
Abstract
The present work aims to provide a high-resolution stratigraphic range of the planktic foraminiferal group during the Campanian/Maastrichtian boundary (C/M) of Gebel Owaina, Nile Valley. We use the base of Planoglobulina acervulinoides (CF7c Subzone) to mark the C/M boundary, where the planktic foraminiferal assemblages in the subzone are proposed as indicative of the early Maastrichtian age. Five zones (CF9-CF5) and three subzones (CF8a, CF8b, and CF8c) were recognized in the present study. A morphogroups scheme for the calcareous benthic foraminifera used in this work comprises five morphogroups, observed in the Owaina section. Three planktic morphogroups can be recognized and indicate open marine conditions with a good connection to the Tethys Ocean due to the highly diverse Maastrichtian assemblage and high P/B ratio (% P > 95).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Previously the Campanian/Maastrichtian (C/M) boundary had been placed at the top of the Radotruncana calcarata Zone [1,2,3,4,5,6,7,8,9]. According to [10], the base of the Maastrichtian is no longer defined by the R. calcarata highest occurrence (HO) but is defined by the lowest occurrence (LO) of the ammonite species Pachydiscus neubergicus [10,11,12].
The C/M boundary GSSP coincides with the LO of the planktic foraminiferal species Rugoglobigerina scotti and Contusotruncana contusa. It is hypothesized that the LO of Contusotruncana contusa could be concurrent with the LO of Rugoglobigerina scotti, as reported in the GSSP for the C/M boundary [13].
The Campanian marks the transition between the mid-Cretaceous “hot greenhouse” and the ʻcool greenhouseʼ of the Maastrichtian and early Paleocene [14,15,16,17,18,19]. The Campanian–Maastrichtian (~ 83–66 Ma) became a duration of worldwide climate cooling, throughout the Late Campanian Event (LCE) and the Campanian–Maastrichtian Transition Event (CMTE) [20]. The cooling was observed by changes in the global oceanic circulation in high southern latitudes [21,22,23,24,25,26,27,28,29,30,31]. The Campanian Maastrichtian was a time of global climate cooling [16, 22]. The culmination of this event was accompanied by third-order global sea level falls [9].
The benthic foraminifera tests are environmentally sensitive and facies-dependent [32,33,34]. The morphogroups analyses of calcareous benthic assemblages are applied for paleoenvironmental studies [e.g., 35,36,37].
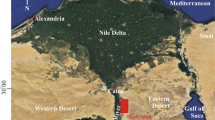
The upper Campanian–Maastrichtian succession of the Dakhla Formation (i.e. Sharawna Shale Member) in the Gebel Owaina section (Fig. 1) is still poorly documented. Dakhla Formation termed by [38] is represented by thick siliciclastic facies which are underlain by the Duwi Formation (phosphate deposits) and overlain by the Tarawan Formation (carbonate facies). The Campanian/Maastrichtian boundary was recorded at the middle part of the middle Sharawna Marl unit (Fig. 2). Biostratigraphic correlation based on planktic and benthic foraminifera has received little attention due to the scarcity of low diversity of microfossils in these, mostly shallow marine sequences.

Geological map of the studied area (After the EGPC/CONOCO, 1987, Luxor sheet) [131]

Measured stratigraphic sections showing the lithostratigraphic units in the studied area
1.1 History of biostratigraphic studies
Said and Sabry (1964) [39] subdivided the Upper Cretaceous Lower Tertiary succession of Gebel Oweina, Nile Valley according to the planktonic foraminifera into several zones. These are as follows from the top to the base; Globigerina rex Zone (Lower Eocene), Globigerina velascoensis Zone (Landenian), Globigerina pseudomenardii Zone (Landenian), Globigerina pusilla pusilla Zone (Landenian), Globigerina uncinata Zone (Landenian), Globigerina daubjergensis-Globigerina pseudobulloides Zone (Danian), and Globotruncana-Heterohelix Zone (Maastrichtian).
El Naggar (1966) [40] made a detailed study on the stratigraphy and planktic foraminifera of some Upper Cretaceous-Lower Tertiary sections along the Esna-Idfu regions, the Nile Valley. He stated that the Esna Group is widely distributed in Egypt extending from the Dakhla-Kharga-Kurkur scarp into the Nile Valley, resting over the Phosphate Formation and underlying the Thebes Formation. He recognized several zones from the base upwards, as follows; Globotruncana fornicate Zone, Globotruncana gansseri Zone, the Globotruncana esnehensis Zone, Globorotalia copressa/Globigerina daubjergensis Zone, and Globorotalia angulata Zone.
El-Younsy et al. (2015) [41] studied the sedimentological and biostratigraphical detailed studies of the Campanian-Eocene succession in the Gebel Oweina area, Upper Egypt, and helped in reaching a high-resolution sequence stratigraphic classification that led to the recognition of the first depositional sequence in the studied Gebel Oweina stratigraphic section coincides with the Campanian of both the Duwi Formation (~ 33 m thick) and the Oweina Shale Member (~ 57 m thick) of the lower part of the Dakhla Formation. It is difficult to determine the precise age dating of this sequence because most of its sediment is barren of any diagnostic planktic foraminifera, except the basal part of the Oweina Shale Member, which contains planktic foraminifera of Globotruncana aegyptiaca. The second depositional sequence in the studied stratigraphic section is made up of both the Sharawna Marl Member (~ 16 m thick) and the lower part of the Oweina Shale Member (~ 52 m thick) of the middle part of the Dakhla Formation. It represents the Maastrichtian zones Gansserina gansseri and Pseudogumbelina palpebra that extends from ~ 70.4 to ~ 65.2 Ma [52]. The third depositional sequence represents the Late Campanian G. aegyptiaca Zone (~ 9 m thick) followed upward by the Maastrichtian G. gansseri, P. hariaensis, P. palpebra, and P. Hantkeninoides zones (~ 21 m thick) that extends from ~ 70.4 to ~ 65.0 Ma [53].
El-Younsy et al. (2017) [42] reviewed the Upper Cretaceous-Lower Paleogene succession at the Gebel Qareiya area and they investigated several distinctive benthonic foraminiferal assemblages, these foraminiferal assemblages, and their paleobathymetric interpretation are as follows; the pre-Campanian Shale of the Quseir Formation is characterized by the presence of Haplophragmoides calculus benthonic assemblage, which points to the inner neritic paleodepths (~ 10–50 m) setting, the lower Campanian shale and marl of the Duwi Formation are characterized by the presence of Lagena sulcata and Tritaxia midwayensis benthonic assemblage, which points to inner to middle neritic paleodepths (~ 40–80 m) setting, the Upper Campanian–Maastrichtian of the Hamama Marl Member is characterized by the occurrence of Anomalina umboniferus, Rectuvigerinastriata, and Bolivina midwayensis benthonic assemblage, which points to outer neritic-upper bathyal paleodepths (~ 150–600 m) setting, coincides with an increase in the P/B ratio of ~ 30–80%.
Farouk et al. (2019) [43] studied Maastrichtian‐early Paleocene foraminiferal palaeobathymetry, palaeodiversity, and vertical facies changes of Gebel El Sharawna, south Luxor, Egypt, and determine the depositional sequences, their relationships to global records and/or tectonic signatures. Five benthonic assemblages are recorded and replicated in the present study to reflect fluctuation in palaeo‐water depth from restricted marginal marine to outer shelf palaeoenvironments. Four sequence boundaries that coincide with the Campanian/Maastrichtian, intra‐early Maastrichtian, Early/Late Maastrichtian, Cretaceous/Palaeogene (K/ Pg), and intra‐Danian were recognized based upon sharp vertical facies changes, foraminiferal assemblage changes, hiatuses, mineral hard ground, and reworking.
This paper aims to document the morphology of the foraminifera and stratigraphic range of the planktic foraminiferal group during the Campanian/Maastrichtian boundary that has prevailed during the sedimentary deposition across the Campanian/Maastrichtian boundary and evaluate the paleoecology of the Gebel Owaina section. The new detailed subzonation of the standard Gansserina gansseri Zone [5, 13, 44,45,46,47] involves the consecutive origination of rugoglobigerinids and multiserial heterohelicids.
2 Geological setting and lithostratigraphy
During Campanian–Maastrichtian, Egypt was part of a vast epicontinental shelf at the edge of the southern Tethys Ocean on the northwestern margin of the Arabian-Nubian massif [48]. Tectonics played an important role in controlling the configuration of the Egyptian Campanian–Maastrichtian depositional environment [48,49,50,51]; leading to great vertical and lateral complexities in the facies distribution and thicknesses. The Campanian–Maastrichtian succession in the studied area shows great lateral changes in both lithofacies and biofacies.
The shelf area of Egypt, during the Late Cretaceous and Early Paleogene times, can be divided into the Stable Shelf surrounding the basement exposures of the Arabo Nubian craton and the Unstable Shelf [38]. The Late Cretaceous-Early Eocene sediments in central and southern Egypt are characterized by gradual facies [52].
Gebel Oweina area (Latitude 25°15'N; Longitude 32°45' E) is located on the eastern side of the upper Nile Valley, about 25 km SE of Esna town and 8.5 km NE of the Sibaiya railway station. It rises about 450 m above sea level [38] and has a height: of ~ 154 m.
The detailed lithostratigraphic description of the encountered Upper Cretaceous rock units in the studied area is given below. The samples examined in the present study were obtained from one stratigraphic surface section (Fig. 2)., and about 72 samples were collected from the Nile Valley facies (Quseir, Duwi, and Dakhla formations).
2.1 Quseir Formation [53]
Attains ~ 5 m thickness and consists of varying color shales alternating with sandstone and siltstone beds (Campanian) (Fig. 3).

Photograph of fieldwork. A Bed of Pecten farafrensis of the upper Sharawna Shale unit. B Hematitic shale of the lower Sharawna Shale unit. C Gypsum and anhydrite veinlets of the lower Sharawna Shale unit. D Campanian/Maastrichtian boundary recorded at the middle part of the middle Sharawna Marl unit. E Middle Sharawna Marl unit overlies the lower Sharawna Shale unit
2.2 Duwi Formation [53]
Attains ~ 10 m thickness and consists of phosphates deposits interbedded with siltstones, marls, and shales. In the south of the Western Desert, [54, 55] considered the Duwi Formation is of the late Campanian to early Maastrichtian age. Stratigraphically, the Duwi Formation is underlain by the Quseir Formation (Quseir Variegated Shale) and overlain by the Dakhla Formation [e.g., 38, 56, 57] and assigned to late Campanian-early Maastrichtian (Fig. 3).
2.3 Dakhla Formation [38, 56]
The Dakhla Formation in the studied succession has a thickness of about 154 m and is made mainly up of shale, light grey at the top, and green at the base, including marl bands. Generally, the Dakhla Formation overlies and underlies the Duwi and Tarawan formations, respectively, with two unconformity surfaces at the base and the top. The Dakhla Formation was assigned to the early Campanian-middle Paleocene, [57], in the studied area the two-thirds part of the Dakhla Formation attains a thickness of 117 m and is assigned to the late Campanian-early Maastrichtian (Fig. 3).
Lithologically, the Dakhla Formation consists of two members; Sharawna Shale Member (Cretaceous) and Owaina Shale Member (Paleocene) [55]. The Sharawna Shale Member consists of three distinct units; a lower shale unit (about 57 m thick), a middle marl unit (12 m thick), and an upper shale unit (about 48 m thick). The Sharawna Shale Member [58] can be correlated with the Hamama Member [59] in the Qena-Luxor region. The middle marl unit is composed of biscuit-colored, hard, and highly fossiliferous. This unit can be correlated with the Sharawna Marl unit [55, 60].
The Owaina Shale Member [55] of the Dakhla Formation (Paleocene) is composed of dark grey, greenish grey, green, greyish violet, and greenish-yellow shales with clay ironstone bands, it is intercalated with gypsum vines. The famous disconformity surface between the Cretaceous and Paleocene occurs near the base of the Owaina Shale Member and is represented by a 50 to 120-cm thick phosphatic conglomerate layer. The Owaina Shale Member can be correlated with the Beida Member [60]. This shale member is disconformably overlain by the Paleogene sediments.
3 Material and methods
3.1 Foraminiferal studies
All collected 72 samples were dried for at least 24 h in an oven at 60˚C. About 50 g from each sample were soaked in Hydrogen Peroxide solution (10% H2O2), then washed over a 63 μm mesh sieve, and then dried and sieved into fractions greater than 250, 125, and 63 μm. The washed residue from each sample was studied under a binocular microscope. The foraminifera tests were picked into faunal slides. About 250–300 foraminifera specimens were picked from a random sample in the rich samples but 50 individuals in the poor samples [61]. The following parameters were calculated; TFN = Total Foraminifera Number, number of planktic foraminifera (P), number of benthic foraminifera (B), Planktic ratio P % = (P/ (P + B) × 100), Agglutinated Foraminifera/Calcareous Foraminifera (A/C) %, Epifaunal % = Epifaunal/Epifaunal + Infaunal × 100 (Epi% = E/E + I X 100), diversity = number of species per sample, Fisher’s alpha, and Shannon–Weaver. The most important foraminifera are photographed using scanning electron microscopy (JEOL JSM-5500 LV) of Alexandria University (Figs. 4, 5 and 6).

Scanning Electron Microscope (SEM) of foraminifera. A Spiroplectinella esnaensis (Le Roy), Sample 66, CF5 Zone, Owaina sectionB Gaudryina rugosa (D’ Orbigny), sample 66, CF5 Zone, Owaina section. C Gaudryina cf. ellisorae Cushman, sample 66, CF5 Zone, Owaina section. D Clavulinoides asper (Cushman), sample 44, CF7c Zone, Owaina section. E Pyramidulina latejugata (Gumble), sample 65, CF5 Zone, Owaina section. F Pyramidulina raphinistrum Linne, sample 70, CF5 Zone, Owaina section. G Frondicularia sp, sample 41, CF8a Subzone, Owaina section. H Saracenaria triangularis (D’Orbigny), sample 56, CF5, Owaina section. I Saracenaria saratogona (D’Orbigny), sample 51, CF6, Owaina section. J Neoflabellina jarvisi (Cushman), sample 41, CF8a Subzone, Owaina section. K Marginulina pachygaster (Gumbel), sample 71, CF5 Zone, Owaina section. L Laevidentalina sp., sample 39, CF9 Zone, Owaina section. M Reussella aegyptiaca (Nakkady), samples 69, CF5 Zones, Owaina section. N Orthokaresteina oveyi (Nakkadi), sample 40, CF8a Subzone, Owaina section. O Orthokaresteina parva (Cushman), sample 46, CF7c Subzone, Owaina section. P Bulimina quadrata (Plummer), sample 44, CF7c Subzone, Owaina section

Scanning Electron Microscope (SEM) of foraminifera. A Cibicidoides alleni (Plummer), sample 71, CF5 Zone, Owaina section. B–D Cibicidoides succedens (Brotzen); B umbilical view, sample 58, CF5 Zone, Owaina section; C lateral view with aperture, sample 58, CF5 Zone, Owaina section; D- spiral view, sample 58, CF5 Zone, Owaina section. E, F Osangularia plummerae (Brotzen); E- lateral view with aperture; sample 41, CF8a Subzone, Owaina section; F umbilical view; sample 41, CF8a Subzone, Owaina section. G–I Anomalinoides umboniferus (Schwager): G Involute view; sample 47, CF6 Zone, Owaina section; H lateral view with aperture; sample 47, CF6 Zone, Owaina section. I Evolute view; sample 47, CF6 Zone, Owaina section; J Heterohelix globulosa (Ehrenberg), sample 65, CF5 Zone, Owaina section. K Heterohelix labellosa Nederbragt, sample 65, CF5 Zone, Owaina section. L, M Heterohelix striata (Ehrenberg), sample 42, CF8b Subzone, Owaina section. N Planoglobulina acervulinoides (Egger), sample 65, CF5 Zone, Owaina section. O, P Planoglobulina carseyae (Plummer), sample 63 and sample 43, CF5 Zone and CF8b Subzone, Owaina section. Q, R Pseudotextularia elegans (Rzehak), sample 54, CF5 Zone; Owaina section. S Pseudoguembelina costulata (Cushman), sample 67, CF5 Zone, Owaina section

Scanning Electron Microscope (SEM) of foraminifera. A Contusotruncana contusa (White), sample 47, CF6 Zone, Owaina section. B–D Gansserina gansseri (Bolli), B spiral view, sample 44, CF7c Subzone, Owaina section; C lateral view with aperture, sample 44, CF7c Subzone, Owaina section; D umbilical view, sample 44, CF7c Subzone, Owaina section. E–G Globotruncana aegyptica Nakkady, E spiral view, sample 46, CF7c Zone, Owaina section; F lateral view with aperture, sample 46, CF7c Zone, Owaina section; G umbilical view, sample 46, CF7c Zone, Owaina section. H–J Globotruncana arca (Cushman), H spiral view, sample 56, CF5 Zone, Owaina section; I lateral view with aperture, sample 58, CF5 Zone, Owaina section; J umbilical view, sample 66, CF5 Zone, Owaina section. K, L Globotruncanita conica (White), K umbilical view, sample 63, CF5 Zone, Owaina section; L lateral view with aperture, sample 64, CF5 Zone, Owaina section. M–O Globotruncanita stuarti (De Lapparent), M- spiral view, sample 63, CF5 Zone, Owaina section; N lateral view with aperture, sample 63, CF5 Zone, Owaina section; O umbilical view, sample 63, CF5 Zone, Owaina section. P Rugoglobigerina macrocephala (Brönnimann), sample 43, CF8b Subzone, Owaina section. Q–S Rugoglobigerina rugosa (Plummer), Q spiral view, sample 53, CF6 Zone, Owaina section; R lateral view with aperture, sample 53, CF6 Zone, Owaina section; S umbilical view, sample 53, CF6 Zone, Owaina section
4 Biostratigraphy
Several zonal schemes for the Late Cretaceous planktic foraminifera (Campanian–Maastrichtian) were proposed in many parts of the world. The planktic zones used here are based on the schemes of [9, 13, 62, 63]. Based on the first occurrence (FO), and last occurrence (LO) of the identified index planktic foraminiferal species, eight planktic foraminiferal zones, and subzones are recognized in the Campanian–Maastrichtian sequence. The range charts of the identified planktic foraminiferal species of the studied section are shown in Fig. 7, while the distribution of the planktic and benthic foraminiferal species counted in Campanian–Maastrichtian is shown in Figs. 8, and 9. The following is the description of the established planktic foraminiferal zones arranged from older to younger.

Range charts of the identified Campanian–Maastrichtian planktic foraminifera

Distribution of the planktic foraminiferal species recorded in Campanian–Maastrichtian

Distribution of the benthic foraminiferal species recorded in Campanian–Maastrichtian
4.1 Campanian–Maastrichtian zones
The predominant zones recognized in this section are five zones only; CF9, CF8, CF7, CF6, and CF5, and three subzones CF 8a, CF 8b, and CF7c. The following zones and subzones discuss from older to younger:
4.1.1 Globotruncanella havanensis (CF9) Zone
Author: Ref. [6].
Definition: The base of the Globotruncanella havanensis Zone designated by [6] is traced at the last occurrence of Radotruncana calcarata.
Thickness: It attains about ~ 9.5 m of the Dakhla Formation from samples 38 to 39 in the Gebel Owaina section (Fig. 7).
Planktic Assemblage: This zone includes Globotruncanella havanensis, other common constituents are Globotruncanella petaloidea, Planoglobulina multicamerata, Contusotruncana walfischensis, Globotruncanita stuarti, Glo. pettersi, and Gublerina sp., the latter two species just appeared before the upper boundary and at the uppermost top respectively. Globigerinelloides alvarezi, Glob. bollii, Globotruncana arca, G. linneiana, Heterohelix globulosa, H. striata, and Pseudotextularia elegans are quite common throughout the zone.
Benthic Assemblage: Bulimina quadrata, Lenticulina sp., Cibicidoides pseudoacutus, C. pseudoperlucides, Gyroidinoides subangulatus. Anomalinoides praeacutus and A. acuta.
Equivalent and age: This zone is equivalent to the Globotruncanella subcrinatus (CF9) Zone [8, 9, 64], this zone belonged to the late Campanian according to [9].
4.1.2 Globotruncana aegyptiaca (CF8a) subzone
Definition: It is defined as the interval from the (FO) of Globotruncana aegyptiaca Nakkady to the (FO) of Gansserina gansseri.
Thickness: It attains about ~ 19.5 m of the Dakhla Formation from samples 40 to 41 at the Gebel Owaina section (Fig. 7).
Planktic Assemblage: In addition to the nominate species Globotruncana aegyptiaca; other common constituents include Heterohelix planata, H. globulosa, H. striata, H. reussi, Pseudotextularia nuttalli, P. elegans, Archaeoglobigerina cretacea, A. blowi, and Rugoglobigerina rugosa.
Benthic Assemblage: nearly the same as in the previous zone, in addition to the presence of Osangularia plummerae.
Equivalent and age: This zone is probably equivalent to the Rugoglobigerina hexacamerata zones of [9, 64]. Therefore, this zone is assigned to late Campanian.
Remarks: The (FO) of Globotruncana aegyptiaca was considered the marker of the base of the earliest Maastrichtian age while others considered the (FO) Globotruncana aegyptiaca to lie within the late Campanian.
In Tunisia, [65] used this zone as a marker for the late Campanian age, but [6, 66, 67] assigned this zone to the early Maastrichtian. Globotruncana aegyptiaca Zone is conformably overlain by the Gansserina gansseri zone.
4.1.3 Rugoglobigerina hexacamerata (CF8b) subzone
Author: Ref. [9].
Definition: This subzone is defined as the interval from the (FO) of the R. hexacamerata (Brönnimann) to the (FO) of Gansserina gansseri (Bolli).
Thickness: It attains about ~ 3.50 m of the Dakhla Formation from samples 42 to 43 at the Gebel Owaina section (Fig. 7).
Planktic Assemblage: In addition to the nominate species Rugoglobigerina hexacamerata; other common constituents include R. macrocephala, R. rugosa, Globotruncana arca, G. aegyptiaca, G. orientalis, G. bulloides, Globotruncanita stuarti, Gl. stuartiformis, Heterohelix reussi, H. navarroensis, H. globulosa and H. striata.
Benthic Assemblage: Bulimina quadrata, Lenticulina sp., Cibicidoides pseudoacutus, Gyroidinoides subangulatus, Anomalinoides midwayensis, A. praeacutus, A. acuta, and Gaudryina ellisorae.
Equivalent and age: This zone is probably equivalent to the R. hexacamerata zone of [13, 44], and Rug. rotundata [10, 68], and is assigned to the late Campanian, and R. hexacamerata (CF8b) of the earliest Maastrichtian (71.0–70.39 Ma) of [69].
Remarks: This zone is equivalent to the upper part of Globotruncana lapparenti tricarinata Zone of [70], the upper part of Globotruncana tricarinata Zone of [71], the upper part of Globotruncana tricarinata Zone of [72], the upper part of G. stuartiformis Zone of [1], the upper part of G. falsostuarti Zone of [5, 73], the upper part of G. havanensis Zone of [74], the upper part of Globotruncana aegyptiaca Zone of [5, 6], the upper part of G. aegyptiaca Zone (CF8) of [8, 62]. Also, it is equivalent to the Rugoglobigerina hexacamerata Subzone (CF8b) of [9].
The FO of R. hexacamerata is utilized by numerous authors in different localities to detect the Campanian/Maastrichtian boundary (C/M) [75,76,77].
4.1.4 Gansserina gansseri (CF7) zone
Author: Ref. [39].
Definition: This zone is defined from the (FO) of Gansserina gansseri (Bolli) to the (FO) of Abathomphalus mayaroensis.
Remarks: Ref. [78] divided the Gansserina gansseri Zone into three subzones: the late Campanian Rugoglobigerina rotundata subzone (CF7a), and the two early Maastrichtian Rugoglobigerina scotti Subzone (CF7b) and Planoglobulina acervulinoides Subzones (CF7c). In the existing work, the (FO) of Gansserina gansseri was reported in sample No. 43.
4.1.4.1 Planoglobulina acervulinoides (CF7c) subzone
Definition: This subzone spans the interval between the (FO) of Planoglobulina acervulinoides and the (FO) of Contusotruncana contusa at the top.
Thickness: It attains about ~ 7 m of the Dakhla Formation from samples 44 to 46 at the Gebel Owaina section (Fig. 7).
Planktic Assemblage: The predominant species of the zone are Globotruncana orientalis, G. mariei, G. linneiana, Archaeoglobigerina blowi, Rugoglobigerina rugosa, R. scotti, R. macrocephala, Planoheterohelix planata, and Laeviheterohelix dentata.
Benthic Assemblage: Gaudryina ellisorae, Ga. rugosa, Spiroplectinella spectabilis, Bulimina quadrata, Lenticulina sp., Cibicidoides alleni, C. pseudoacutus, Gyroidinoides subangulatus, Gy. girardanus, Anomalinoides midwayensis, and Osangularia plummerae.
Equivalent and age: This subzone is probably equivalent to the Gansserina gansseri [6, 70].
Age: Early Maastrichtian.
Remarks: Remarks [79] utilized the FO of Planoglobulina acervulinoides as a biostratigraphic marker and located it within the upper part of the G. gansseri Zone. It was also recorded at a relatively similar position in several localities including Kalaat Senan in Tunisia [45, 78]; Black Nose [63]; Zumaia, northern Spain [13]; [80] and Exmouth Plateau of the northwestern Australian margin [81]. Ref. [82] assigned the FO of this species marks the upper part of the early Maastrichtian. Here we use the Planoglobulina acervulinoides to mark the C/M boundary [78], where the planktic foraminiferal assemblages in this subzone are indicative of the early Maastrichtian age.
This subzone can be correlated with the upper part of the standard G. gansseri Zone, and the heterohelicids show a gradual diversification expressed by the emergence of complex multiserial and coarsely striate forms (e.g., Racemiguembelina powelli) [8, 79].
4.1.5 Contusotruncana contusa (CF6) Zone
Author: Ref. [83].
Definition: It is defined from the (FO) of Contusotruncana contusa at the base and the (LO) of the Globotruncana linneiana (D’ Orbigny) at the top of the biozone. This zone is equal to the CF6 Zone of [8, 9, 62].
Thickness: It attains about ~ 10 m of the Dakhla Formation from samples 47 to 53 at the Gebel Owaina section (Fig. 7).
Planktic assemblage: In addition to the nominate species Contusotruncana contusa; other common constituents include Globotruncanita conica (White), Gansserina gansseri (Bolli), and Heterohelix globulosa (Ehrenberg).
Benthic Assemblage: Gaudryina ellisorae, Gaudryina pyramidata, Gaudryina rugosa, Spiroplectinella spectabilis, Orthokarestenia oveyi, Orthokarestenia parva, Bulimina quadrata, Lenticulina sp., Cibicidoides alleni, C. pseudoacutus, C. succedens, Gyroidinoides subangulatus, Gy. girardanus, Anomalinoides midwayensis, A. praeacutus, A. acuta, Osangularia plummerae, and Oridorsalis plummerae.
Equivalent and age: This zone is probably equivalent to the Contusotruncana contusa Zone of [8, 44, 62, 62, 84, 85]. Thus, this zone is assigned to the late early Maastrichtian.
4.1.6 Pseudotextularia intermediata (CF5) Zone
Author: Ref. [79].
Definition: This zone is defined from the (LO) of the Globotruncana linneiana at the base to the (FO) of Racemiguembelina fructicosa at the top, [79].
Thickness: It attains about ~ 31.50 m within the Dakhla Formation from samples 54 to 71 at the Gebel Owaina section (Fig. 7).
Planktic assemblage: In addition to the nominate species Pseudotextularia intermediate, other common constituents include Pseudotextularia elegans (Rzehak), Globotruncana arca (Cushman), Heterohelix globulosa (Ehrenberg), and Globotruncanita conica (White).
Benthic Assemblage: Gaudryina ellisorae, Gaudryina pyramidata, Gaudryina rugosa, Spiroplectinella spectabilis, Orthokarestenia oveyi, Or. parva, Bulimina quadrata, Lenticulina sp., Cibicidoides alleni, C. pseudoacutus, C. succedens, Gyroidinoides subangulatus, Gy. girardanus, Anomalinoides midwayensis, A. praeacutus, A. acuta, Osangularia plummerae, Valvulineria scrobiculata, and Marginulinopsis tuberculate.
Equivalent and age: This zone is probably equivalent to the upper part of the Gansserina gansseri Zone of [5, 6, 65, 86], the Ps. intermedia Zone (CF5) of [8, 9, 62]. In Egypt, this biozone is equivalent to the upper-middle part of the Gansserina gansseri Zone of [49, 55, 87,88,89,90,91,92]. It is assigned to the late early Maastrichtian age.
Remarks: Ref. [9] used the top of this biozone to place the early/late Maastrichtian boundary in Tunisia. In this study, this zone is equivalent to the upper middle part of the G. gansseri Zone of [5, 6, 65, 86], the Ps. intermedia Zone (CF5) of [8, 9, 62].
5 Results
5.1 Benthic parameters
Even more surprisingly, the total foraminifera number (TFN) values reflect the state of water depth, usually the increase of the water depth, the increase in TFN. The benthic foraminifera number (B) is a useful proxy to estimate oxygen content and organic matter flux in the past [e.g., 93, 94]. The (B) generally decreases in oxygen-depleted sediments [95, 96]. In contrast, a higher (B) is observed with increasing organic matter flux to the seafloor [e.g., 94, 97, 98]. Consequently, the (B) is controlled by both oxygen content and organic matter flux. According to [99], the benthic Foraminifera Number is inversely related to water depth. The Benthic foraminiferal parameters of the studied successions would be briefly discussed below:
In the Owaina section, samples 38–41 (CF9 Zone-CF8a Subzone) the TFN values increase from 265 to 306 individuals. TFN values decrease from 265 to 261 individuals in the CF8b Subzone and CF7 Zone then increases again upward in CF6 and CF5 Zones to reach a maximum value of 367 individuals in sample 61 and at the end of the CF5 Zone.
The benthic number (B) ranges from 3 to 305 individuals in samples 38–70, while samples 1–37 are barren. In samples, 38–41 (B) values increase from 182 to 233 individuals (CF9 Zone—CF8a Subzone). (B) values decrease from 233 to 66 individuals in the CF8b Subzone and CF7 Zone then increases again upward in CF6 and CF5 zones to reach a maximum value of 305 individuals in sample 56 at the end of the CF5 Zone (Fig. 10).

Benthic and planktic foraminiferal parameters in the Owaina section
5.1.1 Diversity Indices
In the present study, biodiversity is measured by many diversity indices, such as species richness, Fisher’s alpha, and Shannon–Weaver. In general, the species diversity increases with paleo-water depth [100,101,102].
5.1.2 Species richness (S)
In the Owaina section, the species richness of benthic foraminiferal tests ranged from barren to 46 species per sample. It is worth mentioning that samples, 1–38, and 72–74, are barren intervals but other samples range from 2 to 46 species/sample. The species richness of benthic foraminifera in samples 39–42 (CF9 Zone, CF8a, CF8b subzones) ranges from 19 to 26 species per sample. In samples, 43 to 53 (CF7, CF6 zones) species richness of benthic foraminifera tests is increasing upward from 18 to 25 species/sample. In samples, 54 to 71 (CF5 Zone) species richness of benthic foraminifera are fluctuating from 14 to 28 species/sample at the end of the CF5 Zone (Fig. 10).
5.1.3 Fisher α values
In the present study, the Fisher α values are ranging from 0.856 to 15.32 in samples 38–70. The highest values are ranging from 7 to 15.32 in samples 39 to 43 (CF9 Zone, CF8a, CF8b, CF7c subzones). The lowest values are ranging from less than 1 to 7 in samples 45 to 71 of zones CF6 and CF5 (Fig. 10).
5.1.4 Arenaceous/Calcareous (A/C %)
The A/C% benthic foraminiferal ratio of the present study is illustrated in (Fig. 10). Ref. [103] stated that the dominance of calcareous foraminifers indicates deposition largely above the CCD line, in an area high in calcium carbonate, well-oxygenated, and characterized by normal salinity and/or high temperature. The calcareous foraminiferal tests are dominated by irregular distribution ranging from dominant ranging from 63 to 100%.
5.1.5 Epifaunal/Infaunal Ratio (E/I):
Several studies have demonstrated the relationship between the different microhabitats of benthic foraminifera and the epifaunal % [e.g., 94, 104, 105]. The Epifaunal % (E/I) is ranging from 8 to 94% in samples 38 and 128. The dominance of epifaunal species often indicates high oxygen conditions and/or low nutrient concentration [106].
5.1.6 Benthic foraminiferal Morphogroups
The benthic foraminifera distribution, abundance, and evolution are governed by several parameters such as depth, the flux of organic nutrients to the seafloor, oxygen levels of the bottom, salinity, food quality, and quantity, substrate, temperature, and others [35,36,37, 102, 107, 108]. The beds of the Owaina section yielded two agglutinated genera and more than nine calcareous benthic genera. The morphogroups scheme for benthic foraminifera used in this work was adopted after [108], where five morphogroups are observed in the Owaina section (Fig. 11).

Distribution of benthic foraminiferal morphogroups in the Owaina section
5.1.6.1 Paleoecology of benthic foraminifera
The presence of Valvulineria in the CF5 Zone inhabited inner shelf to upper slope environments [109], where the abundance of Cibicidoides in the CF5 Zone as epifaunal form and recognized in the middle-outer shelf to upper slope deposits [95, 110, 111]. Lenticulina presents in all zones and represents epifaunal to deep infaunal microhabitats and is observed in the sublittoral to upper bathyal [110, 112]. Orthokarestenia and Bulimina. suggests that they are shallow to deep infaunal, and live in the neritic to the upper bathyal environment [111, 112]. These faunas are indicative of a well-oxygenated environment and are usually determined by increased organic flux [113, 114].
Gaudryina and Spiroplectinella, these taxa are in shallow to deep infaunal conditions, with low levels of oxygen, and are found in a wide range of marine environments from the inner shelf to upper bathyal [106].
5.2 Planktic parameters
5.2.1 Planktic percent (P%)
Planktic percent (P%) or the index of oceans of a foraminiferal population (Gibson, 1989) indicates an inner shelf (10-50 m) characterizes by rare planktic (< 8%) with low species diversity. The relative P% of 8–25% characterizes the middle shelf (50-100 m) with increased species diversity. On the other hand, the outer shelf environment (100-200 m) is characterized by a high P% of 70%, while the P% of 90% is attributed to the middle slope (400-800 m) [115].
The P% is ranging from 1 to 99% in samples 39–71 (Fig. 10). In the Dakhla Formation, the P% ranges from 1 to 20% in samples 70, and 71. In the lower part of the studied section, P% is decreasing from 31 to 20% in samples 39–41 (CF8a Subzone). In CF8b Subzone and CF7 Zone (samples 42–43), P% increases from 37–75%. P% oscillates again in samples 44- 53 (CF6 Zone). In CF5 Zone P% fluctuates (30–40%) but it drops to 1% in sample 70 (Fig. 10).
The planktic number (P) is ranging from 1 to 312 individuals in samples 39–70. In samples, 39–41 (P) values decrease from 83 to 60 individuals (CF9 Zone-CF8a Subzone). (P) values increase from 96 to 195 individuals in the CF8b Subzone and CF7c Subzone then decrease again upward in CF6 and CF5 zones to reach a minimum value of 2 individuals in sample 70 and at the end of CF5 Zone, it is barren except sample 72 (110 individuals).
5.2.2 Planktic foraminiferal morphogroups
Planktic foraminifera tests are adapted to changes in temperature, nutrient supply, and oxygen content [116]. We applied the classification of comparable Cretaceous morphotypes by [117,118,119,120,121], whereas Fig. 12 shows the distribution of planktic morphogroups in the Owaina section. In this matter, three groups can be recognized; (1) Shallow-water fauna (0–50 m) epipelagic is characterized by non-keeled tests of the genera, Rugoglobigerina, Globigerinelloides, Pseudotextularia, and Heterohelix with no peripheral keel, (2) Intermediate-water fauna (50–100 m) mesopelagic as a single-keeled and includes genera Globotruncanella, Globotruncanita, and Gansserina, with only one peripherally keeled, and (3) Deeper-water fauna (with adults normally below 100 m) bathypelagic and has a double-keeled test (Contusotruncana, and Globotruncana) [118, 119]. Ref. [122] reported that Globotruncana arca is more common in deeper water deposits. Ref. [123] found that Contusotruncana is a shallower dweller. During the Cretaceous time the warmest temperature is recorded in tests of rugoglobigerinids and globigerinellids [123, 124].

Distribution of planktic morphogroups in the Owaina section
5.3 Campanian/Maastrichtian boundary
[78] divided the Gansserina gansseri Zone into three subzones; the late Campanian Rugoglobigerina rotundata Subzone (CF7a), and the two early Maastrichtian Rugoglobigerina scotti Subzone (CF7b) and Planoglobulina acervulinoides Subzone (CF7c). In the present work, the authors use the first occurrence of Planoglobulina acervulinoides to mark the C/M boundary. The planktic foraminiferal assemblages in the subzone are proposed as indicative of the early Maastrichtian age. These three subzones across the upper Campanian and Maastrichtian correlate with the previously proposed zonal schemes for the Tethyan realm [125, 126] (Fig. 13).

A comparison of the most relevant planktic foraminiferal biozonations across the upper Campanian and Maastrichtian biozonations is proposed in this study
At the end of the Campanian, some planktic foraminifera became extinct, which might have been related to two volcanic events in the Atlantic Ocean, the Sierra Leone Rise, and the Maud Rise [127]. These had been accompanied by a regional oxygen-reduction event [128] occurring throughout the Campanian, and was accompanied by third-order global sea level falls [9, 120].
The beginning of the Maastrichtian was a period of high turnover, where the planktic foraminifera taxa were highly developed with morphological features such as double peripheral keels that allowed them to sink into deeper waters [129]. The diversity and size of the foraminifera increased during the Maastrichtian pointing to warm temperatures and stable environments [130].
6 Conclusion
Here we use the Planoglobulina acervulinoides Subzone (CF7c) to mark the Campanian/Maastrichtian boundary. The planktic foraminiferal assemblages in the subzone are proposed as indicative of the early Maastrichtian age. This zone is probably equivalent to the Gansserina gansseri [6, 68].
The occurrence of Orthokarstenia and Lenticulina and well-preserved in the Campanian sediments of Gebel Owaina indicate the deepening of the sea, where. this fauna is indicative of a well-oxygenated environment. The abundant occurrence of Orthokarstenia clarki in the late Campanian suggests a middle to outer shelf slightly deeper than 100 m.
The highly double-keeled planktic foraminiferal assemblages of the Maastrichtian sediments of the Owaina section indicate deep-water deposition. Meanwhile, the increased numbers of well-preserved benthic foraminiferal assemblage Cibicidoides and Gyroidinoides in the CF6 Zone imply a tropical to sub-tropical climate.
Data availability
Data will be made available on reasonable request.
References
Postuma JA (1971) Manual of planktonic foraminifera. Elsevier Publishing Company, Amsterdam, p 420
Sigal J (1977) Essai de zonation du Crétacé méditerranéan á laide des foraminiferés planktonique. Géol Méditerr 4(2):99–108
Salaj J (1980) Microbiostratigraphie du Crétacé et du Paléogène de la Tunisie septentrionale et orientale (Hypostratotypes Tunisiens). State Geological Institute of Dionyz Stur, Bratislava (in French)
Bellier JP (1983) Foraminifères planctoniques du Crétacé de Tunisie septentrionale: Systématique, Biozonation, Utilisation stratigraphique de l'Albien au Maastrichtien. (Ph.D.). Université Pierre et Marie Curie, Paris, France (in French)
Robaszyński F, Caron M, Gonzalez Donoso M, Wonders AAH (1984) Atlas of Late Cretaceous Globotruncanids. Rev Micropaleontol 26:145–305
Caron M (1985) Cretaceous planktic foraminifera, in Plankton Stratigraphy. In: Bolli HM, Sautiders JB, Perch-Nielsen K, Caron M, Homewood P (eds) Evolution of early planktonic foraminifera marine micropaleontology, vol 7. Cambridge Univ. Press, Cambridge, pp 453–462
Rami A, Zaghbib-Turki D, El Ouardi H (1997) Biostratigraphie (foraminifères) et contrôle tectono-sédimentaire du Crétacé supérieur dans la région de Mejez el bab (Tunisie septentrionale). Géol Méditerr 24:101–123 (in French)
Li L, Keller G (1998) Abrupt deep-sea warming at the end of the Cretaceous. Geology 26:995–998
Li L, Keller G, Stinnesbeck W (1999) The Late Campanian and Maastrichtian in northwestern Tunisia: paleoenvironmental inferences from lithology, macrofauna and benthic foraminifera. Cretac Res 20:231–252
Odin GS (2001) The Campanian-Maastrichtian boundary: Definition at Tercis (Landes, SW France) principle, procedure, and proposal. In: Odin GS (ed) The Campanian-Maastrichtian stage boundary: Characterisation at Tercis-Les-Bains (France) and correlation with Europe and other continents. Developments in palaeontology and stratigraphy, vol 19. Elsevier, Amsterdam, pp 820–833
Odin GS, Lamaurelle MA (2001) The Global Campanian-Maastrichtian stage Boundary. Episodes 24:229–238
Ogg JG, Ogg G, Gradstein FM (2008) The concise geologic time scale. Cambridge University Press, p 150
Arz JA, Molina E (2002) Bioestratigrafíay cronoestratigrafía con foraminíferos planctónicos del Campaniense superior y Maastrichtiense de latitudes subtropicales y templadas (España, Francia, y Tunicia). Neues Jb Geol Paläontol Abh 224:161–195
Clarke LJ, Jenkyns HC (1999) New oxygen isotope evidence for long-term Cretaceous climatic change in the Southern Hemisphere. Geology 27:699–702
Zachos JC, Dickens GR, Zeebe RE (2008) An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 451:279–283
Friedrich O, Norris RD, Erbacher J (2012) Evolution of middle to Late Cretaceous oceans—a 55 m.y. record of Earth’s temperature and carbon cycle. Geology 40:107–110
Ando A, Woodard SC, Evans HF, Littler K, Herr-Mann S, MacLeod KG, Kim S, Khim B-K, Robinson SA, Huber BT (2013) An emerging palaeoceanographic ʻmissing linkʼ: Multidisciplinary study of rarely recovered parts of a deep-sea Santonian-Campanian transition from Shatsky Rise. J Geol Soc 170:381–384
Linnert C, Robinson SA, Lees JA, Brown R, Pérez-Rodríguez I, Petrizzo MR, Falzoni F, Littler K, Arz A, Russel EE (2014) Evidence for global cooling in the Late Cretaceous. Nat Commun 5:4194. https://doi.org/10.1038/ncomms5194
Falzoni F, Petrizzo MR, Clarke LC, MacLeod KG (2016) Long-Term Cretaceous carbon and oxygen isotope trends and planktonic foraminiferal turnover: A new record from the southern mid-latitudes. Geol Soc Am Bull 128:1725–1735
Huber BT, MacLeod KG, Watkins DK, Coffin MF (2018) The rise and fall of the Cretaceous Hot Greenhouse climate. Glob Planet Change 167:1–23
Barrera E, Savin SM (1999) Evolution of Campanian-Maastrichtian marine climates and oceans. In: Barrera E, Johnson CC (eds) Evolution of the Cretaceous Ocean-Climate System, vol 332. Geological Society of America. Special Paper, pp 245–282
Huber BT, Norris RD, MacLeod KG (2002) Deep-sea paleotemperature record of extreme warmth during the Cretaceous. Geology 30:123–126
Cramer BS, Toggweiler JR, Wright JD, Katz ME, Miller KG (2009) Ocean overturning since the Late Cretaceous: inferences from a new benthic foraminiferal isotope compilation. Paleoceanography 24:PA4216. https://doi.org/10.1029/2008PA001683
Robinson SA, Murphy DP, Vance D, Thomas DJ (2010) Formation of “Southern Component Water” in the Late Cretaceous: Evidence from Nd-isotopes. Geology 38:871–874
MacLeod KG, Londoño CI, Martin EE, Jiménez Berrocoso Á, Basak C (2011) Changes in North Atlantic circulation at the end of the Cretaceous greenhouse interval. Nat Geosci 4:779–782
Martin EE, MacLeod KG, Jiménez Berrocoso Á, Bourbon E (2012) Water mass circulation on Demerara Rise during the Late Cretaceous based on Nd isotopes. Earth Planet Sci Lett 327:111–120
Robinson SA, Vance D (2012) Widespread and synchronous change in deep-ocean circulation in the North and South Atlantic during the Late Cretaceous. Paleoceanography. https://doi.org/10.1029/2011PA002240
Jung C, Voigt S, Friedrich O, Koch MC, Frank M (2013) Campanian-Maastrichtian Ocean circulation in the tropical Pacific. Paleoceanography 28:1–12. https://doi.org/10.1002/palo.20051
Voigt S, Jung C, Friedrich O, Frank M, Teschner C, Hoffmann J (2013) Tectonically restricted deep-ocean circulation at the end of the Cretaceous greenhouse. Earth Planet Sci Lett 369–370:169–177
Moiroud M, Pucéat E, Donnadieu Y, Bayon G, Guiraud M, Voigt S, Deconinck JF, Monna F (2016) Evolution of neodymium isotopic signature of seawater during the Late Cretaceous: Implications for intermediate and deep circulation. Gondwana Res 36:503–522
Donnadieu Y, Pucéat E, Moiroud M, Guillocheau F, Deconinck J-F (2016) A better-ventilated ocean triggered by Late Cretaceous changes in continental configuration. Nat Commun 7:10316. https://doi.org/10.1038/ncomms10316
Gusic I, Jelaska V, Velic I (1988) Foraminiferal assemblages, facies, and environments in the upper cretaceous of the island of Brac, Yugoslavia. Revue de Palébiologie 2 (Benthos’86), 447–456
Velic I, Vlahovic I (1994) Foraminiferal assemblages in the Cenomanian of the Buzet-Savudrija area (northwestern Istria, Croatia). Geol Croat 47(1):25–43
Cause E, Bernaus JM, Boix C, Calonge A, Perez R (2003) Upper cretaceous shallow benthic biozones (KSBZ): a preliminary report. In: AAPG International Conference Barcelona, Spain, 21–24
Koutsoukos EAM, Hart MB (1990) Cretaceous foraminiferal morphogroup distribution patterns, palaeocommunities and trophic structures: a case study from the Sergipe Basin, Brazil. Trans R Soc Edinb Earth Sci 81(3):221–246
Setoyama E, Kaminski MA, Tyszka J (2011) The Late Cretaceous-Early Paleocene palaeobathymetric trends in the southwestern Barents Sea-palaeoenvironmental implications of benthic foraminiferal assemblage analysis. Palaeogeogr Palaeoclimatol Palaeoecol 307:44–58
Setoyama E, Kaminiski MA, Tyszka J (2017) Late Cretaceous–Paleogene foraminiferal morphogroups as palaeoenvironmental tracers of the rifted Labrador margin, northern proto-Atlantic. In: Kaminski MA, Alegret L (eds) Proceedings of the Ninth International Workshop on Agglutinated Foraminifera, vol 22. Grzybowski Foundation Special Publication, pp 179–220.
Said R (1962) The Geology of Egypt. Elsevier Publication Co., Amsterdam, p 377
Said R, Sabry H (1964) Planktonic foraminifera from the type locality of the Esna shale in Egypt. Micropaleontology 3:375–395
El-Naggar ZRM (1966) Stratigraphy and classification of type Esna Group of Egypt. Bull Am Assoc Pet Geolog 50:1455–1477
El-Younsy ARM, Obaidalla NA, Philobbos ER, Salman AM (2015) Depositional environments and sequence stratigraphy of the Campanian-Eocene succession, Gabal Oweina area, Upper Egypt. The 8th International Conference on the Geology of Africa, Assiut University, Egypt, I, pp 73–97
El-Younsy ARM, Nageh A, Obaidalla NA, Philobbos ER, E. R., Salman, A. M., (2017) High-resolution sequence stratigraphy of the Upper Cretaceous-Lower Paleogene succession, Gabal Qreiya area, Upper Egypt. Arab J Geosci 10:531. https://doi.org/10.1007/s12517-017-3319-z
Farouk S, Aaskalany M, El-Sorogy A, Youssef M, Taha S (2019) Maastrichtian-early Paleocene foraminiferal palaeobathymetry and depositional sequences at Gebel ElSharawna, south Luxor, Egypt. Lethaia 53:316–331
Brönnimann P (1952) Trinidad Paleocene and Lower Eocene Globigerinidae. Bull Am Paleontol 34(143):1–34
Robaszyński F, Caron M (1995) Cretaceous planktonic foraminifera: Comments on the Europe-Mediterranean zonation. Bull Soc Géolog Fr 166:681–692
Arz JA (1996) Los foraminiferos planctonicos del Campaniense Maastrichtiense. Bioestratigrafia, cronoestratigrafia y eventos paleoecologicos. (Ph.D.). Universidad de Zaragoza, Zaragoza, Spain (in Spanish)
Robaszyński F, González-Donoso JM, Linares D, Amédro F, Caron M, Dupuis C, Gartner SJM (2000) Le Crétacé supérieur de la region de Kalaat Senan, Tunisie Centrale. Litho-biostratigraphie intégrée: Zones d’ammonites, deforaminifers planctoniques et de nannofossiles du Turonien supérieur au Maastrichtien. Bulletin des Centres de Recherche et d’Explor-Prod Elf-Aquitaine 22:359–490
Said R (1990) The geology of Egypt. Balkema, Rotterdam, p 721
Kerdany MT, Cherif OH (1990) Mesozoic. In: Said R (ed) The geology of Egypt. Balkema, Rotterdam, pp 407–438
Guiraud R, Bosworth W (1999) Phanerozoic geodynamic evolution of northeastern Africa and the northwestern Arabian platform. Tectonophysics 315:73–108
Guiraud R, Bosworth W, Thierry J, Delplanque A (2005) Phanerozoic geological evolution of Northern and Central Africa: an overview. J Afr Earth Sci 43:83–143
Issawi B (1972) Review of Upper Cretaceous-Lower Tertiary stratigraphy in central and southern Egypt. AAPG Bull 56:1448–1463
Youssef (1957) Upper Cretaceous rocks in Kosseir area. Bull Inst Desert Egypt 7(2):35–53 (Cairo)
Luger P (1985) Stratigraphie der marinen Oberkreide und des Alttertiärs im südwestlichen Obernil-Beeken (W-Ägypten) under besonderrer Berücksichtigung der Mikropaläontologie, Palökologie und Paläogeographie. Berliner geowissenschaftlichen Abhandlungen (63), 151 pp., 22 fig., 11 tab., 24 pl., Berlin.
Farouk S, Ahmed F, Powell JH, Marzouk A (2016) Integrated microfossil biostratigraphy, facies distribution, and depositional sequences of the upper Turonian to Campanian succession in northeast Egypt and Jordan. Facies 62:8
Said R (1961) Tectonic framework of Egypt and its influence on distribution of foraminifera. AAPG Bull 45:198–218
El Gammal RMH, Orabi HO (2019) Coniacian-late Campanian planktonic events in the Duwi formation, Red Sea region, Egypt. J Geol Geophys 8(1):16
El-Naggar, Z.R.M., 1970. On a proposed lithostratigraphic subdivision for the late cretaceous-early Paleocene succession in the Nile Valley, Egypt. In: Proceedings of the Arab petroleum congress, Kuwait, 1–64.
Abdel Razik TM (1972) Comparative studies on the upper Cretaceous early Paleogene sediments on the Red Sea coast, Nile Valley and Western Desert, Egypt. In: Proceedings of the 8th Arab petroleum congress, Algeria. 71(B-3): 1–23
Philobbos ER (1996) The phosphatic sediments of the Nile Valley and Eastern Desert in the view of the Upper Cretaceous —lower tertiary sedimentation tectonics. Geol Soc Egypt Spec Publ 2:313–352
Fatela F, Taborda R (2002) Confidence limits of species proportions in microfossil assemblages. Mar Micropaleontol 45:169–174. https://doi.org/10.1016/S0377-8398(02)00021-X
Li L, Keller G (1998) Maastrichtian climate, productivity and faunal turnover in planktic foraminifera in south Atlantic DSDP Sites 525 A and 21. Mar Micropaleontol 33:5–86
Huber BT, Macleod KG, Tur N (2008) Late Campanian-Maastrichtian chronostratigraphic framework for Campanian– Maastrichtian sediments on Blake Nose (subtropical North Atlantic). J Foraminiferal Res 38:162–182
Al-Kahtany K (2017) Planktic foraminiferal biostratigraphy of the Maastrichtian-Paleocene succession at the north Farafra Oasis, Western Desert, Egypt. Arab J Geosci 10:10. https://doi.org/10.1007/s12517-017-3128-4
Keller G (1988) Biotic turnover in benthic foraminifera across the Cretaceous/Tertiary boundary at El Kef, Tunisia. Palaeogeogr Palaeoclimatol Palaeoecol 66:153–171
Bralower TJ, Leckie RM, Sliter WV, Thierstein HR (1995) An integrated Cretaceous microfossil biostratigraphy. In: Berggren WA, Kent DV, Aubry MP, Hardenbol J (eds) Geochronology, time scales and global stratigraphic correlation, vol 54. SEPM Special Publication, Tulsa, pp 65–79
Guerrero J, Sarmiento G, Navarrete R (2000) The stratigraphy of the W side of the Cretaceous Colombian Basin in the Upper Magdalena Valley. Reevaluation of selected areas and type localities including Aipe, Guaduas, Ortega, and Piedras. Geolog Colomb 25:45–110
Odin GS (2002) Définition d’une limite multicritère; stratigraphie du passage Campanien-Maastrichtien du site géologique de Tercis (Landes, SW France). CR Geoscience 334:409–414 (in French)
Farouk S (2014) Maastrichtian carbon cycle changes and planktonic foraminiferal bioevents at Gebel Matulla, west-central Sinai, Egypt. Cretaceous Res 50:238–251
Bolli HM (1966) Zonation of the Cretaceous to Pliocene marine sediments based on planktonic foraminifera. Chacao 9:3–32
Boersma A (1984) Campanian through Paleocene paleotemperature and carbon isotope sequence and the Cretaceous-Tertiary boundary in the Atlantic Ocean. In: Berggren WA, Van Couvering JA (eds) Catastrophes and earth history. Princeton University, Press, Princeton, pp 247–277
Barr FT (1972) Cretaceous biostratigraphy and planktonic Foraminifera of Libya. Micropaleontology, 1B (1) l-46, pl. 1–10, l0 text fig., New York
Almogi-Labin A, Luz B, Duplessy JC (1986) Quaternary paleo-oceanography, pteropod preservation and stable-isotope record of the Red Sea. Paleogeogr Paleoclimatol Paleoecol 57:195–211
Huber BT (1990) Maastrichtian planktonic foraminifer biostratigraphy of the Maud Rise (Weddell Sea, Antarctica), ODP Leg 113 Hole 689B and 690C. In: Barker PF, Kennett JP (eds) Proceeding of the Ocean Drilling Program, Scientific Results 113, 489-514.3
Zepeda MA (1998) Planktonic foraminiferal diversity, equitability and biostratigraphy of the uppermost Campanian-Maastrichtian, ODP Leg 122, Hole 762, Exmouth Plateau, NW Australia, eastern Indian Ocean. Cretac Res 19:117–152
Tantawy AA, Keller G, Adatte T, Stinnesbeck W, Kassab A, Schulte P (2001) Maastrichtian to Paleocene (Dakhla Formation) depositional environment of the Western Desert in Egypt: Sedimentology, mineralogy and integrated micro and macrofossils biostratigraphies. Cretac Res 22:795–827
Jaff RBN, Wiliams M, Wilkinson IP, Lawa F, Zalasiewicz J (2014) A refind foraminiferal biostratigraphy for the Late Campanian-Early Maastrichtian succession of northeast Iraq. Geo-Arabia 19:161–180
Saïdi E, Zaghbib-Turki D (2016) Planktonic foraminiferal biostratigraphy and quantitative analysis during the Campanian-Maastrichtian transition at the Oued Necham section (Kalâat Senan, central Tunisia). Turk J Earth Sci 25:538–572
Nederbragt AJ (1991) Late Cretaceous biostratigraphy and development of Heterohelicidae (planktic foraminifera). Micropaleontology 37:329–372
Pérez-Rodríguez I, Lees JA, Larrasoaña JC, Arz JA, Arenillas I (2012) Planktonic foraminiferal and calcareous nannofossil biostratigraphy and magnetostratigraphy of the uppermost Campanian and Maastrichtian at Zu - maia, northern Spain. Cretac Res 37:100–126
Thibault N, Harlou R, Schovsbo N, Schiøler P, Minoletti F, Galbrun B, Lauridsen BW, Sheldon E, Stemmerik L, Surlyk F (2012) Upper Campanian-Maastrichtian nannofossil biostratigraphy and high-resolution carbon-isotope stratigraphy of the Danish Basin: towards a standard δ13C curve for the Boreal Realm. Cretaceous Res 33:72–90
Farouk S, Faris M, Bazeen YS, Elamri Z, Ahmad F (2021) Upper Campanian-lower Maastrichtian integrated carbon isotope stratigraphy and calcareous microplankton biostratigraphy of North-central Tunisia. Mar Micropaleontol 166:102003. https://doi.org/10.1016/j.marmicro.2021.102003
Dalbeiez F (1955) The genus Globotruncana in Tunisia. Micropaleontology 1(2):161–171
Allameh M, Nejad RRY (2017) Late Cretaceous biostratigraphy planktonic foraminifera of the Farokhi Formation, Iran. Open J Geol 7:320–334
Sarigül V, Hakyemez A, Tüysüz O, Can Genç Ş, Yilmaz İÖ, Özcan E (2017) Maastrichtian-Thanetian planktonic foraminiferal biostratigraphy and remarks on the K-Pg boundary in the southern Kocaeli Peninsula (NW Turkey). Turk J Earth Sci 26:1–29
D’Hondt S, Keller G (1991) Some patterns of planktic foraminiferal assemblage turnover at the Cretaceous-Tertiary boundary. Mar Micropaleontol 17:77–118
Cherif OH, Hewaidy AA (1986) Maastrichtian foraminifera from four sections in the Dakhla Shale of Central Egypt and their bathymetric implications. In: 3 eme Symposium international Sur Les Foraminifers benthiques, Genève,1–20. 2 photographs
Shahin AM (1988) Tertiary Planktonic Foraminiferal Biostratigraphy and Paleobathymetry at Gebel Withr, Southwestern Sinai, Egypt. Neues Jb Geol Paläontol Abh 209(3):323–348
Shahin A (1992) Contribution to the foraminiferal biostratigraphy and paleopathymetery of the Late Cretaceous and Early Tertiary in west Central Sinai, Egypt. Revue de Micropaleontol 35(2):157–175
Anan HS (1994) Contribution to the stratigraphy and paleobiogeography of some diagnostic Upper Cretaceous and Paleogene foraminifera- Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen 5:257–266
Abdel-Kireem MR, Samir AM, Ibrahim MI (1995) Upper Cretaceous planktonic foraminiferal zonation and correlation in the northern part of the Western Desert, Egypt. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 198, 3, 329–361, Stuttgart
Lüning S, Marzouk AM, Morsi AM, Kuss J (1998) Sequence stratigraphy of the Upper Cretaceous of south-east Sinai, Egypt. Cretaceous Res 19:153–196
Kaiho K, Hasegawa T (1994) End-Cenomanian benthic foraminiferal extinction and oceanic dysoxic events in the northwestern Pacific Ocean. Palaeogeogr Palaeoclimatol Palaeoecol 111:29–43
Jorissen FJ, De Stigter HC, Widmark JGV (1995) A conceptual model explaining benthic foraminiferal microhabitats. Mar Micropaleontol 22:3–15
Coccioni R, Galeotti S (1993) Orbitally induced cycles in benthonic foraminiferal morphogroups and trophic structure distribution patterns from the L Albian Amadeus segment (Central Italy). J Micropaleontol 12:227–239
Friedrich O, Reichelt K, Herrle JO, Lehmann J, Pross J, Hemleben C (2003) Causes for the formation of the Late Aptian Niveau Fallot black shales in the Vocontian Basin (SE France): evidence from planktic and benthic foraminifera and stable isotopes. Mar Micropaleontol 49:65–85
Kaiho K (1994) Benthic foraminiferal dissolved-oxygen index and dissolved-oxygen levels in the modern ocean. Geology 22:719–722
Murray JW (2000) When Does environmental variability become environmental change? The proxy record of Benthic Foraminifera. In: Martin RE (ed) Environmental micropaleontology: the application of microfossils to environmental geology, topics in geobiology, 15. Plenum Press, New York, pp 7–37
Mendes I, Gonzalez R, Dias JMA, Lobo F, Martins V (2004) Factors influencing recent benthic foraminifera distribution on the Guadiana shelf (Southwestern Iberia). Mar Micropaleontol 51:171–192
Murray JW (2006) Ecology and applications of benthic foraminifera. Cambridge University Press
Murray JW, Alve E (2011) The distribution of agglutinated foraminifera in NW European seas: Baseline data for000000000000000 the interpretation of fossil assemblages. Palaeontolog Electron 14(2) (ISSN 1935-3952)
Nagy J, Kaminski MA, Kuhnt W, Bremer MA (2000) Agglutinated foraminifera from neritic to bathyal facies in the Palaeogene of Spitsbergen and the Barents Sea. In: Hart MB, Kaminski MA (eds) Proceedings of the Fifth International Workshop on Agglutinated Foraminifera, vol 7. Grzybowski Foundation Special Publication, 333–361
Saint-Marc P (1986) Qualitative and quantitative analysis of benthic foraminifers in Paleocene deep-sea sediments of the Sierra Leone Rise, central Atlantic. J Foraminiferal Res 16(3):244–253
Corliss BH (1985) Microhabitats of benthic foraminifera within deep-sea sediments. Nature 314:435. https://doi.org/10.1038/314435a0
Corliss BH, Chen C (1988) Morphotype patterns of Norwegian Sea deep-sea benthic foraminifera and ecological implications. Geology 16:716–719
Bernhard JM (1986) Characteristic assemblages and morphologies of benthic foraminifera from anoxic, organic-rich deposits; Jurassic through Holocene. J Foraminifera Res 16:207–215. https://doi.org/10.2113/gsjfr.16.3.207
Nagy J (1992) Environmental significance of foraminiferal morphogroups in Jurassic North Sea deltas. Palaeogeogr Palaeoclimatol Palaeoecol 95:111–134
Stojanova V, Petrov G (2018) Paleoecological significance of benthic foraminiferal fauna from the Ovče Pole Basin. Republic of Maced Geolog Maced 32(1):45–57
Olsson RK (1988) Foraminiferal modeling of sea-level change in the Late Cretaceous of New Jersey. In: Wilgus CK, Hastings BS, Kendall CGSC, Posamentier HW, Ross CA, Van Wagoner JC (eds) Sea level changes: an integrated approach. The Society of Economic Paleontologists and Mineralogists. Special Publication, Tulsa, pp 289–297
Widmark JGV (1997) Deep-sea benthic foraminifera from Cretaceous-Paleogene boundary strata in the South Atlantic -taxonomy and paleoecology. Fossils Strata 43:1–94
Grunert P (2011) Integrated facies analysis and stratigraphy of the Early Miocene North Alpine foreland basin (Upper Austria). Unpublished Ph.D. Thesis, Karl Franzens Universität Graz
Rückheim S, Bornemann A, Mutterlose J (2006) Planktic foraminifera from the mid-Cretaceous (Barremian-early Albian) of the North Sea Basin: Palaeoecological and palaeoceanographic implications. Mar Micropaleontol 58:83–102
Nagy J, Gradstein FM, Kaminski MA, Holbourn AE (1995) Foraminiferal morphogroups, palaeoenvironments and new taxa from Jurassic to cretaceous strata of Thakkhola, Nepal. In: Kaminski MA, Geroch S, Gasiński MA (eds) Proceedings of the Fourth International Workshop on Agglutinated Foraminifera, 3. Grzybowski Foundation Special Publication, pp 181–209
Nagy J, Kaminski MA, Johnsen K, Mitlehner AG (1997) Foraminiferal, palynomorph, and diatom biostratigraphy and paleoenvironments of the Tork Formation: a reference section for the Paleocene-Eocene transition in the western Barents Sea. In: Hass HC, Kaminski MA (eds) Contributions to the micropaleontology and paleoceanography of the northern North Atlantic, vol 5. Grzybowski Foundation Special Publication, pp 15–38
Olsson RK, Nyong EE (1984) A paleoslope model for Campanian—lower Maastrichtian foraminifera of New Jersey and Delaware. J Foramin Res 14:50–68
Bé AWH (1977) An ecological, zoogeographic and taxonomic review of recent planktonic foraminifera. In: Ramsay ATS (ed) Oceanic micropalaeontology. Academic Press, London, p 100
Hart MB, Bailey HW (1979) The distribution of planktonic foraminiferida in the mid-Cretaceous of NW Europe. In: Wiedmann J (ed) Aspekte der Kreide Europas, vol 6. International Union Geological Society, pp 527–542
Hart MB (1980) A water depth model for the evolution of planktonic foraminiferida. Nature 286:252–254
Wonders AAH (1980) Middle and Late Cretaceous planktonic foraminifera of the western Mediterranean area. Utrecht Micropaleontol Bull 24:157
BouDagher-Fadel MK (2013) Biostratigraphic and geological significance of planktonic Foraminifera, 2nd edn. University College, Office of the Vice Provost Research, London, p 271
Leckie RM (1989) A paleoceanographic model for the early evolutionary history of planktonic foraminifera. Palaeogeogr Palaeoclimatol Palaeoecol 73:107–138
Sliter WV (1972) Cretaceous foraminifers-depth habitats and their origin. Nature 239:514–515
Boersma A, Shackleton NJ (1981) Oxygen and carbon isotope variations and planktonic foraminifera depth habitats, Late Cretaceous to Paleocene, Central Pacific, Deep Sea Drilling Project Sites 463 and 465. In: Theide J, Vallier TL et al (eds) Initial Reports of the deep-sea drilling Project 62. U. S Government Printing Office, Washington, pp 513–526
Douglas RG, Savin SM (1978) Oxygen isotopic evidence for the depth stratification of Tertiary and Cretaceous planktic foraminifera. Mar Micropalaeontol 3:175–196
Odin GS, Arz JA, Caron M, Ion J, Molina E (2001) Campanian-Maastrichtian planktonic foraminifera at Tercis les Bains (Landes, France); synthetic view and potential for global correlation. In: Odin GS (ed) The Campanian-Maastrichtian stage boundary: characterisation at Tercis-Les-Bains France) and correlation with Europe and other continents. Developments Les Palaeontology and Stratigraphy 19. Elsevier, Amsterdam, pp 379–395
Arz JA, Molina E (2001) Planktic foraminiferal quantitative analysis across the Campanian/Maastrichtian boundary at Tercis (Landes, France). In: Odin GS (ed) The Campanian-Maastrichtian stage boundary: characterisation at Tercisles-Bains (France) and correlation with Europe and Other Continents. Developments in palaeontology and stratigraphy 19. Elsevier, Amsterdam, pp 339–348
Eldholm O, Coffin MF (2000) Large igneous provinces and plate tectonics. In: Richards MA, Gordon RG, van der Hilst RD (eds) The history and dynamics of global plate motions. American Geophysical Union Geophysical Monograph, 121 American Geophysical Union, Washington, DC, pp 309–326
Walliser OH (1996) Global events and event stratigraphy in the phanerozoic. In: Walliser OH (ed) Global events and event stratigraphy. Springer, Berlin, pp 7–19
Djoulah B, Hamimed M, Belghar N, Boulemia S (2023) Sepiolite as paleo-CO2 barometer of dramatic climate change, and a basic element for paleogeographic reconstitution across Cretaceous-Paleogene, Asserdoune boundary, El Kouif region (Algerian-Tunisian border). Carbonates Evaporites 38:6
Nallamuthu M, Orabi HO, Sharma B, Shashtri C, Srinivas CK (2021) Planktonic foraminiferal quantitative analysis across the Campanian/Maastrichtian boundary at the Pondicherry area, Cauvery Basin, Southern India. Geol J, pp 1–19
Geological Map of Egypt. (1987) NG 36 SW Sheet of Luxor, 1: 500 000. Egyptian General Petroleum Corporation and Conoco, Cairo, Egypt
Acknowledgements
Great thanks are due to Prof Dr. Jere H. Lipps, the University of California for the useful discussion and revised version of the manuscript. We greatly appreciate the organizations that funded our research.
Funding
No funding was received to assist with the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
OHO collecting previous literature—reviewing and continuous follow-up of all parts of the research. AMS participation in determining the title of research—collecting previous studies—writing research content—identifying and organizing references-adjusting research formats in line with the journal.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Ethical approval
Ethical approval is Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Orabi, O.H., El-Sabbagh, A., Mansour, A.S. et al. Foraminifera study for the characterization of the Campanian/Maastrichtian boundary in Gebel Owaina, Nile Valley, Egypt. J.Umm Al-Qura Univ. Appll. Sci. 9, 341–359 (2023). https://doi.org/10.1007/s43994-023-00043-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s43994-023-00043-8