
Abstract
The responses of wildlife to environmental factors are of conservation importance. However, the absence of relevant information due to inadequate studies, and lack of understanding of the influences of environmental factors on wildlife, particularly in the Zambezian bioregion, remain a conservation concern. For instance, there is a shortage of knowledge on the relationship between fixed effects of environmental factors and behaviour as well as nest group sizes of Smith’s bush squirrels, Paraxerus cepapi Smith, 1836. Our study examined the relationship between fixed effects of several environmental factors (i.e., with focus on ecological factors) and the behaviour as well as nest group sizes of the bush squirrels in and around Zambia’s Chembe Bird Sanctuary (CBS). Flight initiation distances (FIDs) as an index of bush squirrel behaviour, and nest group sizes were simultaneously surveyed to understand the drivers of biological and population responses, respectively. The results revealed that higher tree height (m) and larger canopy coverage (%) could increase FIDs for bush squirrels. The nest group sizes of bush squirrels could also increase with canopy coverage (%) and presence of termite mounds. By focusing on the two different squirrels' responses, the results of this study highlight the most important environmental factors to consider in minimizing the impacts of human activities on bush squirrels, especially in conservation planning and management by taking into consideration the bush squirrels’ natural history, habitat protection and safe distance between humans and bush squirrels.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Land-use modifications have often resulted in habitat fragmentation (Lambin and Meyfroidt 2011) and biodiversity loss, especially in small and isolated fragments (Haddad et al. 2015). The habitat fragments limit the availability of nesting sites for tree nesting mammals, such as bats and flying lemurs, leading to a risk of local extinction (Crooks et al. 2017). The nests serve a protective role against disturbances and adversaries, such as climatic conditions and predation (Kolbe and Janzen 2002). In addition, the altered fire regimes by local communities during wildlife hunting, preparation of crop fields, and improper gathering of non-timber forest products, such as fruits, cause late fires, occurring way after the rains, in the hot dry season. Such fires are destructive to nests for birds, reptiles, and mammals, such as African savanna-dwelling rodents (Burgess et al. 2004; Eriksen 2007). Excessive fires may damage caches made by rodents, which would otherwise be useful during the dry season as secure food reserves at times of scarcity. The rodents also face other survival threats, such as road kills (Collinson et al. 2015) and bushmeat harvesting (Hoffman and Cawthorn 2012). Furthermore, increased human population growth and unsustainable land use, such as unmanaged agriculture, human settlements, plantations, timber harvesting and charcoal production, may threaten the survival of rodents by destroying their habitats through deforestation and fragmentation (Kataoka and Tamura 2005; Walpole and Bowman 2011). The loss of trees resulting from deforestation and habitat fragmentation may extirpate tree-dwelling rodents, dependent on trees for nesting, refuge, resting and foraging.
The Smith's bush squirrels (Paraxerus cepapi Smith, 1836; hereafter referred to as bush squirrels), also known as yellow-footed bush squirrels, are diurnal, arboreal, congregatory, socially cohesive, communally nesting, and territorial medium-sized mammalian rodents, with an estimated average body mass of 223 g (n = 69; Viljoen 1986). They live in varying family sizes of up to 12 individuals on 0.3–1.26 ha (Thorington et al. 2012). In the wild, bush squirrels can breed throughout the year (Viljoen 1975) but peak in March (Viljoen 1986). The group sizes can be as small as two prior to breeding and can increase to as many as 12 post-parturition (Viljoen 1977a). The bush squirrels use spacing calls to maintain their home ranges and territories, which contain caches of food (Viljoen 1983a). They inhabit an array of habitats, such as miombo and mopane woodland, especially with termitaria (Linzey and Kesner 1997; Fleming and Loveridge 2003; Anderson and Lurz 2019). They construct dreys or opportunistically use tree cavity nests. Dreys are made commonly from natural materials, such as leaves and sticks, while suitable tree cavities found on appropriate tree height away from dangers, such as predation and fires, are occupied. The bush squirrels depend on the availability of host trees for nesting as tree-dwelling rodents (Viljoen 1986). According to Viljoen (1981, 1986), the bush squirrel intraspecific competition and food availability could be the key limiting factors to a demographic increase in the wild. Bush squirrels are omnivorous, feeding on an array of dietary items, such as insects, seeds, leaves, fruits and flowers (Viljoen 1977b, 1983b). Furthermore, bush squirrels contribute to ecosystem functioning and biodiversity through seed and fungal dispersal, seed predation, and as prey for meso-predators, such as birds, monkeys, and canids (Mendes et al. 2019). Despite their ecological role and wide distribution in southern Africa, little is known of their ecology (Anderson and Lurz 2019). Although the International Union for Conservation of Nature (IUCN) Red List considers bush squirrels as of ‘Least Concern’ conservation status (Cassola 2016), it may require constant monitoring, particularly in rapidly changing environments. Bush squirrels are losing suitable habitats to ever-increasing agricultural and plantation activities by tree cutting. Besides causing wildfires during crop field preparations, farmers around Chembe Bird Sanctuary (CBS) use considerable amounts of chemicals in growing a variety of crops, such as maize (Zea mays), cotton (Gossypium hirsutum) and tobacco (Nicotiana tabacum), which may have negative impacts on ecosystem functioning.
Available historical population information of a particular species can guide human perceptions and resultant actions (Soga and Gaston 2018). From this general perspective, this study particularly motivates people’s conservation concerns for this species given limited available historical population information, making evidence-based conservation planning and management decisions practically difficult. Therefore, by highlighting relationships between FID or nest group size and environmental factors based on the study results, the human impacts on this species may be minimised in CBS and beyond. There is a need for the provision of biological and population information to stakeholders, who would then use human perceptions of change to inform conservation policy-making or management (Papworth et al. 2009). Our study provides a baseline to motivate future monitoring of bush squirrels in CBS. Given that bush squirrels’ coherent social organisation shapes their behaviour and nest group sizes (Viljoen 1977a, 1986), understanding these biological and population responses can be of great value to bush squirrel conservation.
There are several environmental factors that might impact squirrels’ behaviour and nest group sizes. For example, squirrels’ behaviour and nest group sizes may be influenced by wariness levels towards humans (Parker and Nilon 2008; Rodríguez-Prieto et al. 2009; Uchida et al. 2016). Land-use types, particularly with high human activities, may influence the antipredator behaviour by squirrels (Cooper et al. 2008; Engelhardt and Weladji 2011). Squirrels may select the host tree species based on food abundance (Krauze-Gryz et al. 2016; Reher et al. 2016) and nest site attributes, such as host tree height and nest status, such as active or abandoned nests (Ocaña-Mayorga et al. 2018).
Presence of termite mounds, supporting host trees for bush squirrels, may also encourage their higher abundance (Fleming and Loveridge 2003). Other potential environmental factors that might influence the behaviour of bush squirrels may include seasonal variations as is the case with Eurasian red squirrels (Uchida et al. 2016). The area with high canopy cover is likely to provide more food for squirrels, which may result in bigger nest group size (Trapp et al. 2017). Though environmental factors may singularly or collectively act to influence the behaviour and nest group sizes of bush squirrels, such information is often lacking to guide adaptive management. In this study, we explored the relationship between fixed effects of environmental factors and behaviour as well as nest group sizes of bush squirrels from human–squirrel coexistence perspective.
There is still a dearth of knowledge on the extent to which environmental factors influence behaviour and nest group sizes of medium-sized mammals (Rovero et al. 2017), particularly bush squirrels, although much useful information on the species natural history, specifically the interactions of bush squirrels with their environment in southern Africa has been documented (e.g., Viljoen 1977a, 1986). The aim of this study is to understand how extrinsic environmental factors influence both the tolerance to human activities shown by behavior, and population responses expressed by nest group sizes of bush squirrels. In this study, the research question is: (i) which among the identified key environmental variables influences the behaviour and nest group sizes of bush squirrel in the savanna environment? The behavioural responses by bush squirrels are determined by the use of FIDs. FID is generally defined as "the distance at which an animal moves away from an approaching threat" (Blumstein 2003). Thus, FID is a measure of antipredator behaviour that reflects the tolerance towards approaching threats, such as humans (Møller et al. 2013; Cooper and Blumstein 2015; Young et al. 2020), and employed in this study to understand the impact of human activity on bush squirel’s behaviour. There are several applications of FID by wildlife managers. For instance, FID is often used by wildlife managers: (1) to evaluate the wariness (or stress/risk perceived) of a particular species or population to human activities (Parker and Nilon 2008; Rodríguez-Prieto et al. 2009); and (2) to quantify human disturbance or determine "buffer zones" beyond which people are not allowed to go to minimise the disturbance on wild populations (Blumstein 2003; Weston et al. 2012; Guay et al. 2016). In such conservation applications, understanding the environmental influences on species, especially in ecosystems that have suffered disturbance and/or degradation, is important for restoration purposes (Jones et al. 2018). By examining FID as the measurement of tolerance to humans contributes to the actual conservation actions, such as buffer zonations and wariness assessments, which may promote protection of the species and its habitats for breeding and foraging (Mayer et al. 2019).
In this study, we used FID (tolerance to the human activity of individual behaviour) and nest group size (proximity of population-level response) simultaneously as dependent variables because the two measures may benefit to detect the environment where there is decrease in the FID (increase in the tolerance to human disturbance) and increase in nest group size (increase in the abundance). Our study involves three habitat types: miombo woodland, human settlement and exotic pine plantation. A human settlement refers to an assemblage of houses and farms. Squirrels may tolerate human presence (Uchida et al. 2016), decreasing their stress response and perception of risk, resulting in shorter FIDs at the proximity of human settlements than in more natural or wild habitats. Since predator risk, predator presence and human activity are directly related to FID, our study focuses on bush squirrel’s response to humans. We predict FID would be shorter in habitat which has high human activities because prior studies (e.g., Parker and Nilon 2008; Rodríguez-Prieto et al. 2009; Uchida et al. 2016) have shown that individuals in highly disturbed environments represent higher tolerance to humans than conspecifics in less disturbed environments. The larger the FID, the higher the wariness and perceived human disturbance as risk (Uchida et al. 2016). Furthermore, we predicted that there would be larger nest group sizes of bush squirrels in woodland (a dry, tropical deciduous forest) than human settlements and exotic pine plantations by seasons because while the bush squirrels are exposed to various threats, such as predation and wildfires in the study area, there are limited physical interactions with humans. The wildlife is better protected from seasonal human activities in woodland than elsewhere within the study area. However, changes in seasonal conditions, such as food availability in habitats, might result in changing rodent populations, as sporadic movements by rodents in between habitats would affect abundances (Mortelliti and Boitani 2006), amplifying the fixed effects of seasonal change from dry to the wet season. Therefore, it can be expected that there would be more bush squirrels in human settlements in the wet season than in the dry season. Further, it is expected that with such trans-seasonal movements, the FIDs in respective habitats would be dynamic.
Materials and methods
Study area
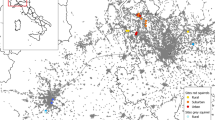
This study was conducted in Chembe Bird Sanctuary (CBS; 539 ha; located 27.9955º E, 12.8302º S) and its adjacent areas in Kalulushi District, Zambia (Fig. 1). The area is located in the Zambezian bioregion, characterised by a dearth of information on biodiversity (Linder et al. 2012). Predominant vegetation mosaics in the study area comprise miombo woodland and grasslands. The miombo woodland is the largest and dominant savanna in southern Africa and forms Zambia’s predominant dry tropical deciduous forests (Ryan and Williams 2011). The miombo woodland is characterised by the presence of genera Brachystegia, Julbernardia and Isoberlinia, utilised as wildlife habitat (Dewees et al. 2010), including bush squirrels.

Chembe Bird Sanctuary landscape study area in Kalulushi District, Copperbelt Province, Zambia
The area experiences temperatures in excess of 30 °C in the hot season, September–October (Conservation Farming Unit 2009). Bush squirrels are known to adjust to temperatures as high as 34 °C and have a low oxygen consumption of 0.65 ml O2/g h, indicative of low energy metabolism rate and activity (Viljoen 1985). Thus, the bush squirrels in the CBS landscape may have adjusted to such elevated temperatures. The bush squirrels construct leaf-and-stick dreys in selected trees and alternatively use existing tree cavities as dens and sleeping sites. Smith's bush squirrel (P. cepapi) is the only species of bush squirrel in CBS, and it is protected by the Zambian law (Chomba et al. 2011; GRZ 2015).
CBS receives a number of day visitors for eco-tourism throughout the year, and bush squirrels are among the fauna attracting such visitors to the area. In the adjoining areas of CBS, there are also exotic pinus spp. plantations, and remnant miombo woodlots in the human settlements utilised by bush squirrels. An estimated human agrarian population of 66,575 resides in these adjacent areas (CSO 2012). The subsistence farmers cultivate cereals, such as maize (Zea mays), millet (Eleusine coracana), cotton (Gossypium hirsutum) and tobacco (Nicotiana tabacum) and nut crops such as groundnuts (Arachis hypogaea), soya beans (Glycine max), and Bambara groundnuts or under-ground beans (Vigna subterranea) in addition to a variety of other crops. To exclude anthropogenic threats, such as wildfires, human encroachment and intrusion, CBS has been wire-fenced and vegetation around the fenced area cleared and fortified by Zambia’s Department of National Parks and Wildlife (DNPW). The predation risk for the bush squirrels in the study area arises principally from humans, vervet monkeys (Chlorocebus pygerythrus), slender mongoose (Galerella sanguinea), banded mongoose (Mungos mungo), snakes, such as puff adders (Bitis arietans arietans) and birds of prey, such as steppe buzzards (Buteo vulpinus). While there are no human settlements in the sanctuary and plantations, there is often the human presence in these areas. Human settlements, constituting clusters of houses and farms, are situated outside the sanctuary. Little is known about the abundance of predators, as well as specific human densities.
Data collection protocols
To examine the relationship between fixed effects of environmental factors and behaviour as well as nest group sizes of bush squirrels in the African savanna, data were collected in wet (February–March) and dry season (September–October) in 2018 and 2019 on three land-use types. The land-use types under this study were: (1) miombo woodland in CBS; (2) human settlements, where agrarian activities are practiced on the peripherals of CBS, and (3) pine plantations, comprising Pinus kesiya located adjacent to the sanctuary, where timber production and harvesting take place.
We undertook a multi-stage selection of sampling units. First, using a grid map covering the study area (Sutherland 2006), a total of 30 sampling plots of 200 m × 200 m each in dimension (i.e., ten plots per land-use type per season) were randomly selected. Woodland plots were only located within the sanctuary boundary, and the isolated small woodlots around the sanctuary were associated with anthills within the farmlands and human settlements. The human settlement and plantation plots were all outside the sanctuary as there were no human settlements and plantations within the sanctuary boundary. The plot sizes were small enough to allow easy detection of bush squirrels but big enough for its home range of up to 1.26 ha (own pers. obs.; Thorington et al. 2012). The plots were located physically by the use of Geographic Positioning System (GPS) central coordinates. The coordinates indicated the geographical locations of the centre points for each sampling plot. Secondly, these coordinates served as random starting points of the north-oriented 500 m line transects.
The distance sampling techniques using ground line-transect surveys were adopted from Uchida et al. (2016) and applied in 2-year consecutive periods of wet and dry seasons. The 30 line transects across the study sites were surveyed only once per season (total line-transect runs, n = 120) to avoid the bush squirrels getting accustomed to observers. Along the line transects, we used Tasco binoculars with a magnification of 20 × to locate the squirrels. As starting distance can affect FID (Cooper, 2005; Chamaillé-Jammes and Blumstein 2012; Dumont et al. 2012; Bonnot et al. 2017), to account for the variations in starting distances (i.e., the total distance between the observer at the start of approach and the animal when sighted), we measured the distance between the starting point (i.e., a point where the approach towards the animal started upon sighting it) and the location of the observer when the focal individual started to flee (i.e., the distance equivalent to the difference between the total distance and FID), also referred to as SD-FID. We targeted only the seemingly isolated foraging bush squirrels, as the bush squirrels displayed social organisation (Viljoen 1986), and would potentially affect others’ vigilance and consequently bias the FID outcomes. Furthermore, to avoid double-counting of the same bush squirrels, observers took note of the direction of flight, numbers and age classes (adults/juveniles) of the fleeing bush squirrels. The line transects were conducted from 09:00 to 12:00 h in the morning and 14:00 to 17:00 h in the afternoon, when visibility was optimal, coinciding with their diurnal peaks of activity (own pers. obs.). During the survey period, environmental conditions were favourable, such as being neither a rainy day in wet seasons nor an excessively hot day (> 30 °C) in dry seasons. Environmental factors potentially affecting FID, such as seasons (wet and dry seasons), canopy coverage (%), tree height (m), distance (m) from the point where the target bush squirrel was initially sighted (i.e., focal point) to the tree climbed by the approached bush squirrel upon fleeing (Blumstein 2006; Engelhardt and Weladji 2011) and land-use types were recorded. Visual observation method developed by Freitas et al. (2002) for estimating canopy coverage (%) was applied. Tree height was estimated using Suunto hypsometer clinometers (Leverett 2010) from the average of three randomly selected focal positions within 2 m radius from the focal point. Distances to bush squirrels, nests and points of flight by bush squirrels from humans within the sampling plots were measured using invar measuring tape. Further, as shorter FID is often considered as a proxy of animal tolerance to humans (Rodríguez-Prieto et al. 2010; Uchida et al. 2016), we searched for evidence of bush squirrels fed by humans, which would attract bush squirrels to humans. Given that in the context of our study, there were no specific sites/habitats where such a practice was typically taking place, we considered that any habitat with people might potentially have squirrels fed by humans.
Third, the nest group size was surveyed at sunsets and sunrises in accordance with Viljoen (1977a), in the same randomised sampling plots (i.e., the 30 sampling plots across the study sites; see Fig. 1). The bush squirrels came out of their nests to feed at sunrise and made their final retreat for the day to nests at sunset to rest and sleep. Thus, observers arrived at the nests at least 30 min before sunrise in readiness to count the bush squirrels emerging from the nest as light intensified, and at least 2 h before sunset as the bush squirrels retreated to their nests before sunset, and counted the respective nest group members to establish nest group sizes. The bush squirrels appear sensitive to light intensity (Viljoen 1983c), becoming more active during the day and less active before sunrise and after sunset. Further, observers concealed themselves from bush squirrels behind vegetation cover to avoid being seen because bush squirrels tend to evade potential predators, including humans (McCleery 2009; Greene et al. 2016), which would otherwise jeopardise the observations of the bush squirrels. With the use of 200 m × 200 m grids, counts of bush squirrel were adopted for estimating densities in the three land-use types, but not necessarily to indicate habitat quality (Van Horne, 1983), considering that testing for correlation between densities and habitat qualities were outside this studies’ scope. To determine the density, a number of squirrels per unit area (i.e., no. of squirrels/ha), all the individuals counted from the nest groups were summed up and divided by 4, as the size of the grid (i.e., 200 m × 200 m) was equivalent to 4 ha. The population sizes and densities per land-use types were determined from nest group sizes, as the bush squirrel’s natural history can contribute to an understanding of how bush squirrel’s interaction with its environment influences its behaviour and population (Gurnell 1987). In our study, nest group size referred to the number of bush squirrels hosted by a particular nest, and the break-away kin members with their own nests were included in the analysis as separate individuals. Thus, a nested group was not based on kinship, constituting a family. Distance sampling could not be adopted for bush squirrel density estimates because of the high possibility of bush squirrels hiding from observers and those in nests being missed during line-transect surveys (Viljoen 1986).
Environmental factors potentially affecting nest group sizes, inter alia; seasons (wet and dry seasons), canopy coverage (%), presence or absence of termite mounds, nest types (i.e. dreys and cavities), host tree height (m), nest status (i.e. active or abandoned), host tree species, tree diameter at breast height (DBH) in centimetres estimated at 1.3 m of host trees, and land-use types were recorded. At the nest site, we limited observation of natural causes of nest abandonment, such as nest invasions to the presence or absence of tree climbing ants. We surveyed abandoned nests because they could be potentially indicative of disturbances that would influence the squirrel’s occurrence, and consequently, affecting their nest group sizes. Fire scars were not included in the study because of the potential confounding effects in the area, which was widely burnt (own pers. obs.). Micro-variations in tree diameters over the study period were not considered on the assumption that there were insignificant between and within season variations caused by tree contractions and expansions at the short peak seasons (Krauss et al. 2007; O’Brien et al. 2008).
Host tree identification within the sample plots was made using a tree identification guide (Newman and Read 2008) and local expertise. We placed metal plates on surveyed trees as markers for ease identification in subsequent survey visits, as each nest was visited four times during the study period. With the help of the local people's expert knowledge and physical inspection for the presence of signs, such as loose fur, fresh nesting materials, and scratches around the bark of the host trees (Stuart and Stuart 2013), we established the nest status.
Data analysis
We modelled FIDs and nest group sizes of bush squirrels as functions of environmental covariates. All the numeric data tested for normality had significantly normal distribution, following their natural logarithmic transformation (Fig. S1). To detect the relationship between fixed effects of environment factor (i.e., explanatory variables) and FIDs as dependent variable, we used generalised linear mixed models (GLMMs), fitted by maximum likelihood for Laplace approximation (Zuur et al. 2009), with Gaussian as the model family. The three land-use types (i.e., miombo woodland, human settlements and pine plantations), seasons (wet and dry seasons), canopy coverage (%), tree heights (m), DBH (cm), and distance (m) from focal point to trees climbed by bush squirrels were considered as fixed effects, the distance (in meters) equivalent to the difference between the total distance (i.e., between observer and animal at the beginning of the approach) and FID as an independent variable, and year (2018 and 2019) as a random effect. Land-use types and seasons were applied as categorical variables with 3 and 2 levels, respectively. The dependent variable in this model was the FIDs, whereas the environmental factors were the independent variables. We tested FIDs a priori for normality (Borcard et al. 2011), using Shapiro–Wilk test (Shapiro and Wilk 1965), and then natural logarithmically transformed as the numeric data exhibited the skewed rather than normal distributions (Logan 2010; Fig. S1). In Fig. 2, environmental correlates of both FIDs and nest group sizes is visualised using ggplot2 package in R-statistical software. To meet the aim of this study, we employed likelihood ratio test (LRT)-based model selection to ascertain the predictions for the relationship between FID or nest group size and fixed effects of environmental factors. The LRT would be essential for detection of environmental factors influencing behaviors and nest group sizes, and how environmental factors can be managed to minimize the human impact. To determine which pair of models was better than others in explaining the relationship between FID and fixed effects of environmental factors, we applied anova function and tested with LRT the significance of the pair of models. For fitting the model for FIDs measured in metres as a continuous variable, we applied glmer function from lme4 package (Bates et al. 2015) and applied Gaussian as the model family.

Environmental correlates of FIDs and nest group sizes for bush squirrels in and around Chembe Bird Sanctuary, Zambia
We repeated the same GLMM procedure to analyse the fixed effects of different environmental factors on bush squirrel nest group sizes (i.e., number of individuals per nest), with Poisson as the model family. The nest group sizes were considered as a function of nest characteristics and environmental variables in CBS and its surrounding areas. The land-use types (miombo woodland, human settlements and pine plantations), seasons (wet and dry seasons), tree heights (m), nest status (active and abandoned), canopy coverage (%), presence or absence of termite mounds, and host tree species were considered as fixed effects, and the year (2018 and 2019) as a random effect. For FIDs and nest group sizes, the variance inflation factors (VIF) were used to determine multicollinearity of potential explanatory covariates prior to model fitting (Zuur et al. 2009; Logan 2010). Using faraway package in R-statistical software, the multicollinearity was not detected (VIF < 3) for FIDs, except for land-use types and DBH where VIF values were greater than the acceptable limit of 5 (Table S1). In nest group sizes, all the VIF values were also below 2, except for land-use types and nest height (Table S2). The variables with higher VIF values than the standard VIF of 5 were discarded from the variable selection. In the final GLMM building, lme4 package in R-statistical software was used. As in FID, we applied anova function and tested with LRT the significance of the pair of models for the nest group size. Data analyses were performed in R-statistical software, version 3.6.1 (R Core Development Team 2019).
Results
When the multicollinearity test was applied to all potential variables for FIDs, VIF values for land-use types and DBH were higher than 5, and removing these two variables significantly improved the VIF values for the retained variables at VIF < 3 (Table S1). Similarly, removing land-use types and nest height improved the VIF values to less than 2 for the remainder of the variables (Table S2).
The model containing three valuables: canopy coverage, tree height and distance to the climbed tree would potentially best explain bush squirrel’s FIDs, whereas nest group sizes would be potentially best explained by model with two variables: canopy coverage and presence or absence of termite mounds (Tables S3 and S4). The results of the final fitted GLMMs are presented in Tables 1 and 2. Host tree height and canopy coverage seemed to positively and significantly influence FIDs for bush squirrels (Table 1), while distance to climbed trees did not. Canopy coverage and presence of termite mounds also seemed to positively and significantly influence bush squirrel’s nest group sizes (Table 2). In addition, the bush squirrel densities derived from the nest group sizes are given as supplementary materials in Table S5.
Discussion
There was varied significance in the relationship between dependent variables and each fixed effect. Among several environmental factors, the FIDs of bush squirrels were influenced significantly by the tree height and canopy coverage (Table 1). As trees play a critical role in the protection of bush squirrels from adversaries like predation and fires as well as provision of nesting sites, tall trees (27.13 m [median], or taller) are the most preffered host trees. Therefore, large trees need to be conserved for bush squirrels. The bush squirrel also depends on the canopy coverage for concealment from a potential predator (Viljoen 1986). However, the canopy coverage may reduce because of natural and anthropogenic causes, such as land clearance for human settlements and agriculture (Maitima et al. 2009). For instance, improper agricultural practices in the human settlements that emphasised mass tree clearance may have contributed to the reduced canopy coverage for animal refuge. In CBS areas, farmers only left fragments, such as termite mounds, uncultivated as varying land contours were typically difficult for them to negotiate (own pers. obs.). In the pine plantations adjacent to CBS, the canopy coverage was determined by tree spacing. Further, though the distance to the climbed trees may affect FIDs as the animal has to flee to safety away from perceived risk based on optimal escape theory (Engelhardt and Weladji 2011; Samia et al. 2016), it appears the bush squirrels were more affected by tree height and canopy coverage than other considered variables in and around CBS (Tables 1 and S3). The more the canopy cover and the higher the tree height, the greater the FIDs (Fig. 2). We attribute the shorter FIDs in human settlements to levels of tolerance by the local farming community, evidenced from their traditional belief system that killing the bush squirrels would bring about a bad omen (own pers. obs.). Besides, there was no evidence of the deliberate feeding of bush squirrels in the area. Therefore, this coexistence between humans and bush squirrels in the study area where no deliberate feeding occurred could also be as a result of bush squirrels using humans as "shields"—a concept where some species actually perceive less predation threat in proximity to humans because predators tend to avoid humans (Sarmento and Berger 2017). Elsewhere in Japan, Uchida et al. (2016) attributed shorter FIDs to animal tolerance of humans caused by deliberate animal feeding.
The larger canopy coverage and presence termite mounds also seem to increase nest group sizes for bush squirrels (Table 2). Consequently, tree cutting, resulting in loss of canopy coverage and nest site depletion, might reduce bush squirrel nest group sizes. Although rodent populations may rapidly adjust to anthropogenic disturbances (Puig-Gironès et al. 2018), tree-dwelling bush squirrels are known to respond differently to habitat fragmentation (Koprowski 2005). The bush squirrel population response and their behavioural plasticity may depend on the severity of the perturbation impacts. Nonetheless, reduction of canopy coverage by anthropogenic activities around protected areas poses a significant risk to biodiversity (Joppa et al. 2016), particularly for a small protected area, such as CBS. While protected areas are usually biodiversity hotspots and important to biodiversity conservation beyond species of interest (Vina and Liu 2017), their immediate surroundings are often hotspots of environmental conflicts that threaten human–wildlife coexistence (Vedeld et al. 2012). The wanton removal of tall trees with large canopy coverage because of agriculture and timber logging in areas adjoining CBS, which could otherwise have been helpful for nest placement by bush squirrels, may cause extirpation of bush squirrel populations. Presence of termite mounds also seem to encourage larger nest group sizes (Tables 2 and S4), probably because of termite mounds being resource-rich of food and nesting sites sought by bush squirrels (Fleming and Loveridge 2003). The relatively larger number (60.9%, n = 39) of termite mounds in woodland than in the settlements and exotic pine plantations combined might have significantly affected the nest group sizes (Table S5) by providing additional resources to bush squirrels.
Conservation implications
The understanding of the relationship between FID or group nest size and environmental factors may be useful for designing the habitat and protecting the vegetation to minimize human impact. Based on the FIDs, wildlife managers may establish and implement buffer zones in the habitat to minimize disturbances to bush squirrels (Weston et al. 2012; Guay et al. 2016). Based on our data, tourist routes could be placed at greater than 9.4 m (median) away from the areas usually utilised by bush squirrels in the wild (i.e., miombo woodland). Spatial segregation of at least 2.1 and 5.3 m (medians) could be allowed between human activities and bush squirrels in human settlements and exotic pine plantations, respectively. Tree cutting should be controlled to promote tree growth for higher canopy coverage and greater tree height. Thus, understanding the relative importance of habitat quality for animals and their biological responses to perceived risks is essential for their management (Haughland and Larsen 2004). Further, high habitat quality decreases the impact of human activities and maintains the population of bush squirrels as depicted in the woodland. An intrinsic increase in nest group size may also positively impact flight behaviour by bush squirrels as their collective vigilance increases with vocalisation (Viljoen 1977a), thereby increasing population responses of bush squirrels.
Further, we also provide indirect conservation implications of this study because of their relevance to the study area. The understanding of the selection of nest tree characteristics by bush squirrels can later inform restoration approaches, such as selective preservation of trees or planting of specific tree species. Though bush squirrels are still categorised as species of least conservation concern (Cassola 2016), the protection of bush squirrels and their habitats should be given elevated management consideration to ensure their persistence in the contemporary dynamic environment. We propose enriching indigenous ecological knowledge about squirrels by increasing awareness creation among local people of the associated traditional beliefs through engagement and traditional leadership involvement, who are the respected curators and custodians of traditions. Further, the wildlife agency could promote community targeted extension service to integrate bush squirrel ecological importance in the environment. Our findings could be applied towards integration of life histories and habitat conditions through the protection of farm trees preferred by wildlife, such as bush squirrels (Ye et al. 2014; Oborn et al. 2017). Thus, there will be a need to implement multiple tools, such as co-designed role-playing games for conservation planning, strong law enforcement, and effective communication among the stakeholders (Röttgers 2016; Perrotton et al. 2017), for both natural and disturbed areas (Winter et al. 2018), focussing on nest level. Both bush squirrels and human inhabitants will benefit from conservation initiatives, such as eco-tourism and limited development, particularly since Zambia's rural poverty levels are above 60% (Snyman 2012; UNDP 2016). Furthermore, there is also a need to collectively explore and implement effective long-term restoration programmes to ensure speedy rehabilitation of habitats (Benayas and Bullock 2012), particularly in human settlements and plantations. Restoration of plantations to suitable bush squirrel habitat can, in part, be achieved by the use of prescribed fires while avoiding herbicide application (Boone et al. 2017). We also propose adding to plantations tree species preferred by bush squirrels to improve their nest group sizes. The restoration actions, such as regulating the cutting of trees which are preferred by bush squirrels in farm woodlots for wood fuels, construction and agriculture (Du Plessis 1995; Oborn et al. 2017), planting and caring for indigenous trees favoured by bush squirrels could be implemented within human settlements to ensure continued bush squirrel coexistence. Furthermore, restoration programmes may take the form of committed collaborative biodiversity offsets as a mechanism to encourage stakeholder participation in biodiversity conservation (Maron et al. 2012; Suding et al. 2015). Continued awareness creation and social learning among subsistence farmers and other stakeholders, such as wildlife managers and policy-makers on bush squirrels and associated benefits to the public, are critical for bush squirrel conservation. Through such FIDs and nest group size-based knowledge, necessary conditions for bush squirrel conservation may be generated.
Conclusions
Our study reveals that FIDs for bush squirrels increase significantly with tree height and canopy coverage, while nest group sizes also increase significantly with canopy coverage and presence of termite mounds. Thus, canopy coverage supports both biological and population responses of bush squirrels for FIDs and nest group sizes, respectively. The approaches used in this study allow a better understanding of environmental factors that significantly influence FIDs, and which nest characteristics favour bush squirrels (i.e., the conditions under which the bigger nest group sizes are found), highly social medium-sized mammalian tree-dwelling rodents. We postulate that specific bush squirrel conservation interventions should consider these insights on environmental factors influencing bush squirrel behaviour and nest group sizes, which may be site-specific. In particular, knowledge on the relationship between fixed effects of environmental factors and bush squirrel behaviour as well as nest group sizes can inform bush squirrel conservation planning and management.
Availability of data and material
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Code availability
Not applicable.
References
Anderson NE, Lurz PWW (2019) The density of Smith’s bush squirrel, Paraxerus cepapi, in Luambe National Park, Zambia. Afr J Ecol 58(2):332–336
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1–48
Benayas JMR, Bullock JM (2012) Restoration of biodiversity and ecosystem services on agricultural land. Ecosystems 15:883–899
Blumstein DT (2003) Flight-initiation distance in birds is dependent on intruder starting distance. J Wildl Manage 67(4):852–857
Blumstein DT (2006) The multipredator hypothesis and the evolutionary persistence of antipredator behaviour. Ethology 112:209–217
Bonnot NC, Hewison AJM, Morellet N, Gaillard J-M, Debeffe L, Couriot O, Cargnelutti B, Chaval Y, Lourtet B, Kjellander P, Vanpé C (2017) Stick or twist: Roe deer adjust their flight behaviour to the perceived trade-off between risk and reward. Anim Behav 124:35–46
Boone WW IV, McCleery RA, Reichert BE (2017) Fox squirrel response to forest restoration treatments in longleaf pine. J Mamm 98(6):1594–1603
Borcard D, Gillet F, Legendre P (2011) Numerical ecology with R. Springer, New York
Burgess N, Hales JD, Underwood E, Dinerstein E, Olson D, Itoua I, Schipper J, Ricketts T, Newman K (2004) Terrestrial eco-regions of Africa and Madagascar : a conservation assessment. Island Press, Washington DC
Cassola F (2016) Paraxerus cepapi. In: The IUCN red list of threatened species 2016:e.T16205A115131842. Downloaded on 12 October 2017: IUCN
Chamaillé-Jammes S, Blumstein DT (2012) A case for quantile regression in behavioural ecology: getting more out of flight initiation distance data. Behav Ecol Sociobiol 66:985–992
Chomba C, Mwenya AN, Nyirenda VR (2011) Wildlife legislation and institutional reforms in Zambia for the period, 1912–2011. J Sustain Dev Afr 13(6):218–236
Collinson WJ, Parker DM, Bernard RTF, Reilly BK, Davies-Mostert HT (2015) An inventory of vertebrate roadkill in the Greater Mapungubwe Transfrontier Conservation Area, South Africa. Afr J Wildl Res 45(3):301–311
Conservation Farming Unit (2009) Conservation farming and conservation agriculture handbook for hoe farmers in agro-ecological regions I and IIa. Conservation Farming Unit, Lusaka
Cooper WE Jr (2005) When and how do predator starting distances affect flight initiation distances? Can J Zool 83:1045–1050
Cooper WE Jr, Blumstein DT (2015) Escaping from predators: an integrative view of escape decisions. Cambridge University Press, Cambridge
Cooper CA, Neff AJ, Poon DP, Smith GR (2008) Behavioural responses of Eastern gray squirrels in suburban habitats differing in human activity levels. Northeastern Nat 15(4):619–625
Crooks KR, Burdett CL, Theobald DM, King SRB, Di Marco M, Rondinini C, Boitani L (2017) Quantification of habitat fragmentation reveals extinction risk in terrestrial mammals. Proc Natl Acad Sci 114:7635–7640
CSO (2012) The 2010 Census of population and housing. Central Statistical Office, Lusaka
Dewees PA, Campbell BM, Katerere Y, Sitoe A, Cunningham AB, Angelsen A, Wunder S (2010) Managing the miombo woodlands of Southern Africa: policies, incentives, and options for the rural poor. J Nat Resour Policy Res 2:57–73
Du Plessis MA (1995) The effects of fuel wood removal on the diversity of some cavity-using birds and mammals in South Africa. Biol Conserv 74:77–82
Dumont F, Pasquaretta C, Réale D, Bogliani G, von Hardenberg A (2012) Flight initiation distance and starting distance: biological effect or mathematical artefact? Ethology 118(11):1051–1062
Engelhardt SC, Weladji RB (2011) Effects of levels of human exposure on flight initiation distance and distance to refuge in foraging eastern gray squirrels (Sciurus carolinensis). Can J Zool 89:823–830
Eriksen C (2007) Why do they burn the ‘bush’? Fire, rural livelihoods, and conservation in Zambia. Geogr J 173:242–256
Fleming PA, Loveridge JP (2003) Miombo woodland termite mounds: resource islands for small vertebrates? J Zool 259(2):161–168
Freitas SR, Cerqueira R, Vieira MV (2002) A device and standard variables to describe microhabitat structure of small mammals based on plant cover. Braz J Biol 62(4B):795–800
Greene DU, McCleery RA, Wagner LM (2016) A comparison of four survey methods for detecting fox squirrels in the Southeastern United States. J Fish Wildl Manag 7(1):99–106
GRZ (2015) The Zambia Wildlife Act, 2015. Government of Republic of Zambia, Lusaka
Guay P-J, van Dongen WFD, Robinson RW, Blumstein DT, Weston MA (2016) AvianBuffer: an interactive tool for characterizing and managing wildlife fear responses. Ambio 45(7):841–851
Gurnell J (1987) Natural history of squirrels. Christopher Helm, London, UK
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A, Holt RD, Lovejoy TE, Sexton JO, Austin MP, Collins CD et al (2015) Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci Adv 1:1–9
Haughland DL, Larsen KW (2004) Exploration correlates with settlement: red squirrel dispersal in contrasting habitats. J Anim Ecol 73(6):1024–1034
Hoffman LC, Cawthorn DM (2012) What is the role and contribution of meat from wildlife in providing high quality protein for consumption? Anim Front 2(4):40–53
Jones HP, Jones PC, Barbier EB, Blackburn RC, Benayas JMR, Holl KD, McCrackin M, Meli P, Montoya D, Mateos DM (2018) Restoration and repair of Earth’s damaged ecosystems. Proc R Soc B 285:20172577
Joppa L, Baillie JEM, Robinson JG (2016) Protected areas: are they safeguarding biodiversity? Wiley-Blackwell, West Sussex
Kataoka T, Tamura N (2005) Effects of habitat fragmentation on the presence of Japanese squirrels, Sciurus lis, in suburban forests. Mamm Stud 30(2):131–137
Kolbe JJ, Janzen FJ (2002) Impact of nest-site selection on nest success and nest temperature in natural and disturbed habitats. Ecology 83:269–281
Koprowski JL (2005) The response of tree squirrels to fragmentation: a review and synthesis. Anim Conserv 8:369–376
Krauss KW, Keeland BD, Allen JA, Ewel KC, Johnson DJ (2007) Effects of season, rainfall and hydrogeomorphic setting on mangrove tree growth in Micronesia. Biotropica 39:161–170
Krauze-Gryz D, Mazur K, Gryz J (2016) Density of red squirrels and their use of non-native tree species in the Rogow Arboretum. For Res Papers 77(1):42–49
Lambin EF, Meyfroidt P (2011) Global land use change, economic globalization, and the looming land scarcity. Proc Natl Acad Sci 108:3465–3472
Leverett RT (2010) Measuring tree height by tape and clinometers scenarios. Bull East Nativ Tree Soc 5:3–12
Linder HP, de Klerk HM, Born J, Burgess ND, Fjeldsa J, Rahbek C (2012) The partitioning of Africa: statistically defined biogeographical regions in sub-Saharan Africa. J Biogeogr 39:1189–1205
Linzey AV, Kesner MH (1997) Small mammals of a woodland-savannah ecosystem in Zimbabwe II Community Structure. J Zool 243(1):153–162
Logan M (2010) Biostatistical design and analysis using R: a practical guide. Wiley-Blackwell, West Sussex
Maitima JM, Mugatha SM, ReidRS GLN, Majule A, Lyaruu H, Pomery D, Mathai S, Mugisha S (2009) The linkages between land use change, land degradation and biodiversity across East Africa. Afr J Environ Sci Technol 3:310–325
Maron M, Hobbs RJ, Moilanen A, Matthews JW, Christie K, Gardner TA, Lindenmayer DB, McAlpine CA (2012) Faustian bargains? Restoration realities in the context of biodiversity offset policies. Biol Conserv 155:141–148
Mayer M, Natusch D, Frank S (2019) Water body type and group size affect the flight initiation distance of European waterbirds. PLoS One 14(7):e0219845
McCleery RA (2009) Changes in fox squirrel anti-predator behaviour across the urban-rural gradient. Landsc Ecol 24:483–493
Mendes CP, Koprowski JL, Galetti M (2019) Neosquirrel: a data set of ecological knowledge on neotropical squirrels. Mamm Rev 49(3):210–225
Møller AP, Grim T, Ibáñez-Álamo JD, Markó G, Tryjanowski P (2013) Change in flight initiation distance between urban and rural habitats following a cold winter. Behav Ecol 24(5):1211–1217
Mortelliti A, Boitani L (2006) Patterns of rodent species diversity and abundance in a Kenyan relict tropical rainforest. Biodivers Conserv 15:1425–1440
Newman K, Read C (2008) The wildlife of Southern Africa: the larger illustrated guide to the animals and plants of the region. Random House Struik, Cape Town, pp 114–163
O’Brien JJ, Oberbauer SF, Clark DB, Clark DA (2008) Phenology and stem diameter increment seasonality in a Costa Rican wet tropical forest. Biotropica 40:151–159
Oborn I, Vanlauwe B, Phillips M, Thomas R, Brooijmans W, Atta-Krah K (2017) Sustainable intensification in smallholder agriculture: an integrated systems research approach. Routledge, London
Ocaña-Mayorga S, Lobos SE, Crespo-Pérez V, Villacís AG, Miguel-Pinto C (2018) Influence of ecological factors on the presence of a triatomine species associated with the arboreal habitat of a host of Trypanosoma cruzi. Parasites Vectors 11:567
Papworth SK, Rist J, Coad L, Milner-Gulland EJ (2009) Evidence for shifting baseline syndrome in conservation. Conserv Lett 2(2):93–100
Parker TS, Nilon CH (2008) Gray squirrel density, habitat suitability, and behaviour in urban parks. Urban Ecosyst 11(3):243–255
Perrotton A, de Garine-Wichatitsky M, Valls-Fox H, Le Page C (2017) My cattle and your park: codesigning a role-playing game with rural communities to promote multistakeholder dialogue at the edge of protected areas. Ecol Soc 22(1):35
Puig-Gironès R, Clavero M, Pons P (2018) Importance of internal refuges and the external unburnt area in the recovery of rodent populations after wildfire. Int J Wildland Fire 27:425–436
R Core Development Team (2019) R: A Language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Reher S, Dausmann KH, Warnecke L, Turner J (2016) Food availability affects habitat use of Eurasian red squirrels (Sciurus vulgaris) in a semi-urban environment. J Mamm 97:1543–1554
Rodríguez-Prieto I, Fernández-Juricic E, Martín J, Regis Y (2009) Antipredator behaviour in blackbirds: habituation complements risk allocation. Behav Ecol 20(2):371–377
Rodríguez-Prieto I, Fernández-Juricic E, Martín J (2010) Habituation to low-risk predators improves body condition in lizards. Behav Ecol Sociobiol 64(12):1937–1945
Röttgers D (2016) Conditional cooperation, context and why strong rules work—a Namibian common-pool resource experiment. Ecol Econ 129:21–31
Rovero F, Owen N, Jones T, Canteri E, Iemma A, Tattoni C (2017) Camera trapping surveys of forest mammal communities in the Eastern Arc Mountains reveal generalized habitat and human disturbance responses. Biodivers Conserv 26(5):1103–1119
Ryan CM, Williams M (2011) How does fire intensity and frequency affect miombo woodland tree populations and biomass. Ecol Appl 21:48–60
Samia DSM, Blumstein DT, Stankowich T, Copper WE Jr (2016) Fifty years of chasing lizards: new insights advance optimal escape theory. Biodivers Rev 91:349–366
Sarmento WM, Berger J (2017) Human visitation limits the utility of protected areas as ecological baselines. Biol Conserv 212:316–326
Shapiro S, Wilk MB (1965) An analysis of variance test for normality (complete samples). Biometrika 52:591–611
Snyman S (2012) The role of eco-tourism employment in poverty reduction and community perceptions of conservation and tourism in Southern Africa. J Sustain Tour 20:395–416
Soga M, Gaston KJ (2018) Shifting baseline syndrome: causes, consequences, and implications. Front Ecol Environ 16(4):222–230
Stuart C, Stuart M (2013) A field guide to the tracks and signs of Southern Central and East African wildlife. Struik Nature, Cape Town
Suding K, Higgs E, Palmer M, Callicott JB, Anderson CB, Baker M, Gutrich JJ, Hondula KL, LaFevor MC, Larson BMH, Randall A, Ruhl JB, Schwartz KZS (2015) Committing to ecological restoration. Science 348(6235):638–640
Sutherland WJ (2006) Ecological census techniques: a handbook, 2nd edn. Cambridge University Press, Cambridge
Thorington R, Koprowski J, Steele M, Whatton J (2012) Squirrels of the world. The Johns Hopkings University Press, Baltimore, Maryland
Trapp SE, Smith WP, Flaherty EA (2017) Diet and food availability of the Virginia northern flying squirrel (Glaucomys sabrinus fuscus): implications for dispersal in a fragmented forest. J Mammal 98(6):1688–1696
Uchida K, Suzuki K, Shimamoto T, Yanagawa H, Koizumi I (2016) Seasonal variation of flight initiation distance in Eurasian red squirrels in urban versus rural habitat. J Zool 298:225–231
UNDP (2016) Zambia Human Development Report: industrialisation and human development-poverty reduction through wealth and employment creation. United Nations Development Programme, Lusaka
Van Horne B (1983) Density as a misleading indicator of habitat quality. J Wildl Manage 47(4):893–901
Vedeld P, Jumane A, Wapalila G, Songorwa (2012) A protected areas, poverty and conflicts: a livelihood case study of Mikumi National Park, Tanzania. For Policy Econ 21:20–31
Viljoen S (1975) Aspects of the ecology, reproductive physiology and ethology of the bush squirrel, Paraxerus cepapi cepapi. M.Sc Thesis. University of Pretoria
Viljoen S (1977a) Behaviour of the bush squirrel, Paraxerus cepapi cepapi (A. Smith, 1836). Mammalia 41(2):119–166
Viljoen S (1977b) Feeding habits of the bush squirrel Paraxerus cepapi cepapi (Rodentia: Sciuridae). Afr Zool 12(2):459–467
Viljoen S (1981) Environment and reproduction in tree squirrels with special reference to the genus Paraxerus. In: Gilmore DP, Cooke B (eds) Environmental factors in mammal reproduction. Palgrave Macmillan, London, pp 178–186
Viljoen S (1983a) Communicatory behaviour of southern African tree squirrels, Paraxerus palliatus, ornatus, P. p. tongensis, P. c. cepapi and Funisciurus congicus. Mammalia 47:441–461
Viljoen S (1983b) Feeding habits and comparative feeding rates of three southern African arboreal squirrels. Afr Zool 18(4):378–387
Viljoen S (1983c) Activity parameters relative to habitat in four southern African tree squirrels. South Afr J Zool 18:134–138
Viljoen S (1985) Comparative thermoregulatory adaptations of southern African tree squirrels from four different habitats. South Afr J Zool 20(1):28–32
Viljoen S (1986) Use of space in southern African tree squirrels. Mamm 50(3):293–309
Vina A, Liu J (2017) Hidden roles of protected areas in the conservation of biodiversity and ecosystem services. Ecosphere 8:e01864
Walpole AA, Bowman J (2011) Patch occupancy by squirrels in fragmented deciduous forest: effects of behaviour. Acta Theriol 56:63–72
Weston MA, McLeod EM, Blumstein DT, Guay P-J (2012) A review of flight-initiation distances and their application to Australian birds. Emu-Austral Ornithol 112(4):269–286
Winter E, Gronau S, Grote U (2018) Sustaining rural livelihoods through an integrated landscape approach. Biodivers Ecol 6:288–294
Ye X, Skidmore AK, Wang T (2014) Joint effects of habitat heterogeneity and species’ life-history traits on population dynamics in spatially structured landscapes. PLoS One 9(9):e107742
Young JK, Coppock DL, Baggio JA, Rood KA, Yirga G (2020) Linking human perceptions and spotted hyena behavior in Urban areas of Ethiopia. Animals 10:2400
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer Verlag, New York
Acknowledgements
This research was commissioned by the Copperbelt University, and the Department of National Parks and Wildlife. We are also grateful to many people who took part in the fieldwork. The anonymous reviewers of the initial draft are greatly thanked for providing constructive contributions used to shape the manuscript.
Funding
This study was partly supported by the JSPS grant (21K01027).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Ethics approval
The study was approved by the Copperbelt University and the Department of National Parks and Wildlife.
Consent to participants
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Handling editor: Raquel Monclús.
Supplementary Information
Below is the link to the electronic supplementary material.
42991_2021_159_MOESM1_ESM.docx
Supplementary file1 Fig. S1 Normality test for quantitative data in this study (a-FIDs; b-Nest group sizes; c-SD-FIDs; d-Tree height; e-Nest height; f-Diameter at breast height). Table S1 Multi-colinearity tests for FIDs regressed on potential independent environmental covariates in and around CBS, Zambia, in 2018 and 2019. Table S2. Multi-colinearity tests for nest group counts regressed on potential independent environmental covariates in and around CBS, Zambia, in 2018 and 2019. Table S3 Model selection for determination of environmental factors influencing FIDs in and around CBS, Zambia, in 2018 and 2019. Table S4 Model selection for determination of environmental factors influencing nest group sizes in and around CBS, Zambia, in 2018 and 2019. Table S5 The bush squirrel densities based on nest group sizes by seasons and land-use types in CBS landscape, 2018 and 2019 (DOCX 216 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nyirenda, V.R., Sakala, S., Simwanda, M. et al. Effects of environmental factors on the behaviour and nest group sizes of Smith's bush squirrels, Paraxerus cepapi, in a Zambezian bioregion. Mamm Biol 101, 555–566 (2021). https://doi.org/10.1007/s42991-021-00159-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-021-00159-6