
Abstract
In an effort to ameliorate the impacts of climate change, forest managers in Central Europe increasingly turn to conifer species that produce higher yields and are better adapted to projected future climatic conditions. Though small mammals are an important component of the forest ecosystem, the impacts of enriching native broadleaf forests with conifers on small mammal communities are not well understood. We conducted mark–recapture surveys of small mammals to ascertain differences in their community structure among stands of two conifers (native Norway spruce Picea abies and non-native Douglas fir Pseudotsuga menziesii) and the dominant broadleaf in the region, European beech (Fagus sylvatica). After estimating the density of two common species, the yellow-necked mouse Apodemus flavicollis and bank vole Clethrionomys glareolus, we found that the population density of each is positively related to the proportion of beech and negatively to the proportion of conifers in each stand, though these effects of stand composition are smaller than the positive effect of herb cover. Increasing Norway spruce proportion reduced monthly survival of small mammals, while Douglas fir proportion had a positive effect on survival. We conclude that the two conifer species have similar impacts on small mammal density, though overall small mammal survival was significantly lower on plots with Norway spruce. This suggests that increasing the proportion of Douglas fir at the expense of Norway spruce may be possible without significantly changing local patterns of small mammal population density, but further research is necessary to elucidate the exact impacts of these two conifer species on small mammal demography and behavior.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Stands of Norway spruce (Picea abies) and other conifers were established across much of central Europe over the past two centuries in the face of wood shortages, as they were relatively easy to establish on degraded land and promised increased production volume relative to broadleaf European beech (Fagus sylvatica) forests (Spiecker 2003). In recent years, Norway spruce has fared poorly in the face of drought, heat waves, and pest infestations, leading to extensive sanitary and salvage harvests (Mezei et al. 2017). This has increased forest managers’ interest in other tree species and management methods as they plan replacements, including the creation of multi-species forest stands which may be more resilient to these pressures than traditional monocultures (Griess et al. 2012; Dobor et al. 2020; Schnabel et al. 2021). Cultivating multiple tree species in mixed stands has also been shown to increase biodiversity, water quality, and aesthetic value (Felton et al. 2016), result in higher levels of ecosystem services (Gamfeldt et al. 2013), and can stabilize or even increase timber production (Jucker et al. 2014; Pretzsch and Schütze 2016).
In terms of alternatives to Norway spruce, non-native Douglas fir (Pseudotsuga menziesii), originally from North America, is increasingly utilized due to its high productivity, being well-adapted to predicted future central European climatic conditions and resistant to many common European forest pests (Roques et al. 2006; Vitali et al. 2017). Douglas fir may even reduce pest damage to European beech in mixed stands (Matevski et al. 2023). However, the ecosystem impacts of replacing native Norway spruce with exotic Douglas fir are not fully understood, especially on native fauna (Thomas et al. 2022). Little is known about how Central European small mammals may be affected by increasing cultivation of Douglas fir, which has been found to support different assemblages of birds (Schuldt et al. 2022), arthropods (Goßner and Ammer 2006; Kriegel et al. 2021), and soil microfauna, fungi, and microbes (Podrázský et al. 2020; Likulunga et al. 2021; Lu and Scheu 2021), as compared to Norway spruce.
Yellow-necked mice (Apodemus flavicollis) and bank voles (Clethrionomys glareolus) are two of the most common small mammal species found across Central European forests (Niedziałkowska et al. 2010; Suchomel et al. 2012). Seed and seedling predation by these and other small mammals plays an important role in European beech regeneration patterns (Zwolak et al. 2016b) and has a significant impact on regeneration patterns of many other tree species (Bílek et al. 2009; Wagner et al. 2010). Caching of beech seeds by small mammals, in particular, has been cited as an important component of beech germination success, as winter frost can kill un-cached seeds on the soil surface (Jensen 1985). Both species play an important role in ectomycorrhizal fungal spore dispersal in Central Europe (Schickmann et al. 2012; Komur et al. 2021), providing inoculum of symbiotic fungi to tree seedlings which is crucial for forest regeneration. Pupal predation by small mammals can regulate insect population densities (Kollberg et al. 2014), and bank voles have been found to affect outbreak patterns of forest pests (Hanski and Parviainen 1985). Furthermore, these common small mammals provide an important prey resource to sustain increased predator diversity (Sandom et al. 2013), especially for avian (Salamolard et al. 2000; Sundell et al. 2004) and smaller-bodied mammalian predators such as mustelids and foxes (Dell’Arte et al. 2007; Sundell et al. 2013).
Research on small mammal communities in Central European forests suggests that understory structure and canopy cover are important for many species (Ecke et al. 2001; Benedek et al. 2021). Understory structures such as downed wood, rocks, and vegetation provide small mammals with shelter as well as food (Harmon et al. 1986; Hallett et al. 2003; Juškaitis et al. 2013). The bank vole prefers forests with relatively dense herb cover (Mazurkiewicz 1994), while the yellow-necked mouse prefers closed canopy forests which often have less ground vegetation (Marsh and Harris 2000). Both species inhabit a variety of forests from mature beech stands to young spruce plantations, though interspecific competition may lead to occupancy of less suitable habitats or changes in activity patterns by bank voles (Zárybnická et al. 2017). While both are considered generalist species, yellow-necked mice rely heavily on tree seeds, while bank voles consume more green vegetation including herbs and seedlings when seeds are not available (Selva et al. 2012). Arthropod prey make up a seasonally important part of both species’ diets, especially when breeding (Obrtel 1974).
While it has been shown that deciduous trees support greater small mammal abundance and diversity relative to conifers (Niedziałkowska et al. 2010), few studies investigate the impact of introduced tree species or mixtures on small mammals. To better understand how enriching native European beech forests with conifers impacts small mammals, and to ascertain whether a non-native conifer, Douglas fir, has a different impact than native Norway spruce, we conducted surveys of small mammal communities in managed forests across the state of Lower Saxony in Northern Germany. We assessed the relationship between small mammal densities and forest stand composition across five stand types: pure Douglas fir, pure European beech, pure Norway spruce, and mixtures of European beech with each of the two conifer species.
We addressed two primary questions: (i) Do the densities of the two most common species, the yellow-necked mouse and the bank vole, vary between pure beech, mixed beech-conifer, and pure conifer stands, or with the proportions of each conifer in a stand? (ii) Does the apparent survival of each species vary between stand types or with the proportion of each of the conifer species?
We expected that (i) pure conifer stands would have lower yellow-necked mouse densities than pure beech or mixed stands, due to their seed-focused diet, while bank vole densities would be similar across stand types, and that densities of both species in non-native Douglas-fir stands would be reduced compared to stands with Norway spruce. Similarly, we expected that stands with a lower proportion of conifers, and thus a higher proportion of European beech, would have a higher density of yellow-necked mice along the gradient of increasing beech proportion, while expecting bank vole densities to be less affected by tree species proportion and more influenced by herb cover. We expected that both species would again have relatively reduced densities in stands with non-native Douglas fir as compared to stands with similar proportions of Norway spruce. (ii) With regard to apparent survival, we expected that pure conifer stands would have lower apparent survival rates than mixed or pure beech stands, due to the higher availability of seeds from beech masting in pure beech and mixed stands, and that survival would be highest in stands with high proportions of native beech and lowest in stands with high proportions of non-native Douglas fir.
Methods
Study area
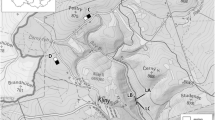
Our study was conducted within the framework of the Research Training Group 2300 project in the Northern German state of Lower Saxony, in state-managed forests ranging from the Solling uplands and Harz mountains in the south to lowland areas near Nienburg in the northwest and the Elbe River in the northeast (Fig. 1). The climate in Northern Germany is cool and wet punctuated by hot, dry weeks over the summer, but varies with elevation and latitude. The southern half of our study area is characterized by loamy soils, a mean annual temperature of 8.5 °C, elevations from 277 to 493 m and 815 to 1029 mm annual rainfall. The northern half has sandy soils, a mean annual temperature of 9.3 °C, ranges in elevation from 47 to 138 m and receives from 672 to 746 mm of annual rainfall (Foltran et al. 2023).

Study area and location of study sites (black diamonds) within the federal state of Lower Saxony (dark gray), Germany (light gray). Lower right inset depicts the arrangement of the five study plots at our site in the Harz mountains
Forests in Lower Saxony encompass over 1.2 million hectares, and are managed for wood production, hunting, and other outdoor recreation. Pine species make up the largest proportion (Pinus spp. 28.6%), followed by spruce (Picea spp. 16.4%), beech (Fagus sylvatica 13.5%), and oak (Quercus spp. 12.3%). Douglas fir is present in a limited area, making up only 2.4% of the forested area, but has increased its share over the past decades relative to other conifers (Thünen-Institut 2012). Forests are typical of managed stands in Central Europe and Germany, with mature trees of a single age-class.
To obtain a representative sample of forests across our study area, eight study sites were established, half in the northern and half in the southern portion, with each site consisting of five 0.5-hectare study plots. Each of these plots, in turn, represents one of five forest stand types: (1) pure Douglas fir, (2) a mixture of Douglas fir and European beech, (3) pure European beech, (4) a mixture of European beech and Norway spruce, and (5) pure Norway spruce. All stands selected for our study plots were older than 60 years, and each species in mixed stands contributed over 20% of the total basal area. Each quintet of five plots that make up a study site were located as close to each other as possible, usually within a few kilometers (Fig. 1), while maintaining a minimum separation of 100 m between plots.
Small mammal surveys
At each of our 40 study plots, a small mammal survey grid was established, with trapping points spaced 10 m apart in an eight-by-eight configuration covering approximately 0.5 ha, with a minimum distance of 50 m between survey grids within a site. Small mammal surveys were conducted at all sites during the summer (July, August, and early September) of 2018, 2019, and 2020. Additionally, a subset of sites was surveyed in May and June of 2018 and 2019. Each survey lasted four nights, with traps set on the evening of the first day and removed after being checked the morning of the fifth day; all five plots at a site were surveyed concurrently. During each survey, one Sherman live trap (LFA, 8 cm × 9 cm × 23 cm) was set at each of the trapping points, for a total 64 active traps per plot. An exception was the summer 2019 survey, where a subset of the grid in a 6 × 6 configuration of 36 traps covering ¼ hectare was used. Traps were baited with a mixture of sunflower seeds, oats, millet, peanuts, raisins, and peanut butter; a handful of natural wool at the back of each trap provided nesting material. Traps were checked at least twice per day, once at dawn and once in the late afternoon, with a third check added at mid-day if the maximum temperature was forecast to exceed 30 °C.
Captured small mammals were identified to species and weighed, had their hind-foot measured, and were visually inspected to determine sex, age, and breeding status. We additionally measured head–body length and tail length and recorded any further morphometric measurements diagnostic for differentiating between species when necessary. These additional measurements were primarily used to discriminate between wood mice (Apodemus sylvaticus) and yellow-necked mice (Jenrich et al. 2010). Each animal had a small tissue sample taken from the ear, was injected with a passive integrated transponder (PIT) tag (Peddymark 1.4 mm × 10 mm ISO 11784/5 FDX-B) for identification, and had a small patch of fur on the right rear haunch trimmed as a secondary mark. Individuals recaptured during the same survey session had only their PIT tag number read and recorded; inter-session recaptures were evaluated as a new capture, less tissue sampling and marking.
Forest stand and vegetation surveys
The location of each tree on the plot relative to the plot center was recorded, and the diameter at breast height (DBH) of each tree was measured. All trees were identified to species, and the heights of a random sample of each tree species were measured. These stand surveys were updated yearly to account for any tree falls or other changes between years, but most stands did not experience significant changes in tree composition on the timescale of our study. Vegetation surveys were conducted on all plots by dividing the half-hectare into quarters of an eighth-hectare each, and visually estimating the percentage of herbaceous cover within each of these sub-plots. The mean of these sub-plot surveys was used as a plot-level estimate of herb cover.
Stand structure
To obtain a more realistic estimate of stand composition by tree species that is based on the area occupied by individual trees, we utilized the area potentially available (APA) measure and calculated the proportion of the plot occupied by each tree species (Dirnberger et al. 2017). The area potentially available is a spatially explicit index, taking into account the interaction of each tree in with its neighbors and the competitive ability of each species. The competitive ability was approximated with the 95% quantile of the crown radius (Pretzsch et al. 2015; Glatthorn 2021a). We used the statistical software application R (R Core Team 2022) to perform this and all subsequent data transformations and analyses. APA proportions for each of the three tree species were calculated using the APAtree package (Glatthorn 2021b) for R as described in Glatthorn (2021a).
Density estimation
We estimated the density of adult small mammals on our plots using spatially explicit capture–recapture (SECR) methods (Efford et al. 2009) as implemented in the R package secr version 4.5.7 (Efford 2022a) throughout. Spatially explicit measures allow for estimation of density, expressed as the number of individuals per hectare, that are comparable between studies with different survey methods and effort (Tourani 2022). Using the capture locations of each individual to estimate the probability of detection and scale of movement around their home range centers, SECR models can directly estimate the effective sampling area and account for behavioral differences between species or study sites that may otherwise lead to over- or under-estimation of population densities (Efford and Fewster 2013).
We conducted a model-selection procedure, described below, to determine the best model for each of the two most common small mammal species, as well as for all small mammals without regard to species identity. As there may be differences in animal behavior between study sites, best models were selected separately for each site.
Density, with the exception of the null model, was always set to vary by trapping session, as this was the primary output variable of interest and is known to vary by season and year. A half-normal detection function was used to describe the decline in detection probability with increasing distance from an individual’s home range center. Models considering the impact of learned and transient responses to previous capture, trapping location, and previous capture of an individual at a specific trapping location (implying a trap-happy or trap-shy individual) on detection parameters g0 (probability of detection) and/or sigma (spatial scale parameter) were tested. Additionally, a time-of-day factor variable on the occasion level, corresponding to whether a capture period occurred during the day or overnight, was included in our model evaluation. A null model with density (D), g0, and sigma all being invariant across trapping sessions was included as a reference.
The models with the lowest AICc value were selected for each species and site combination, and these models were then used to generate all subsequent density estimates (see Supplementary Table 1).
Survival estimation
Apparent monthly survival rates were estimated for all marked adult small mammals, as well as for adult yellow-necked mice and bank voles using non-spatial Jolly–Seber–Schwarz–Arnason (JSSA) models (Schwarz and Arnason 1996; Efford and Schofield 2020) implemented in the R package openCR version 2.2.5 (Efford 2022b). We allowed survival (phi) to vary between surveys, and evaluated models which held encounter probability (p) and per-capita recruitment (f) constant, or allowed them to vary between surveys, and chose the best model for each plot based on the models’ AICc values (Supplementary Table 2). Only half of the sites, which were surveyed twice per summer in both 2018 and 2019, were included, as the species considered here are relatively short-lived and a few individuals survive between years. Due to extremely low small mammal population density during the spring survey of 2018, we only considered survival estimates from the summer of 2019 in subsequent analyses.
Statistical analyses
To assess the impact of stand composition on small mammal densities, we used negative binomial generalized linear mixed models; the negative binomial distribution is typically used for count data that is overdispersed (Lindén and Mäntyniemi 2011). Primary fixed covariates were either stand type (categorical with five levels) or APA proportion of Douglas fir and Norway spruce (each a proportion between 0 and 1). We considered both the stand type and proportion of conifers, because mixture proportions varied and we were interested both in the overall effect of stand type and the more precise impact of each conifer proportion on small mammal density.
We included herb cover (continuous between 0, i.e., total lack of herbaceous ground vegetation and 1, i.e., complete coverage of the ground by herbaceous vegetation) to account for its influence on small mammal density (Suchomel et al. 2012). Survey campaign was included as a random effect to incorporate temporal dependency, and site and plot as nested random effects to account for the spatial dependency among observations.
To evaluate the impact of stand composition on apparent monthly survival rates of small mammals, we used beta generalized linear models; the beta distribution is appropriate for a proportion such as survival rate (Damgaard and Irvine 2019). Due to the lower sample size of our apparent survival estimates, we could not include random effects. As in our density models, stand type or proportion of Douglas fir and Norway spruce were our primary fixed effects, while we included herb cover and site as additional fixed effects to account for the impact of herb cover and geographic location on survival. We used the R package glmmTMB (Brooks et al. 2017) to fit our models, as it allows for the use of the negative binomial and beta distributions, as well as the flexibility to specify zero-inflation and dispersion models which are often important to account for variance patterns in field data.
Model assumptions were checked using diagnostic plots from the DHARMa package (Hartig 2022), which uses simulation to create interpretable residuals for fitted GLMMs; we ran 1000 simulations for each of our fitted models and used these for model diagnostics. To ensure there was no multicollinearity between fixed effects terms, we used the variance inflation factor (VIF), as implemented in the performance package (Lüdecke et al. 2021), and considered a VIF < 3 to indicate low correlation between fixed effects (Zuur et al. 2010).
We estimated both the marginal (proportion of total variance explained by fixed effects) and conditional (proportion of total variance explained by both fixed and random effects) R2 for each negative binomial linear mixed model per Nakagawa et al. (2017), as implemented in the MuMIn package (Bartoń 2023), and the marginal R2 for each beta linear model per Efron (1978), as implemented in the performance package.
To assess the significance of each fixed effect, we used likelihood ratio tests to compare a model with and without each effect; though as we were interested in the effect of each predictor on density and survival, we did not remove non-significant terms. To determine the relative effect and significance of each stand type on the density and apparent survival of small mammals, we conducted post hoc pairwise Tukey tests; to determine the effect of conifer proportion on the same, we applied post hoc general linear hypothesis tests, each as implemented in the multcomp (Hothorn et al. 2008) package. We used the R packages ggplot2 (Wickham 2016) and ggeffects (Lüdecke 2018) to visualize our results.
Results
Trapping results
Total trapping effort from 2018 to 2020, after accounting for sprung or otherwise inactive traps, was 35,759 trap nights and 27,097 trap days. 3885 individuals of 7 species were captured over the course of the study, with the majority of captured individuals (3692) belonging to two common species: the yellow-necked mouse (47% of marked individuals) and the bank vole (48% of marked individuals) (Table 1). Additionally, 378 shrews (Soricidae spp.) were captured, though shrews were not targeted by our trapping methods and we did not mark them. There was no evidence that species richness or Shannon diversity varied between stand types.
Inter-year, seasonal, and regional effect on small mammal density
Small mammal density varied widely between years and seasons. Density of both yellow-necked mice and bank voles was lowest in the spring of 2018 (mean ± SD, 0.92 ± 1.67 mice/ha, 2.6 ± 3.49 voles/ha) and highest in the spring (voles) and summer (mice) of 2019 (mean ± SD, 40.46 ± 25.58 mice/ha, 29.51 ± 23.22 voles/ha). Surveys earlier in the summer generally found lower densities than surveys later in the summer. We found large differences in density between northern and southern plots within years as well, with bank voles generally present at higher densities in the southern plots, and yellow-necked mice at higher densities at the northern plots (Table 2).
Effect of stand type and APA proportion on small mammal density
Stand type was a significant component in models of yellow-necked mouse density and small mammal density overall, though not in bank vole density models. Relative to pure beech stands, pure stands of both Douglas fir and Norway spruce had a significant negative effect on yellow-necked mouse density, while mixed stands of Douglas fir and beech (estimate ± SE = 0.42 ± 0.14, z = 3.04, p = 0.020) and Norway spruce and beech (estimate ± SE = 0.45 ± 0.16, z = 2.87, p = 0.033) had a significant positive effect on yellow-necked mice relative to their respective pure conifer stands. Mixed Douglas fir/beech plots also had a positive effect on yellow-necked mouse density relative to pure Norway spruce plots (estimate ± SE = 0.53 ± 0.14, z = 3.71, p = 0.002). We did not find significant relationships between stand type pairs and bank vole density nor overall small mammal density (Fig. 2, Table 3a).

Model predicted density (ind/ha) and 95% confidence intervals for a yellow-necked mice (A. flavicollis) and b bank voles (C. glareolus) for each stand type. Stars represent significant differences at the levels (***) p < 0.001, (**) p < 0.01, (*) p < 0.05. Predictions were conditioned on mean herb cover
We found strong evidence that the proportion of Douglas fir or Norway spruce was an important component of our models for the density of yellow-necked mice, as well as small mammal density overall, while the proportion of each conifer was not a significant component in our bank vole density models (Table 3b). Yellow-necked mouse and overall small mammal densities were negatively affected by increasing proportion of Douglas fir or Norway spruce, while bank vole density showed a similarly negative but not significant trend (Fig. 3).

Model predicted density (ind/ha) and 95% confidence intervals for yellow-necked mice (A. flavicollis, a, b) and bank voles (C. glareolus, c, d) in relation to the APA proportion of Douglas fir (red lines) or Norway spruce (blue lines) in the left two panels (a, c), and to herb cover proportion in the right two panels (b, d). Solid lines indicate significant relationships (p < 0.05), dashed lines indicate non-significant relationships. Predictions based on herb cover effects were conditioned on average spruce and Douglas APA proportions, and predictions based on conifer APA proportions were each conditioned on mean herb cover and zero APA proportion for the other conifer
Herb cover was a significant component in all density models for yellow-necked mice, bank voles, and small mammals overall, whether including categorial stand type or continuous conifer proportions (Table 3).
Effect of stand type and APA proportion on summer apparent survival rates
Stand type was a significant component in models of monthly apparent survival rates for bank voles and small mammals overall, but not yellow-necked mice. Norway spruce stands had a significant negative effect on overall small mammal apparent survival relative to stands of beech (estimate ± SE = − 0.84 ± 0.25, z = − 3.39, p = 0.006), Douglas fir (estimate ± SE = − 1.10 ± 0.23, z = − 4.72, p < 0.001), and mixed Douglas-beech (estimate ± SE = − 0.74 ± 0.22, z = − 3.34, p = 0.007), and on bank vole apparent survival relative to Douglas fir (estimate ± SE = − 1.62 ± 0.37, z = − 4.41, p < 0.001) and mixed Douglas-beech (estimate ± SE = − 1.09 ± 0.29, z = − 3.78, p = 0.001) stands (Table 4a).
The proportion of Norway spruce had a significant negative effect on apparent survival of yellow-necked mice, bank voles, and small mammals overall, while the proportion of Douglas fir had a significant positive effect on bank vole and overall small mammal apparent survival (Table 4b). Interestingly, despite having a larger effect on density than stand composition, herb cover did not appear to have an impact on apparent survival (Table 4, Fig. 4).

Model predicted apparent survival rates and 95% confidence intervals for yellow-necked mice (A. flavicollis, a–c) and bank voles (C. glareolus, d–f) by stand type (a, d), tree APA proportion (b, e) of Douglas fir (red) and Norway spruce (blue), or herb cover proportion (c, f). Stars represent significant differences at the levels (***) p < 0.001, (**) p < 0.01, (*) p < 0.05. Dashed lines represent non-significant relationships (p > 0.05). Predictions based on tree APA proportions were each conditioned on mean herb cover and a proportion of zero for the other conifer
Discussion
We surveyed small mammal communities for 3 years at 40 plots across Northern Germany to assess the relationship between these communities and the composition of managed forest stands. Overall, we found that small mammal density, in particular of yellow-necked mice, was affected negatively by conifers relative to European beech. Small mammal apparent survival, especially of bank voles, was negatively affected by Norway spruce, while Douglas fir had either no significant impact on survival (yellow-necked mice) or a significant positive impact (bank voles, all species). Herb cover had a positive effect on the density of all species, but no clear effect on apparent survival.
We observed a strong effect of year and season on small mammal density, the cyclical boom-bust nature of which is well documented (Korpimäki et al. 2004; Lambin et al. 2006). Our first surveys in the summer of 2018 revealed very low densities of all species on nearly every plot, even late in the summer when populations normally reach their yearly peak. In the autumn of 2018, beech forests across Northern Germany experienced a strong masting event, which led to a substantial increase in small mammal densities the following year. By the summer of 2020, the density of small mammals was greatly reduced compared to 2019, but still noticeably higher than in 2018 (Table 2).
Effects of stand composition on small mammal density
Despite the significant effect of stand composition on the density of yellow-necked mice and small mammals overall, stand composition had only weak effects on the density of bank voles, the density of which was most affected by herb cover. The strong negative effect of both conifers on the density of yellow-necked mice (Fig. 3a) supported our first hypothesis; this is likely related to the proportion of high-caloric value tree seeds in their diet (Selva et al. 2012) and their well-known affinity for forests with deciduous seed-bearing trees (Niedziałkowska et al. 2010). We found that mixed stands supported densities of yellow-necked mice that were not significantly different than pure beech stands (Fig. 2a), which suggests that mixing conifers with native beech may allow for increased timber production and forest resilience to climate change while avoiding a large decrease in yellow-necked mouse populations.
While we did see a slight negative effect of both conifers on bank vole density, (Fig. 3c), it was not strongly affected by stand composition, supporting our hypothesis that bank vole density would be similar across stand types. The wide diet breadth of bank voles, which is uniquely intermediate and overlapping between European arvicoline and murine rodents (Butet and Delettre 2011), likely allows bank voles to fully exploit available resources in each stand type.
Norway spruce and Douglas fir had similar effects on small mammal density, which did not support our hypothesis that Douglas fir would have a more negative impact on densities than Norway spruce. This was unexpected for several reasons. Most conifers produce defense compounds that can reduce the palatability of their seeds and seedlings to rodent predators (Lobo 2014), and Norway spruce produces different defensive compounds than Douglas fir (Mamoci et al. 2022); the novelty of compounds present in non-native Douglas fir tissues could result in lower consumption of Douglas fir seeds compared to those of Norway spruce. In addition, Norway spruce stands are preferred by foraging red squirrels (Sciurus vulgaris) over silver fir (Abies alba) stands, suggesting that rodents may prefer Norway spruce seeds over the seeds of other conifers (Rubino et al. 2012). Though a study in Norway found that abundances of the most common small mammal species were lower in non-native spruce stands relative to stands of native birch, and that abundances in mixed stands were similar to native birch stands (Pedersen et al. 2010), this could be attributed to the differences between coniferous and broadleaf species rather than introduced vs. native species. Despite these indications that Norway spruce could be preferred to non-native Douglas fir, our results suggest that the differences between the two conifers do not translate into differential impacts on small mammal population densities.
Though the importance of herb cover to many small mammal species is well known (Ecke et al. 2001), and both yellow-necked mouse and bank vole densities increased significantly with increasing herb cover on our plots (Fig. 3), we found that, as hypothesized, herb cover has a stronger effect on bank voles than yellow-necked mice. This is likely due to green vegetation making up a much greater proportion of the bank vole’s diet as compared to the yellow-necked mouse, which depends more heavily on seeds year-round (Selva et al. 2012). The positive effect of understory herbs, especially in bank voles, can mask the negative effect of conifers on small mammal density; it is therefore important to consider the impact of understory herb cover or a similar understory cover measure in addition to stand composition. The more open canopy, and thus higher herb cover, found in managed conifer forests is primarily due to silviculture practices, specifically thinning, which has been found to influence understory cover more than tree species (Augusto et al. 2003). These silvicultural choices, therefore, likely have a large impact on the small mammal community in managed forests.
Effects of stand composition on apparent survival
We found that Norway spruce had a significant negative effect on the apparent survival of small mammals on our plots (Fig. 4). This did not support our hypothesis that stands of both conifers would have lower apparent survival rates than beech or mixed stands, nor that apparent survival would be lowest in Douglas fir stands. Interestingly, the apparent survival of small mammals was similar in stands of both Douglas fir and European beech, as well as their mixture. We expected apparent survival to be significantly higher in stands with higher proportions of beech, especially after the previous autumn’s major beech mast, but this was not supported by our data. It is important to note, however, that apparent survival does not allow for differentiation between emigration and death, and small mammals in Norway spruce stands could simply exhibit lower site fidelity than those in Douglas fir or European beech stands.
While the positive effect of Douglas fir proportion on survival was unexpected, the lack of a clear relationship between herb cover and survival was even more surprising given its importance in predicting small mammal population density. Zwolak et al. (2016a) found that both yellow-necked mice and bank voles had weaker associations with vegetative cover following a mast year when abundance was high, suggesting that when a rich food resource such as beech mast is available, understory vegetation may have less impact on survival. As we were only able to consider apparent survival following a mast year, this shift to beech seeds as a primary food source could explain the weak relationship between herb cover and survival in our data.
Given the similar negative effect of both conifers on small mammal density, the differences between the two conifers with regard to apparent survival were unexpected, though an Italian study on the effects of forestry practices on the density and survival of yellow-necked mice and bank voles also found that survival and density estimates were not always clearly linked (Gasperini et al. 2016). These differences between survival and density patterns suggest that plots with a higher proportion of Douglas fir may simply have a lower carrying capacity than plots with a high proportion of beech, while plots with a high proportion of Norway spruce could be population sinks for some small mammal species.
To investigate this further, we included per-capita recruitment in our apparent survival models and found that, in the case of bank voles, there was a significant increase in the per-capita recruitment rate with increasing Norway spruce proportion (Supplementary Table 5). As bank vole apparent survival exhibited the strongest negative response to Norway spruce proportion, the corresponding higher recruitment rate suggests that one plausible explanation for the observed differences in density and apparent survival patterns is Norway spruce stands serving as population sinks.
Conclusions
In central European managed forests, Douglas fir in pure and mixed stands with European beech is likely to support a community of common fossorial small mammals similar to that found in pure or mixed stands of Norway spruce. Though other ecological differences between the two conifers may give pause to managers considering Douglas fir, there does not appear to be any clear difference between them with regard to the population density of common small mammals. Reduced small mammal apparent survival rates in stands including Norway spruce relative to Douglas fir or European beech warrant further investigation, and could indicate differences in the underlying demographic processes of small mammal communities in Norway spruce stands. Further studies are needed to confirm our findings and continue the process of discerning the ecological impacts of non-native tree species in central European managed forests, especially with regards to under-studied groups such as small mammals.
Data availability
The datasets generated during this study are available upon reasonable request from the corresponding author.
References
Augusto L, Dupouey J-L, Ranger J (2003) Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann For Sci 60:823–831
Bartoń K (2023) MuMIn: multi-model inference. R package version 1.47.5. https://CRAN.R-project.org/package=MuMIn. Accessed 22 Mar 2023
Benedek AM, Sîrbu I, Lazăr A (2021) Responses of small mammals to habitat characteristics in Southern Carpathian forests. Sci Rep 11:12031
Bílek L, Remeš J, Zahradník D (2009) Natural regeneration of senescent even-aged beech (Fagus sylvatica L.) stands under the conditions of Central Bohemia. J For Sci 55:144–145
Brooks ME, Kristensen K, van Benthem KJ, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Maechler M, Bolker BM (2017) GlmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9:378–400
Butet A, Delettre YR (2011) Diet differentiation between European arvicoline and murine rodents. Acta Theriol 56:297–304
Damgaard CF, Irvine KM (2019) Using the beta distribution to analyse plant cover data. J Ecol 107:2747–2759
Dell’Arte GL, Laaksonen T, Norrdahl K, Korpimäki E (2007) Variation in the diet composition of a generalist predator, the red fox, in relation to season and density of main prey. Acta Oecol 31:276–281
Dirnberger G, Sterba H, Condés S, Ammer C, Annighöfer P, Avdagić A, Bielak K, Brazaitis G, Coll L, Heym M, Hurt V, Kurylyak V, Motta R, Pach M, Ponette Q, Ruiz-Peinado R, Skrzyszewski J, Šrámek V, de Streel G, Svoboda M, Zlatanov T, Pretzsch H (2017) Species proportions by area in mixtures of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.). Eur J For Res 136:171–183
Dobor L, Hlásny T, Zimová S (2020) Contrasting vulnerability of monospecific and species-diverse forests to wind and bark beetle disturbance: the role of management. Ecol Evol 10:12233–12245
Ecke F, Löfgren O, Hörnfeldt B, Eklund U (2001) Abundance and diversity of small mammals in relation to structural habitat factors. Ecol Bull 49:165–171
Efford M (2022a) secr: spatially explicit capture-recapture models. R package version 4.5.7. https://CRAN.R-project.org/package=secr. Accessed 8 Oct 2022
Efford M (2022b) OpenCR: open population capture-recapture models. R package version 2.2.5. https://CRAN.R-project.org/package=openCR/. Accessed 25 Sept 2022
Efford MG, Fewster RM (2013) Estimating population size by spatially explicit capture-recapture. Oikos 122:918–928
Efford MG, Schofield MR (2020) A spatial open-population capture-recapture model. Biometrics 76:392–402
Efford MG, Borchers DL, Byrom AE (2009) Density estimation by spatially explicit capture–recapture: likelihood-based methods. In: Thomson DL, Cooch EG, Conroy MJ (eds) Modeling demographic processes in marked populations. Springer, Boston, pp 255–269
Efron B (1978) Regression and ANOVA with zero-one data: measures of residual variation. J Am Stat Assoc 73:113–121
Felton A, Nilsson U, Sonesson J, Felton AM, Roberge J-M, Ranius T, Ahlström M, Bergh J, Björkman C, Boberg J, Drössler L, Fahlvik N, Gong P, Holmström E, Keskitalo ECH, Klapwijk MJ, Laudon H, Lundmark T, Niklasson M, Nordin A, Pettersson M, Stenlid J, Sténs A, Wallertz K (2016) Replacing monocultures with mixed-species stands: ecosystem service implications of two production forest alternatives in Sweden. Ambio 45:124–139
Foltran EC, Ammer C, Lamersdorf N (2023) Do admixed conifers change soil nutrient conditions of European beech stands? Soil Res. https://doi.org/10.1071/SR22218
Gamfeldt L, Snäll T, Bagchi R, Jonsson M, Gustafsson L, Kjellander P, Ruiz-Jaen MC, Fröberg M, Stendahl J, Philipson CD, Mikusiński G, Andersson E, Westerlund B, Andrén H, Moberg F, Moen J, Bengtsson J (2013) Higher levels of multiple ecosystem services are found in forests with more tree species. Nat Commun 4:1340
Gasperini S, Mortelliti A, Bartolommei P, Bonacchi A, Manzo E, Cozzolino R (2016) Effects of forest management on density and survival in three forest rodent species. For Ecol Manag 382:151–160
Glatthorn J (2021a) A spatially explicit index for tree species or trait diversity at neighborhood and stand level. Ecol Indic 130:108073
Glatthorn J (2021b) APAtree: computation of the area potentially available (APA) to trees. R package version 1.0.1. https://doi.org/10.5281/zenodo.5204412. Accessed 17 Aug 2021
Goßner M, Ammer U (2006) The effects of Douglas-fir on tree-specific arthropod communities in mixed species stands with European beech and Norway spruce. Eur J For Res 125:221–235
Griess VC, Acevedo R, Härtl F, Staupendahl K, Knoke T (2012) Does mixing tree species enhance stand resistance against natural hazards? A case study for spruce. For Ecol Manag 267:284–296
Hallett JG, O’Connell MA, Maguire CC (2003) Ecological relationships of terrestrial small mammals in western coniferous forests. In: Zabel CJ, Anthony RG (eds) Mammal community dynamics: management and conservation in the coniferous forests of western North America. Cambridge University Press, Cambridge, pp 120–156
Hanski I, Parviainen P (1985) Cocoon predation by small mammals, and pine sawfly population dynamics. Oikos 45:125
Harmon ME, Franklin JF, Swanson FJ, Sollins P, Gregory SV, Lattin JD, Anderson NH, Cline SP, Aumen NG, Sedell JR, Lienkaemper GW, Cromack K, Cummins KW (1986) Ecology of coarse woody debris in temperate ecosystems. In: MacFadyen A, Ford ED (eds) Advances in ecological research. Academic Press, Cambridge, pp 133–302
Hartig F (2022) DHARMa: residual diagnostics for hierarchical (multi-level/mixed) regression models. R package version 0.4.6. https://CRAN.R-project.org/package=DHARMa. Accessed 8 Sept 2022
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biom J 50:346–363
Jenrich J, Löhr P-W, Müller F, Lang J (2010) Kleinsäuger: körper- und schädelmerkmale, ökologie, beiträge zur naturkunde in Osthessen. Imhof, Petersberg
Jensen TS (1985) Seed-seed predator interactions of European beech, Fagus silvatica and forest rodents, Clethrionomys glareolus and Apodemus flavicollis. Oikos 44:149
Jucker T, Bouriaud O, Avacaritei D, Coomes DA (2014) Stabilizing effects of diversity on aboveground wood production in forest ecosystems: linking patterns and processes. Ecol Lett 17:1560–1569
Juškaitis R, Balčiauskas L, Šiožinytė V (2013) Nest site selection by the hazel dormouse Muscardinus avellanarius: is safety more important than food? Zool Stud 52:53
Kollberg I, Bylund H, Huitu O, Björkman C (2014) Regulation of forest defoliating insects through small mammal predation: reconsidering the mechanisms. Oecologia 176:975–983
Komur P, Chachuła P, Kapusta J, Wierzbowska IA, Rola K, Olejniczak P, Mleczko P (2021) What determines species composition and diversity of hypogeous fungi in the diet of small mammals? A comparison across mammal species, habitat types and seasons in Central European mountains. Fungal Ecol 50:101021
Korpimäki E, Brown PR, Jacob J, Pech RP (2004) The puzzles of population cycles and outbreaks of small mammals solved? Bioscience 54:1071
Kriegel P, Matevski D, Schuldt A (2021) Monoculture and mixture-planting of non-native Douglas fir alters species composition, but promotes the diversity of ground beetles in a temperate forest system. Biodivers Conserv 30:1479–1499
Lambin X, Bretagnolle V, Yoccoz NG (2006) Vole population cycles in northern and southern Europe: is there a need for different explanations for single pattern? J Anim Ecol 75:340–349
Likulunga LE, Rivera Pérez CA, Schneider D, Daniel R, Polle A (2021) Tree species composition and soil properties in pure and mixed beech-conifer stands drive soil fungal communities. For Ecol Manag 502:119709
Lindén A, Mäntyniemi S (2011) Using the negative binomial distribution to model overdispersion in ecological count data. Ecology 92:1414–1421
Lobo N (2014) Conifer seed predation by terrestrial small mammals: a review of the patterns, implications, and limitations of top-down and bottom-up interactions. For Ecol Manag 328:45–54
Lu J-Z, Scheu S (2021) Response of soil microbial communities to mixed beech-conifer forests varies with site conditions. Soil Biol Biochem 155:108155
Lüdecke D (2018) ggeffects: tidy data frames of marginal effects from regression models. J Open Source Softw 3:772
Lüdecke D, Ben-Shachar MS, Patil I, Waggoner P, Makowski D (2021) Performance: an R package for assessment, comparison and testing of statistical models. J Open Source Softw 6:3139
Mamoci E, Andrés MF, Olmeda S, González-Coloma A (2022) Chemical composition and activity of essential oils of Albanian coniferous plants on plant pests. Chem Proc 10:15
Marsh ACW, Harris S (2000) Partitioning of woodland habitat resources by two sympatric species of Apodemus: lessons for the conservation of the yellow-necked mouse (A. favicollis) in Britain. Biol Conserv 92:275–283
Matevski D, Foltran E, Lamersdorf N, Schuldt A (2023) Introduction of non-native Douglas fir reduces leaf damage on beech saplings and mature trees in European beech forests. Ecol Appl. https://doi.org/10.1002/eap.2786
Mazurkiewicz M (1994) Factors influencing the distribution of the bank vole in forest habitats. Acta Theriol 39:113–126
Mezei P, Jakuš R, Pennerstorfer J, Havašová M, Škvarenina J, Ferenčík J, Slivinský J, Bičárová S, Bilčík D, Blaženec M, Netherer S (2017) Storms, temperature maxima and the Eurasian spruce bark beetle Ips typographus—an infernal trio in Norway spruce forests of the Central European High Tatra Mountains. Agric For Meteorol 242:85–95
Nakagawa S, Johnson PCD, Schielzeth H (2017) The coefficient of determination R2 and intra-class correlation coefficient from generalized linear mixed-effects models revisited and expanded. J R Soc Interface 14:20170213
Niedziałkowska M, Kończak J, Czarnomska S, Jędrzejewska B (2010) Species diversity and abundance of small mammals in relation to forest productivity in northeast Poland. Écoscience 17:109–119
Obrtel R (1974) Comparison of animal food eaten by Apodemus flavicollis and Clethrionomys glareolus in a lowland forest. Zool Listy 23:35–46
Pedersen ÅØ, Yoccoz NG, Ims RA, Sigurdsen T (2010) Effects of non-native spruce plantations on small mammal communities in subarctic birch forests. For Ecol Manag 260:331–338
Podrázský V, Kupka I, Prknová H (2020) Substitution of Norway spruce for Douglas-fir: changes of soil microbial activities as climate change induced shift in species composition—a case study. Cent Eur For J 66:71–77
Pretzsch H, Schütze G (2016) Effect of tree species mixing on the size structure, density, and yield of forest stands. Eur J For Res 135:1–22
Pretzsch H, del Río M, Ammer Ch, Avdagic A, Barbeito I, Bielak K, Brazaitis G, Coll L, Dirnberger G, Drössler L, Fabrika M, Forrester DI, Godvod K, Heym M, Hurt V, Kurylyak V, Löf M, Lombardi F, Matović B, Mohren F, Motta R, den Ouden J, Pach M, Ponette Q, Schütze G, Schweig J, Skrzyszewski J, Sramek V, Sterba H, Stojanović D, Svoboda M, Vanhellemont M, Verheyen K, Wellhausen K, Zlatanov T, Bravo-Oviedo A (2015) Growth and yield of mixed versus pure stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) analysed along a productivity gradient through Europe. Eur J For Res 134:927–947
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Version 4.2.2. https://www.R-project.org/. Accessed 31 Oct 2022
Roques A, Auger-Rozenberg M-A, Boivin S (2006) A lack of native congeners may limit colonization of introduced conifers by indigenous insects in Europe. Can J for Res 36:299–313
Rubino FM, Alessio M, Pitton M, Di Fabio D, Caruso E, Banfi S, Tosi G, Wauters LA, Adriano M (2012) Food choice of Eurasian red squirrels and concentrations of anti-predatory secondary compounds. Mamm Biol 77:332–338
Salamolard M, Butet A, Leroux A, Bretagnolle V (2000) Responses of an avian predator to variations in prey density at a temperate latitude. Ecology 81:2428–2441
Sandom C, Dalby L, Fløjgaard C, Kissling WD, Lenoir J, Sandel B, Trøjelsgaard K, Ejrnæs R, Svenning J-C (2013) Mammal predator and prey species richness are strongly linked at macroscales. Ecology 94:1112–1122
Schickmann S, Urban A, Kräutler K, Nopp-Mayr U, Hackländer K (2012) The interrelationship of mycophagous small mammals and ectomycorrhizal fungi in primeval, disturbed and managed Central European mountainous forests. Oecologia 170:395–409
Schnabel F, Liu X, Kunz M, Barry KE, Bongers FJ, Bruelheide H, Fichtner A, Härdtle W, Li S, Pfaff C-T, Schmid B, Schwarz JA, Tang Z, Yang B, Bauhus J, von Oheimb G, Ma K, Wirth C (2021) Species richness stabilizes productivity via asynchrony and drought-tolerance diversity in a large-scale tree biodiversity experiment. Sci Adv 7:eabk1643
Schuldt A, Huke P, Glatthorn J, Hagge J, Wildermuth B, Matevski D (2022) Tree mixtures mediate negative effects of introduced tree species on bird taxonomic and functional diversity. J Appl Ecol 59:3049–3060
Schwarz CJ, Arnason AN (1996) A general methodology for the analysis of capture-recapture experiments in open populations. Biometrics 52:860
Selva N, Hobson KA, Cortés-Avizanda A, Zalewski A, Donázar JA (2012) Mast pulses shape trophic interactions between fluctuating rodent populations in a primeval forest. PLoS ONE 7:e51267
Spiecker H (2003) Silvicultural management in maintaining biodiversity and resistance of forests in Europe—temperate zone. J Environ Manag 67:55–65
Suchomel J, Purchart L, Čepelka L (2012) Structure and diversity of small-mammal communities of lowland forests in the rural central European landscape. Eur J For Res 131:1933–1941
Sundell J, Huitu O, Henttonen H, Kaikusalo A, Korpimäki E, Pietiäinen H, Saurola P, Hanski I (2004) Large-scale spatial dynamics of vole populations in Finland revealed by the breeding success of vole-eating avian predators. J Anim Ecol 73:167–178
Sundell J, O’Hara RB, Helle P, Hellstedt P, Henttonen H, Pietiäinen H (2013) Numerical response of small mustelids to vole abundance: delayed or not? Oikos 122:1112–1120
Thomas FM, Rzepecki A, Werner W (2022) Non-native Douglas fir (Pseudotsuga menziesii) in Central Europe: Ecology, performance and nature conservation. For Ecol Manag 506:119956
Thünen-Institut (2012) Third National Forest Inventory - results database. https://bwi.info/. Accessed 23 Sept 2021
Tourani M (2022) A review of spatial capture–recapture: ecological insights, limitations, and prospects. Ecol Evol 12:e8468
Vitali V, Büntgen U, Bauhus J (2017) Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob Change Biol 23:5108–5119
Wagner S, Collet C, Madsen P, Nakashizuka T, Nyland RD, Sagheb-Talebi K (2010) Beech regeneration research: From ecological to silvicultural aspects. For Ecol Manag 259:2172–2182
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer, New York
Zárybnická M, Riegert J, Bejček V, Sedláček F, Šťastný K, Šindelář J, Heroldová M, Vilímová J, Zima J (2017) Long-term changes of small mammal communities in heterogenous landscapes of Central Europe. Eur J Wildl Res 63:89
Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 1:3–14
Zwolak R, Bogdziewicz M, Rychlik L (2016a) Beech masting modifies the response of rodents to forest management. For Ecol Manag 359:268–276
Zwolak R, Bogdziewicz M, Wróbel A, Crone EE (2016b) Advantages of masting in European beech: timing of granivore satiation and benefits of seed caching support the predator dispersal hypothesis. Oecologia 180:749–758
Funding
Open Access funding enabled and organized by Projekt DEAL. Funding was provided by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) grant no. 316045089/GRK2300.
Author information
Authors and Affiliations
Contributions
SMA and NB conceived of and designed the study. Material preparation, data collection, and analysis were performed by SMA. The first draft of the manuscript was written by SMA, and NB commented on previous versions of the manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Ethical approval
Small mammal surveys were conducted in compliance with the German Animal Welfare Act under the “Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit” permit number 33.9-42502-04-18/2790. All procedures involving animals were approved by the internal animal welfare committee of the University of Göttingen.
Additional information
Handling editor: Stephanie Schai-Braun.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Appleby, S.M., Balkenhol, N. Douglas fir and Norway spruce have similar effects on small mammal density, but not survival, in Central European managed forests. Mamm Biol 104, 25–39 (2024). https://doi.org/10.1007/s42991-023-00384-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-023-00384-1