
Abstract
El Cierro Cave (Ribadesella, Asturias, Spain) possesses one of the most complete Upper Palaeolithic stratigraphic sequences in northern Spain. Magdalenian occupations, particularly the lower Magdalenian, are well represented in its full sequence. This article presents the zooarchaeological analysis of the levels Cierro G1, Cierro G and Cierro F, dated by 14C to between 20,000 and 17,000 cal BP. The remains correspond to vertebrates (mainly large mammals and to a lesser extent fish and birds) and invertebrates (almost exclusively marine molluscs). The taphonomic analysis of the bones and shells has determined the role that the different animal resources played in the diet of the Magdalenian hunter-gatherers at El Cierro. The results obtained in each level of the sequence are compared and included in their regional context. This study also considers the way in which the Magdalenian groups at El Cierro processed the meat and fat of the different animal species and establishes the operational chain in the strategies of procurement, preparation and consumption of the prey.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The animal resources that formed part of the diet of the human groups that lived in northern Spain between ca. 20,000 and 17,000 cal BP have been a common topic of study since the 1970s (e.g. Altuna, 1972 and 1976; Castaños, 1980; Eastham, 1984; Morales, 1984; Madariaga & Fernández, 1987). Although taphonomic analyses were not carried out in the region before the 1990s and were little developed until the change of millennium, several studies are now available about the strategies involved in hunting large mammals (e.g. Carvalho et al., 2021; Portero, 2022; Portero et al., 2019 and 2024), gathering molluscs (e.g. Álvarez-Fernández, 2011 and 2017), fishing (e.g. Pereda & Doadrio, 2016; Roselló & Morales, 2017) and fowling (e.g. Elorza, 2014; Llorente & Morales, 2016; Rufà et al., 2022) during the above-mentioned period in the north of Spain.
This research has succeeded in determining the importance of the different animal resources in the diet of the hunter-gatherers, although few of them offer a picture that encompasses all the zoological resources they consumed (Álvarez-Fernández et al., 2019, 2020 and 2022a; Martínez-Villa et al., 2022). Similarly, few works analyse the role of terrestrial and marine fauna in the diet of Magdalenian groups and most of them form part of the integral study of particular archaeological levels (e.g. Álvarez-Fernández et al., 2019, for Cova Rosa, and Arias et al., 2005, for La Garma A). Additionally, no studies have explored changes in subsistence strategies in the course of a Magdalenian sequence in a deposit and compared the intra-site results.
The present study provides new data about subsistence at El Cierro Cave (Ribadesella, Asturias, Spain) obtained from three lower Magdalenian levels (Cierro G1, Cierro G and Cierro F) through zooarchaeological and taphonomic analysis of vertebrates (large mammals, fish and birds) and invertebrates (marine molluscs). The results obtained in the three levels are compared with the information available from other contexts in the Cantabrian region of Spain that are dated to the lower Magdalenian.
El Cierro Cave (Ribadesella, Asturias, Spain)
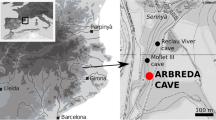
El Cierro is a limestone cave at the eastern end of the Asturian Massif in the Cantabrian Mountains (Fig. 1). It is located at 83 m above sea level in a pre-littoral depression. The cave has two entrances that are connected to each other behind a doline. The roof of the main room, where the site is located, collapsed, which led to the opening of a zenithal window that allows sunlight to enter. When the cave was discovered at the end of the 1950s, the main room was partially filled with archaeological deposits crowned by a shell midden (Álvarez-Fernández et al., 2016; Jordá Pardo et al., 2018a).

a) Map showing the location of the El Cierro Cave in Cantabrian Spain. b) Map of the Magdalenian sites mentioned in the text
El Cierro is located 3.1 km from the mouth of the river Sella and 2.1 km from the modern coastline. During the lower Magdalenian, the marine regression, about 75 m below the current one, would place El Cierro cave at 6 km from the coast (Jordá Pardo et al., 2018b). Paleoenvironmental and anthracological studies carried out at the site suggest the presence of an open landscape and a cold climate in the area of El Cierro during the lower Magdalenian, due to the presence of moor taxa, particularly Leguminosae, and rodents such as the water rat and the field vole (Uzquiano, 2019; García-Ibaibarriaga et al., 2019; Álvarez-Fernández et al., 2022a). However, the presence of several forested areas can also be deduced from the existence of micromammals such as Sorex araneus-coronatus in other nearby sites (Garcia-Ibaibarriaga et al., 2019). Regarding climate, isotopic studies (δ13C and δ18O) of the Magdalenian levels indicate that the average annual precipitation during this period was above 500 mm year−1, and the temperature was between 5 and 8 °C less than the current temperature (Lecuyer et al., 2021).
The lower Magdalenian faunal remains studied here come from the excavations performed by F. Jordá Cerdá and A. Gómez Fuentes from 1977 to 1979. This excavation was carried out in the main room, which has an extension of about 4 × 5 m where the Magdalenian habitat at El Cierro was located. Given the characteristics of the intervention area, the excavation was carried out in the northwest area of the room, in an area of 200 × 60 cm; they excavated to a depth of about 50 cm below the shell midden that forms the top of the sequence. This intervention area, although it is a marginal area of the site, was well protected from the erosive processes resulting from the entry of water through the zenithal opening of the room (Álvarez-Fernández et al., 2016; Jordá Pardo et al., 2018a).
The excavated sediment was screened with 5 to 2 cm mesh sizes (Álvarez-Fernández et al., 2016). The stratigraphic sequence obtained in this excavation was composed of 14 stratigraphic levels that were identified with letters from B to N (Fig. 2) (Álvarez-Fernández et al., 2018a; Jordá Pardo et al., 2018a).

Drawing of the stratigraphic section in El Cierro Cave by Francisco Jordá Cerda and Alejandro Gómez Fuentes, in which the studied levels are highlighted (apud Álvarez-Fernández et al., 2016 )
In 2014, an intervention was carried out on a core from the excavation of the 1970s in order to determine the characteristics of the occupations, for which, among other works, the lithostratigraphic sequence was revised and different samples were taken (pollen, sedimentology and biotic samples for radiocarbon dating) (Álvarez-Fernández et al., 2018a, 2022a). These samples led to the appearance of, among others, faunal remains that will also be studied in this work.
The three stratigraphic units belonging to the lower Magdalenian display the following lithostratigraphic characteristics, from the base to the top of the sequence (Álvarez-Fernández et al., 2018a; Jordá Pardo et al., 2018a):
-
Cierro G1: formed by rounded fine and very fine quartz sand, with black silt and clay. It is 30 cm thick to the west and 20 cm to the east. In the west, a large autochthonous limestone boulder 125 cm long and 60 cm thick deforms the unit
-
Cierro G: formed by two pockets of sediment up to 30 cm thick that are intercalated with Level F
-
Cierro F: level formed by light brown silt and clay with fine rounded quartz sand. It contains clasts of autochthonous limestone with angular and rounded edges in a chaotic arrangement. The level forms an irregular pocket between the two layers of Cierro G
Four AMS radiocarbon dates have been obtained at Oxford Radiocarbon Accelerator Unit for these three levels. One of these is a new date not published previously (Table 1). They indicate a period between 19,735 and 18,504 cal BP, which is similar to the age of other deposits, such as Levels CR2 and CR1 at Cova Rosa (Álvarez-Fernández et al., 2021) in the same Sella valley as El Cierro and in other caves in Asturias, like Level B4 in the cave of Coímbre B (Álvarez-Alonso & Jordá Pardo, 2017), Levels G and F in Arangas Cave in the Cares valley (Álvarez-Fernández et al., 2020) and Levels XII and XI in Chamber II in Las Caldas Cave in the Nalón valley (Corchón, 2017a). In Cantabria, the dates are closely correlated with, for example, Levels 19 and 17 in the outer vestibule in El Mirón Cave (Hopkins et al., 2021), Levels 3 to 5 in El Rascaño Cave (Barandiarán & González Echegaray, 1981), Levels 4 to 11 in El Juyo Cave (González Echegaray, 1987) and Level 2 at Altamira, 3c at El Linar, Level B in Las Aguas Cave and Level E at Cualventi (Rasines del Río, 2016). In the Basque Country, these lower Magdalenian dates resemble the chronology of the occupations in Santimamiñe Cave (Level Csn-Camr) (López Quintana & Guenga, 2011), in Ekain Cave (Level VII) (Altuna, 1984 and 2012) and in Erralla Cave (Level V) (Altuna, 1985a).
Method
All the faunal remains recovered in the 1970s excavations and in 2014 have been studied. However, it should be noted that the sediment extracted in the 1977–1979 fieldwork was not screened with small mesh sizes, unlike in the 2014 fieldwork in which 1 mm and 0.5 mm mesh were used, and therefore, the information about small vertebrates and invertebrates is partial (Álvarez-Fernández et al., 2022a).
The large mammals were identified anatomically and taxonomically using the reference collections in the Zooarchaeology Laboratories at the University of Salamanca and Aranzadi Science Society and with different anatomical atlases for large mammals (Barone, 1966; Schmid, 1972; Pales & Garcia, 1981; Varela & Rodríguez, 2004; Hillson, 2005; López-Plana et al., 2011), birds (Bacher, 1967; Kraft, 1972; Hoyo et al., 1992) and fish (Le Gall, 1984). Uniquely, for the taxonomic identification of three teeth from Cierro G, the peptide fingerprint was identified with the ZooMS method (Buckley et al., 2009) at the Autonomous University of Barcelona.
For the study of the skeletal profiles, the remains were classified into anatomical regions, differentiating between cranial elements (antler, skull, maxilla and mandible), axial elements (vertebra, rib, sternum, scapula, pelvis, sacrum and flat bone), anterior appendicular elements (humerus, radius, ulna, carpal and metacarpal) and posterior elements (femur, patella, tibia, fibula, tarsal and metatarsal). When the appendicular region could not be determined, the bone was included in the generic category of limb (metapodial, phalanx, sesamoid and long bone shaft or epiphysis) (Yravedra, 2006). In those cases when the faunal remain could not be assigned a taxonomic and anatomic classification, it was included in one of three mammal size categories: large mammals (> 300 kg), medium-size (100–300 kg) and small (5–100 kg), based on criteria of the thickness and robustness of the cortical bone (Bunn & Kroll, 1986; Yravedra, 2006; Portero, 2022).
The nomenclature of FAUNA EUROPAEA was followed for large mammals, birds and fishes (De Jong et al., 2014). Fish classification followed Fishbase (Froese & Pauly, 2024).
The terrestrial faunal assemblages were quantified in terms of the number of remains (NR) (Reitz & Wing, 2003), number of identified specimens (NISP) (Grayson, 1984; Lyman, 2019), minimum number of elements (MNE) (Bunn, 1983; Marean & Spencer, 1991) and minimum number of individuals (MNI) (Lyman, 1994).
To determine whether the skeletal representation of ungulates responds to the selective transport of certain anatomical elements (e.g. Grayson, 1989; Costamagno, 2002; Madrigal & Holt, 2002; Faith & Gordon, 2007; Saladié et al., 2011), we calculated Pearson’s correlation coefficient between the percentage of the skeletal representation of ungulates and the minimum animal units (MAU), the modified general utility index (MGUI) and the meat, marrow and grease indices for caribou and sheep, which we have applied to red deer and Iberian ibex in this work, as these are the most abundant taxa at El Cierro. For the red deer, we have also used the standardized food utility index (SFUI) proposed by Metcalfe & Jones (1988). From these correlations, we have determined the type of relationship between the variables, the strength of the relationship and whether it is statistically significant (p-value < 0.05).
The age of death of the mammals was established based on the patterns of tooth eruption and wear and epiphysis fusion for the different species (Noddle, 1974; Grigson, 1982; Levine, 1982; Mariezkurrena, 1983; Mariezkurrena & Altuna, 1983; Wegrzyn & Serwatka, 1984; Eisenmann et al., 1988; Pérez Ripoll, 1988; Brown & Chapman, 1991; Azorit et al., 2002 and 2003; Steele, 2002; Tomé & Vigne, 2003). For the birds, the studies of Bacher (1967) and Kraft (1972) have been followed.
Once the age of the individuals was established, we classified the mammals according to five age categories: neonate (deciduous dentition not yet fully erupted and without wear, or bones with early ossification), immature (animals with complete deciduous dentition and/or with permanent teeth germs and epiphyses of long bones not yet fused), juvenile (some deciduous teeth present, but also with permanent teeth with little wea, and the bones do not have fused epiphyses and/or maintain the fusion line of the epiphyses), adult (animals with complete permanent dentition and fully fused epiphyses) and senile (individuals whose permanent dentition shows a high degree of wear). In order to establish the mortality patterns of the prey hunted by the Magdalenian groups of El Cierro, we grouped the mortality profiles based on ternary graphs (Stiner, 1990), taking into account the four zonations established by Discamps & Costamagno (2015) and adapted to the life cycle of each species:
-
“JPO” (juvenile-prime-old) zone: includes sites with the following ratio: juvenile > adult > senile. Mainly refers to L-shaped mortality profiles (but also to a small U-shaped group)
-
“JOP” (juvenile-old-prime) zone: determined by the following class ratio: juveniles > senile > adults. Characteristic of the attritional profiles (“U” shape)
-
“P” (prime) zone: dominated by adult individuals
-
“O” (old) zone: includes age profiles showing a mortality pattern with a predominance of senile animals
The seasonality of the Magdalenian occupations was estimated based on the age profiles and reproduction biological cycles of modern wild species, considering the times of mating, gestation, birth, growth and life expectancy of those species (e.g. Costamagno, 1999; Discamps & Costamagno, 2015).
For the taphonomic analysis, the bone surfaces were observed with hand magnifying glasses of different magnifications (× 5, × 10 and × 15), combined with a Leica EZ4 HD microscope (6.5–32 ×). Cutmarks were analysed in terms of their morphology, location and orientation on the bone in order to determine their function in the butchery process (e.g. Binford, 1981; Shipman & Rose, 1983; Bunn & Kroll, 1986; Yravedra, 2006; Costamagno et al., 2019). To establish the butchery processes, the classification proposed by Binford (1981) was followed, and each cut mark on the bone was counted individually, and where possible the position was indicated on anatomical templates.
The frequency of the cutmarks was also analysed based on the NISP to determine which elements were most often processed for each species and size category. The anthropogenic breakage of the bones considered the angle, line and type of fracture as well as the shape of the percussion marks (Villa & Mahieu, 1991; Pickering & Egeland, 2006; Vettese et al., 2020). Wherever possible, the exact position of the impact on the bone was noted to detect any breakage standardised patterns (Masset et al., 2016). The degree of burning of the bones was calculated from their colour (Stiner et al., 1995), which is a proxy for the temperature they were exposed to (Nicholson, 1993). The burnt remains have been grouped into six categories according to the degree of burning as proposed by Stiner et al. (1995):
-
Grade 1: partially burned with brown hue (< from half of the bone)
-
Grade 2: burnt with brownish hue (> of half of the bone)
-
Grade 3: completely charred (black shade)
-
Grade 4: black and white shade (< half charred)
-
Grade 5: black and white shade (> half charred)
-
Grade 6: completely charred (totally white)
Differential thermal alteration processes have also been documented by noting bones with two or three colours on the same surfaces, as well as their position on the bone and the number of surfaces with those alterations. This can help to determine the function of the exposure to fire: either roasting meat (Cáceres et al., 2002; Costamagno et al., 2005; McKinley, 2008 and 2016) or use of bones as fuel (Costamagno et al., 2005; Yravedra et al., 2005; Yravedra & Uzquiano, 2013).
Marks caused by carnivores were also taken into account, classifying them according to their morphology (Haynes, 1980; Selvaggio, 1994). Finally, alterations related to the formation of the archaeological deposit have been considered, such as weathering, precipitation of calcium carbonate, the presence of manganese oxide and post-depositional processes (root marks, trampling and polish) (Behrensmeyer, 1978; Andrews & Cook, 1985; Behrensmeyer et al., 1986; Fernández-Jalvo & Andrews, 2016).
The taxonomic classification of the marine molluscs was carried out with the reference collection in the Zooarchaeology Laboratory at the University of Salamanca and publications that specify the habitat of the species living on the northern Spanish coast today (e.g. Palacios & Vega, 1997; Trigo et al., 2018). WoRMS nomenclature was used for the marine invertebrates (WoRMS Editorial Board, 2024).
The molluscs were classified to species level, wherever possible, based on their general attributes (shape, ornamentations, etc.), and the MNI was calculated from the NR and NISP, taking into account the different fragmentation categories. To calculate the MNI, the apices of the Patella genus were counted, and the apex and umbilical zone in the case of the other genera (e.g. Madariaga, 1975a and 1975b; Moreno, 1994; Álvarez-Fernández, 2009 and 2013; Álvarez Fernández et al., 2019).
The various alterations to the shells, caused by epifauna, marine perforating organisms, marine erosion, anthropogenic action, etc. (e.g. Fretter & Graham, 1962; O’Connor & Lamont, 1978; Laukner, 1980; Baxter, 1984; Fernández López, 1998; Álvarez-Fernández, 2006 and 2013; Southward, 2008) were noted to discern whether the molluscs were taken to the site as food or for other purposes, such as raw material to make objects of adornment.
Results
The study has documented a total of 40,940 remains of vertebrates (mammals, birds and fish) and invertebrates (marine molluscs and a crustacean) from the lower Magdalenian levels in El Cierro Cave. Of these, 98.2% come from the 1977–1979 excavations, while 1.8% come from the 2014 intervention. A total of 23,023 (56% of the total) come from Cierro G1: remains of large mammals are the most numerous (NR = 15,281), followed by molluscs (NR = 7729), fish (NR = 10) and birds (NR = 3). From Cierro G, 8608 remains (21% of the total) consist of 8151 remains of mammals, 456 of molluscs and one of a bird. The 9309 remains (23% of the total) from Cierro F correspond to mammals (NR = 8890), molluscs (NR = 417), one bird and one crustacean. No remains of fish were identified in this level (Tables 2 and 3).
Vertebrates
Large Mammals
In Cierro G1, 2592 remains (17%) were identified to taxonomic level, and the rest were assigned to their corresponding size categories. Based on the NISP, red deer is the most common animal, with 2363 remains (91.2%), followed by Iberian ibex with 139 (5.4%), large bovids with 51 (2%), roe deer with 23 (0.9%) and horse with 14 (0.5%). Carnivores are present with two elements belonging to a canid. In the size categories, medium-sized mammals are most abundant (NR = 3212) followed by the small (NR = 835) and large categories (NR = 88).
In Cierro G, 6.8% (NR = 558) of the remains were identified taxonomically. Red deer is the best represented species in the NISP, with 509 remains (91.2%), followed by Iberian ibex with 34 (6.1%), roe deer and Bos/bison with six remains each and horse with three. In this level, three teeth were initially thought to belong to an indeterminate marine mammal (Portero et al., 2019). However, ZooMS analyses of one of them concluded that they correspond to a cervid species other than reindeer. The morphology of the teeth resembles the first premolar of red deer (Mech et al., 1970), a dental anomaly that is absent in most modern animals and which may explain their scarcity in this archaeological deposit. In the size categories, medium-sized mammals are the most abundant (NR = 1295), followed by small (NR = 271) and large animals (NR = 29).
In Cierro F, 1088 remains (12.2%) were classified taxonomically. Red deer is again the main species, with 1005 remains (92.4% of the NISP), followed by Iberian ibex with 70 (6.4%), roe deer with seven (0.6%) and large bovid with six (0.5%). Medium-sized mammals are again the most abundant in the size categories (NR = 498), followed by small (NR = 357) and large animals (NR = 9).
The MNE displays a very similar distribution to the NISP in the three levels as red deer is the species with the largest number of elements, followed by Iberian ibex, large bovid, roe deer, horse and canid. All the skeletal parts of red deer are represented in the three levels and cranial elements (mainly teeth) and the hind limbs are the most abundant in all of them while axial elements are the least well-represented. For Iberian ibex, the representation patterns differ considerably from one level to the other. In Cierro G and Cierro G1, cranial elements are abundant, as teeth are very common. However, in Cierro F the best represented elements are the limbs, especially the hind limbs, and no axial elements have been recorded. Similarly, in Cierro G and Cierro G1, a high percentage of cranial elements of roe deer have been documented, mainly teeth and mandibles, whereas in Cierro F only limb elements were identified. In the three levels, distal appendicular elements of this animal are very abundant, especially the phalanges. Large bovids are represented mainly by distal appendicular elements, mostly phalanges and carpal and tarsal bones. Cranial elements are abundant in Cierro G1 and F, but absent in Cierro G. Horse, present in Cierro G and G1, is represented mainly by teeth.
In the size categories, the medium and small mammals are represented by a large number of axial elements, mainly ribs and vertebrae, although limb elements are also quite abundant and in the majority in Cierro G. A large number of appendicular elements are observed in the large mammal category, mostly long bone shafts. Axial elements are also abundant, whereas cranial remains are very scarce (Table 4).
In terms of the MNI, red deer is the most abundant species in all levels. The mortality patterns indicate that animals of practically all age ranges are present. Although young adults are most common in all levels, there is a significant number of infantile and juvenile individuals. This indicates that hunting focused mainly on younger animals. Iberian ibex, the second most common species, is represented by both adult and juvenile individuals, except in Cierro F, with only two adult individuals. For the few individuals of roe deer and large bovids, there is only an immature individual of each taxon in Cierro G, and the rest are young adults. All the remains of horse and canid in Cierro G1 belong to adult animals. In the size categories, the medium and small mammals are represented by a large number of young adults and immature individuals. No senile animals have been identified in those categories. For the large mammals, except for an immature specimen in Cierro G1, the other remains belong to adult individuals, as observed in the case of large bovid and horse (Table 5).
The population distribution of the different species and size categories based on the zonations established by Discamps & Costamagno (2015) for ternary diagrams (Stiner, 1990) shows for Cierro G1 a general “JPO” type profile. The mortality of young and adult individuals dominates over senile individuals, except for the roe deer, which presents a “P” profile, with adults predominating over the rest of the age categories. In the case of Cierro G, we have a “JPO” profile where the mortality of young and adult individuals predominates over that of senile individuals. Outside this profile, we have the species represented by a single adult or juvenile individual and the small mammals that show a “P” profile, with young adults predominating over immature and senile individuals. For Cierro F, we find a general profile where the mortality of adult individuals predominates over juveniles and senile individuals. Outside this range would be roe deer and Bos/bison consisting only of adult individuals (Fig. s1, s2 and s3).
The correlation analysis between %MAU and the different general utility indices (MGUI and SFUI) in Cierro G1 shows a negative coefficient of weak intensity (r < 0) for the red deer, while for the marrow and fat utility indices the relationships are direct and of moderate and weak intensity, respectively. These indices could reveal the transport to the cavity of those skeletal elements with higher marrow and grease utilisation. In this sense, the only relationship that shows statistical significance (p-value < 0.001) is the one that relates %MAU with the marrow index (Table s1 and Fig. s4). For Iberian ibex, all correlation data with utility indices are direct. In the case of grease and marrow utility, the intensity of the relationship is weak, while for MGUI and the meat utility index, the intensity is moderate. Only for the correlation between %MAU and meat utility index we have a p-value < 0.05 (Table s4 and Fig. s7).
In Cierro G, red deer show an inverse and weak relationship between the general food utility indices (MGUI and SFUI) and the percentage of anatomical representation (%MAU). This negative relationship holds for meat utility values and becomes positive, but not very strong for grease and marrow utility values. For this taxon, none of the correlations is statistically significant (Table s2 and Fig. s5). For Iberian ibex, we find a weak to moderate inverse relationship between food utility indices and anatomical representation. However, none of the resulting correlations is statistically significant (Table s5 and Fig. s8).
Red deer shows in all cases a direct relationship between the different utility indices and the anatomical representation in Cierro F. This relationship is practically null between the %MAU, the general utility indices (MGUI and SFUI) and the meat utility index. This relationship is stronger when correlating %MAU and medullary utility, the latter being the only one that is also statistically significant (p-value < 0.01) (Table s3 and Fig. s6). For Iberian ibex, we observed a direct correlation between the general utility indices and %MAU. In this taxon, only the relationship between %MAU and grease utility index is statistically significant (p-value < 0.05) (Table s6 and Fig. s9).
The taphonomic study revealed the extent of the anthropogenic manipulation of these faunal remains, attested by cutmarks, intentional breakage of bones and heat alteration processes (Fig. 3 and Table 6). Cutmarks have been documented on 11.6% of the NISP from Cierro G1, 11.4% in Cierro G and 6% in Cierro F. The processing of carcasses has been observed on the remains of red deer, Iberian ibex, roe deer, large bovid and horse, and also on the large, medium and small mammals.

Anthropogenic marks on remains from the lower Magdalenian levels in El Cierro. 1 Red deer mandible with evisceration marks (Cierro G1). 2 Red deer metatarsal with defleshing marks (Cierro G). 3 Roe deer astragalus with disarticulation marks (Cierro F). 4 Red deer metacarpal with a percussion mark (Cierro G). 5 Red deer tibia with consecutive percussion marks and an opposite notch (Cierro F). 6 Flakes (Cierro G1). 7 Burnt bones with different tones of colour
Red deer was the main prey processed in all the Magdalenian levels. The abundance of the anthropogenic marks has enabled an appreciation of the specific treatment given to this ungulate. Evidence has been observed of the complete processing of all the anatomical parts of the animal (cranial, axial and appendicular). The tasks involved skinning, disarticulating, dismembering and defleshing the body and also scraping the periosteum before breaking the bones to access the marrow (Table 7). Most of the cutmarks in these levels are located on the autopodia; even though in Cierro F most marks are on the tibiae and femora, the autopodia also display a large percentage of them (Fig. s10 , s11 and s12). The percentages of the frequencies of cutmarks on red deer bones show that in Cierro G1, axial elements display a high proportion of marks. This may indicate that the axial skeleton was also processed intensely although, owing to the low NISP of those skeletal elements, they may be over-represented. In Cierro G and F, cutmarks are most frequent on autopodia and zeugopodia (Fig. s13).
For the mammals in the medium-size category, the butchery patterns are similar to those of red deer in all the Magdalenian levels. In this case, owing to the large number of long bone shafts and ribs, those are the elements with the largest percentage of cutmarks. However, according to the frequency of the marks, femora and metapodia display a high percentage of marks (Fig. s14). As in the case of red deer, tasks of skinning, evisceration, disarticulation, dismembering, filleting and scraping the periosteum have been identified.
Few Iberian ibex remains display cutmarks in the three levels (NR = 17). They are mainly defleshing marks on femur shafts and vertebrae and of disarticulation on the pelvis and phalanges. Only three marks have been recognised on roe deer remains: disarticulation marks on a talus and a phalanx and a defleshing mark on a metatarsal.
The whole meat-processing sequence is present in the small mammal category in the three levels. They appear in all the skeletal regions, although they are most usual on ribs, vertebrae and long bones. According to the frequency of these cutmarks, limb bones are, in many cases, processed more often than axial and cranial elements (Fig. s15).
The few cutmarks documented on large bovid remains (NR = 10) are located on the autopodia and are disarticulation and filleting marks on the limbs of animals from Cierro G1 and G. In contrast, the only cutmarks identified on horse remains are on cranial elements (NR = 2) from Cierro G1. Mammals in the large size category display a similar pattern to that of large bovid, as mostly defleshing marks are observed on long bones and appendicular elements, i.e. the elements that provide the most meat on the animal. Only one rib with evisceration marks was documented.
In addition to the butchery process, the intentional breakage of fresh bones to access the marrow has been observed in the three levels. These fractures were made by humans, as shown by the angles, planes, texture and direction of the fractures, and above all by the impact notches, percussion pits, opposite notches and bone flakes documented on the long bones of red deer, Iberian ibex and large bovid, and on remains in the large, medium and small categories (Table 8).
The fractures appear mainly on metapodia and first and second phalanges, which display a large number of percussion notches (in 19.4% of metacarpal, 18.8% of metatarsal, 11.6% of metapods, 19.2% of first phalanges and 16% of second phalanges). However, a wide variety of axial and cranial elements, mostly mandibles and ribs were also broken; the latter by bending them by hand. The number of remains that were fractured, especially of red deer, has enabled a systematisation of the way that the marrow was extracted from each bone, by analysing the morphology and location of the impact. In addition, percussion notches have been observed repeatedly on specific sides and portions of bone within a high degree of standardisation (Fig. s16, s17 and s18).
Thus, red deer femora were mainly struck on both lateral faces of the shaft and less often on the anterior and posterior faces. Humeri were broken mostly on the anterior and posterior faces of the shaft and the metaphysis, in both its proximal part (mainly in Cierro G and F) and distal part (especially in Cierro G1). On the radii, the impacts were mainly applied to the anterior and posterior faces of the proximal part of the shaft. In Cierro G, the fracture pattern changes to the distal part, where both lateral faces were struck. Metapodia are the elements on which the most impacts were documented (NR = 235). Two types of breakage can be defined: one is a preference for striking the proximal part of the shaft and metaphysis on the upper face of the bone, near the longitudinal groove, while the other type is affected mainly by the sides of the shaft. Different fracture patterns are observed on the first and second phalanx. The first phalanx was struck mostly on the anterior and posterior faces of the shaft, which created longitudinal fracture planes. In contrast, the second phalanges were impacted mainly on the lateral faces, producing oblique fracture planes. A series of refits were made of fractured specimens, and this has shown how the marrow was extracted from those bone elements (Fig. 4). In some cases, an anvil was used to break long bones, which caused numerous opposite impacts on the bone shafts (on between 10 and 25% of the bones). Additionally, a large number of remains display isolated percussion notches, especially phalanges. A total of 271 flakes broken off bones during the percussion process were recovered from the three levels (Cierro G1 = 176; Cierro G = 25; Cierro F = 70).

Impacts on the anterior and posterior faces of the red deer first phalanges from Cierro G1 and their respective refits
Less evidence of fractures has been observed on the axial and cranial skeleton. Most of the fractures on cranial elements are in the mandible ramus, mainly in the vestibular region beneath the molar line. The objective was to break the lower part of the ramus crosswise. Less often, the mandible was broken longitudinally after the third molar, separating the ramus from the gonion. On the axial skeleton, the fractures appear mainly on parts of the pelvis and the neck of scapulae.
Ribs and flat bones were also broken by flexion. However, owing to the difficulty in the taxonomic identification of those elements, most of them were included in the medium-size category.
For Iberian ibex, roe deer and large bovid, although some elements were fractured intentionally, they are not sufficient in number to establish fracture patterns. All the Iberian ibex remains with impact marks correspond to limb bones, mainly femora, phalanges and metapodia. All the fractured large bovid bones are limb bones, particularly autopodia, whereas for roe deer only a mandible from Cierro G1 displays a percussion mark.
In the size categories, although fractured elements are very common among the medium and small mammals, most percussion marks appear on limb bone shafts that are difficult to classify. On the few elements that could be studied, they do not differ in the form of breakage from the pattern observed for red deer and Iberian ibex. In contrast, among the large mammals, all the fractured elements are limb bones, except in Cierro G1, where a fractured mandible, pelvis and rib have been documented.
Thermal alterations have been observed on 7.9% of the remains from Cierro G1, 14% from Cierro G and 10.7% from Cierro F. They affect red deer, Iberian ibex, roe deer, large bovids and all size categories. Most of the altered bones display brown or brown-black colouring, which indicates exposure to low temperatures (Table 9). To be exact, between 54.5 and 67.4% of the burnt remains can be classified in stage 1 of Stiner et al. (1995). They would have been exposed to heat below 400 °C. While this alone is not evidence of roasting, and other factors influence the colour of a bone, it is compatible with the exposure of bones with soft tissue to sources of heat during a relatively short time.
Other taphonomic agents that have altered the remains include carnivores, which have affected between 5.1% and 10.6%. They left marks of scores, pits, furrowing, gnawing, crenulated edges and digestion on bones of red deer, Iberian ibex, roe deer, horse and large bovid. These marks equally appear in all the size categories (Tables 10 and 11). In over 40 cases, cutmarks and carnivore marks are seen on the same bone. The affected remains are mostly axial elements, including a considerable number of scapulae, vertebrae, pelvis and ribs. Rodents also modified the faunal remains and left facetted marks with their teeth on the surface of the bones.
Other modifications to the faunal record in all the Magdalenian levels at El Cierro are the vermiculations caused by roots, which have been documented on between 16 and 26.9% of the remains. These alterations have affected all the taxa and size categories and indicate the constant entry of light and humidity in the cave during the formation of the strata. In connection with the humidity, a large number of bones are rounded by water action (from 3.7 to 6.2% of the NR), although in most cases low energy flow that on very few occasions has totally polished their surface. Some remains have been documented with precipitated calcium carbonate, or with manganese oxides that have stained them. Weathering has also been an important factor of alteration and has affected from 6.3 to 10.1% of all the remains (Fig. s19). Finally, the activity of different biological agents in the cave has caused trampling marks in the form of fine and short striations distributed at random on the surface of a very small percentage of the bones (< 3.5% of the NR).
The length and season of the year of the Magdalenian occupations differ depending on each level. In general terms, the occupations tended to take place in spring and summer in all levels. However, this tendency is conditioned by the large number of fawns killed in their first months of life. Nonetheless, the wide age range of red deer in the three levels may indicate that hunting was carried out from the site throughout the year, even if it was more intense in the summer. The other taxa display age ranges that also cover the summer while some indicate from autumn to winter; in the latter case, an Iberian ibex in Cierro G was hunted between autumn and spring (Fig. 5).

Seasonality of the occupations based on the age of death of large mammals in the Magdalenian levels at El Cierro Cave
Birds
Birds have been documented in Cierro G1 through a grey partridge coracoid, another of Anser/Branta spp., and a tibiotarsal of an indeterminate bird (see Tables 2 and 3). Only a Perdix perdix coracoid was found in Cierro G and a carpometacarpal of Corvus corax in Cierro F. No taphonomic alterations have been identified on any of these remains that might suggest they were processed or cooked.
Fish
Of the few fish remains (NR = 10), all from Cierro G1, only two vertebrae have been identified as belonging to the salmonid family (Álvarez-Fernández et al., 2022a) (see Tables 2 and 3). As in the case of the birds, none of these remains displays evidence of anthropogenic handling.
Invertebrates
The invertebrate remains that were recovered are all shells of marine molluscs (gastropods and bivalves). They have been collected from rocky substrates in the intertidal zone, the area where all documented species currently live. The one exception is a small plate belonging to a crustacean (Balanidae) (see Tables 2 and 3).
Gastropods predominate almost absolutely. In Cierro G1, most of the remains are the common periwinkle Littorina littorea (78.5% of the NR) and limpet (Patella sp.) (21.4% of the NR). Nucella lapillus (dog whelk) and Littorina obtusata (flat periwinkle) (0.01% of the NR) are also represented minimally. The MNI exhibits a similar distribution to the NISP, as Littorina littorea is the most abundant species (MNI = 1724), followed by Patella sp. (MNI = 127) and Patella vulgata (MNI = 60). Mytilus galloprovincialis (mussel), Nucella lapilus and Littorina obtusata are each represented by a single specimen. The winkles and limpets in the three levels are of a large size. Biometrics could only be carried out on gastropods from the G1 level. In the case of Littorina littorea, the maximum height is 28 mm; in the case of Patella vulgata, 39.5 mm.
The proportion of molluscs is similar in Cierro G as periwinkles again predominate in both the NR (400) and MNI (79), followed by limpets (NR = 55; MNI = 15). In Cierro F, although the number of limpets increases (NR = 141; MNI = 28), Littorina littorea is still the most abundant species (NR = 274; MNI = 86). Nucella lapilus and Littorina obtusata only appear with a single remain in Cierro F and G. Bivalves are represented by a mussel shell in Cierro G.
The outer surface of three shells from Cierro G1 (one of Littorina littorea and two of Patella vulgata) are perforated by annelids (Polydora sp.). These perforations are small galleries with two holes that connect with each other on the external surface but would not have harmed the live gastropods. A single plate of an acorn barnacle (Semibalanus balanoides balanoides) was found isolated in the sediment of Cierro F (Carriol & Álvarez-Fernández, 2015). It should be noted that the shells of the gastropod species Littorina obtusata and Nucella lapillus, both present in Cierro G and Cierro G1, are eroded by water action and sand and therefore were probably collected on a beach after the mollusc had died. Only the shells documented in Cierro G1 retain anthropogenic alterations. Thermal alteration is present only in < 0.1% of the NR (n = 4, one limpet and three periwinkles’ shells), and fracturing of the edges of limpets produced when separating their shells from the rock is documented in < 0.2% of the limpets shells (NR = 3). Shell surfaces documented in the three levels are corroded, powdery and whitish, probably because of a reaction to the conditions in which the shells have been buried. Very few displays precipitated calcium carbonate.
Discussion
The zooarchaeological data obtained from the Magdalenian sequence in El Cierro Cave shows that large mammals and, to a lesser extent, marine molluscs, were fundamental in the subsistence of the hunter-gatherer groups.
Red deer was the main prey in all levels, represented by over 90% of the NISP. In northern Spain, other sites with lower Magdalenian levels have been found to contain similar percentages of red deer, such as Las Caldas Level XIII (Altuna & Mariezkurrena, 2017), Cova Rosa Level B6 (Álvarez-Fernández et al., 2019), La Riera Level 19 (Altuna, 1986), El Linar Level 3c (Castaños, 2016), Las Aguas Level B (Castaños, 2016), Cualventi Level E (Castaños, 2016), Altamira Level 2 (Castaños & Castaños, 2014), El Juyo Levels 4 and 7 (Klein & Cruz Uribe, 1987) and Ekain Level VIII (Altuna & Mariezkurrena, 1984). Iberian ibex, roe deer, large bovid and horse were also present in the diet of the Magdalenian groups at El Cierro, but in small percentages. In northern Spain, sites with a significant predominance of red deer are usually located in open valleys near the modern coastline and at low altitudes (Portero et al., 2019).
All skeletal parts are represented among the red deer remains even though limb bones and those with larger marrow content are the most abundant. Regarding the transport strategy of this ungulate, we observe that the Magdalenian groups from El Cierro transported more frequently those elements with a higher medullary content. This is corroborated by the direct and statistically significant trend (except in Cierro G) between %MAU and marrow index. Furthermore, the correlation between %MAU and general utility indices is inverse for Cierro G1 and Cierro G and direct for Cierro F, although in neither case are they statistically significant. Therefore, it does not seem that the parts with the highest feed utilisation are being transported frequently, since in that case, we would find a direct and significant relationship between these variables (Grayson, 1989).
In the case of Iberian ibex, in addition to dental pieces, the main elements that make up the sample are represented by hindlimb and autopodia, although as we have seen, the percentages of representation of this species vary greatly between levels. Furthermore, the correlation between %MAU and the general utility indices (MGUI and SFUI) for the Iberian ibex is direct and moderate in Cierro G1 and Cierro F, being inverse and moderate in Cierro G, although in no case are they statistically significant. The only level where we obtain a significant relationship between %MAU and meat utility index is in Cierro F, which could indicate a transport of those elements with higher meat utilisation.
The study of the age patterns shows that individuals of practically all age ranges are present in the case of red deer, from neonates to immature, juvenile, adult and senile animals. There is a predominance of young adults in all levels, but a significant number of infantile and juvenile individuals have been documented. This indicates that hunting concentrated mainly on younger individuals, with a mortality profile of the “L”-shaped catastrophic type (Discamps & Costamagno, 2015), as can be seen from the ternary graphs. This suggests that hunting focused on herds of hinds and their calves, as observed in lower Magdalenian levels at such sites as La Riera 19 (Altuna, 1986), El Linar 3C (Castaños, 2016), Ekain VII (Altuna & Mariezkurrena, 1984) and Erralla V (Altuna & Mariezkurrena, 1985). This selective strategy did not rule out the occasional capture of other prey in the area: Iberian ibex, roe deer, horse and large bovids in the case of El Cierro, but with no apparent strategy, based on the age profiles that have been documented.
Although El Cierro Cave may have been occupied throughout the year during the lower Magdalenian, according to the documented capture regime, there is an occupational trend during the spring and summer for all levels, and despite the large volume of remains documented in these levels, it is likely that these occupations were not of a lasting nature and were rather a set of seasonal accumulations over a prolonged period of time. This type of seasonal occupation strategy could be related to the hunting pressure on resources and the need for active mobility for the recovery of the environment (Portero et al., 2024). In this sense, some research has developed a series of predictive mathematical models based on the demographic data of current animal populations (Stiner et al., 1999 and 2008; Beaver, 2007; Merkle et al., 2015; Venkataraman et al., 2017). These models predict the maximum number of individuals that can be hunted annually in a location without the species becoming extinct over time. To do this, they establish two predictive ranges, one based on a favourable population growth rate (high growth model - HGM) and the other on a population growth rate under unfavourable conditions (low growth model - LGM). An example applicable to this model on a population of 1000 red deer would indicate that only a total of 31 individuals could be hunted per year in a favourable population growth environment (HGM), whereas only ten individuals could be shot per year in an unfavourable growth environment (LGM) (Beaver, 2007).
However, the seasonality of occupations cannot be established very precisely solely by the age ranges of the individuals in the deposit, so we must be cautious with this type of interpretation.
The processing of the mammals found in the Magdalenian levels in El Cierro has been documented by cutmarks on the carcasses of red deer, roe deer, large bovids, horse and large, medium and small-sized animals. Red deer remains, as the main prey, display marks on all skeletal parts, and the whole processing chain can be followed from skinning to scraping the periosteum so as to access the marrow. While a large proportion of the cutmarks are related to the process of disarticulating the autopodia, in all levels, evidence of access to and consumption of the main sources of meat on femora and humeri have also been documented, as a large number of marks are observed in the places of the main muscles of the animals. Studies carried out in northern Spain at such sites as Coímbre and La Lluera have systematised the way in which large mammal carcasses were processed, based on the orientation, position and morphology of the cutmarks (López-Cisneros et al., 2019, 2020).
The intentional fracture of the bones while fresh to extract the marrow has been attested in the levels in El Cierro Cave through the impact marks on long bones and the flexion of flat bones. The large number of impacted remains, especially of red deer, has allowed a systematisation of how the marrow was obtained from each bone. This was carried out in a highly standardised way in the three Magdalenian levels. In general, the marrow of all the taxa was consumed intensely, and elements such as the phalanges and metapods of red deer were fractured repeatedly following a similar pattern in all levels. This frequent use might indicate a systematised strategy in the acquisition and conservation of this resource. Marrow is a very rich food in caloric energy and keeps in a suitable condition for consumption for weeks following the death of the animals (e.g. Mateos, 1999 and 2003; Blasco et al., 2019). Thus, the occupants of the cave may have consumed the soft parts and meat of the prey first and kept the elements with the largest marrow content to be eaten later or even stored for a period of time as a source of food in times of scarcity. Although the fracture of elements such as phalanges has been used as an indicator of nutritional stress in prehistoric peoples due to the low rate of caloric return (Binford, 1978; Jones & Metcalfe, 1988), experimental and ethnographic studies have shown that the presence of broken phalanges by themselves cannot be taken as evidence of nutritional stress and that their exploitation may be due to their taste and soft texture or to symbolic aspects (Morin, 2007; Costamagno & David, 2009; Jin & Mills, 2011; Costamagno & Rigaud, 2014, Kuntz et al., 2016, Birouste, 2020).
The alterations caused by heat observed on bones are compatible with roasting meat. A large number of bones burnt at low temperatures display double brown and black tones on the same surfaces, indicating the presence of soft tissue on them during their exposure to heat. These double colours have been detected in remains of red deer and large, medium and small mammals and are located mainly on autopodia and zeugopodia, i.e. the distal parts of the limbs. The double tones and their location mean that the burning of the bones after their burial can be ruled out (Bennett, 1999), and they are compatible with roasting the animals’ meat. Moreover, the large number of phalanges and metapodia affected in this way suggests that they were roasted before the bones were broken to access the marrow.
The activity of carnivores at El Cierro has generated a sufficiently high percentage of marks on the determinable remains that sporadic occupation of the cave by these animals cannot be ruled out. The presence of two medium-sized canid remains in Cierro G1 and the size of the marks observed could suggest that these animals were the agents that sporadically modified the Magdalenian bones from El Cierro. In this sense, and although we have not detected any superimposition of carnivore marks over the cut marks, we have documented more than 40 remains with both cut and carnivore marks on the same bone. Most of them are axial elements, metapods and phalanges. This leads us to believe that carnivore access must have been secondary, given the high frequency of cut marks on the main richest elements at all levels and for the main species documented, and the higher incidence of carnivory on those elements of lesser nutritional value. The action of carnivores, although detected in all size categories, shows a greater abundance on medium-sized animals, which are also the most consumed by human groups, being less in the remains of small and large sizes. This leads us to think that these carnivores were secondary agents in the access to animal carcasses, once they had been exploited by human groups, especially considering that medium-sized canids (such as wolves and dogs) tend to have a commensal synanthropic behaviour (Blumenschine & Selvaggio, 1988; Stiner, 1994; Yravedra, 2006; Baumann, 2023).
The bird remains do not display signs of anthropogenic impact, and therefore, it cannot be affirmed that they were consumed by the human groups. Little information is available about anthropogenic processing of birds in the lower Magdalenian in the region, only evidence of cutmarks on a lagopod, an Anatidae and indeterminate bird in Levels 8 and 9 at El Juyo (Rufà et al., 2022), on the humerus of a sea eagle from Cualventi E (Garcia Petit, 2016), on an Anatidae from El Linar 2 (Garcia Petit, 2016) and on an indeterminate bird from Level F at Arangas (Álvarez-Fernández et al., 2020). With a slightly more recent chronology, a Lagopus mutus remain with disarticulation marks was documented in Level 1CS at Tito Bustillo Cave (Álvarez-Fernández et al., 2022b). Some thermally altered remains of birds may indicate their processing in the lower Magdalenian. This is the case of remains of a corvid, an Anatidae, one of Lagopus lagopus and three of indeterminate birds in Levels 4 to 8 at El Juyo (Rufà et al., 2022), an Anatidae remain from Cualventi E (Garcia Petit, 2016), and an Alectoris sp./Perdix sp. radius from Level 1C1 at Tito Bustillo and two Accipitridae phalanges from Level 1CS in the same cave (Álvarez-Fernández et al., 2018b).
Fish would also have formed part of the diet of the occupants of El Cierro Cave as they would catch salmonids in the proximate River Sella, even though no taphonomic marks indicating their processing and consumption have been noted. Nor have we observed the alteration by other agents (Guillaud et al., 2021). No evidence has been found of the processing of fish at lower Magdalenian sites in north Spain, apart from some bones affected by heat. In Cova Rosa B6, salmonid and eel remains were altered thermally (Álvarez-Fernández et al., 2019), and two burnt remains of salmonids were found in Level 1CS at Tito Bustillo (Álvarez-Fernández et al., 2018b).
Molluscs were a complement in the diet of the Magdalenian groups at El Cierro. Although the energy intake of these animals is lower than that of mammals, molluscs may have played an important role in the diet of the groups given their high content of protein, vitamins, carbohydrates and polyunsaturated fatty acids such as omega-3 (Santhanam, 2018). These resource high salt levels may have provided a favourable dietary quality for human groups from an organoleptic point of view. In addition, the ease of harvesting makes seafood an accessible resource.
Periwinkles (L. littorea) and limpets (P. vulgata) were selected on rocky substrates. These are both cold-climate species that live on sheltered weed-covered shores. The presence of annelid perforations on three specimens from Cierro G1 shows that they were collected in sheltered areas. The presence of a plate of S. balanoides balanoides, a boreo-arctic species that lives in cold surface waters, indicates that the water temperature on the coast of Ribadesella was much colder than at present. This crustacean reached the deposit adhered to the surface of a shell and the plate became detached later (Carriol & Álvarez-Fernández, 2015).
The larger proportion of winkles than limpets in the three levels at El Cierro is unusual in northern Spain. Thus, in Layer B6 at Cova Rosa (Álvarez-Fernández et al., 2019), Levels 18–20 at La Riera (Ortea, 1986), Levels 1 and 2 at Altamira (Álvarez-Fernández, 2009), Levels 9 to 4 at El Juyo (Madariaga & Fernández, 1987) and Level V at Erralla (Altuna, 1985b), P. vulgata and L. littorea represent nearly 99% of the mollusc remains, but the former always predominates over the latter. They are all characterised by the selection of large (adult) (Álvarez Fernández, 2011).
Gastropod Shells at El Cierro were collected by hand without difficulty on the rocky shore. In the case of limpets, their shells would be separated from the rocks with poorly elaborate tools (fragments of bones or lithics), causing fractures on their edges. Its form of consumption was not the result of direct contact with fire, given the few remains that are thermoaltered. Perhaps they were consumed boiled or raw. The post-depositional alterations that the remains present, particularly the decalcification, prevent obtaining more information about the collection strategies and their consumption.
This subsistence strategy detected by the faunal resources in the Magdalenian levels at El Cierro resembles the situation at sites in the Sella valley, such as Cova Rosa (Álvarez-Fernández et al., 2019), and at other sites in northern Spain, like Las Caldas (Corchón, 2017b), Arangas (Álvarez-Fernández et al., 2020), Santimamiñe (López Quintana, 2011) and Erralla (Altuna et al., 1985). In all of them, large mammals were the main source of food, while fish, molluscs and birds were complementary resources in the diet of the hunter-gatherer groups.
It should be noted that plant resources were also widely exploited for subsistence by human groups during the lower Magdalenian, as demonstrated by anthracological studies carried out at Cierro G1 (Uzquiano, 2019), which indicate the use of Fabaceae collected in valley areas, especially in siliceous soils. The importance of plant foods in subsistence has also been highlighted for this period based on the analysis of phytoliths (Zurro, 2010) and the analysis of dental calculi from human teeth (Power et al., 2015).
Conclusion
The importance of animal resources in the subsistence of the Magdalenian inhabitants of Levels G1, G and F in El Cierro Cave has been documented through their remains. The data obtained show that in the lower Magdalenian, the hunting strategy focused on red deer as the main prey. This is indicated by both the high percentage of remains and the taphonomic evidence of their use. Other species, like Iberian ibex, roe deer, horse and large bovids, were also consumed. Molluscs and fish were a complement to the part of the diet with an animal origin, as a small number of individuals have been documented compared with the abundance of large mammals. No evidence of anthropogenic manipulation of avifauna supports the processing and consumption of birds.
The taphonomic study has determined that the accumulation of large mammal remains in the lower Magdalenian is the result of anthropogenic activity, since the complete operational chain of the processing of the prey has been documented, including skinning, evisceration, disarticulation, dismembering, filleting and scraping the periosteum, especially of red deer. In addition to the consumption of the meat, the present study has determined a systematic approach to the acquisition of the marrow inside the bones, mainly the long limb bones of ungulates. The pattern is similar in the three levels, which indicates continuity in the way of acquiring and processing animal fats, which would have been of great importance in the diet of the groups because marrow stays in good condition longer than meat and is therefore a resource that can be stored to be consumed when necessary.
Fire was used in the processing of meat, as shown by thermally altered bones in shades of colour associated with exposure to low temperatures. The double colours on many distal and proximal limb bones are due to the variable quantity of meat tissue on those elements during the roasting process.
The zooarchaeological study has also taken into consideration the time of the seasonal occupation of the cave. The levels in the sequence at El Cierro are the result of several occupations during the period from spring to autumn. In those occupations, the hunting strategy concentrated on herds of young adult red deer and their calves, whose complete carcasses could be transported to the site. However, these impressions are based only on the data provided by the skeletal age of the mammals and need to be confirmed more precisely by other seasonality studies in the future (e.g. cementochronology, dental mesowear and microwear analysis, isotopic analysis).
Finally, this study has shown the importance of studying all the biotic resources in a deposit as a whole, as this is the only way to obtain a full panorama of the subsistence strategies of past human groups. It enables an assessment of the relative importance of each of the animal resources in the diet, the way of obtaining nutrients, the places in which the resources were obtained and the complexity of the processing and consumption of animals by the groups.
Data Availability
The data generated and analysed during the present study are available on request from the corresponding author. The faunal remains studied are preserved in the Museo Arqueológico de Asturias (Oviedo).
References
Altuna, J. (1972). Fauna de mamíferos de los yacimientos prehistóricos de Guipúzcoa, con catálogo de los mamíferos cuaternarios del Cantábrico y Pirineo Occidental. Munibe. Sociedad De Ciencias Naturales Aranzadi (san Sebastian), 24(1–4), 1–464.
Altuna, J. (1976). Los Mamíferos del yacimiento prehistórico de Tito Bustillo (Asturias). In Moure, J. A. & Cano, M. (Eds.), Excavaciones en la cueva de Tito Bustillo (Asturias): Trabajos de 1975. (pp. 149–194), Instituto de Estudios Asturianos.
Altuna, J. (1984). Historia de las excavaciones. Descripción del yacimiento. Resumen estratigráfico del relleno. Utilización del espacio. Dataciones absolutas. In Altuna, J. & Merino, J.M. (Eds.), El yacimiento prehistórico de la cueva de Ekain (Deba, Guipúzcoa). (pp. 17–45), Eusko Ikaskuntza
Altuna, J. (1985a). Dataciones del C14. Comparación con las dataciones de otros yacimientos. In Altuna, J., Baldeón, A. & Mariezkurrena, K. (Eds.), Cazadores Magdalenienses de la Cueva de Erralla. (Munibe, 37, pp. 25–28), Sociedad de Ciencias Aranzadi
Altuna, J. (1985b). Los moluscos marinos de Erralla. In Altuna, J., Baldeón, A. & Mariezkurrena, K. (Eds.), Cazadores Magdalenienses de la Cueva de Erralla. (Munibe, 37, pp. 119–121), Sociedad de Ciencias Aranzadi
Altuna, J. (1986). The mammalian faunas from the prehistoric site of La Riera. In Straus, L. G. & Clark, G. (Eds), La Riera Cave. Stone Age Hunter-Gatherer adaptations in Northern Spain. (Anthropological Research Papers, 36, pp. 237–274), Arizona State University
Altuna, J. (2012). Cueva De Ekain. Excavación. Arkeoikuska, 2011, 336–338.
Altuna, J.& Mariezkurrena, K. (1984). Bases de subsistencia de origen animal en el yacimiento de Ekain. In Altuna, J. & Merino, J.M. (Eds.), El yacimiento prehistórico de la cueva de Ekain (Deba, Guipúzcoa). (pp. 211–280), Eusko Ikaskuntza
Altuna, J.& Mariezkurrena, K. (1985). Bases de subsistencia de los pobladores de Erralla: Macromamíferos. In Altuna, J., Baldeón, A., Mariezkurrena, K. (Eds.), Cazadores Magdalenienses de la Cueva de Erralla. (Munibe, 37, pp. 87–117), Sociedad de Ciencias Aranzadi
Altuna, J. & Mariezkurrena, K. (2017). Bases de subsistencia de origen animal durante el Magdaleniense en la cueva de Las Caldas (Priorio, Oviedo). In Corchón, S. (Ed.), La cueva de Las Caldas (Priorio, Oviedo): ocupaciones solutrenses, análisis espaciales y arte parietal. (Estudios Históricos & Geográficos, 166, pp. 55–119), Ediciones Universidad de Salamanca
Altuna, J., Baldeón, A., Mariezkurrena, K. (1985). Cazadores Magdalenienses de la Cueva de Erralla. (Munibe, 37), Sociedad de Ciencias Aranzadi
Álvarez-Alonso, D. & Jordá Pardo, J.F. (2017). Secuencia estratigráfica, radiocarbono y cronoestratigrafía del registro del Pleistoceno superior de la cueva de Coímbre (zona B) (Asturias, España). In Álvarez-Alonso, D. & Yravedra, J. (Eds.). La cueva de Coímbre (Asturias, Spain). Ocupaciones magdalenienses en el valle del Cares. (pp. 194–217), Fundación Masaveu
Álvarez-Fernández, E. (2006). Los objetos de adorno-colgantes del Paleolítico Superior y del Mesolítico en la Cornisa Cantábrica y en el valle del Ebro: una visión europea. Ediciones Universidad de Salamanca, (Colección Vítor, 195)
Álvarez-Fernández, E. (2009). Análisis arqueomalacológico de la cueva de Altamira (Santillana del Mar, Cantabria): Excavaciones de J. González Echegaray y L.G Freeman. Complutum, 20, 55–69.
Álvarez-Fernández, E. (2011). Humans and marine resource interaction reappraised: Archaeofauna remains during the late Pleistocene and Holocene in Cantabrian Spain. Journal of Anthropological Archaeology, 30(3), 327–343.
Álvarez-Fernández, E. (2013). Shellfish gathering in Cantabrian Spain during the Magdalenian: The cave of “Tito Bustillo” (Asturias, Northern Spain). L’Anthropologie, 117(1), 62–93.
Álvarez-Fernández, E. (2017). Los moluscos marinos de la cueva de Las Caldas (San Juan de Priorio, Oviedo). In Corchón, M.S. (Ed.), La cueva de Las Caldas (Priorio, Oviedo): ocupaciones solutrenses, análisis espaciales y arte parietal. (Estudios Históricos & Geográficos, 166, pp. 159–171), Ediciones Universidad de Salamanca
Álvarez-Fernández, E., Álvarez-Alonso, D., Bécares, J., Carral, P., Carriol, R.-P., Chauvin, A., Cubas, M., Cueto, M., Domingo, R., Douka, K., Jordá Pardo, J. F., Murelaga, X., Portero, R., Rivero, O., Tapia, J., Tarriño, A., & Teira, L. C. (2016). Nouvelles données sur le Magdalénien inférieur de la Région Cantabrique : Le Niveau F de la grotte de El Cierro (Ribadesella, Asturies, Espagne). L’Anthropologie, 120(5), 537–567.
Álvarez-Fernández, E., Bécares, J., Jordá Pardo, J.F., Aguirre, A., Álvarez-Alonso, D., Andrés-Herrero, M De., Aparicio, M.T., Barrera, I., Carral, P., Carriol, R.P., Chauvin, A., Cubas, M., Cueto, M., Domingo, R., Douka, K., Duarte, C., Elorza, M., Fernández-Gómez, M.J., Gabriel, S., Haber, M., Iriarte, M.J., Julián, M.-A., Lepage, J., Llave, C., Martín-Jarque, S., Murelaga, X., Osete, M.L., Palencia, A., Portero, R., Rivero, M., Rivero, O., Tapia, J., Tarriño, A., Teira, L.C., Uzquiano, P. & Arias, P. (2018a). Excavaciones arqueológicas en El Cierro (Fresnu, Ribadesella). Campañas de Excavación e Investigación 1977–1979, 2014 y 2016. In Excavaciones Arqueológicas en Asturias 2013–2016. (pp. 93–106.), Consejería de Educación y Cultura del Gobierno del Principado de Asturias
Álvarez-Fernández, E., Cueto, M., Tapia, J., Aparicio, M. T., Douka, K., Elorza, M., Gabriel, S., García-Ibaibarriaga, N., Suárez-Bilbao, A., & Arias, P. (2018b). Nuevos datos cronoestratigráficos y arqueozoológicos de la cueva de Tito Bustillo-Área de Estancia (Ribadesella, Asturias). Kobie (serie Anejo), 18, 109–122.
Álvarez-Fernández, E., Andrés, M., Aparicio, M., Elorza, M., Gabriel, S., García-Ibaibarriaga, N., Murelaga, X., Portero, R., Sensburg, M., Suarez-Bilbao, N., Tapia, J., Uzquiano, P., Becares, J., Cubas, M., & Jordá Pardo, J. F. (2019). Biotic resources in the Lower Magdalenian at Cova Rosa (Sardeu, Asturias, Spain). Quaternary International, 506, 25–34.
Álvarez-Fernández, E., Cubas, M., Aparicio, M. T., Cueto, M., Elorza, M., Fernández, P., Gabriel, S., García-Ibaibarriaga, N., Portero, R., Suarez-Bilbao, N., Tapia, J., Teira, L. C., Uzquiano, P., & Arias, P. (2020). New data for the late Upper Palaeolithic in the Cantabrian region: Arangas Cave (Cabrales, Asturias, Spain). Journal of Archaeological Science: Reports, 29, 1–11.
Álvarez-Fernández, E., Jordá Pardo, J. F., Arias, P., Bécares, J., Martín-Jarque, S., Portero, R., Teira, L., & Douka, K. (2021). Radiocarbon dates for the late Pleistocene and early Holocene occupations of Cova Rosa (Ribadesella, Asturias, Spain). Radiocarbon, 63(3), 1–20.
Álvarez-Fernández, E., Arias, P., Bécares, J., Cubas, M., Elorza, M., Gabriel, S., Martín-Jarque, S., Portero, R. & Jordá Pardo, J.F. (2022a). Intervenciones arqueológicas en la cueva de El Cierro (Fresnu, Ribadesella, Asturias): síntesis de los datos disponibles procedentes de los recientes trabajos realizados en tres zonas del yacimiento. In Jordá Pardo, J.F., Martín-Jarque, S., Portero, R & Álvarez-Fernández, E. (Eds.), Descendiendo el río Sella. Una (re) visión de la Arqueología Prehistórica del valle del Sella (Asturias, España). (Entemu XIX, pp. 133–162), UNED
Álvarez-Fernández, E., Tapia, J., Aguirre-Uribesalgo, A., Arias, P., Camarós, E., Cerezo-Fernández, R., García-Alonso, B., Martín, N., Martín-Jarque, S., Peyroteo-Stjerna, R., Portero, R., Teira, L.C. & Cueto, M. (2022b). La cueva de Tito Bustillo (Ardines, Ribadesella, Asturias, España): intervenciones arqueológicas en el Área de Estancia. In Jordá Pardo, J.F., Martín-Jarque, S., Portero, R. & Álvarez-Fernández, E. (Eds.), Descendiendo el río Sella. Una (re)visión de la Arqueología Prehistórica del valle del Sella (Asturias, España). (Entemu XIX, pp. 247–266). UNED (Asturias)
Andrews, P., & Cook, J. (1985). Natural modifications to bones in a temperate setting. Man, New Series, 20(4), 675–691.
Arias, P., Ontañón, R., Álvarez-Fernández, E., Aparicio, M.T., Chauvin, A., Conte, I., Cueto, M., González Urquijo, J.E., Ibáñez, J.J., Tapia, J. & Teira, L.C. (2005). La estructura magdaleniense de La Garma A: aproximación a la organización espacial de un hábitat paleolítico. In Bicho, N. & Corchón, M.S. (Coords.), O Paleolítico: actas do IV Congresso de Arqueologia Peninsular (Faro, 14 a 19 de Setembro de 2004). (Centro de Estudos de Patrimonio, pp. 123 – 142). Universidade do Algarve
Azorit, C., Analla, M., Carrasco, R., Calvo, J. A., & Muñoz Cobo, J. (2002). Teeth eruption pattern in red deer (Cervus elaphus hispanicus) in southern Spain. Anales De Biología, 24, 107–114.
Azorit, C., Analla, M., Carrasco, R., & Muñoz-Cobo, J. (2003). Determinación de la edad por desgaste dental en el ciervo ibérico (Cervus elaphus hispanicus, Erxleben, 1777) (Mammalia, Cervidae). Boletín de la Real Sociedad Española de Historia Natural (Sec. Biol.), 98(1–4), 123–134.
Bacher, A. (1967). Vergleichend morphologische Untersuchungen an Einzelknochen des postkranialen Skeletts in Mitteleuropa vorkommender Schwäne und Gänse. Ludwig-Maximilians Universität.
Barandiarán, I. & González-Echegaray, J. (1981). Dataciones por el carbono 14. In González-Echegaray, J. & Barandiaran, I. (Eds), El Paleolítico Superior de la cueva del Rascaño. (Centro de Investigación y Museo de Altamira, 3, pp. 212–214), Ministerio de Cultura. Dirección General de Bellas Artes, Archivos y Bibliotecas
Barone, R. (1966). Atlas de mammifères domestiques. Masson.
Baumann, C. (2023). The paleo-synanthropic niche: A first attempt to define animal’s adaptation to a human-made micro-environment in the Late Pleistocene. Archaeological and Anthropological Sciences, 15, 63.
Baxter, J. M. (1984). The incidence of Polydora Ciliata and Cliona Celata Boring the shell of Patella vulgata in Orkney. Journal of the Marine Biological Association of the United of Kingdom, 64(3), 728–729.
Beaver, J.E. (2007). Paleolithic ungulate hunting: Simulation and mathematical modeling for archaeological inference and explanation. (Unpublished PhD) University of Arizona
Behrensmeyer, A. K. (1978). Taphonomic and ecological information from bone weathering. Paleobiology, 4(2), 150–162.
Behrensmeyer, A. K., Gordon, K. D., & Yanagi, G. T. (1986). Trampling as a cause of bone surface damage and pseudocutmarks. Nature, 319, 768–771.
Bennett, J. L. (1999). Thermal alteration of buried bone. Journal of Archaeological Science, 26, 1–8.
Binford, L. R. (1978). Nunamiut Ethnoarchaeology. Academy Press.
Binford, L. R. (1981). Bones: Ancient man and modern myths. Academy Press.
Birouste, C. (2020). Human and animal individuals in the Middle Magdalenian. Journal of Archaeological Method and Theory, 27, 607–630.
Blasco, R., Rosell, J., Arilla, M., Margalidas, A., Villalbas, D., Gopher, A., & Barkai, R. (2019). Bone marrow storage and delayed consumption at Middle Pleistocene Qesem Cave, Israel (420 to 200 ka). Science Advances, 5, 1–12.
Blumenschine, R. J., & Selvaggio, M. M. (1988). Percussion marks on bone surfaces as a new diagnostic of hominid behavior. Nature, 333, 763–765.
Brown, W. A. B., & Chapman, G. N. (1991). The dentition of red deer (Cervus elaphus): A scoring scheme to assess age from wear of the permanent molariform teeth. Journal of Zoology, 224, 519–536.
Buckley, M., Collins, M., Thomas-Oates, J., & Wilson, J. C. (2009). Species identification by analysis of bone collagen using matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry. Rapid Communications in Mass Spectrometry, 23, 3843–3854.
Bunn, H. T., & Kroll, E. M. (1986). Systematic butchery by Plio-Pleistocene hominids at Olduvai Gorge, Tanzania. Current Anthropology, 27, 123–149.
Bunn, H.T. (1983). Comparative analysis of modern bone assemblages from a San hunter gatherer camp in the Kalahari Desert, Botswana, and from spotted hyena den near Nairobi, Kenya. In Clutton-Brock, J. & Grigson, C. (Eds.), Animals and Archaeology. Vol. 1. Hunters and their prey. (International Series, 163, pp. 143–148). BAR Publishing
Cáceres, I., Bravo, P., Esteban, M., Expósito, I. & Saladié, P. (2002). Fresh and heated bones breakage. An experimental approach. In Rienzi, M. De, Pardo Alonso, M.V., Belinchón, M., Peñalver, E., Montoya, P. & Márquez-Aliaga, A. (eds.), Current topics on taphonomy and fossilization. (pp. 471–481), Ayuntamiento de Valencia
Carriol, R.-P., & Álvarez-Fernández, E. (2015). Balanomorphs from late Upper Pleistocene and Holocene caves in northern Spain, with a new genus and species, and their palaeoclimatological implications. Annales De Paléontologie, 101(1), 21–27.
Carvalho, M., Jones, E. L., Marín, A. B., Geiling, J. M., Straus, L. G., & González Morales, M. R. (2021). Initial and Lower Magdalenian large mammal faunas and human subsistence at El Mirón Cave (Cantabria, Spain). Journal of Paleolithic Archaeology, 4(15), 1–34.
Castaños, P. (1980). Macrofauna de la cueva de la Paloma. In Hoyos, M., Martínez Navarrete, I., Chapa, T., Castaños, P. & Sanchiz, F.B. (Eds.), La Cueva de La Paloma: Soto de Las Regueras (Asturias). (Excavaciones Arqueológicas en España, 116, pp. 65–100), Ministerio de Cultura. Dirección General de Bellas Artes, Archivos y Bibliotecas
Castaños, P. (2016). Estudio arqueozoológico de la macrofauna de los yacimientos del proyecto «Los tiempos de Altamira». In Lasheras Corruchaga, J.A. (Dir.), Proyecto de investigación los tiempos de Altamira. Actuaciones arqueológicas en las cuevas de Cualventi, El Linar y Las Aguas (Alfoz de Lloredo, Cantabria, España). (Monografías del Museo Nacional y Centro de Investigación de Altamira, 26, pp. 196–218), Ministerio de Educación, Cultura y Deporte
Castaños, P. & Castaños, J. (2014). Estudio y revisión de la fauna de la cueva de Altamira (Santillana del Mar, Cantabria). Sautuola, XIX, 27–64
Corchón, M.S. (2017a). El yacimiento de Las Caldas en el contexto del valle del Nalón (17.000 – 11.500 cal BP). In Corchón, M.S. (Ed.), La cueva de Las Caldas (Priorio, Oviedo): ocupaciones magdalenienses en el valle del Nalón. (Estudios Históricos & Geográficos, 164, pp. 21–34), Ediciones Universidad de Salamanca
Corchón, M.S. (2017b), La cueva de Las Caldas (Priorio, Oviedo): ocupaciones magdalenienses en el valle del Nalón. (Estudios Históricos & Geográficos, 164), Ediciones Universidad de Salamanca
Costamagno, S. (1999). Stratégies de chasse et fonction des sites au Magdalénien dans le sud de la France. Université Bordeaux I.
Costamagno, S. (2002). Laboratory taphonomy—material loss and skeletal part profiles: The example of Saint–Germain–la–Rivière (Gironde, France). Archaeometry, 44(3), 495–504.
Costamagno, S., & David, F. (2009). Comparaison des practiques bouchères et culinaires de différents groupes sibériens vivant de la renniculture. Archaeofauna, 18, 9–25.
Costamagno, S., & Rigaud, J.-P. (2014). L’exploitation de la graisse au Paléolithique. In S. Costamagno (Ed.), Histoire de l’alimentation humaine: Entre choix et contraintes (pp. 134–152). CTHS.
Costamagno, S., Soulier, M.-C., Val, A., & Chong, S. (2019). The Reference Collection of Cutmarks. Palethnologie, 10, 186–280.
Costamagno, S. Théry-Parisot, I., Brugal, J-P. & Guilbert, R. (2005). Taphonomic consequences of the use of bone as fuel. Experimental data and archaeological applications. In O’Connor, T. (Ed.), Biosphere to Lithosphere: New studies in vertebrate taphonomy. Proceedings of the 9th ICAZ Conferences (Durham, August 2002). (pp. 51–62.), Oxbow Books
De Jong, Y., Verbeek, M., Michelsen, V., De Place Bjørn, P., Los, W., Steeman, F., Bailly, N., Basire, C., Chylarecki, P., Stloukal, E., Hagedorn, G., Wetzel, F. T., Glöckler, F., Kroupa, A., Korb, G., Hoffmann, A., Häuser, C., Kohlbecker, A., Müller, A., … Penev, L. (2014). Fauna Europaea - all European animal species on the web. Biodiversity Data Journal, 2, e4034. https://doi.org/10.3897/BDJ.2.e4034
Discamps, E., & Costamagno, S. (2015). Improving mortality profile analysis in zooarchaeology: A revised zoning for ternary diagrams. Journal of Archaeological Science, 58, 62–76.
Eastham, A. (1984). The Avifauna of the cave of Ekain. In Altuna, J. & Merino, J.M. (Eds.), El yacimiento prehistórico de la cueva de Ekain (Deba, Guipúzcoa). (pp.331–345), Eusko Ikaskuntza
Eisenmann, V., Alberdi, M.T., De Giuli, C. & Staeche, U. (1988). Studying fossil horses. Volume I: Methodology. Brill
Elorza, M. (2014). Explotación de aves marinas en el Tardiglacial del Golfo de Bizkaia: Las aves de Santa Catalina. Kobie (excavaciones Arqueológicas En Bizkaia), 4, 263–296.
Faith, J. T., & Gordon, A. D. (2007). Skeletal element abundances in archaeofaunal assemblages: Economic utility, sample size, and assessment of carcass transport strategies. Journal of Archaeological Science, 34, 872–882.
Fernández López, S., (1998). Tafonomía y fosilización. In Meléndez, B. (Ed.), Tratado de Paleontología. Tomo I. (Colección Textos Universitarios 29, pp. 51–107). CSIC
Fernández-Jalvo, Y. & Andrews, P. (2016). Atlas of taphonomic identifications. 1001+ images of fossil and recent mammal bone modification. (Vertebrate Paleobiology and Paleoanthropology Series), Springer
Fretter, V., & Graham, A. (1962). British prosobranch molluscs; Their functional anatomy and ecology. Royal Society.
Froese, R. & Pauly, D. (2024). FishBase. World Wide Web electronic publication. www.fishbase.org. Accessed 2 Dec 2024.
Garcia Petit, L. (2016). Aportación de las aves al conocimiento del entorno de la cueva de Altamira. In Lasheras Corruchaga, J.A. (dir.), Proyecto de investigación los tiempos de Altamira. Actuaciones arqueológicas en las cuevas de Cualventi, El Linar y Las Aguas (Alfoz de Lloredo, Cantabria, España). (Monografías del Museo Nacional y Centro de Investigación de Altamira, 26, pp. 320–340), Ministerio de Educación, Cultura y Deporte
Garcia-Ibaibarriaga, N., Suarez-Bilbao, A., Iriarte-Chiapuso, M. J., Arrizabalaga, A., & Murelaga, X. (2019). Palaeoenvironmental dynamics in the Cantabrian Region during Greenland stadial 2 approached through pollen and micromammal records: State of the art. Quaternary International, 506, 14–24.
González Echegaray, J. (1987). Morfología. Contenidos y aspectos genéticos de la “Cueva del Juyo”. In Barandiarán, I., Freeman, L.G., González-Echegaray, J. & Klein, R.G. (Eds.), Excavaciones arqueológicas en la cueva del Juyo. (Centro de Investigaciones y Museo de Altamira, 14, pp. 8–14), Ministerio de Cultura. Dirección General de Bellas Artes, Archivos y Bibliotecas
Grayson, D. K. (1984). Quantitative zooarchaeology: Topics in the analysis of archaeological faunas. Academic Press.
Grayson, D. K. (1989). Bone transport bone destruction, and reverse utility curves. Journal of Archaeological Science, 16, 643–652.
Grigson, C. (1982). Sex and age determination of some bones and teeth of domestic cattle: A review of the literature. In Wilson, B., Grigson, C. & Payne, S. (Eds.), Ageing and Sexing Animal Bones from Archaeological Sites. (British Series, 109, pp. 7–24), BAR Publishing
Guillaud, E., Bearez, P., Daujeard, C., Defleur, A. R., Desclaux, E., Rosello-Izquierdo, E., Morales-Muniz, A., & Moncel, M.-H. (2021). Neanderthal foraging in freshwater ecosystems: A reappraisal of the Middle Paleolithic archaeological fish record from continental Western Europe. Quaternary Science Review, 252, 10631.
Haynes, G. (1980). Evidence of carnivore gnawing on Pleistocene and recent mammalian bones. Paleobiology, 6, 341–351.
Heaton, T. J., Köhler, P., Butzin, M., Bard, E., Reimer, R. W., Austin, W. E. N., Bronk Ramsey, C., et al. (2020). Marine20—The Marine Radiocarbon Age Calibration Curve (0-55,000 cal BP). Radiocarbon, 62(4), 779–820.
Hillson, S. (2005). Teeth. Cambridge Manuals. in Archaeology.
Hopkins, R. J. A., Straus, L. G., & González Morales, M. R. (2021). Assessing the chronostratigraphy of El Mirón cave. Cantabrian Spain. Radiocarbon, 63(3), 1–32.
Hoyo, J. del, Elliott, A., Christie, D.A. & Sargatal, J.E. (1992). Handbook of the Birds of the World. (15 Vols), Lynx Edicions
Jin, J. J. H., & Mills, E. W. (2011). Split phalanges from archaeological sites: Evidence of nutritional stress? Journal of Archaeological Science, 38, 1798–1809.
Jones, K. T., & Metcalfe, D. (1988). Bare bones archaeology: Bone marrow índices and efficiency. Journal of Archaeological Science, 15, 415–423.
Jordá Pardo, J. F., Carral, P., Álvarez-Alonso, D., Arias, P., Bécares, J., Cubas, M., Martín-Jarque, S., Portero, R., Teira, L. C., & Álvarez-Fernández, E. (2018). Al oeste del Sella. Geoarqueología y cronoestratigrafía del registro del Pleistoceno Superior de la cueva de El Cierro (Fresno, Ribadesella, Asturias, España). Boletín Geológico y Minero, 129(1–2), 207–250.
Jordá Pardo, J.F., Carral, P., Maestro, A., Álvarez-Alonso, D., Arias, P., Bécares, J., Cubas, M., Martín-Jarque, S., Portero, R., Teira, L.C. & Álvarez-Fernández, E. (2018b). La gran transgresión. La secuencia pleistocena-holocena de la cueva de El Cierro (Fresnu, Ribadesella, Asturias): geoarqueología, cronoestratigrafía y paleogeografía. In García-Ibaibarriaga, N., Murelaga, X, Suárez, A. & Suarez, O. (Eds.), Paleoambiente y recursos bióticos del Pleistoceno superior cantábrico. Estado de la cuestión a la luz de las nuevas investigaciones. (Kobie, Serie Anejo, 18, pp. 57–68), Bizkaiko Foru Aldundia
Klein, R.G. & Cruz-Uribe, K. (1987). La fauna mamífera del yacimiento de la cueva de "el Juyo". Campañas de 1978 y 1979. In Barandiarán, I., Freeman, L.G., González-Echegaray, J. & Klein, R.G. (Eds.), Excavaciones arqueológicas en la cueva del Juyo. (Centro de Investigaciones y Museo de Altamira, 14, pp. 99–120), Ministerio de Cultura. Dirección General de Bellas Artes, Archivos y Bibliotecas
Kraft, E., 1972. Vergleichend morphologische Untersuchungen an Einzelknochen Nordund Mitteleuropäischer Kleinerer Hühnervögel. Universität München
Kuntz, D., Costamagno, S., Feyfant, L., & Martin, F. (2016). The exploitation of ungulates in the Magdalenian in the Entre-Deux- Mers (Gironde, France). Quaternary International, 414, 135–158.
Laukner, G. (1980). Diseases of Mollusca: Gastropoda. In O. Kinne (Ed.), Diseases of Marine Annimals (Vol. I, pp. 311–424). John Wiley & Sons.
Lecuyer, C., Hillaire-Marcel, C., Burke, A., Julien, M.-A., & Helie, J.-F. (2021). Temperature and precipitation regime in LGM human refugia of southwestern Europe inferred from d13C and d18O of large mammal remains. Quaternary Science Reviews, 255, 1–21.
Le Gall, O. (1984). L’ichtyofaune d’eau douce dans les sites préhistoriques. (Cahiers du Quaternaire ,8), CNRS
Levine, M.A. (1982). The use of crown height measurements and eruption-wear sequences to age horse teeth. In Wilson, B., Grigson, C. & Payne, S. (Eds.), Ageing and Sexing Animal Bones from Archaeological Sites. (British Series, 109, pp. 223–250), BAR Publishing
Llorente, L. & Morales, A. (2016). Las aves de la zona interior de la cueva de Aitzbitarte III. In Altuna, J., Mariezkurrena, K., Ríos, J. & San Emeterio, A. (Eds,), Ocupaciones humanas en Aitzbitarte III (País Vasco) 26.000 – 13.000 Bp (zona profunda de la cueva). (EKOB 8, pp. 317–330), Gobierno Vasco
López Plana, C., Mayor, P., Navarro, M., Carretero, A., Nacher, V., Aige, V., López, M., & Ruberte, J. (2011). Atlas de Osteología de los Mamíferos Domésticos. Universitat Autònoma de Barcelona.
López Quintana, J.C. (2011). La cueva de Santimamine: revision y actualización (2004–2006). (Kobie, Serie BAI, 1), Bizkaiko Foru Aldundia
López Quintana, J.C. & Guenga, A. (2011). Revisión estratigráfica del depósito arqueológico de la cueva de Santimamine (Kortezubi, Bizkaia): campanas de 2004 a 2006. cronoestratigrafía y paleoambiente. In López Quintana, J.C. (Dir.), La cueva de Santimamine: revision y actualización (2004–2006). (Kobie, Serie BAI, 1, pp. 7–70), Bizkaiko Foru Aldundia
López-Cisneros, P., Yravedra, J., Álvarez-Alonso, D., & Linares-Matás, G. (2019). The exploitation of hunted resources during the Magdalenian in the Cantabrian region. Systematization of butchery processes at Coímbre cave (Asturias, Spain). Quaternary International, 506, 46–58.
López-Cisneros, P. (2020). Patrones culturales en los procesos de carnicería sobre los macrovertebrados en el Paleolítico superior cantábrico. (Unpublished PhD), UNED
Lyman, R. L. (1994). Vertebrate taphonomy. Cambridge Manuals. in Archaeology.
Lyman, R. L. (2019). A critical review of four efforts to resurrect MNI in zooarchaeology. Journal of Archaeological Method and Theory, 26, 52–87.
Madariaga, B. (1975a). Representación malacológica de la Cueva de Tito Bustillo o de El Ramu (Oviedo). In M. A. García Guinea (Ed.), Primeros Sondeos Estratigráficos en la Cueva de Tito Bustillo (Ribadesella, Asturias) (Excavaciones de 1970) (pp. 71–74). Publicaciones del Patronato de las Cuevas Prehistóricas de la Provincia de Santander XII.
Madariaga, B. (1975b). Estudio de la fauna marina de la Cueva de Tito Bustillo (Oviedo). In Moure, J.A. (Ed.), Excavaciones en la Cueva de ‘‘Tito Bustillo’’ (Ribadesella, Asturias): Campañas de 1972 y 1974. (pp. 89–108). Instituto de Estudios Asturianos
Madariaga, B. & Fernández, P. (1987). Estudio malacológico de la cueva de El Juyo. In Barandiarán, I., Freeman, L.G., González-Echegaray, J. & Klein, R.G. (Eds.), Excavaciones arqueológicas en la cueva del Juyo. (Centro de Investigaciones y Museo de Altamira, 14, pp. 75–96), Ministerio de Cultura. Dirección General de Bellas Artes, Archivos y Bibliotecas
Madrigal, T. C., & Holt, J. Z. (2002). White-tailed deer meat & marrow return rates and their application to Eastern Woodlands archaeology. American Antiquity, 67(4), 745–759.
Marean, C. W., & Spencer, L. M. (1991). Impact of carnivore ravaging on zooarchaeology measures of element abundance. American Antiquity, 56, 645–658.
Mariezkurrena, K. (1983). Contribución al conocimiento del desarrollo de la dentición y el desarrollo del esqueleto postcraneal de Cervus elaphus. Munibe, 35, 149–202.
Mariezkurrena, K., & Altuna, J. (1983). Biometría y dimorfismo sexual en el esqueleto de Cervus elaphus würmiense, postwürmiense y actual del Cantábrico. Munibe, 35, 203–246.
Martínez-Villa, A., Álvarez-Fernández, E., Álvarez-Vena, A., Arrojo, L., Ballesteros, D., Cubas, M., Drak, L., Llorente Rodríguez, L., Martín-Jarque, S., & Gil, M. (2022). New insights into upper Palaeolithic and Mesolithic occupations in Les Pedroses cave (Asturies, north Spain). Journal of Archaeological Science: Reports, 45, 103592.
Masset, C., Costamagno, S., Cochard, D., & Laroulandie, V. (2016). La fracturation osseuse: Du fait technique à l’essai d’interprétation sociétale: l’exemple de l’antilope saïga du gisement magdalénien de Saint-Germainla- Rivière (Gironde). Bulletin De La Société Préhistorique Française, 113(4), 691–712.
Mateos, A. (1999). El consumo de grasa en el Paleolítico Superior. Implicaciones paleoeconómicas: Nutrición y subsistencia. Espacio, Tiempo y Forma, Serie i, Prehistoria y Arqueología, 12, 169–181.
Mateos, A. (2003). Estudio de la fragmentación de falanges y mandíbulas en la secuencia temporal del 19000–13000 BP de la cueva de las Caldas (Priorio, Oviedo) Implicaciones Paleoeconómicas: Nutrición y Subsistencia. Gallaecia, 22, 9–20.
McKynley, J. I. (2008). In the heat of the pyre: Efficiency of oxidation in Romano-British cremations – did it really matter? In C. A. Schmidt & S. Symes (Eds.), Beyond recognition: The analysis of burned human remains (pp. 163–184). Levier.
McKynley, J. I. (2016). Complexities of the ancient mortuary rite of cremation: An osteoarchaeological conundrum. In G. Grupe & G. McGlynn (Eds.), Isotopic Landscapes in Bioarchaeology (pp. 17–41). Springer.
Mech, L. D., Frenzel, L. D., Karns, P. D., & Kuehen, D. W. (1970). Mandibular dental anomalies in white tailed deer from Minnesota. Journal of Mammalogy, 51, 804–806.
Merkle, J. A., Cherry, S. G., & Fortin, D. (2015). Bison distribution under conflicting foraging strategies: Site fidelity vs energy maximization. Ecology, 96(7), 1793–1801.
Metcalfe, D., & Jones, K. T. (1988). A reconsideration of animal body-part utility indices. American Antiquity, 53, 486–504.
Monge Soares, A. M., Gutiérrez-Zugasti, I., González-Morales, M., Matos Martins, J. M., Cuenca Solana, D., & Bailey, G. N. (2016). Marine radiocarbon reservoir effect in Late Pleistocene and Early Holocene coastal waters off Northern Iberia. Radiocarbon, 58(4), 869–883.
Morales, A. (1984). Primer informe sobre la ictiofauna magdalaniense de la cueva de Tito Bustillo ( provincia de Asturias). Boletín del Instituto de Estudios Asturianos, 113, 904–929.
Moreno, R. (1994). Análisis arqueomalacológicos en la Península Ibérica. Contribución metodológica y biocultural. Universidad Autónoma de Madrid.
Morin, E. (2007). Fat composition and Nunamiut decision-making: A new look at the marrow and bone grease indices. Journal of Archaeological Science, 34, 69–82.
Nicholson, R. (1993). A morphological investigation of burnt animal bone and an evaluation of its utility in archaeology. Journal of Archaeological Science, 20, 411–428.
Noddle, B. (1974). Ages of epiphyseal closure in feral and domestic goats and ages of dental eruption. Journal of Archaeological Science, 1, 195–204.
O’Connor, R. J., & Lamont, P. (1978). The spatial organization of an intertidal Spirorbis community. Journal of Experimental Marine Biology and Ecology, 32, 143–169.
Ortea, J. (1986). The Malacology of La Riera Cave. In Straus, L. G. & Clark, G. (Eds), La Riera Cave. Stone Age Hunter-Gatherer adaptations in Northern Spain. (Anthropological Research Papers, 36, pp. 289–298), Arizona State University
Palacios, N. & Vega, J.J. (1997). Guía de conchas de las playas y rías de Cantabria. Consejería de Medio Ambiente de la Diputación Regional de Cantabria
Pales, L. & Garcia, M.A. (1981). Atlas ostéologique pour servir à l’identification des mammifères du Quaternaire. Tête, rachis, ceintures scapulaire et pelvienne, membres: Herbivores. Editions du CNRS
Pereda, S. & Doadrio I. (2016). Aproximacion al estudio paleosistematico de la ictiofauna de las cuevas de Cualventi, El Linar y Las Aguas. In Lasheras Corruchaga, J.A. (Dir.), Proyecto de investigación los tiempos de Altamira. Actuaciones arqueológicas en las cuevas de Cualventi, El Linar y Las Aguas (Alfoz de Lloredo, Cantabria, España). (Monografías del Museo Nacional y Centro de Investigación de Altamira, 26, pp. 373–409), Ministerio de Educación, Cultura y Deporte
Pérez Ripoll, M. (1988). Estudio de la secuencia del desgaste de los molares de Capra pyrenaica de los yacimientos prehistóricos. Archivo De Prehistoria Levantina, 18, 83–128.
Pickering, T. R., & Egeland, C. P. (2006). Experimental patterns of hammerstone percussion damage on bones: Implications for inferences of carcass processing by humans. Journal of Archaeological Science, 33, 459–469.
Portero, R. (2022). Estrategias de subsistencia a finales del Pleistoceno superior y comienzos del Holoceno en el Valle del Sella: La Cueva de El Cierro (Ribadesella, Asturias, España). (Unpublished PhD) Universidad de Salamanca
Portero, R., Cueto, M., Jordá Pardo, J. F., Bécares, J., & Álvarez-Fernández, E. (2019). The persistence of red deer (Cervus elaphus) in the human diet during the Lower Magdalenian in northern Spain: Insights from El Cierro cave (Asturias, Spain). Quaternary International, 506, 35–45.
Portero, R., Fernández-Gómez, M. J. & Álvarez-Fernández, E. (2024). Revisiting macromammal exploitation in the Spanish Cantabrian region during the lower Magdalenian (ca. 20-17 ky cal BP). Quaternary Science Reviews, 332, 108651.
Power, R. C., Salazar-García, D. C., Straus, L. G., González Morales, M. R., & Henry, A. G. (2015). Microremains from El Miron Cave human dental calculus suggest a mixed plant-animal subsistence economy during the Magdalenian in Northern Iberia. Journal of Archaeological Science, 60, 39–46.
Rasines del Río, P. (2016). Cronología de los yacimientos arqueológicos de Altamira, Cualventi, El Linar y Las Aguas (Cantabria). In Lasheras Corruchaga, J.A. (Dir.), Proyecto de investigación los tiempos de Altamira. Actuaciones arqueológicas en las cuevas de Cualventi, El Linar y Las Aguas (Alfoz de Lloredo, Cantabria, España). (Monografías del Museo Nacional y Centro de Investigación de Altamira, 26, pp. 132–147), Ministerio de Educación, Cultura y Deporte
Reimer, P., Austin, W., Bard, E., Bayliss, A., Blackwell, P., Bronk Ramsey, C. J., Butzin, M., et al. (2020). The IntCa20 Northern Hemisphere Radiocarbon Age Calibration Curve (0-55 cal kBP). Radiocarbon, 62(4),725–757.
Reitz, E. J., & Wing, E. S. (2003). Zooarchaeology. Cambridge Manuals. in Archaeology.
Roselló, E. & Morales, A. (2017). La ictiofauna solutreo-magdaleniense de la cueva de Las Caldas (Asturias). In Corcchón, M.S. (Ed.), La cueva de Las Caldas (Priorio, Oviedo): ocupaciones solutrenses, análisis espaciales y arte parietal. (Estudios Históricos & Geográficos, 166, pp. 229–236), Ediciones Universidad de Salamanca
Rufà, A., Blasco, R., Menschel, M., & Pokines, J. T. (2022). The avian remains from El Juyo, Lower Magdalenian Cantabrian Spain. Journal of Archaeological Science: Reports, 46, 103713.
Saladié, P., Huguet, R., Díez, C., Rodríguez-Hidalgo, A., Cáceres, C., Vallverdú, J., Rosell, J., Bermúdez de Castro, J. M., & Carbonell, E. (2011). Carcass transport decisions in Homo antecessor subsistence strategies. Journal of Human Evolution, 61(4), 425–446.
Santhanam, R. (2018). Biology and ecology of edible marine gastropod molluscs. Apple Academic Press Inc.
Schmid, E. (1972). Atlas of animal bones for prehistorians, archaeologist and quaternary geologist. Elsevier Publishing Company.
Selvaggio, M. M. (1994). Carnivore tooth marks and stone tool butchery marks on scavenged bones: Archaeological implications. Journal of Human Evolution, 27, 215–228.
Shipman, P., & Rose, J. J. (1983). Early hominid hunting, butchering, and carcass-processing behaviors: Approaches to the fossil record. Journal of Anthropological Archaeology, 2(1), 57–98.
Southward, A.J., (2008). Barnacles. Synopses of the British Fauna (New Series) 57. The Linnean Society of London and The Estuarine and Coastal Sciences Association by Field Studies Council
Steele, T. E. (2002). Red deer: Their ecology and how they were hunted by Late Pleistocene Hominids in Western Europe. Stanford University.
Stiner, M. C. (1990). The use of mortality patterns in archaeological studies of hominid predatory adaptations. Journal of Anthropological Archaeology, 9, 305–351.
Stiner, M. C. (1994). Honor among thieves: A zooarchaeological study of Neandertal ecology. Princeton University Press.
Stiner, M.C., Beaver, J.E., Munro, N.D., & Surovell, T.A. (2008). Modeling paleolithic predator-prey dynamics and the effects of hunting pressure on prey choice. In Bocquet-Appel, J.-P. (Ed.), Recent Advances in Palaeodemography. Data, techniques, patterns. (pp.143–178). Springer
Stiner, M. C., Kuhn, S. L., Weiner, S., & Bar-Yosef, O. (1995). Differential burning, recrystallization, and fragmentation of archaeological bone. Journal of Archaeological Science, 22, 223–237.
Stiner, M. C., Munro, N. D., Surovell, T. A., Tchernov, E., & Bar-Yosef, O. (1999). Paleolithic population growth pulses evidenced by small animal exploitation. Science, 283, 190–194.
Tomé, C., & Vigne, J. D. (2003). Roe deer (capreolus capreolus) age at death estimates: New methods and modern reference data for tooth eruption and ear and ford epiphysial fusion. Arqueofauna, 12, 157–173.
Trigo, J. E., Rolán, E., Díaz Agras, G. J., García-Álvarez, O., Guerra Sierra, A., Moreira da Rocha, J., Pérez Dieste, J., Souza Troncoso, J., & Urgorri, V. (2018). Guía de los moluscos marinos de Galicia. Universidade de Vigo.
Uzquiano, P. (2019). Vegetal landscape and firewood supply strategies in N Spain at the Greenland Stadial 2. Quaternary International, 506, 6–13.
Varela, S., & Rodríguez, J. (2004). Atlas osteológico. Carnívoros ibéricos. CSIC.
Venkataraman, V. V., Kraftb, T. S., Dominyc, N. J., & Endicott, K. M. (2017). Hunter-gatherer residential mobility and the marginal value of rainforest patches. PNAS, 114(12), 3097–3102.
Vettese, D., Blasco, R., Cáceres, I., Gaudzinski-Windheuser, S., Moncel, M.-H., Thun Hohenstein, U., & Daujeard, C. (2020). Towards an understanding of hominin marrow extraction strategies: A proposal for a percussion mark terminology. Archaeological and Anthropological Science, 12(48), 1–19.
Villa, P., & Mahieu, E. (1991). Breakage patterns of human long bones. Journal of Human Evolution, 20, 1–22.
Wegrzyn, M., & Serwatka, S. (1984). Teeth eruption in the European Bison. Acta Theriologica, 29(9), 111–121.
WoRMS Editorial Board (2024). World register of marine species. Available from. www.marinespecies.org de VLIZ. Accessed 24–02–01.
Yravedra, J. (2006). Tafonomía aplicada a la zooarqueología. UNED.
Yravedra, J., & Uzquiano, P. (2013). Burnt bone assemblages from El Esquilleu cave (Cantabria, Northern Spain): Deliberate use for fuel or systematic disposal of organic waste? Quaternary Science Reviews, 68, 175–190.
Yravedra, J., Baena, J., Arrizabalaga, A. & Iriarte, M.J. (2005). El empleo de material óseo como combustible durante el Paleolítico Medio y Superior en el Cantábrico. Observaciones experimentales. In Montes, R. & Lasheras, J.A. (Coords), Actas de la Reunión Científica: Neandertales Cantábricos. Estado de la Cuestión. (Monografías del Museo Nacional y Centro de Investigación de Altamira, 20, pp. 369–383), Ministerio de Educación, Cultura y Deporte
Zurro, D. (2010). Ni carne ni pescado (consumo de recursos vegetales en la Prehistoria): Análisis de la variabilidad de los conjuntos fitolitológicos en contextos cazadores-recolectores. (Unpublished PhD) Universitat Autònoma de Barcelona
Acknowledgements
This research was made possible thanks to a Margarita Salas postdoctoral fellowship awarded to Rodrigo Portero by the University of Salamanca and the Spanish Ministry of Science (Plan de Recuperación, Transformación y Resiliencia) with EU-Next-Generation funds. The authors are very grateful to Jean-Marc Pétillon for sampling and Krista McGrath for the analysis of ZooMS performed in the framework of the project PaleoCet. Thanks to Peter Smith for the translation of the manuscript. We thank the reviewers for their careful reading of the manuscript.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This work was undertaken in the context of the PaleontheMove Project PID2020.114462 GB-I00 (PI: Esteban Álvarez Fernández) and CantabricOIS2 PID2020.115192 GB-I00 (PI: Jesús F. Jordá Pardo) funded by the Programa Estatal de Fomento de Generación de Conocimiento y Fortalecimiento Científico y Tecnológico of the Spanish Ministry of Science and Innovation. ZooMS analysis was funded by the PaleoCet project ANR grant 18-CE27-0018–01 (PI: J.-M. Pétillon).
Author information
Authors and Affiliations
Contributions
R.P.: Conceptualization, Formal analysis, Data curation, prepared figures and tables, Writing – original draft, Writing – review & editing. M.C.: Formal analysis, Data curation, prepared figures, Writing – review. M.E.: Formal analysis, Data curation, Writing – review. A.M.: Formal analysis, Writing – review. J.J.P.: Formal analysis, Data curation, Writing – review. E.A-F.: Formal analysis, Data curation,Writing – review & editing.
Corresponding author
Ethics declarations
Ethical Approval
Not applicable.
Consent to Participate
All authors consent to participate.
Consent for Publication
All authors consent to publish.
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Portero, R., Cueto, M., Elorza, M. et al. Subsistence Strategies in the Lower Magdalenian at El Cierro Cave (Ribadesella, Asturias, Spain). J Paleo Arch 7, 14 (2024). https://doi.org/10.1007/s41982-024-00179-x
Accepted:
Published:
DOI: https://doi.org/10.1007/s41982-024-00179-x