia faba. Environ Pollut 250:831–838
Jiang XJ, Liu W, Wang E, Zhou T, Xin P (2017) Residual plastic mulch fragments effects on soil physical properties and water flow behavior in the Minqin Oasis, northwestern China. Soil and Tillage Research 166:100–107
Judy JD, Williams M, Gregg A, Oliver D, Kumar A, Kookana R, Kirby JK (2019) Microplastics in municipal mixed-waste organic outputs induce minimal short to long-term toxicity in key terrestrial biota. Environ Pollut 252:522–531
Kalbitz K, Popp P, Geyer W, Hanschmann G (1997) β-HCH mobilization in polluted wetland soils as influenced by dissolved organic matter. Sci Total Environ 204(1):37–48
Kalbitz K, Schwesig D, Schmerwitz J, Kaiser K, Haumaier L, Glaser B, Ellerbrock R, Leinweber P (2003) Changes in properties of soil-derived dissolved organic matter induced by biodegradation. Soil Biol Biochem 35(8):1129–1142
Kalbitz K, Solinger S, Park JH, Michalzik B, Matzner E (2000) Controls on the dynamics of dissolved organic matter in soils: a review. Soil Science 165(4):277–304
Keuskamp JA, Feller IC, Laanbroek HJ, Verhoeven JTA, Hefting MM (2015) Short- and long-term effects of nutrient enrichment on microbial exoenzyme activity in mangrove peat. Soil Biol Biochem 81:38–47
Khalid N, Aqeel M, Noman A (2020) Microplastics could be a threat to plants in terrestrial systems directly or indirectly. Environ Pollut 267:115653
Kim SW, Jeong S-W, An Y-J (2021) Microplastics disrupt accurate soil organic carbon measurement based on chemical oxidation method. Chemosphere 276:130178
Kleinteich J, Seidensticker S, Marggrander N, Zarfl C (2018) Microplastics Reduce Short-Term Effects of Environmental Contaminants. Part II: Polyethylene Particles Decrease the Effect of Polycyclic Aromatic Hydrocarbons on Microorganisms. Int J Environ Res Public Health 15(12):287
Kumar M, Xiong X, He M, Tsang DCW, Gupta J, Khan E, Harrad S, Hou D, Ok YS, Bolan NS (2020) Microplastics as pollutants in agricultural soils. Environ Pollut 265:114980
Lee YK, Hur J (2020) Adsorption of microplastic-derived organic matter onto minerals. Water Res 187:116426
Lehmann A, Zheng W, Rillig MC (2017) Soil biota contributions to soil aggregation. Nature Ecology & Evolution 1(12):1828–1835
Li C, Moore-Kucera J, Lee J, Corbin A, Brodhagen M, Miles C, Inglis D (2014) Effects of biodegradable mulch on soil quality. Appl Soil Ecol 79:59–69
Li H, Liu L, Xu Y, Zhang J (2022a) Microplastic effects on soil system parameters: a meta-analysis study. Environ Sci Pollut Res 29(8):11027–11038
Li J, Yu Y, Zhang Z, Cui M (2023) The positive effects of polypropylene and polyvinyl chloride microplastics on agricultural soil quality. J Soils Sediments 23(3):1304–1314
Li K, Du L, Qin C, Bolan N, Wang H, Wang H (2024a) Microplastic pollution as an environmental risk exacerbating the greenhouse effect and climate change: a review. Carbon Research 3(1):9
Li R, Xi B, Tan W, Yuan Y (2022b) Spatiotemporal heterogeneous effects of microplastics input on soil dissolved organic matter (DOM) under field conditions. Sci Total Environ 847:157605
Li X, Wang X, Ren C, Palansooriya KN, Wang Z, Chang SX (2024b) Microplastic pollution: Phytotoxicity, environmental risks, and phytoremediation strategies. Crit Rev Environ Sci Technol 54(6):486–507
Liang C, Schimel JP, Jastrow JD (2017) The importance of anabolism in microbial control over soil carbon storage. Nat Microbiol 2(8):17105
Lincmaierová K, Botyanszká L, Lichner L, Toková L, Zafeiriou I, Bondarev D, Horák J, Šurda P (2023) Assessing Microplastic-Induced Changes in Sandy Soil Properties and Crop Growth. 5(3):1555–1567
Liu H, Yang X, Liang C, Li Y, Qiao L, Ai Z, Xue S, Liu G (2019) Interactive effects of microplastics and glyphosate on the dynamics of soil dissolved organic matter in a Chinese loess soil. CATENA 182:104177
Liu H, Yang X, Liu G, Liang C, Xue S, Chen H, Ritsema CJ, Geissen V (2017) Response of soil dissolved organic matter to microplastic addition in Chinese loess soil. Chemosphere 185:907–917
Liu, Y., Xu, Z., Chen, L., Xun, W., Shu, X., Chen, Y., Sun, X., Wang, Z., Ren, Y., Shen, Q.J.F.M.R. (2024). Root colonization by beneficial rhizobacteria. 48(1):fuad066.
Liu Y, Zhong Y, Hu C, Xiao M, Ding F, Yu Y, Yao H, Zhu Z, Chen J, Ge T, Ding J (2023) Distribution of microplastics in soil aggregates after film mulching. Soil Ecology Letters 5(3):230171
López MV, Gracia R, Arrúe JL (2000) Effects of reduced tillage on soil surface properties affecting wind erosion in semiarid fallow lands of Central Aragón. Eur J Agron 12(3):191–199
Lu S, Hao J, Yang H, Chen M, Lian J, Chen Y, Brown RW, Jones DL, Wan Z, Wang W, Chang W, Wu D (2023) Earthworms mediate the influence of polyethylene (PE) and polylactic acid (PLA) microplastics on soil bacterial communities. Sci Total Environ 905:166959
Lucas N, Bienaime C, Belloy C, Queneudec M, Silvestre F, Nava-Saucedo J-E (2008) Polymer biodegradation: Mechanisms and estimation techniques – A review. Chemosphere 73(4):429–442
Maity S, Chatterjee A, Guchhait R, De S, Pramanick K (2020) Cytogenotoxic potential of a hazardous material, polystyrene microparticles on Allium cepa L. J Hazard Mater 385:121560
Maity S, Pramanick K (2020) Perspectives and challenges of micro/nanoplastics-induced toxicity with special reference to phytotoxicity. Glob Change Biol 26(6):3241–3250
Major, J., Steiner, C., Downie, A. and Lehmann, J. (2009) Biochar effects on nutrient leaching. In: Biochar for Environmental Management: Science and Technology.
McCormick A, Hoellein TJ, Mason SA, Schluep J, Kelly JJ (2014) Microplastic is an Abundant and Distinct Microbial Habitat in an Urban River. Environ Sci Technol 48(20):11863–11871
Meng F, Yang X, Riksen M, Geissen V (2022) Effect of different polymers of microplastics on soil organic carbon and nitrogen – A mesocosm experiment. Environ Res 204:111938
Miao J, Chen Y, Zhang E, Yang Y, Sun K, Gao B (2023) Effects of microplastics and biochar on soil cadmium availability and wheat plant performance. GCB Bioenergy 15(8):1046–1057
Mohan AJ, Sekhar VC, Bhaskar T, Nampoothiri KM (2016) Microbial assisted High Impact Polystyrene (HIPS) degradation. Biores Technol 213:204–207
Navarro E, Baun A, Behra R, Hartmann NB, Filser J, Miao A-J, Quigg A, Santschi PH, Sigg L (2008) Environmental behavior and ecotoxicity of engineered nanoparticles to algae, plants, and fungi. Ecotoxicology 17(5):372–386
Ng EL, Lin SY, Dungan AM, Colwell JM, Ede S, Huerta Lwanga E, Meng K, Geissen V, Blackall LL, Chen D (2021) Microplastic pollution alters forest soil microbiome. J Hazard Mater 409:124606
Nizzetto L, Futter M, Langaas S (2016) Are agricultural soils dumps for microplastics of urban origin? Environ Sci Technol 50(20):10777–10779
Otake Y, Kobayashi T, Asabe H, Murakami N, Ono K (1995) Biodegradation of low-density polyethylene, polystyrene, polyvinyl chloride, and urea formaldehyde resin buried under soil for over 32 years. J Appl Polym Sci 56(13):1789–1796
Paço A, Duarte K, da Costa JP, Santos PSM, Pereira R, Pereira ME, Freitas AC, Duarte AC, Rocha-Santos TAP (2017) Biodegradation of polyethylene microplastics by the marine fungus Zalerion maritimum. Sci Total Environ 586:10–15
Palmer, R.J. (2001). Polyamides, Plastics. Encyclopedia of Polymer Science and Technology 251.
Pan T, Liao H, Yang F, Sun F, Guo Y, Yang H, Feng D, Zhou X, Wang Q (2023) Review of microplastics in lakes: sources, distribution characteristics, and environmental effects. Carbon Research 2(1):25
Pignattelli S, Broccoli A, Renzi M (2020) Physiological responses of garden cress (L. sativum) to different types of microplastics. Sci Total Environ 727:138609
Planchon O, Esteves M, Silvera N, Lapetite J-M (2000) Raindrop erosion of tillage induced microrelief: possible use of the diffusion equation. Soil and Tillage Research 56(3):131–144
PostGate, J.R. (1967) Advances in Microbial Physiology. Rose, A.H. and Wilkinson, J.F. (eds), pp. 1–23, Academic Press.
Powell JR, Rillig MC (2018) Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol 220(4):1059–1075
Qi R, Jones DL, Li Z, Liu Q, Yan C (2020a) Behavior of microplastics and plastic film residues in the soil environment: A critical review. Sci Total Environ 703:134722
Qi Y, Ossowicki A, Yang X, Huerta Lwanga E, Dini-Andreote F, Geissen V, Garbeva P (2020b) Effects of plastic mulch film residues on wheat rhizosphere and soil properties. J Hazard Mater 387:121711
Qi Y, Yang X, Pelaez AM, Huerta Lwanga E, Beriot N, Gertsen H, Garbeva P, Geissen V (2018) Macro- and micro- plastics in soil-plant system: Effects of plastic mulch film residues on wheat (Triticum aestivum) growth. Sci Total Environ 645:1048–1056
Qiu X, Ma S, Pan J, Cui Q, Zheng W, Ding L, Liang X, Xu B, Guo X, Rillig MC (2024) Microbial metabolism influences microplastic perturbation of dissolved organic matter in agricultural soils. The ISME Journal 18(1):wrad017
Ramos L, Berenstein G, Hughes EA, Zalts A, Montserrat JM (2015) Polyethylene film incorporation into the horticultural soil of small periurban production units in Argentina. Sci Total Environ 523:74–81
Rehse S, Kloas W, Zarfl C (2018) Microplastics Reduce Short-Term Effects of Environmental Contaminants. Part I: Effects of Bisphenol A on Freshwater Zooplankton Are Lower in Presence of Polyamide Particles. Int J Environ Res Public Health 15(2):280
Ren X, Tang J, Liu X, Liu Q (2020) Effects of microplastics on greenhouse gas emissions and the microbial community in fertilized soil. Environ Pollut 256:113347
Rillig MC (2012) Microplastic in terrestrial ecosystems and the soil? Environ Sci Technol 46(12):6453–6454
Rillig MC (2018) Microplastic Disguising As Soil Carbon Storage. Environ Sci Technol 52(11):6079–6080
Rillig MC, Bonkowski M (2018) Microplastic and soil protists: A call for research. Environ Pollut 241:1128–1131
Rillig MC, de Souza Machado AA, Lehmann A, Klümper U (2019a) Evolutionary implications of microplastics for soil biota. Environ Chem 16(1):3–7
Rillig MC, Kim SW, Zhu Y-G (2023) The soil plastisphere. Nat Rev Microbiol. 22(2):64–74
Rillig MC, Lehmann A (2020) Microplastic in terrestrial ecosystems. Science 368(6498):1430–1431
Rillig MC, Lehmann A, de Souza Machado AA, Yang G (2019b) Microplastic effects on plants. New Phytol 223(3):1066–1070
Rillig MC, Leifheit E, Lehmann J (2021) Microplastic effects on carbon cycling processes in soils. PLoS Biol 19(3):e3001130
Rillig MC, Ziersch L, Hempel S (2017) Microplastic transport in soil by earthworms. Sci Rep 7(1):1362
Romera-Castillo C, Pinto M, Langer TM, Álvarez-Salgado XA, Herndl GJ (2018) Dissolved organic carbon leaching from plastics stimulates microbial activity in the ocean. Nat Commun 9(1):1430
Rong L, Wang Y, Meidl P, Wang L, Sun H (2023) Microplastics affect soybean rhizosphere microbial composition and function during vegetative and reproductive stages. Ecotoxicol Environ Saf 252:114577
Rousk J, Brookes Philip C, Bååth E (2009) Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Appl Environ Microbiol 75(6):1589–1596
Russell Jonathan R, Huang J, Anand P, Kucera K, Sandoval Amanda G, Dantzler Kathleen W, Hickman D, Jee J, Kimovec Farrah M, Koppstein D, Marks Daniel H, Mittermiller Paul A, Núñez Salvador J, Santiago M, Townes Maria A, Vishnevetsky M, Williams Neely E, Percy VM, N., Boulanger, L.-A., Bascom-Slack, C. and Strobel Scott, A. (2011) Biodegradation of Polyester Polyurethane by Endophytic Fungi. Appl Environ Microbiol 77(17):6076–6084
Santana VT, Gonçalves SPC, Agnelli JAM, Martins-Franchetti SM (2012) Biodegradation of a polylactic acid/polyvinyl chloride blend in soil. J Appl Polym Sci 125(1):536–540
Sarkhel R, Sengupta S, Das P, Bhowal A (2019) Comparative biodegradation study of polymer from plastic bottle waste using novel isolated bacteria and fungi from marine source. J Polym Res 27(1):16
Schimel, J. and Schaeffer, S. (2012). Microbial control over carbon cycling in soil. 3
Sebiomo A, Ogundero VW, Bankole SA (2011) Effect of four herbicides on microbial population, soil organic matter and dehydrogenase activity. Afr J Biotech 10(5):770–778
Sekhar VC, Nampoothiri KM, Mohan AJ, Nair NR, Bhaskar T, Pandey A (2016) Microbial degradation of high impact polystyrene (HIPS), an e-plastic with decabromodiphenyl oxide and antimony trioxide. J Hazard Mater 318:347–354
Sforzini S, Oliveri L, Chinaglia S, Viarengo A (2016) Application of Biotests for the Determination of Soil Ecotoxicity after Exposure to Biodegradable Plastics. Front Environ Sci 4:1–12
Shah AA, Hasan F, Hameed A, Ahmed S (2008) Biological degradation of plastics: A comprehensive review. Biotechnol Adv 26(3):246–265
Shah Z, Krumholz L, Aktas DF, Hasan F, Khattak M, Shah AA (2013) Degradation of polyester polyurethane by a newly isolated soil bacterium, Bacillus subtilis strain MZA-75. Biodegradation 24(6):865–877
Shi J, Wang J, Lv J, Wang Z, Peng Y, Shang J, Wang X (2022a) Microplastic additions alter soil organic matter stability and bacterial community under varying temperature in two contrasting soils. Sci Total Environ 838:156471
Shi J, Wang Z, Peng Y, Fan Z, Zhang Z, Wang X, Zhu K, Shang J, Wang J (2023) Effects of Microplastics on Soil Carbon Mineralization: The Crucial Role of Oxygen Dynamics and Electron Transfer. Environ Sci Technol 57(36):13588–13600
Shi, R., Liu, W., Lian, Y., Wang, Q., Zeb, A. and Tang, J. (2022b). Phytotoxicity of polystyrene, polyethylene and polypropylene microplastics on tomato (Lycopersicon esculentum L.). Journal of Environmental Management 317, 115441.
Six J, Frey SD, Thiet RK, Batten KM (2006) Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci Soc Am J 70(2):555–569
Somaratne NM, Smettem KRJ (1993) Effect of cultivation and raindrop impact on the surface hydraulic properties of an Alfisol under wheat. Soil and Tillage Research 26(2):115–125
Song B, Shang S, Cai FM, Liu Z, Fang J, li, N., Adams, J.M. and Razavi, B.S. (2023) Microbial resistance in rhizosphere hotspots under biodegradable and conventional microplastic amendment: Community and functional sensitivity. Soil Biol Biochem 180:108989
Sun Y, Li X, Li X, Wang J (2022) Deciphering the Fingerprint of Dissolved Organic Matter in the Soil Amended with Biodegradable and Conventional Microplastics Based on Optical and Molecular Signatures. Environ Sci Technol 56(22):15746–15759
Tan, Z., Yi, Y., Wang, H., Zhou, W., Yang, Y. and Wang, C. (2016). Physical and Degradable Properties of Mulching Films Prepared from Natural Fibers and Biodegradable Polymers. Applied Sciences 6(5).
Temminghoff EJM, Van der Zee SEATM, de Haan FAM (1997) Copper mobility in a copper-contaminated sandy soil as affected by pH and solid and dissolved organic matter. Environ Sci Technol 31(4):1109–1115
Tisdall JM, Oades JM (1982) Organic matter and water-stable aggregates in soils. J Soil Sci 33(2):141–163
Tokiwa Y, Calabia BP, Ugwu CU, Aiba S (2009) Biodegradability of Plastics. Int J Mol Sci 10(9):3722–3742
van den Berg P, Huerta-Lwanga E, Corradini F, Geissen V (2020) Sewage sludge application as a vehicle for microplastics in eastern Spanish agricultural soils. Environ Pollut 261:114198
Wagg C, Bender SF, Widmer F, van der Heijden MGA (2014) Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc Natl Acad Sci 111(14):5266–5270
Wan Y, Wu C, Xue Q, Hui X (2019) Effects of plastic contamination on water evaporation and desiccation cracking in soil. Sci Total Environ 654:576–582
Wang B, Liu C, Chen Y, Dong F, Chen S, Zhang D, Zhu J (2018) Structural characteristics, analytical techniques and interactions with organic contaminants of dissolved organic matter derived from crop straw: a critical review. RSC Adv 8(64):36927–36938
Wang F, Feng X, Liu Y, Adams CA, Sun Y, Zhang S (2022) Micro(nano)plastics and terrestrial plants: Up-to-date knowledge on uptake, translocation, and phytotoxicity. Resour Conserv Recycl 185:106503
Wang J, Chen G, Christie P, Zhang M, Luo Y, Teng Y (2015a) Occurrence and risk assessment of phthalate esters (PAEs) in vegetables and soils of suburban plastic film greenhouses. Sci Total Environ 523:129–137
Wang J, Liu X, Li Y, Powell T, Wang X, Wang G, Zhang P (2019) Microplastics as contaminants in the soil environment: A mini-review. Sci Total Environ 691:848–857
Wang J, Lv S, Zhang M, Chen G, Zhu T, Zhang S, Teng Y, Christie P, Luo Y (2016) Effects of plastic film residues on occurrence of phthalates and microbial activity in soils. Chemosphere 151:171–177
Wang J, Zhang M-Y, Chen T, Zhu Y, Teng Y, Luo Y-M, Christie P (2015b) Isolation and Identification of a Di-(2-Ethylhexyl) Phthalate-Degrading Bacterium and Its Role in the Bioremediation of a Contaminated Soil. Pedosphere 25(2):202–211
Wang W, Ge J, Yu X, Li H (2020) Environmental fate and impacts of microplastics in soil ecosystems: Progress and perspective. Sci Total Environ 708:134841
Wang, Y., Chen, Y., Gao, X., Wang, Q., Cui, M., Zhang, D. and Guo, P. (2024). Unveiling the driving role of pH on community stability and function during lignocellulose degradation in paddy soil. 15.
Wang Z, Li X, Shi H, Sun M, Ding T, Wang C (2015c) Effects of residual plastic film on soil hydrodynamic parameters and soil structure. Transactions of the Chinese Society for Agricultural Machinery 46:101–106
Wei R, Breite D, Song C, Gräsing D, Ploss T, Hille P, Schwerdtfeger R, Matysik J, Schulze A, Zimmermann W (2019a) Biocatalytic Degradation Efficiency of Postconsumer Polyethylene Terephthalate Packaging Determined by Their Polymer Microstructures. Advanced Science 6(14):1900491
Wei W, Huang Q-S, Sun J, Wang J-Y, Wu S-L, Ni B-J (2019b) Polyvinyl Chloride Microplastics Affect Methane Production from the Anaerobic Digestion of Waste Activated Sludge through Leaching Toxic Bisphenol-A. Environ Sci Technol 53(5):2509–2517
Xiang Y, Peñuelas J, Sardans J, Liu Y, Yao B, Li Y (2023) Effects of microplastics exposure on soil inorganic nitrogen: A comprehensive synthesis. J Hazard Mater 460:132514
Xiao M, Ding, J.n., Luo, Y., Zhang, H., Yu, Y., Yao, H., Zhu, Z., Chadwick, D.R., Jones, D., Chen, J. and Ge, T. (2022) Microplastics shape microbial communities affecting soil organic matter decomposition in paddy soil. J Hazard Mater 431:128589
Xiao M, Shahbaz M, Liang Y, Yang J, Wang S, Chadwicka DR, Jones D, Chen J, Ge T (2021) Effect of microplastics on organic matter decomposition in paddy soil amended with crop residues and labile C: A three-source-partitioning study. J Hazard Mater 416:126221
Xie H-J, Shi Y-J, Zhang J, Cui Y, Teng S-X, Wang S-G, Zhao R (2010) Degradation of phthalate esters (PAEs) in soil and the effects of PAEs on soil microcosm activity. J Chem Technol Biotechnol 85(8):1108–1116
Yang X, Bento CPM, Chen H, Zhang H, Xue S, Lwanga EH, Zomer P, Ritsema CJ, Geissen V (2018) Influence of microplastic addition on glyphosate decay and soil microbial activities in Chinese loess soil. Environ Pollut 242:338–347
Yang Y, Sun K, Han L, Chen Y, Liu J, Xing B (2022) Biochar stability and impact on soil organic carbon mineralization depend on biochar processing, aging and soil clay content. Soil Biol Biochem 169:108657
Ye Q, Wang Y-H, Zhang Z-T, Huang W-L, Li L-P, Li J, Liu J, Zheng Y, Mo J-M, Zhang W, Wang J-J (2020) Dissolved organic matter characteristics in soils of tropical legume and non-legume tree plantations. Soil Biol Biochem 148:107880
Yi M, Zhou S, Zhang L, Ding S (2021) The effects of three different microplastics on enzyme activities and microbial communities in soil. Water Environ Res 93(1):24–32
Yoshida S, Hiraga K, Takehana T, Taniguchi I, Yamaji H, Maeda Y, Toyohara K, Miyamoto K, Kimura Y, Oda K (2016) A bacterium that degrades and assimilates poly(ethylene terephthalate). Science 351(6278):1196–1199
Yu H, Zhang Z, Zhang Y, Song Q, Fan P, Xi B, Tan W (2021) Effects of microplastics on soil organic carbon and greenhouse gas emissions in the context of straw incorporation: A comparison with different types of soil. Environ Pollut 288:117733
Yu Y, Battu AK, Varga T, Denny AC, Zahid TM, Chowdhury I, Flury M (2023) Minimal Impacts of Microplastics on Soil Physical Properties under Environmentally Relevant Concentrations. Environ Sci Technol 57(13):5296–5304
Zafar U, Houlden A, Robson Geoffrey D (2013) Fungal Communities Associated with the Biodegradation of Polyester Polyurethane Buried under Compost at Different Temperatures. Appl Environ Microbiol 79(23):7313–7324
Zang H, Zhou J, Marshall MR, Chadwick DR, Wen Y, Jones DL (2020) Microplastics in the agroecosystem: Are they an emerging threat to the plant-soil system? Soil Biol Biochem 148:107926
Zhang D, Liu H-B, Hu W-L, Qin X-H, Ma X-W, Yan C-R, Wang H-Y (2016) The status and distribution characteristics of residual mulching film in Xinjiang. China Journal of Integrative Agriculture 15(11):2639–2646
Zhang, E., Chen, Y., Li, Y., Sun, K., Yang, Y., Gao, B. and Xing, B. (2022). The photo-redox of chromium regulated by microplastics (MPs) and MPs-derived dissolved organic matter (MPs-DOM) and the CO2 emission of MPs-DOM. Fundamental Research.
Zhang G, Liu D, Lin J, Kumar A, Jia K, Tian X, Yu Z, Zhu B (2023a) Priming effects induced by degradable microplastics in agricultural soils. Soil Biol Biochem 180:109006
Zhang, G., Zhang, S., Xia, Z., Wu, M., Bai, J. and Lu, H. (2023b) Effects of Biodegradable Film and Polyethylene Film Residues on Soil Moisture and Maize Productivity in Dryland.
Zhang GS, Liu YF (2018) The distribution of microplastics in soil aggregate fractions in southwestern China. Sci Total Environ 642:12–20
Zhang GS, Zhang FX, Li XT (2019) Effects of polyester microfibers on soil physical properties: Perception from a field and a pot experiment. Sci Total Environ 670:1–7
Zhang H, Huang Y, Shen J, Xu F, Hou H, Xie C, Wang B, An S (2024) Mechanism of polyethylene and biodegradable microplastic aging effects on soil organic carbon fractions in different land-use types. Sci Total Environ 912:168961
Zheng W, Morris EK, Lehmann A, Rillig MC (2016) Interplay of soil water repellency, soil aggregation and organic carbon. A Meta-Analysis Geoderma 283:39–47
Zhou B, Wang J, Zhang H, Shi H, Fei Y, Huang S, Tong Y, Wen D, Luo Y, Barceló D (2020a) Microplastics in agricultural soils on the coastal plain of Hangzhou Bay, east China: Multiple sources other than plastic mulching film. J Hazard Mater 388:121814
Zhou QH, Wu ZB, Cheng SP, He F, Fu GP (2005) Enzymatic activities in constructed wetlands and di-n-butyl phthalate (DBP) biodegradation. Soil Biol Biochem 37(8):1454–1459
Zhou Y, Wang J, Zou M, Jia Z, Zhou S, Li Y (2020b) Microplastics in soils: A review of methods, occurrence, fate, transport, ecological and environmental risks. Sci Total Environ 748:141368
Zumstein Michael T, Schintlmeister A, Nelson Taylor F, Baumgartner R, Woebken D, Wagner M, Kohler Hans-Peter E, McNeill K, Sander M (2018) Biodegradation of synthetic polymers in soils: Tracking carbon into CO2 and microbial biomass. Sci Adv 4(7):eaas9024
Funding
This work was financially supported by the National Science Foundation for Distinguished Young Scholars (42125703) and National Natural Science Foundation (41977299).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Yalan Chen: Conceptualization; Data curation; Validation; Visualization; Writing—original draft. Ke Sun: Conceptualization, Supervision, Funding acquisition, Project administration, Writing—review & editing. Yang Li, Xinru Liang, Siyuan Lu, Jiaqi Ren, Yuqin Zhang, Zichen Han: Conceptualization, Writing—review & editing. Bo Gao: Supervision, Writing—review & editing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
Ke Sun is an editorial board member for Carbon Research and was not involved in the editorial review, or the decision to publish, this article. All authors declare that there are no competing interests.
Additional information
Handling editor: Fangbai Li.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, Y., Li, Y., Liang, X. et al. Effects of microplastics on soil carbon pool and terrestrial plant performance. Carbon Res. 3, 37 (2024). https://doi.org/10.1007/s44246-024-00124-1
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s44246-024-00124-1