
Abstract
A stratigraphic study of the carbonate deposits from the interval between the uppermost Sinemurian and the base of the Upper Pliensbachian is presented herein. For this purpose, four expanded sections of the Asturian Basin, in Northern Spain, were sampled. The sections are located between Rodiles E section (Villaviciosa municipality) and Playa de Vega section (Ribadesella municipality). The ammonites recorded in over 100 successive levels of this sector enabled the substantial improvement of the biochronostratigraphic scale of reference; making it comparable with those defined for other basins of the Iberian Peninsula and other areas of the western Tethys. The lower boundary of the Pliensbachian is indicated by the first appearance of Gemmellaroceras aff. aenigmaticum (Gemmellaro) below the first appearance of the genus Apoderoceras. The upper boundary is defined by the first record of the Protogrammoceras. (Matteiceras) subgenus which in some levels is associated with Amaltheus stokesi (Sowerby). The boundaries between the zones and subzones were able to be established accurately in most cases. The ammonite assemblages are similar to those established for the NW European Province. Some intervals, however, include species typical of the Mediterranean Province. Among these, the Taylori Subchronozone (Gemmellaroceras) and the transition between the Jamesoni Subchronozone and the Valdani Subchronozone (Tropidoceras) can be highlighted. Additionally, taking into consideration the succession between of ammonite genera and species, 27 horizons were identified, 24 of which correspond to the Lower Pliensbachian, and the other 3 to the uppermost Sinemurian and the lowermost part of the Upper Pliensbachian. Moreover, the identification of 3 of the Lower Pliensbachian horizons were based upon the evolution of the Mediterranean Tropidoceras.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
The Lower Pliensbachian biochronostratigrapy has been established in many sections of Europe i.e. Dean et al. (1961), Mouterde (1967), Mouterde et al. (1983, 2007), Gèczy (1976), Oates (1978), Comas-Rengifo (1982), Braga (1982), Braga et al. (1984, 1985), Phelps (1985), Meister (1986), Dommergues (1987), Dommergues et al. (1997), Blau & Meister (2000), Page (2003), Meister & Friebe (2003), Meister et al. (2006, 2009, 2017) and Fauré & Bohain (2017). However, no studies at high resolution, at chronohorizon or zonula scale, have been performed in Iberia.
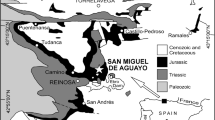
The objective of the present study is to establish the highest biochronostratigraphic resolution of the Lower Pliensbachian succession in Northern Spain. For this purpose, a detailed study and sampling of four sections of the Asturian Basin (AB) were completed, (Fig. 1), the Rodiles E (ER) was taken as a reference section where the Lower Pliensbachian deposits are 42.8 m thick. The obtained data were completed with information from other sections, such as those located in Punta La Llastra (LL), Santa Mera (SM) and Playa de Vega (PV).

Geological map of central-eastern area of Asturias (N Spain) modified from García-Ramos & Gutiérrez Claverol (1995) and location of different study sections: Rodiles E, Punta La Llastra and Santa Mera (Villaviciosa), Huerres (Colunga), Playa de Vega (Ribadesella) and Vega de Sariego (Sariego).
Analysis of the ammonite succession of the Lower Pliensbachian on the central-western coast of Asturias (North Spain) enabled us to achieve a detailed biostratigraphic and chronostratigraphic study. For this purpose, based on these sections and some local data (i.e., Huerres; Fig. 1), we established the stratigraphic distribution of the identified species, in an attempt to improve the biochronostratigraphic scale of reference obtained by Suárez Vega (1974). A complementary objective involves the correlation of the Asturian Basin (AB) with other basins of the Iberian Peninsula, such as the Basque-Cantabrian Basin (BC), the Pyrenean Basin (PB), the Iberian Basin (IB), the Betic Basin (BB) and the Portuguese Lusitanian Basin (LB), between the uppermost Sinemurian and the base of the Upper Pliensbachian. Other goals of interest are to establish the lower boundary of the Pliensbachian stage between Villaviciosa and Ribadesella, and to determine, with the highest resolution, the boundary between the chronozones and subchronozones of ammonites from the Lower Pliensbachian in an attempt to contribute to a better understanding of the palaeogeography during the Pliensbachian of the Iberian Peninsula.
Previous studies on the biochronostratigraphy of the Pliensbachian in the Asturian Basin were carried out in the second half of the XIX century and at the beginning of the XX century (i.e. Schulz, 1858; Barrois, 1882; Mallada, 1885; Jiménez de Cisneros, 1904 among others, see Suárez Vega, 1974. Lately, the biostratigraphic distribution of the ammonites and the chronostratigraphy of the Pliensbachian in Asturias were studied by Dubar (1925), Dubar & Mouterde (1957) and Dubar et al. (1972), who studied the Lower Jurassic between Ribadesella and Gijón. These authors indicate that the Lower Pliensbachian shows an increase in thickness (15 to 40 m) between Ribadesella (PV section) and the Playa de Peñarrubia, East of Gijón.
Suárez Vega (1974) attributes a maximum of approximately 42.8 m to the Lower Pliensbachian, providing detailed biostratigraphic information. In the Lower Pliensbachian he characterises the Jamesoni Zone with the Taylori, Polymorphus-Brevispina and Jamesoni horizons, the Ibex Zone with the Maugenesti, Valdani-Ellipticum, Stahli and Beaniceras spp. horizons and the Davoei Zone with the Oistoceras spp. horizon in the upper part. He also characterises the Valdani Subzone of the Ibex Zone and the Maculatum Subzone of the Davoei Zone, recognised by different authors from France and England. Some years later Braga et al. (1984) conducted further studies of the ammonite successions, limits and correlations of the Pliensbachian in Spain, making use of the data provided by Suárez Vega to study different aspects of the Lower Pliensbachian in Asturias.
2 Materials and methods
All the sections were studied and sampled bed by bed, and the classified ammonites were collected by the authors of the present paper over the last fifteen years. In the studied sections, the relevant discontinuities identified with the available biochronostratigraphy, always encompass less than one subchronozone.
The specimens collected are from the interval between the Upper Sinemurian (top of the Raricostatum Chronozone) and the base of the Upper Pliensbachian (lowermost Margaritatus Chronozone). In the most complete succession (ER), ammonites from 96 successive levels were collected, 83 of which correspond to the Lower Pliensbachian and the remaining to the Sinemurian and Upper Pliensbachian.
In most cases, the infill of the shells is very similar to the matrix in which they were embedded, indicating thata they are accumulated, and only in some exceptional cases evidence indicating that the ammonites were re-elaborated can be found.
In the ER and PV sections, virtually all the layers were numbered (Fig. 2). In many cases the lowermost part (b) and the topmost part (t) were differentiated and when the thickness was relatively significant, three parts were distinguished: lower: (i), middle (m) and upper (s). In some cases the specimens were labelled with the corresponding level number, followed by the distance in centimetres from the base or top of the limestone layers to the point at which the sample was obtained.

a Rodiles E partial section with Jamesoni Chronozone (Polymorphus, Brevispina and Jamesoni subchronozones) and Ibex Chronozone (Masseanum and Valdani subchronozones). b Punta La Llastra partial section with Jamesoni Chronozone (Polymorphus, Brevispina and Jamesoni subchronozones) and Ibex Chronozone (Masseanum and Valdani subchronozones). c Santa Mera partial section with Jamesoni Chronozone (Polymorphus Subchronozone). d Playa de Vega partial section with Jamesoni Chronozone (Polymorphus, Brevispina and Jamesoni subchronozones), Ibex Chronozone (Masseanum, Valdani and Luridum subchronozones), Davoei Chronozone (Maculatum, Capricornus and Figulinum subchronozones) and Margaritatus Chronozone (Stokesi Subchronozone).
In the SM and LL sections, given the particular features of the outcrops, easily-identifiable limestone layers were established, and the label of each specimen indicated the distance in cm below the base or above the top followed by the reference number of the layer, for example, SM (-15) or LL (+ 10) respectively.
The collected ammonites have been completed with the review of the Lower Pliensbachian ammonites deposited in the Jurassic Museum of Asturias (MUJA), under the title “Suárez Vega Collection”, corresponding to the PhD of this author, along with some specimens of the “Mouterde Collection”, studied by Dubar & Mouterde (1957) and subsequently by Suárez Vega (1974). Other Spanish specimens from the “Mouterde Collection” are from the Catholic Faculties of Lyon, and are deposited in the Palaeontology Dept. of the Madrid’s Complutense University (DPUCM). Research has also been conducted on the specimens corresponding to the work of Comas-Rengifo et al. (2010), Comas-Rengifo & Goy (2010) and Gómez et al. (2016a, b), as well as the ones from Asturias studied by Rodríguez-Luengo et al. (2012).
3 Stratigraphic succession
3.1 Stratigraphy and sedimentology
The Lower Pliensbachian marine succession cropping out at the Asturian coast is characterized by rhythmic marl-limestone alternations included in the Santa Mera Member of the Rodiles Formation (Valenzuela et al., 1986). These coastal cliffs containing Jurassic rocks run from the localities of Gijón to Ribadesella (Fig. 1). This carbonate succession was deposited in an intrashelf basin displaying an irregular floor controlled by differential subsidence. From a palaeobiogeographic point of view, it represents an intermediate area between the Boreal and the Tethyan domains.
The whole succession likely accumulated in a relatively shallow water environment with a depth of up to 100 m, which most of the time was situated below the fair-weather wave base (Bandel & Knitter, 1986; Bjerrum et al., 2001; French et al., 2014; Leonowicz, 2015). The carbonate content of the limestone beds ranges from 65 to 97%, fluctuating from 8 to 65% in the marl interbeds (Bádenas et al., 2012). TOC values range from 0.75 to 7.32%, with the higher scores found in the laminated black shale intervals (Borrego et al., 1997, updated following biostratigraphic corrections; Gómez et al., 2016a, b; Deconinck et al., 2020).
Prevailing among the invertebrate macrofauna are molluscs such as ammonites, belemnites and bivalves (Fig. 3a and b), although brachiopods (Fig. 3c) and crinoids are also well represented. A characteristic interval of the Taylori Subchronozone black shales includes high concentrations of oil-infilled parautochthonous brachiopods deposited by storm currents and buried alive (Fig. 3c). During a late burial stage, the oil migration filled the porous interior of the empty or drusy calcite-draped specimens.

a Concentration by storms of Gryphaea specimens in the erosive base (bioclastic layer) of an elementary cycle, Jamesoni Chronozone, Huerres. b Inoceramid shell with several oyster encrusters, Jamesoni Chronozone (top of Jamesoni Subchronozone), Punta La Llastra. c Brachiopod shellbed, some of them with oil infill, Jamesoni Chronozone (Taylori Subchronozone), Santa Mera. d Infaunal bivalve resting (Lockeia, L) and locomotion traces (Ptychoplasma, Pt) associated to Chondrites(Ch) and Planolites (Pl), Jamesoni Chronozone, Punta La Llastra. e Internal mould of an ammonites traversed by Chondrites (Ch) in a marl interbed, Jamesoni Chronozone, Punta La Llastra. f Trace fossil association including Chondrites (Ch), Diplocraterion (D), Phymatoderma (Ph), Planolites (Pl) and Thalassinoides (T) in a marl interbed, Punta La Llastra. g Phymatoderma (Ph) specimens in a marl interbed, Jamesoni Chronozone (Taylori Subchronozone), Huerres. h Skeletal remains of the ichthyosaur Leptonectes sp. Jamesoni Chronozone (Taylori Subchronozone), Rodiles E.
Likewise, the trace fossils exhibit a relatively high diversity and belong to the Skolithos, Cruziana and Glossifungites ichnofacies. The most representative ichnogenera, (albeit appearing in varying proportions) are: Arenicolites, Asterosoma, Bergaueria, Chondrites (Fig. 3e and f), Conichnus, Diplocraterion (Fig. 3f), Gyrolithes, Kulindrichnus, Lockeia (Fig. 3d), Multina, Oravaichnium Palaeophycus, Phycosiphon, Phymatoderma (Fig. 3f and g), Planolites (Fig. 3f), Protovirgularia, Ptychoplasma (Fig. 3d), Rhizocorallium, Skolithos, Taenidium, Teichichnus, Thalassinoides (Fig. 3f) and Trichichnus (García-Ramos et al., 2011).
The vertebrate fauna consists of two partial ichthyosaur skeletons from the Rodiles E (Fernández et al., 2018; Fig. 3h) and Playa de Vega sections, and another incomplete plesiosaur specimen from the Santa Mera section (Bardet et al., 2008); they are all located in the black shale intervals of the Jamesoni Chronozone (Taylori Subchronozone).
3.1.1 Cyclical organization
In the studied rhythmic succession, as a general trend, an increase in both the limestone/marl ratio and the irregularity of the limestone bed surfaces (often by scouring or erosive amalgamation; Fig. 4a) represent shallowing episodes, and vice versa (García-Ramos, 2010; García-Ramos et al. 2011). The marl/limestone couplets are arranged according to the scale considered in two types of cycles: elementary, on one hand, and small transgressive-regressive on the other. The elementary cycles consist of centimetre- to decimetre-thick intervals including a basal bioclastic layer with an erosive base and a fining-upward trend, a middle layer of laminated or massive marl and an upper bioturbated layer (García-Ramos et al., 2011; Bádenas et al., 2012). These elementary cycles are found throughout the succession (Fig. 4b-d), but it is difficult to detect them in the Ibex Chronozone black shales. The small-scale transgressive-regressive cycles, with a thickness from several tens of centimetres to 1.5 m, include several (up to 6) elementary cycles. Cycle boundaries are characterised by condensed elementary cycles and frequent scouring events, erosive amalgamation processes and burrow mottling with complex tiering (time-averaging effect). These cycle boundaries cannot always be placed at a discrete bed surface and actually represent cycle-boundary zones associated with a general loss of accomodation. Moreover, these bounded zones often exhibit firmgrounds associated with Glossifungites ichnofacies and a relatively more diverse assemblages of trace fossils. Herein the limestone/marl ratio is also higher.

a Gryphaea concentrations in the base of a scoured limestone bed excavated by a storm current, Jamesoni Chronozone (Taylori Subchronozone), Santa Mera. b Two consecutive elementary cycles, Jamesoni Chronozone (Jamesoni Subchronozone), Playa de Vega. c Two consecutive elementary cycles, Jamesoni Chronozone (Jamesoni Subchronozone), Punta La Llastra. d The top of scale indicates the base of the bioclastic layer below the laminated layer in an incomplete, at the top, elementary cycle, Jamesoni Chronozone (Jamesoni Subchronozone), Punta La Llastra. e Erosive amalgamation of limestone beds interpreted as a sudden shallowing episode, Jamesoni Chronozone (Polymorphus Subchronozone), Playa de Vega. f Nodular and discontinuous limestone beds and grey marls including frequent phosphatic faunal fragments reworked by erosion, Davoei Chronozone (Maculatum Subchronozone), Playa de Vega. g Laminated black shale interval, Ibex Chronozone (Valdani Subchronozone), Playa de Vega. h Detail of g with crushed bivalve shell beds.
On the other hand, these Lower Pliensbachian successions present several sudden shallowing (forced regression) and deepening episodes with apparently sharp lower and upper boundaries. Two of the most representative of these correlatable episodes throughout the Asturian basin outcrops include erosive amalgamated limestone beds 41 to 45 (upper half of the Polymorphus Subchronozone) in the playa de Vega section (Fig. 4e), and beds 360 to 370 (lower half of the Taylori Subchronozone) in the Rodiles E section. Also characteristic of the Playa de Vega section is an interval displaying alternations of nodular or discontinuous limestone beds and grey marls, containing frequent phosphatic faunal fragments, included predominantly in the Maculatum Subchronozone (Davoei Chronozone). This reveals a condensation episode related to a shallowing event, as evidenced also by a relative increase in amalgamated bioclastic deposits (Fig. 4f).
Some of main transgressive surfaces occur around the Sinemurian-Pliensbachian boundary (top of bed 356, base of the Taylori Subchronozone, in the Rodiles E section), at the top of bed 7 (lower half of the Taylori Subchronozone) in the Santa Mera section, and at the top of bed 63 (near the base of the Valdani Subchronozone) in the Punta La Llastra section. The graphic representation and the possible origin of cyclicity are not within the scope of the present paper; this constitutes the objective of a forthcoming publication.
3.1.2 Diagenetic aspects
Rhythmic marl-limestone alternations are usually interpreted as a consequence of periodic oscillations in the orbital parameters of the Earth, even in greenhouse times.
In the present case, the diagenetic modifications are only partially controlled by the primary (sedimentary) processes. Consequently, several orders of cycles and their boundaries do not match exactly with the present lithologic changes between marl interlayers and limestone beds. Specifically, each marl interbed or limestone bed includes several elementary cycles (Fig. 4d-f).
On the other hand, the boundary between both lithologies crosscuts obliquely the time lines, thus merging diachronous deposits into one single limestone or marl layer, a phenomenon previously indicated by Nohl & Munnecke (2019) in the Silurian rhythmic series of Sweden (See also: Biernacka et al., 2005; Gani, 2017; Nohl et al., 2021).
Light to medium-grey massive and/or bioturbated limestone beds are only slightly compacted and generated during a shallow-burial environment (early diagenesis) (Westphal, 2006; Westphal et al., 2010). Within limestone beds, the macrofossils are most frequently undeformed due to early cementation, while in the marls, many of them are crushed with the exception of those possessing a thick calcitic shell. The greatest values of flattening were found in the internal moulds of ammonites preserved in episodes of dark marls and black shales. These differences result from carbonate redistribution, differential compaction and cementation processes between the limestone beds and the marl interbeds. The local presence in the marl interbeds of infilled internal moulds of ammonites traversed by Chondrites also suggest an early and shallow subsurfacial aragonite dissolution of the cephalopod shells. On the contrary, the dark grey laminated limestones (frequently marly limestones) occur exclusively within the black shale intervals and are generated after the main compaction phase (later diagenesis), as evidenced by the enclosed crushed both ammonites and the scarce trace fossils.
Another problem arising in the use of limestone-marl couplets for cyclostratigraphic analysis in our case involves the lateral wedging or bifurcation of many limestone beds and the apparently sharp boundaries between both calcareous lithologies controlled by present-day weathering processes in the outcrop. The primary (sedimentary) boundaries are very gradual, as evidenced previously by Ricken (1986), with the highest carbonate content usually located in the middle part of the limestone beds and vice versa in the marl interbeds. Moreover, the frequent incomplete tiering of trace fossils by erosive processes suggests multiple small gaps in the rhythmic succession below the present-day biostratigraphic resolution.
These and other additional diagenetic processes call into question the indiscriminate use of limestone-marl alternations for orbital-related climatic cyclicity and astrochronology. See also: Ricken (1986), Munnecke et al. (2001), Westphal (2006), Westphal et al. (2008, 2010), Bádenas et al. (2009, 2012), García-Ramos (2010), García-Ramos et al. (2010, 2011), Nohl & Munnecke (2019), Nohl et al. (2020), Su et al. (2020).
3.2 Chronostratigraphy
In order to establish the biochronostratigaphy of the Lower Pliensbachian, the ER section has been taken as a reference; because the whole interval considered shows a consistent ammonite record (Suárez Vega,1974; Comas-Rengifo et al., 2010; Gómez et al., 2016a, b). This section is the most complete and representative of the AB (Figs. 2a and 5) and the biochronostratigraphic intervals are commonly more expanded. Nonetheless, it was complemented with the information provided on certain intervals in other sections (Figs. 2b-d and 6), such as LL (Jamesoni Chronozone, Brevispina Subchronozone to Ibex Chronozone, Luridum Subchronozone), SM (Jamesoni Chronozone, Taylori Subchronozone to Jamesoni Subchronozone) and PV (Jamesoni Chronozone, Jamesoni Subchronozone to Margaritatus Chronozone, base of the Stokesi Subchronozone). As has been pointed out, this last section presents less thickness than the other three (Valenzuela et al., 1986; García-Ramos et al., 1992; Comas-Rengifo & Goy, 2010).

Lithological section of Rodiles E (ER) with the stratigraphical distributions of the identified species of ammonites (after Comas-Rengifo & Goy, 2010; Gómez et al., 2016a, b). Abbreviations: CZ Chronozone. SCZ Subchronozone. M Meters. LV Level. RAR Raricostatum Chronozone. AP Aplanatum Subchronozone. MA Masseanum Subchronozone. MC Maculatum Subchronozone. CA Capricornus Subchronozone. FI Figulinum Subchronozone.

Lithological section of LLastra (LL), Santa Mera (SM) (after García-Ramos, 2010; García-Ramos et al., 2011) and Playa de Vega (PV) (after Comas-Rengifo & Goy, 2010) sections. Abbreviations: Z Chronozone. SZ Subchronozone. M Meters. LV Level. PO Polymorphus Subchronozone. BR Brevispina Subchronozone. MA Masseanum Subchronozone. MC Maculatum Subchronozone. CA Capricornus Subchronozone. FI Figulinum Subchronozone.
For the zonal scales of reference, we compare the obtained results with those previously proposed for the NW European and Mediterranean Provinces. Among others, by Dean et al. (1961), Mouterde (1967), Gèczy (1976), Oates (1978), Comas-Rengifo (1982), Braga (1982), Mouterde et al. (1983, 2007), Braga et al. (1984, 1985), Phelps (1985), Meister (1986), Dommergues (1987), Dommergues et al. (1997), Blau et al. (2000), Page (2003), Meister & Friebe (2003), Meister et al. (2006, 2009, 2017), and Fauré & Bohain (2017).
Figure 7 shows the distribution of the identified species in relationship to the most recently used standard scales (Dommergues et al., 1997; Page, 2003). The succession of these species was used to characterise 26 ammonite horizons in the uppermost Sinemurian and the base of the Upper Pliensbachian interval.

Stratigraphic distribution of the identified taxa of ammonites. Abbreviations: Z Chronozone. SZ Subchronozone. HZ Horizon. RAR Raricostatum Chronozone. MAR Margaritatus Chronozone.
Supplementary Data, Appendix A shows the systematic position of the ammonites and the levels in which the studied specimens were collected in each section; the specimens included in Figs. 8, 9, 10 and 11 are indicated. Appendix B indicates the base of the Horizons (= Zonules) identified, along with their main elements of correlation with the Mediterranean Province.

Representative ammonites of Jamesoni and Ibex chronozones. a Apoderoceras cf. subtriangulare (Young & Bird) SM7(+ 33). b Apoderoceras dunrobinense Spath ER384. c Uptonia jamesoni (J. de C. Sowerby) LL60T. d Acanthopleuroceras valdani (d’Orbigny) LL63T. e Acanthopleuroceras maugenesti (d’Orbigny) LL63(-5). f Uptonia 66 regnardi (d’Orbigny) LL60(-50). g Tropidoceras erythraeum (Gemmellaro) LL62(-15). h Tropidoceras flandrini (Dumortier) LL62(-22). i Beaniceras centaurus (d’Orbigny) 68 LL64(+ 160). Scale bar = 1 cm, except Fig. c.

Representative ammonites of the Raricostatum, Jamesoni and Ibex chronozones. a Paltechioceras tardecrescens (Hauer) ER342. b Gemmellaroceras aff. aenigmaticum (Gemmellaro) ER387m. c Polymorphites lineatus (Quenstedt) MUJA2828. d Polymorphites trivialis (Simpson) SM52. e Polymorphites gr. polymorphus morphotype mixtus (Quenstedt) MUJA2027. f Platypleuroceras caprarium (Quenstedt) SM52. g Platypleuroceras aureum (Simpson) LL45(+ 55). h-i Phricodoceras taylori (J. de C. Sowerby) MUJA2032. j Platypleuroceras rotundum (Quenstedt) LL45(+ 30). k Platypleuroceras brevispinoides (Tutcher & Trueman) LL45(+ 40). l Platypleuroceras submuticum (Oppel) SM55. m Uptonia involuta Meister SM66(+ 20). MUJA5091a. n Platypleuroceras brevispina (J. de C. Sowerby) LL45(+ 30). o Uptonia bronni (Roemer) ER481(+ 20). p Uptonia aff. confusa (Quenstedt) SM62(-33). MUJA5114b. q Uptonia lata (Quenstedt) ER481(+ 10). r Tropidoceras calliplocum (Gemmellaro) ER495s. s Uptonia costosa (Quenstedt) ER473(+ 70). All full-size specimens, except Fig. a and Fig. b which are for 1.5. Scale bar = 2 cm.

Representative ammonites of the Ibex y Davoei chronozones. a Tropidoceras demonense (Gemmellaro) LL62(-15). b Tropidoceras stahli (Oppel) ER481b. c Acanthopleuroceras alisiense (Reynès in Haug) PV357T. d Acanthopleuroceras actaeon (d’Orbigny) ER509. e Acanthopleuroceras arietiforme (Oppel) LL63(-5). f Beaniceras cf. rotundum Buckman ER521m. g Beaniceras crassum Buckman ER521m. h Beaniceras luridum (Simpson) ER525m. i Aegoceras maculatum (Young & Bird) ER527B. j Aegoceras lataecosta (J. de C. Sowerby) ER527s. k Aegoceras capricornus (Schlotheim) ER528. l Oistoceras angulatum (Quenstedt). ER530i. m Prodactylioceras davoei (J. Sowerby) PV371s. n Prodactylioceras rectiradiatum (Wingrave) ER527m. All full-size specimens. Scale bar = 2 cm.

a Reynesocoeloceras praeincertum Dommergues & Mouterde ER527i. b Gemmellaroceras aenigmaticum (Gemmellaro) ER487m. c Radstockiceras gemmellaroi (Pompeckj) ER513s. d Tragophylloceras loscombi (J. Sowerby) PV375. e Protogrammoceras (Matteiceras) occidentale Dommergues, 1982 ER535m. All full-size specimens. Scale bar = 2 cm.
The Raricostatum Chronozone and the Aplanatum Subchronozone are well represented in ER and PV sections (Comas-Rengifo et al., 2010; Comas-Rengifo & Goy, 2010). In the uppermost part of the Aplanatum Subchronozone we have characterised a Tardecrescens Horizon (Corna et al., 1997). P. tardecrescens associated with Leptechioceras macdonnelli, at the base, where Leptonotoceras abnorme, Gleviceras cf. guibalianum and Epideroceras (?) sp. (= Villania, según Howarth, 2013) are also to be found. This assemblage is similar to those found in the N of the Iberian Peninsula (Comas-Rengifo, 1982; Braga et al., 1988; Comas-Rengifo et al., 2010, 2013; Duarte et al., 2014) and in the French-Spanish Pyrenees (Fauré, 2002; Fauré & Teodori, 2019), as well as in some basins in the Euroboreal and NW European areas (Corna et al., 1997; Blau & Meister, 2000; Howarth, 2002; Page, 2003, 2009, etc.).
The succession obtained in the Lower Pliensbachian is similar to the one used in publications that consider the concept of Zonule (= chronohorizon, sensu Hedberg, 1976 and in accordance with Meister, 2010). Moreover, in addition to the studies cited above in order to correlate these, we considered the work of the following authors to be of interest: Dommergues (1979), Braga et al. (1982), Dommergues et al. (1983), Meister (1995, 2010), and Dommergues & Meister (2008a, b).
The lower boundary of the Pliensbachian has been situated between the level ER348, which contains the last P. tardecrescens recorded, and the level ER357 with Gemmellaroceras aff. aenigmaticum a species mainly known in the Mediterranean Province and in the southern areas of the NW European Province. Above this level, in sections ER and SM, there are several levels containing Apoderoceras. Some correspond to A. subtriangulare, and other more recent ones, which are large and badly preserved as a result of marine abrasion, are very close to A. dunrobinense. They could be associated with P. taylori and they characterise the Taylori Subchronozone. In levels equivalent to intervals ER398-ER418 and SM19-SM24 pyritised fragments of P. gr. taylori were found. The “ex situ” specimens obtained by Suárez Vega (1974, p. 65) might have come from these levels or others located in the vicinity. With the exception of these Gemmellaroceras, this succession is similar to the one described by Meister et al. (2006) in Wine Haven, Yorkshire, and very close to the one found in San Pedro de Moel in Portugal (Meister et al., 2012; Duarte et al., 2014).
In the Jamesoni Chronozone, whenever posible the horizons were based on the evolution of Polymorphytinae species, sporadically associated with Phricodoceratidae, Oxynoticeratidae and Coeloceratidae. The Taylori Subchronozone starts with the Horizon aff. Aenigmaticum, which had already been mentioned in the NE of Spain by Comas-Rengifo (1982), followed by the Subtriangulare and Taylori horizons. In the Polymorphus, Brevispina and Jamesoni subchronozones, the horizons were established also considering the evolution of Polymorphytinae species. In the lower part of the Polymorphus Subchronozone, almost no significant ammonites were recorded, and consequently, the lower boundary could not be established. In the upper part, albeit scarce, Polymorphites trivialis and Polymorphites sp. were found; in ER and SM sections. They are associated with P. caprarium. In Vega de Sariego (only in this locality, situated around 12 km to the S of the coast), Suárez Vega (1974) cites the following species in equivalent levels: P. gr. polymorphus (e.g., P. mixtus, P. costatus, P. polymorphus) and P. lineatus. Additionally, the species P. caprarium was cited by some authors in the upper part of the Polymorphus Subchronozone in Germany and Switzerland (Schlatter, 1977, 1991; Meister & Loup, 1989; Meister, 1991; Blau et al., 2000), the UK (Oates, 1978; Howarth, 2002), NE of Spain (Comas-Rengifo, 1982), France (Dommergues, 1987; Fauré & Bohain, 2017) and Denmark (Donovan & Surlyk, 2003).
The Platypleuroceras are dominant in the Brevispina Subchronozone, which can be associated with G. aenigmaticum. However, Phricodoceras, Parinodiceras and Metaderoceras specimens are scarce. One single specimen of the latter genus, M. cf. muticum D’Orbigny (sensu Fauré & Bohain, 2017), has been found in Huerres (Fig. 1), a locality situated between SM and PV. A Brevispina Horizon has been characterised in this subchronozone, followed by a Submuticum Horizon, as occurs in the NW of Europe.
The species of Uptonia dominate in the Jamesoni Subchronozone: The Bronni Horizon was characterised with the index species associated with U. jamesoni and with scarce Gemmellaroceras, along with the Lata Horizon, with the oldest Tropidoceras: T. erythraeum (Braga & Rivas, 1985; Page, 2003).
In the uppermost part of the Jamesoni Subchronozone and in the lower part of the Ibex Chronozone (Masseanum Subchronozone), Tropidoceras typical of the Mediterranean Province (Linares & Rivas, 1973; Gèczy, 1976; Braga & Rivas, 1985) appear in the Asturian Basin, such as T. erythraeum, followed by T. demonense-flandrini, and T. calliplocum-mediterraneum(?), up to the Valdani Subchronozone. Also recorded in this basin were: T. masseanum and T. stahli, species that are known in the lower-middle part of the Ibex Chronozone in the NW of Europe (Dean et al., 1961; Schlatter, 1977, 1980; Dommergues & Mouterde, 1978; Meister, 1986; Dommergues et al., 1997; Page, 2003; Fauré & Bohain, 2017). The mentioned species were also cited in Submediterranean areas (Kollarova-Andrusova, 1966; Patrulius & Popa, 1971; Popa & Patrulius, 1996; Fauré, 2002; Fauré & Teodori, 2019) and T. stahli, previously cited in Asturias by Suárez Vega (1974, pl. 6 A, Fig. 5), is a relatively generalist species from the NW European Province and adjacent areas.
In the Ibex Chronozone (Masseanum and Valdani subchronozones) the Masseanum Subchronozone includes a Masseanum Horizon, where Uptonia persists, and the Valdani Subchronozone comprises five successive horizons based on species of Acanthopleuroceras: Arietiforme, Maugenesti, Valdani, Actaeon and Alisiense. In the last two horizons the first Beaniceras are recorded (Dommergues & Mouterde, 1978, 1981; Dommergues, 1979; Phelps, 1985; Dommergues et al., 1997; Fauré, 2002, among others). The Liparoceratidae of the genera Beaniceras, such as B. rotundum, B. crassum and B. luridum, associated with L. heptangulare are common in the Ibex Chronozone, Luridum Subchronozone. We characterised a thin Rotundum Horizon, presenting, a Crassum Horizon that includes L. heptangulare and R. gemmellaroi, and a Luridum Horizon with the nominal species associated with R. praeincertum and L. fimbriatum.
The Davoei Chronozone presents a succession of several species of Aegoceras. The presence of A. sparsicosta (cited by Trueman, 1919; Spath, 1938; Phelps, 1985; Dommergues & Meister, 1991; Dommergues et al., 1997, 2008a, b; Page, 2003, etc.) is uncertain. Nonetheless, there is a record of Aegoceras sp. (close to A. truemani Fauré) followed by A. maculatum, A. lataecosta and A. capricornus. In a level equivalent to the Crescens Horizon in the standard scale, we found A. cf. crescens and Oistoceras sinuosiforme (Spath, 1938; Howarth, 2002). Lastly, we also found O. angulatum and O. sp. gr. figulinum. Furthermore, in the Capricornus Subchronozone (lower and middle parts) Prodactylioceras rectiradiatum occurs, and between the middle part of the Capricornus Subchronozone and the lower part of the Figulinum Subchronozone, P. davoei is relatively frequent.
In the Upper Pliensbachian (Margaritatus Chronozone) the Amaltheidae are frequent and the index species was previously cited in the Stokesi Subchronozone (Suárez Vega, 1974; Comas-Rengifo & Goy, 2010). At the base of the Upper Pliensbachian, some Protogrammoceratinae, such as Protogrammoceras (Matteiceras) occidentale and P. (M.) monestieri, are common associated with A. stokesi in some levels of PV and ER sections (Rodríguez-Luengo et al., 2012; Gómez et al., 2016a).
4 Discussion
4.1 Correlation with the chronostratigraphic schemes proposed in other european areas
The succession of the assemblages recognised is very similar to that observed in the basins of NW Europe (Figs. 12 and 13) (Dean et al., 1961; Meister, 1986, 1995, 2010; Corna et al., 1997; Dommergues et al., 1997; Howarth, 2002; Page, 2003, 2009; Meister et al., 2006, 2012; Dommergues et al., 2008b; Fauré & Bohain, 2017), particularly to those described in Dorset (S of England) where the stratigraphy also exhibits some episodes that clearly resemble those in Asturias (Lang et al., 1928; Palmer, 1972; Hesselbo & Jenkyns, 1995; Price et al., 2016; Schöllhorn et al., 2020). There are also numerous elements of correlation with the NE and N of Iberia: IB basin (Braga et al., 1982; Comas-Rengifo, 1982), BC basin (Braga et al., 1984, 1985) and PI basin (Fauré, 2002; Fauré & Téodori, 2019).

Correlation with other basins. 1 Dean et al., 1961. 2 Page, 2003. 3 Gómez et al., 2016a. 4 this work, (A) proposed horizons and (B) position of the Mediterranean elements recorded in Asturias. 5 Braga et al., 1982. 6 Comas-Rengifo, 1982. 7 Braga, 1982. 8 Braga et al., 1982. 9 Braga et al., 1984. 10 Braga & Rivas, 1980. 11 Braga & Rivas, 1985. 12 Mouterde et al., 1983. 13 Mouterde et al., 2007. 14 Dommergues & Mouterde, 1981. 15 Dommergues et al., 2011. 16 Duarte et al., 2013. Abbreviations: RAR Raricostatum Chronozone. AP Aplanatum Subchronozone. TA Taylori Subchronozone. PO Polymorphus Subchronozone. BR Brevispina Subchronozone. JA Jamesoni Subchronozone. MA Masseanum Subchronozone. VA Valdani Subchronozone. LU Luridum Subchronozone. MC Maculatum Subchronozone. CA Capricornus Subchronozone. FI Figulinum Subchronozone. MAR Margaritatus Chronozone. ST Stokesi Subchronozone.

Palaeogeographical map of Western Tethys and the Proto-Atlantic Ocean for the Pliensbachian-Toarcian (modified after Ziegler, 1990, Vera, 2001, Stampfli & Borel, 2002, 2004, Osete et al., 2011, Gómez & Goy, 2011) showing the position of the basins cited in the text. LB-Lusitanian Basin, AB-Asturian Basin, BB-Betic Basin, BCB-Basque-Cantabrian Basin, PB-Pyrenean Basin, IB-Iberian Basin, SFB-Southern France Basin. Other abbreviations: EH-Ebro High, FC-Flemish Cap, GB-Grand Banks, LBM-London–Brabant Massif, MC-Massif Central High, NFB-East Newfoundland Basin, mA-Middle Atlas.
In the Portuguese LB, the elements of correlation are remarkable in the Taylori Subchronozone. They are rare between the Polymorphus Subchronozone and the Valdani Subchronozone, and the successions are similar from the Luridum Subchronozone up to the end of the Lower Pliensbachian (Mouterde, 1967; Dommergues & Mouterde, 1981; Mouterde et al., 1983, 2007; Dommergues et al., 1983, 1997; Phelps, 1985; Duarte et al., 2014).
On the contrary, the succession obtained only occasionally resembles those described in Southern Spain and in other basins of the Tethys (Supplementary Data). The main elements of correlation consist of the existence of Gemmellaroceras, Tropidoceras and scarce Metaderoceras and Reynesocoeloceras typical of Mediterranean basins (Cantaluppi & Montanari, 1971; Gèczy, 1976; Rivas, 1972, 1981; Ferretti, 1975, 1991; Wiedenmayer, 1980; Braga et al., 1984; Braga & Rivas, 1985; Hillebrandt, 1987; Ferretti & Meister, 1994; Venturi & Ferri, 2001; Rakus & Guex, 2002; Gèczy & Meister, 2007; Meister et al., 2011, 2017; Dommergues & Meister, 2017, etc.).
In the Polymorphus Subchronozone in Asturias, as occurs in the IB and the BC (Braga et al., 1982, 1985; Comas-Rengifo, 1982), few ammonites were recorded. They rarely corresponded to the index species which, however, have been cited in other areas of Western Europe (Dean et al., 1961; Schlatter, 1980; Dommergues et al., 1997; Howarth, 2002; Page, 2003; Fauré & Bohain, 2017). Nonetheless, the presence in ER454 and SM52 of Polymorphites (P. trivialis, Polymorphites spp.) and Platypleuroceras (P. caprarium), in levels previous to the first record of P. brevispina, enables the levels cited to be attributed to the upper part of the Polymorphus Subchronozone, as has been justified in Sect. 3.2.
The species of Platypleuroceras (P. rotundum, P. brevispina, P. brevispinoides and P. aureum, followed by P. submuticum) and Uptonia (U. costosa, U. confusa, U. bronni, U. jamesoni, U. lata, among others) dominate in the Jamesoni Subchronozone, similar to what occurs in the NW of Europe and the N of Spain (Schlatter, 1980, 1991; Comas-Rengifo, 1982; Braga et al., 1984; Meister, 1986; Dommergues et al., 1997; Fauré, 2002; Sciau, 2004; Fauré & Bohain, 2017; Fauré & Teodori, 2019) and no different from what can be observed in the LB (Mouterde et al., 1983).
In Asturias (ER, LL, PV), Tropidoceras has been recorded in the transition between the Jamesoni and the Ibex chronozones. This fact enables three successive horizons to be characterised: Erythraeum, Demonense-Flandrini and Calliplocum-Mediterraneum, which can be recognised throughout much of the Mediterranean Province (Gèczy, 1976; Braga et al., 1984; Braga & Rivas, 1985; Ferretti, 1991; Meister, 1995, 2010; Page, 2003; Meister et al., 2009, 2011). According to Dommergues et al. (1997, p. 17), these horizons are successively located in the Jamesoni Subchronozone above the Valdani Subchronozone.
In the Valdani Subchronozone, despite the dominance of assemblages presenting elements from the NW of Europe, species typical of the basins of southern Europe arrived to AB. This might have been facilited by the fact that the interval (Lata Horizon to the Valdani Horizon) corresponds to the transgressive máximum of the T3-R3 cycle described by Quesada et al. (2005) in the BC and shows no significantly effective margins to prevent communication between both areas.
The Valdani Subchronozone is well represented by the succession of several species of Acanthopleuroceras, as occurs in other localities in the NW of Europe (Dommerges et al., 1997; Page, 2003) and in basins in the S of France, N of Iberia and the Austroalpine region (Dommergues & Mouterde, 1981; Comas-Rengifo, 1982; Mouterde et al., 1983; Braga et al., 1985, 1988; Meister, 1986; Meister et al., 1994; Fauré, 2002; Meister & Friebe, 2003; Fauré & Téodori, 2019, etc.). In addition, in the Luridum Subchronozone, the succession of Beaniceras (B. rotundum-B. crassum-B. luridum) is very similar to that of NW Europe and is also recorded in the IB and the BC (Dommergues et al., 2008b; Fauré & Bohain, 2017; Fauré & Téodori, 2019), and with slight modifications in the LB (Phelps, 1985).
The Davoei Chronozone is not very thick (approx. 4 m. in ER and 2.5 m. in PV sections). It is represented by the succession of seven species of Aegoceras-Oistoceras, five of which are associated with two successive species of Prodactylioceras: P. rectiradiatum and P. davoei. The transition to the Upper Pliensbachian is well represented in ER and PV. In the uppermost part of the Davoei Chronozone the last Oistoceras (O. sp.) were recorded; they are situated below the first P. (Matteiceras) but, unlike what occurs in other basins in the N of Iberia (BV, PB, IB and LB), a typical O. figulinum was not found. The basal part of the Margaritatus Chronozone is characterised by the presence of P. (M.) occidentale and P. (M.) monestieri, which are common in the Stokesi Subchronozone and appear sporadically with the nominal species of the subchronozone (Comas-Rengifo & Goy, 2010; Rodríguez-Luengo et al., 2012).
4.2 Black shales
Two main metre-scale episodes of dark grey to black organic-rich marls and laminated shales were identified into the Lower Pliensbachian succession within the Rodiles Formation. The lower interval was deposited during a part of the Taylori Subchronozone (Jamesoni Chronozone). The upper interval was deposited in much of the Valdani and Luridum subchronozones (Ibex Chronozone; Fig. 4 g and h), as previously published by Gómez et al. (2016a, b). Both intervals are associated with episodes of basin deepening that are probably influenced by tectonic pulsations, as evidenced by synsedimentary differential subsidence of the ramp into short-lived swells and troughs. In the most subsident depressions these enhanced episodes of tectonic activity, partially controlled by initial halokinesis (García-Senz et al., 2019), generate several organic-rich deposits like the ones identified in other neighbouring palaeogeographic areas (Basque-Cantabrian and Lusitanian basins; Quesada et al., 2005; Silva & Duarte, 2015; Silva et al., 2021). Consequently, all basins in the northwestern half of the Iberian microplate domain shared a similar tectono-stratigraphic evolution. However, they exhibit a certain degree of diachronism resulting from local or regional tectonics.
Furthermore, the deposition of these organic-rich and black shale facies neither seems to be related to large scale palaeoclimatic oscillations, as deduced from data in Price et al. (2016), Bougeault et al. (2017) and Deconinck et al. (2019, 2020), nor nor seems to be associated with water temperature (Gómez et al., 2016a, b). The most likely drivers involve an irregular bottom topography of the ramp, high rates of differential subsidence and highly variable water circulation.
4.3 Modifications in the ammonite assemblages linked to palaeoenvironmental changes
In the interval studied, several palaeoclimatic and environmental changes took place (Robles et al., 2004; Bádenas et al., 2009; Armendáriz et al., 2012; Gómez et al., 2016a, b), which affected the different ammonite groups recorded in the AB.
In the Sinemurian-Pliensbachian transition, the Echioceratidae became extinct shortly after the end of the R2 regressive episode (Quesada et al., 2005), equivalent to the regressive part of the LJ-2.1 cycle described by Gómez & Goy (2004, 2005); this coincides with the beginning of the T3 transgressive episode described by the same authors. In the lower part of the Taylori Subchronozone, a significant transgressive episode took place, probably at a global scale, coinciding with the first episode of black shales. It is characterised by a clearly negative Carbon (δ13C) excursion that is well recorded in the ER section (Gómez et al., 2016b). Some authors postulate that it might have been caused by volcanic activity and the corresponding hydrothermalism associated with the Central Atlantic Magmatic Province (CAMP) and by the resulting opening up of the Hispanic Corridor (Price et al., 2016; Ruhl et al., 2016). This coincides with the arrival to the Asturian Basin of the Polymorphitinae (G. aff. aenigmaticum), typical of the southern areas of the western Tethys.
The boundary between the Jamesoni Chronozone and the Ibex Chronozone is situated over a long deepening stage coinciding with the upper part of the mentioned negative δ13C excursion, before the appearance of the black shale facies present in the Ibex Chronozone. The palaeotemperatures calculated for this boundary can be considered to be normal for this latitude of around 27ºN during the Pliensbachian (Deconinck et al., 2020). Polymorphitinae (Tropidoceras), typical from Southern Europe, are registered in the AB in the uppermost part of Jamesoni Subchronozone.
The boundary between the Ibex Chronozone and the Davoei Chronozone, which marks the substitution of Beaniceras by Aegoceras within the Liparoceratidae family, is located in the upper part of the deepening megasequence that developed during much of the Lower Pliensbachian and it could be linked to a positive δ13C excursion in a very similar position (Luridum Subchronozone) to that cited by Silva et al. (2015) in the LB (Rabaçal and Peniche sections). The positive peak of δ13C in belemnites from the upper part of Ibex Chronozone and the beginning of a relatively short-lived warming in the Lower Pliensbachian detected by Gómez et al. (2016a, b) in Rodiles E are clearly marked by a negative excursion in the values of δ18O recorded in belemnite rostra unaffected by the diagenesis. However, only scarce specimens typical from Mediterranean areas, such as Lytoceras fimbriatum and Reynesocoeloceras praeincertum in the Luridum Subchronozone, R. praeincertum in Maculatum Subchronozone and Tragophylloceras loscombi in the Figulinum Subchronozone, are registered occasionally in the AB. On the other hand, the absence of A. sparsicosta (typical) before the first record of A. maculatum could be linked to the existence of a small hiatus in the lower part of the Maculatum Subchronozone.
In the uppermost part of the Davoei Chronozone, the Aegoceras-Oistoceras became extinct and were substituted, at the base of the Margaritatus Chronozone, by Amaltheidae, such as A. stokesi, and by Hildoceratidae, such as P. (M.) occidentale.
4.4 Palaeobiogeographic situation of the Asturian basin
Throughout the Jamesoni Chronozone, the assemblages obtained (Fig. 12) differ slightly from those of the Northwest European Province (Dommergues & Mouterde, 1978, 1981; Dommergues, 1979, 1987, 2003; Dommergues et al., 1997, 2008b; Blau et al., 2000; Howarth, 2002; Page, 2003, 2009; Dommergues & Meister, 2008b; Meister et al., 2012; Fauré & Bohain, 2017, 2022). The most significant difference involves the existence in Asturias of G. aff. aenigmaticum, in the lower part of the Taylori Subchronozone. In the upper part of this subchronozone the Gemmellaroceras are scarce, with large-sized (> 25 cm) more or less generalist species dominating, such as Phricodoceras and Apoderoceras (A. dunrobinense). From the upper part of the Polymorphus Subchronozone, with Polymorphites trivialis, P. gr. polymorphus etc. and Platypleuroceras caprarium, to the upper part of the Jamesoni Subchronozone, the assemblages are almost identical to those described in the NW of Europe (UK, France and Germany). Moreover, G. aenigmaticum has only been recorded sporadically and, excepcionally, Metaderoceras.
As from the Ibex Chronozone, and until the end of the Lower Pliensbachian, the assemblages are similar to those of the Northwest European Province, with few elements typical of the Mediterranean Province, such as R. praeincertum, in the Luridum-Maculatum transition. Although up to the Upper Pliensbachian, a Submediterranean Province has not yet been established, according to Page (2003), some areas of the W of Europe, such as Subbriançonais, Australpine, Slovakia, French-Spanish Pyrenees, N Iberia (IB, BC and AB in Spain and LB in Portugal, Fig. 13), show enough elements of correlation to establish the Submediterranean Province (Braga et al., 1982; Comas-Rengifo, 1982; Dommergues et al., 1983, 1990, 1995; Mouterde et al., 1983; Blau & Meister, 1991; Dommergues & Meister, 1991; Meister, 1991, 1995, 2010; Fauré & Téodori, 2019).
They are not very similar to the assemblages recorded in the Mediterranean Province and common genera and species are only recorded in some intervals. The principal elements of correlation are: Gemmellaroceras and rare Metaderoceras in the Jamesoni Chronozone, Tropidoceras in the lower and middle parts of the Ibex Chronozone and Reynesocoeloceras in the Ibex-Davoei chronozones transition. In Iberia, in the BB (Subbetic area), species of the four aforementioned genera are to be found (Fig. 12). The succession of Tropidoceras enables relatively accurate correlations to be established with different areas of the Mediterranean. The AB contains species of Tropidoceras, such as: T. erythraeum, T. demonense (m) - flandrini (M) y T. calliplocum (m) that are also recorded in other basins of southern Europe and the N of Africa (Rivas, 1972, 1981; Dommergues, 1979; Braga & Rivas, 1985; Dommergues & Meister, 1990, 2008, 2017; Dommergues et al., 1997; Page, 2003; Gèczy & Meister, 2007; Meister et al., 2011, etc.).
In Italy (Appenines) and Spain (Betic Basin), the Gemmellaroceras are present up to the upper part of the Jamesoni Subchronozone. Several species of Tropidoceras are particularly frequent between the uppermost part of the Jamesoni Subchronozone and the Valdani Subchronozone (Rivas, 1981; Dommergues et al., 1983, 1994; Braga et al., 1984; Braga & Rivas, 1985; Dommergues & Meister, 1990; Donovan, 1990; Ferretti, 1991; Ferretti & Meister, 1994; Venturi et al., 2005, 2007; Blau & Meister, 2011; Meister & Blau, 2014).
In Hungary (Bakony), at the genus level, the main common elements are: Leptotonoceras and Phricodoceras in the transition between the Raricostatum and the Jamesoni chronozones, Platypleuroceras and Uptonia in the Brevispina and Jamesoni subchronozones, respectively, Tropidoceras and Acanthopleuroceras in the Ibex Chronozone, and Aegoceras in the Davoei Chronozone. The most common species are: P. taylori, U. jamesoni, T. flandrini, T. cf. maugenesti, R. praeincertum, A. maculatum, A. capricornus, O. cf. sinuosiforme and P. davoei (Gèczy, 1976; Dommergues et al., 1983; Dommergues & Gèczy, 1989; Gèczy & Meister, 2007).
In Turkey the ammonite succession exhibits noteworthy differences in relation to Asturias (Bremer, 1965; Alkaya & Meister, 1995), although the Ibex Chronozone is also the interval providing a better correlation.
Correlations with the southern margin of the western Tethys (Morocco, Algeria and Tunisia), are difficult after the uppermost Sinemurian, because there are very few common elements up to the upper part of the Jamesoni Chronozone (Meister, 1995, 2010; El Hariri et al., 1996; Rakus & Guex, 2002; Wilmsen et al., 2002; Fauré et al., 2007; Meister et al., 2011; Dommergues & Meister, 2017).
In the Upper Pliensbachian (Stokesi Subchronozone), the succession obtained corresponds to the Submediterranean Province. The species A. stokesi is associated with P. (M.) occidentale and P. (M.) monestieri, and there is a conspicuous absence of Fuciniceras, which, albeit scarce, are present in the BC and IB (Braga et al., 1985).
5 Conclusion
The study of the ammonite species from the uppermost Sinemurian to the lowermost Upper Pliensbachian interval in expanded sections of the Asturian Basin between Ribadesella and Villaviciosa provided new data on the biostratigraphy and chronostratigraphy of the Lower Pliensbachian in the western part of the Cantabrian Range. The position of the biosedimentary events have been established accurately. For instance, the precise situation of two specific anoxic episodes and two warming intervals, the latter coinciding with the positive excursion of δ13C in the Upper Sinemurian and the transition between lower and Upper Pliensbachian. Characterisation of over 95 successive levels containing ammonites in the ER section and other sections, such as LL, SM and PV enabled the scale of reference to be improved. It is now more detailed, of much higher resolution and closer to the standard zoning of the NW of Europe.
The succession obtained highlights some particularities typical of the central sector of the Asturian Basin. The lower boundary of the Pliensbachian is marked by the first appearance of Gemmellaroceras aff. aenigmaticum, first recorded below the first Apoderoceras found. The upper boundary is marked by the first record of Protogrammoceras (Matteiceras), associated in some levels with Amaltheus stokesi. With the exception of the lower boundary of the Pliensbachian and of the base of the Polymorphus Chronozone, all the boundaries between chronozones and subchronozones were accurately established. The assemblages recorded are very similar to those established in the Northwest European Province, but in some intervals taxa typical of the Mediterranean Province are recorded. These assemblages only occur at the base of the Jamesoni Chronozone (Taylori Subchronozone) as well as in the transition between the Jamesoni and the Ibex chronozones, which is prolonged until the Valdani Subchronozone. There are also some sporadic levels showing a small number of Mediterranean ammonites, such as Metaderoceras in the late Polymorphus Subchronozone and Reynesocoeloceras in the transition between the Luridum and Maculatum subchronozones. Additionaly, taxa from the Submediterranean Province have been recorded, e.g. P. (M.) occidentale and P. (M.) nitescens in the Margaritatus Chronozone, Stokesi Subchronozone, from the Upper Pliensbachian.
Moreover, fundamentally based upon the evolution of the species of several families such as Echioceratidae, Phricodoceratidae, Polymorphitidae, Liparoceratidae, Hildoceratidae, twenty-seven ammonoid horizons were characterised for the considered interval: one for the uppermost Sinemurian (Raricostatum Chronozone, Aplanatum Subchronozone), twenty-four for the Lower Pliensbachian and two for the Upper Pliensbachian (Margaritatus Chronozone, Stokesi Subchronozone).
References
Alkaya, F., & Meister, C. (1995). Liassic ammonites from the central and eastern pontides (Ankara and Kelkit áreas, Turkey). Revue de Paléobiologie, 14, 125–193.
Armendáriz, M., Rosales, I., Bádenas, B., Aurell, M., García-Ramos, J. C., & Piñuela, L. (2012). High-resolution chemostratigraphic records from Lower Pliensbachian belemnites: Palaeoclimatic perturbations, organic facies and water mass exchange (asturian basin, northern Spain). Palaeogeography Palaeoclimatology Palaeoecology, 333–334, 178–191. https://doi.org/10.1016/j.palaeo.2010.10.042.
Bádenas, B., Aurell, M., García-Ramos, J. C., González, B., & Piñuela, L. (2009). Sedimentary vs. diagenetic control on rhythmic calcareous successions (Pliensbachian of Asturias, Spain). Terra Nova, 21, 162–170. https://doi.org/10.1111/j.1365-3121.2009.00869.x.
Bádenas, B., Aurell, M., Armendáriz, M., Rosales, I., García-Ramos, J. C., & Piñuela, L. (2012). Sedimentary and chemostratigraphic record of climatic cycles in Lower Pliensbachian marl-limestone platform successions of Asturias (North Spain). Sedimentary Geology, 281, 119–138. https://doi.org/10.1016/j.sedgeo.2012.08.010.
Bandel, K., & Knitter, H. (1986). On the origin and diagenesis of the bituminous Posidonia Shale (Toarcian) of Southern Germany. Mitteilungen aus dem geologish-Palaöntologischen Institut der Universität Hamburg, 60, 151–177.
Bardet, N., Fernández, M., García-Ramos, J. C., Suberbiola, P., Piñuela, X., Ruiz-Omeñaca, L., J. I., & Vincent, P. (2008). A juvenile plesiosaur from the Pliensbachian (lower jurassic) of Asturias, Spain. Journal of Vertebrate Paleontology, 28, 258–263.
Barrois, C. (1882). Recherches sur les terrains anciens des Asturies et de la Galice. Mémoires de la Societé Géologique du Nord, 2, 1–630.
Biernacka, J., Borysiuk, K., & Raczyński, P. (2005). Zechstein (Ca1) limestone-marl Dommergues, 1979 alternations from the North-Sudetic Basin, Poland: Depositional or diagenetic rhythms? Geological Quarterly, 49, 1–14.
Bjerrum, C. J., Surlyk, F., Callomon, J. H., & Slingerland, R. L. (2001). Numerical paleoceanographic study of the early jurassic transcontinental Laurasian Seaway. Paleoceanography, 16, 390–404. https://doi.org/10.1029/2000PA000512.
Blau, J., & Meister, C. (1991). Liassic (Pliensbachian) ammonites from the Lienz Dolomites (Eastern Tyrol, Austria). Jahrbuch der Geologischen Bundesanstalt, A 134, 171–204.
Blau, J., & Meister, C. (2000). Upper Sinemurian Ammonite Succesions based on 41 Faunal Horizons: An attempt at Worlwide correlation. GeoResearch Forum, 6, 3–12.
Blau, J., & Meister, C. (2011). Resolving the Monte di Cetona biostratigraphical enigma – a revision of R. FISCHER´S Sinemurian and Pliensbachian (Liassic) ammonite collection from the Central Apennines (Tuscany, Italy). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 261, 257–287.
Blau, J., Meister, C., Ebel, R., & Schlatter, R. (2000). Upper Sinemurian and Lower Pliensbachian ammonite faunas from Herford-Diebrock area (NW Germany). Paläontologische Zeitschrift, 74, 259–280.
Borrego, A. G., Hagemann, H. W., Blanco, C. G., Valenzuela, M., & de Suárez, C. (1997). The Pliensbachian (early jurassic) “anoxic” event in Asturias, northern Spain: Santa Mera Member, Rodiles formation. Organic Geochemistry, 25, 295–309. https://doi.org/10.1016/S0146-6380(96)00121-0.
Bougeault, C., Pellenard, P., Deconinck, J. F., Hesselbo, S. P., Dommergues, J. L., Bruneau, L., Cocquerez, T., Laffont, R., Huret, E., & Thibault, N. (2017). Climatic and palaeoceanographic changes during the Pliensbachian (early jurassic) inferred from clay mineralogy and stable isotope (C-O) geochemistry (NW Europe). Global and Planetary Change, 149, 139–152. https://doi.org/10.1016/j.gloplacha.2017.01.005.
Braga, J. C. (1982). Ammonites del Domerense de la Zona Subbética (Cordilleras Béticas, Sur de España) PhD thesis, Universidad de Granada. Published for Palaeontology Department, 410 pp. (1983).
Braga, J. C., Comas-Rengifo, M. J., Goy, A., & Rivas, P. (1982). Comparaciones faunísticas y correlaciones en el Pliensbachiense de la Zona Subbética y Cordillera Ibérica. Boletín de la Real Sociedad Española de Historia Natural (Sección Geológica), 80, 221–244.
Braga, J. C., Comas-Rengifo, M. J., Goy, A., & Rivas, P. (1984). The Pliensbachian of Spain: Ammonite successions, boundaries and correlations. In O. Michelsen & A. Zeiss (Eds), International Symposium on Jurassic Stratigraphy (pp. 160–176). Erlangen.
Braga, J. C., Comas-Rengifo, M. J., Goy, A., & Rivas, P. (1985). Le Pliensbachien dans la chaîne cantabrique orientale entre Castillo Pedroso et reinosa (Santander, Espagne). Les Cahiers de l’Institut Catholique de Lyon, 14, 69–83.
Braga, J. C., Comas-Rengifo, M. J., Goy, A., Rivas, P., & Yébenes, A. (1988). El Lías inferior y medio en la zona central de la Cuenca Vasco-Cantábrica (Camino, Santander). III Coloquio de Estratigrafía y Paleogeografía del Jurásico de España. Ciencias de la Tierra, 11, 17–45.
Braga, J. C., & Rivas, P. (1980). Protogrammoceras y Fuciniceras (Ammonoidea, Hildoceratacea) del Carixiense superior en las Cordilleras Béticas. España) Estudios geológicos, 36, 169–176. Andalucía.
Braga, J. C., & Rivas, P. (1985). The mediterranean Tropidoceras (Ammonitina) in the Betic Cordilleras. Eclogae geologicae Helvetiae, 78, 567–605.
Bremer, H. (1965). Zur Ammonitenfauna und Stratigraphie des unteren Lias (Sinemurium bis Carixium) in des Umgebung von Ankara (Türkei). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 122, 127–221.
Cantaluppi, G., & Montanari, L. (1971). Quadro bioestratigráfico conclusivo del Carixiano e suo passagio al Domeriano nelle Prealpi lombarde occidentali. Bolletino della Societá Paleontologica Italiana, 10, 43–74.
Comas-Rengifo, M. J. (1982). El Pliensbachiense de la Cordillera Ibérica. PhD thesis, Universidad Complutense de Madrid. Colección Tesis Doctorales, 19/85, 1–591 (published in 1985).
Comas-Rengifo, M. J., García-Martínez, J. C., & Goy, A. (2010). Sinemuriense Superior en Rodiles (Asturias): Biocronoestratigrafía y biohorizontes de ammonoideos. Comunicaciones del V Congreso del Jurásico de España. Museo del Jurásico de Asturias (MUJA) (pp. 49–56). Colunga.
Comas-Rengifo, M. J., & Goy, A. (2010). Caracterización biocronoestratigráfica del Sinemuriense Superior y el Pliensbachiense entre los afloramientos de Playa de Vega y Lastres (Asturias). In J. C. García-Ramos (Coord.). Las sucesiones margo-calcáreas marinas del Jurásico Inferior y las series fluviales del Jurásico Superior. Acantilados de la playa de Vega (Ribadesella). V Congreso Jurásico de España. Museo del Jurásico de Asturias (MUJA), Colunga, 10 – 18.
Comas-Rengifo, M. J., Duarte, L. V., Goy, A., Paredes, R., & Silva, R. (2013). El Sinemuriense Superior (cronozonas Oxynotum y Raricostatum) en la región de S. Pedro de Moel (Cuenca Lusitánica, Portugal). Comunicações Geológicas, 100, 15–19.
Corna, M., Dommergues, J. L., Meister, C., & Mouterde, R. (1997). Sinemurian. In E. Cariou &. P. Hantzpergue (Coord.). Biostratigraphie du Jurassique ouest-européen et méditerranéen: zonations parallèles et distribution des invertébrés et microfossiles. Bulletin du Centre de Recherches Elf Exploration Production, Mem. 17, 9–14.
Dean, W. T., Donovan, D. T., & Howarth, M. K. (1961). The Liassic Ammonite Zones and Subzones of the North-West European Province. Bulletin of the British Museum of Natural History, Geology, 4, 435–505.
Deconinck, J. F., Gómez, J. J., Baudin, F., Biscay, H., Bruneau, L., Cocquerez, T., Mathieu, O., Pellenard, P., & Santoni, A. L. (2020). Diagenetic and environmental control of the clay mineralogy, organic matter and stable isotopes (C, O) of jurassic (pliensbachian-lowermost Toarcian) sediments of the Rodiles section (Asturian Basin, Northern Spain). Marine and Petroleum Geology, 115, 104286. https://doi.org/10.1016/j.marpetgeo.2020.104286
Deconinck, J. F., Hesselbo, S. P., & Pellenard, P. (2019). Climatic and sea-level control of jurassic (pliensbachian) clay mineral sedimentation in the Cardigan Bay Basin, Llanbedr (Mochras Farm) borehole, Wales. Sedimentology, 66, 2769–2783. https://doi.org/10.1111/sed.12610
Dommergues, J. L. (1979). Le Carixien bourguignon: biostratigraphie, paléogéographie, approches paléontologique et sedimentologique. PhD thesis, Dijon University, France, 195p.
Dommergues, J. L. (1987). L’évolution chez les ammonitina du lias moyen (Carixien,Domérien basal) en Europe occidentale. Documents des Laboratoires de Géologie de Lyon, 98, 1–297.
Dommergues, J. L. (2003). Nouvelles données sur les ammonites du Carixien basal (jurassique inférieur) en Europe du Nord-Ouest: Les faunes de corbigny (Nièvre, Bourgogne, France). Bulletin scientiphique de Bourgogne, 51, 12–36.
Dommergues, J. L., Cattaneo, G., Aïte, R., & Gélard, J. P. (2008a). Les ammonites de l’Hettangien, du Sinemurien et du Pliensbachien inférieur de la Dorsale de Grande Kabylie (Algérie). Geodiversitas, 30, 539–576.
Dommergues, J. L., Dugué, D. J. L., Gauthier, O., Meister, H., Neige, C., Raynaud, P., Savary, D., X., & Trevisan, M. (2008b). Les ammonites du Pliensbachian et du Toarcian basal dans la carrière de la Roche Blain (Fresnay-le-Puceux, Calvados, Basse Normandie, France). Taxonomie, implications stratigraphiques et paléobiogeographiques. Révue de Paléobiologie, 27, 265–329.
Dommergues, J. L., Ferretti, A., Gèczy, B., & Mouterde, R. (1983). Eléments de corrélation entre faunes d’ammonites mésogéennes (Hongrie, Italie) et subboréales (France, Portugal) au Carixien et au Domérien inférieur. Geobios, 16, 471–499. https://doi.org/10.1016/S0016-6995(83)80106-5.
Dommergues, J. L., Ferretti, A., & Meister, C. (1994). Les faunes d’ammonites du Sinémurien de l’Apennin Central (Marches et toscane, Italie). Bollettino della Società Paleontologica Italiana, 33, 13–42.
Dommergues, J. L., & Gèczy, B. (1989). Les faunes d’ammonites du Carixien basal de Villany (Hongrie): Un toin paléobiogéographique des peuplements de la Marge méridionalle du continent Euro-Asiatique. Revue de Paléobiologie, 8, 21–37.
Dommergues, J. L., & Meister, C. (1990). Les faunes d’ammonites de l’Austroalpin moyen dans les alpes rhétiques italiennes (région de Livigno); biostratigraphie et implications paléogéographiques. Revue de Paléobiologie, 9, 291–307.
Dommergues, J. L., & Meister, C. (1991). Succession des faunes d’ammonites du Sinémurien et du pliensbachien dans les Chablais septentrional (Préalpes médianes, Suisse et France). Bollettino della Società Paleontologica Italiana, 30, 303–324.
Dommergues, J. L., & Meister, C. (2017). Ammonites du Jurassique inférieur (Hettangien, Sinémurien, Pliensbachien) d’Afrique du Nord (Algérie, Maroc et tunisie). Atlas d’identification des especès. Revue de Paléobiologie, 36, 189–367.
Dommergues, J. L., Meister, C., & Böhm, F. (1995). New data an Austroalpine Liassic ammonites from the Adnet quarries and adjacents áreas (Oberösterreich, Northern Calcareous Alps). Jahrbuch der Geologischen Bundesanstalt, 138), 161–205.
Dommergues, J. L., Meister, C., & Mettraux, M. (1990). Succession des faunes d’ammonites du Sinémurien et du pliensbachien dans les Préalpes romandes (Vaud et Fribourg). Geobios, 23, 307–341. https://doi.org/10.1016/0016-6995(90)80004-Y.
Dommergues, J. L., Meister, C., & Mouterde, R. (1997). Pliensbachien. In E. Cariou &. P. Hantzpergue (Coord.). Biostratigraphie du Jurassique ouest-européen et méditerranéen. Zonations parallèles et distribution des invértebrées et microfossiles. Bulletin du Centre de Recherches Elf Exploration Production, Mem. 17, 15–23.
Dommergues, J. L., Meister, C., & Rocha, R. B. (2011). The Pliensbachian ammonites of the Algarve Basin (Portugal) and their palaeobiogeographical significance for the ‘‘Iberia-Newfoundland’’ conjugate margins. Swiss Journal of Geosciences, 104, 81–96. https://doi.org/10.1007/s00015-011-0056-2.
Dommergues, J. L., & Mouterde, R. (1978). Les faunes d’ammonites du Carixien inférieur et moyen du gisement des cottards (Cher). Geobios, 11(3), 345–365.
Dommergues, J. L., & Mouterde, R. (1981). Les acanthopleurocératinés portugais et leurs relations avec les formes subboréales. Ciências da Terra (UNL), 6, 77–100.
Donovan, D. T. (1990). Sinemurian and pliensbachian ammonites faunas of Central Italy. Atti del Secondo Convegno Internazionale Fossili Evoluzioni Ambiente, 2, 253–262.
Donovan, D. T., & Surlyk, F. (2003). Lower jurassic (pliensbachian) ammonites from Bornholm, Baltic Sea, Denmark. Geological Survey of Denmark and Greenland Bulletin, 1, 555–583.
Duarte, L. V., Comas-Rengifo, M. J., Silva, R. L., Paredes, R., & Goy, A. (2014). Carbon isotope stratigraphy and ammonite biochronostratigraphy across the sinemurian-pliensbachian boundary in the western Iberian margin. Bulletin of Geosciences, 89(4), 719–736. https://doi.org/10.3140/bull.geosci.1476.
Dubar, G. (1925). Sur les formations du Lias et du Jurassique supérieur dans les Asturies. Comptes Rendues hebdomadaires des Séances de l’Académie des Sciences, 180, 215–217.
Dubar, G., & Mouterde, R. (1957). Précisions sur le lias des asturies. Comptes Rendues hebdomadaires des Séances de l’Académie des science. Paris Ser D, 244, 1234–1236.
Dubar, G., Mouterde, R., Virgili, C., & Suárez, L. C. (1972). El Jurásico de Asturias (Norte de España). Cuadernos de Geología Ibérica, 2, 561–580.
El Hariri, K., Dommergues, J. L., Meister, C., Souhel, A., & Chafiki, D. (1996). Les ammonites du Lias inférieur et moyen du Haut-Atlas de Béni Méllal (Maroc): Taxinomie, et biostratigraphie à haute résolution. Geobios, 29, 537–576. https://doi.org/10.1016/S0016-6995(96)80025-8.
Fauré, P. (2002). Le Lias des Pyrénées. Strata Ser II, 39, 761.
Fauré, P., Alméras, Y., Sekatni, M., & Zargouni, F. (2007). Le Pliensbachien de Jebel Zaghouan (Tunisia). Nouvelles données fauniques. Implications biostratigraphiques et paléobiogéographiques. Geodiversitas, 29, 473–506.
Fauré, P., & Bohain, P. (2017). Les ammonites du Pliensbachien inférieur de la Vendée méridionale (France). Étude taxonomique. Implications stratigraphiques et paléogéographiques. Strata, 54, 1–147.
Fauré, P., & Bohain, P. (2022). Pliensbachian ammonites from Southern Vendée (France). Toward the individualization of an Atlantic paleobiogeographic region. Comptes Rendus Géoscience-Sciences de la Planète, 354, 5–25. https://doi.org/10.5802/crgeos.136.
Fauré, P., & Téodori, D. (2019). Les ammonites du Pliensbachien des Pyrénées ariégeoises (Zone nord-pyrénéenne, France): Taxonomie, stratigraphie et implications paléogéographiques. Revue de Paléobiologie, 38, 269–361. https://doi.org/10.5281/zenodo.3579349.
Fernández, M. S., Piñuela, L., & García-Ramos, J. C. (2018). First report of Leptonectes (Ichthyosauria: Leptonectidae) from the Lower Jurassic (Pliensbachian) of Asturias, northern Spain. Palaeontologia Electronica, 21.2.29A, 1–15. https://doi.org/10.26879/802
Ferretti, A. (1975). Ricerche biostratigrafiche sul Sinemuriano-Pliensbachiano nella Gola del F. Bosso (Appennino marchigiano). Rivista Italiana di Paleontología e Stratigrafia, 81, 161–194.
Ferretti, A. (1991). Introduzione ad uno studio morfometrico degli ammoniti pliensbachiani della catena del catria (Appennino Marchigiano). Rivista Italiana di Paleontología e Stratigrafia, 97, 49–98.
Ferretti, A., & Meister, C. (1994). Composition des faunes d’ammonites dans les Apennins des Marches et comparaison avec les principales régions téthysiennes et subboreales. In G. Pallini (Ed.) 3ème Convegno Internazionale Fossili, Evoluzione, Ambiente. – Palaeopelagos, 143–153.
French, K. L., Sepúlveda, J., Trabucho-Alexandre, J., Gröcke, D. R., & Summons, R. E. (2014). Organic geochemistry of the early Toarcian oceanic anoxic event in Hawsker Bottoms, Yorkshire, England. Earth and Planetary Science Letters, 390, 116–127. https://doi.org/10.1016/j.epsl.2013.12.033.
Gani, M. R. (2017). Mismatch between time surface and stratal surface in stratigraphy. Journal of Sedimentary Research, 87, 1226–1234. https://doi.org/10.2110/jsr.2017.67.
García-Senz, J., Rosales, I., Robador, A., & Pedrera, A. (2019). The Basque-Cantabrian rift basin. In J. Martin-Chivelet et al., The Late Jurassic-Early Cretaceous Rifting, Quesada, C. & J. T. Oliveira (Eds.), The Geology of Iberia: A Geodynamic Approach, Springer Nature, 171–182. https://doi.org/10.1007/978-3-030-11295-0
García-Ramos, J. C. Coord. (2010). Las sucesiones margo-calcáreas marinas del Jurásico Inferior y las series fluviales del Jurásico Superior. Acantilados de la playa de Vega (Ribadesella). Guía de campo, excursión A, V Congreso del Jurásico de España. Museo del Jurásico de Asturias, Colunga, 85 p.
García-Ramos, J. C., & Gutiérrez Claverol, M. (1995). In C. Aramburu, & F. Bastida (Eds.), La geología de la franja costera oriental y de la depresión prelitoral de Oviedo-Cangas de Onís (pp. 247–258). Geología de Asturias, Editorial Trea.
García-Ramos, J. C., Piñuela, L., Bádenas, B., & Aurell, M. (2010). Ciclos elementales de escala milenaria en una ritmita de marga-caliza del Pliensbachiense de Asturias (Formación Rodiles). Comunicaciones del V Congreso del Jurásico de España. Museo del Jurásico de Asturias (MUJA), Colunga, 73–82.
García-Ramos, J. C., Piñuela, L., & Rodríguez-Tovar, F. J. (2011). Post-workshop field trip guide of the XI International Ichnofabric Workshop (p. 89). Colunga: Museo del Jurásico de Asturias (MUJA).
García-Ramos, J. C., Valenzuela, M., & Suárez de Centi, C. (Eds.). (1992). Icnofósiles, procesos sedimentarios y facies en una rampa carbonatada del Jurásico de Asturias. Reunión Monográfica sobre Biosedimentación, Guía de Campo. Excursión B. Sociedad Geológica de España y Universidad de Oviedo, 1–89.
Gèczy, B. (1976). Les ammonites du Carixien de la montagne du Bakony (p. 220). Budapest: Akadémiai Kiado.
Gèczy, B., & Meister, C. (2007). Les ammonites du Sinémurien et du Pliensbachien inférieur de la montagne du Bakony (Hongrie). Revue de Paléobiologie, 26, 137–305.
Gómez, J. J., Comas-Rengifo, M. J., & Goy, A. (2016a). Palaeoclimatic oscillations in the Pliensbachian (early jurassic) of the Asturian Basin. Climate in the Past, 12, 1199–1214. https://doi.org/10.5194/cp-12-1199-2016.
Gómez, J. J., Comas-Rengifo, M. J., & Goy, A. (2016b). The hydrocarbon souce rocks of the Pliensbachian (early jurassic) in the Asturian Basin (northern Spain): Their relationship with the palaeoclimatic oscillations and gamma-ray response. Journal of Iberian Geology, 42, 259–273. https://doi.org/10.5209/JIGE.53265.
Gómez, J. J., & Goy, A. (2004). Cordilleras Ibérica y Costero-Catalana. Jurásico Inferior. In J. A. Vera (Ed.), Geología de España (pp. 495–500). Madrid: SGE-IGME.
Gómez, J. J., & Goy, A. (2005). Late Triassic and Early Jurassic palaeogeographical evolution and depositional cycles of the Western Tethys Iberian platform system (Eastern Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 222, 77–94. https://doi.org/10.1016/j.palaeo.2005.03.010
Gómez, J. J., & Goy, A. (2011). Warming-driven mass extinction in the early Toarcian (early jurassic) of northern and central Spain. Correlation with other time-equivalent european sections. Palaeogeography Palaeoclimatology Palaeoecology, 306, 176–195. https://doi.org/10.1016/j.palaeo.2011.04.018.
Hedberg, H. D. (1976). International Stratigraphic guide. A guide to stratigraphic classification, terminology, and procedure. New York: John Wiley & Sons.
Hesselbo, S. P., & Jenkyns, H. C. (1995). A comparison of the Hettangian to Bajocian successions of Dorset and Yorkshire. In P. D. Taylor (Ed.), Field Geology of the british jurassic (pp. 105–150). London: Geological Society.
von Hillebrandt, A. (1987). Liassic ammonite zones of South America and correlations with others provinces. With description of new genera and species of ammonites. In W. Volkheimer (Ed.), Bioestratigrafia de los Sistemas Regionales del Jurásico y Cretácico en América del Sur (pp. 111–157). Mendoza.
Howarth, M. K. (2002). The Lower Lias of Robin Hood’s Bay, Yorkshire, and the work of Leslie Bairstow. Bulletin of the Natural History Museum Geology, 58, 81–152. https://doi.org/10.1017/S0968046202000037.
Howarth, M. K. (2013). Mollusca 4, Part L (revised), volume 3B, triassic and jurassic Ammonoidea. In P. A. Selden (Ed.), Treatise on Invertebrate Paleontology Online, 57 (pp. 1–139). Paleontological Institute. University of Kansas Press.
Jiménez de Cisneros, D. (1904). Datos para el estudio del sistema liásico de Asturias. –Boletín de la Real Sociedad Española de Historia Natural, 4, 298–305.
Kollarova-Andrusova, V. (1966). Les céphalopodes du Lias du Slovensky Kras (Karst Slovaque). Nauka Zemi Sér Geologica, 3, 7–77.
Lang, W. D., Spath, L. F., Cox, L. R., & Muir-Wood, H. M. (1928). The Belemnite Marls of Charmouth, a series in the Lias of the Dorset coast. Quarterly Journal of the Geological Society of London, 84, 179–257. https://doi.org/10.1144/GSL.JGS.1928.084.01-04.06.
Leonowicz, P. (2015). Storm-influenced deposition and cyclicity in a shallow-marine mudstone succession-example from the middle jurassic ore-bearing clays of the polish Jura (southern Poland). Geological Quarterly, 59, 325–344. https://doi.org/10.7306/gq.1221.
Linares, A., & Rivas, P. (1973). El Lías medio en la parte sur de la Zona Subbética (Sierra Elvira, Illora e Iznalloz, prov. De Granada). Cuadernos de Geología Universidad de Granada, 4, 141–154.
Mallada, L. (1885). Sinopsis de las especies fósiles que se han encontrado en España. Tomo 2. Terreno Mesozoico (Sistemas Triásico y Jurásico). Boletín de la Comisión del Mapa Geológico de España, 2, 1–150.
Meister, C. (1986). Les ammonites du Carixien des Causses (France). Mémoires suisses de Paléontologie, 109, 1–209.
Meister, C. (1991). Biostratigraphie des ammonites liasiques des alpes de Glaris: Comparaisons et corrélations avec la région de ferdenrothorn (Valais). Eclogae geologicae Helvetiae, 84, 223–243.
Meister, C. (1995). Essai de corrélations au Lias moyen (Sinémurien supérieur et carixien) entre les Pontides et les principales régions adjacentes de la Tethys occidentale et de l’Europe du Nord-Ouest. Hantkeniana, 1, 75–82.
Meister, C. (2010). Worldwide ammonite correlation at the Pliensbachian Stage and Substage boundaries (lower jurassic). Stratigraphy, 7, 83–101.
Meister, C., Aberhan, M., Blau, J., Dommergues, J. L., Feist-Burkhardt, S., Hailwood, E. A., Hart, M., Hesselbo, S. P., Hounslow, M. W., Hylton, M., Morton, N., Page, K., & Price, G. D. (2006). The global Boundary Stratotype Section and Point (GSSP) for the base of the Pliensbachian Stage. Episodes, 29, 93–106. (Lower Jurassic), Wine Haven, Yorkshire, UK.
Meister, C., & Blau, J. (2014). Pliensbachian ammonites from the Central Apennines, Italy (Acquasparta section) – A revisión of Fischer’s collection and new data. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 273, 253–275. https://doi.org/10.1127/0077-7749/2014/0430
Meister, C., Blau, J., & Böhm, F. (1994). Ammonite biostratigraphy of the pliensbachian stage in the Upper Australpine jurassic. Eclogae Geologicae Helvetiae, 87, 139–155.
Meister, C., Dommergues, J. L., Dommergues, C., Lachkar, N., & Hariri, E., K (2011). Les ammonites du Pliensbachien du jebel Bou Rharraf (Haut Atlas oriental, Maroc). Geobios, 44, 117e1–117e60. https://doi.org/10.1016/j.geobios.2010.06.006.
Meister, C., Dommergues, J. L., & Rocha, R. B. (2012). Ammonites from the Apoderoceras beds (early pliensbachian) in São Pedro de Muel (Lusitanian Basin, Portugal). Bulletin of Geosciences, 87, 407–430. https://doi.org/10.3140/bull.geosci.1325.
Meister, C., & Friebe, J. G. (2003). Austroalpine Liassic ammonites from Vorarlberg (Austria, Northern Calcareous Alps). Beiträge Paläontologie, 28, 9–99.
Meister, C., & Loup, B. (1989). Les gisements d’ammonites liasiques (Hettangien à Pliensbachien) du Ferdenrothorn (Valais, Suisse): Analyses paléontologiques, biostratigraphiques et aspects lithostratigraphiques. Ecoglae Geologicae Helvetiae, 82, 1003–1041.
Meister, C., Schirolli, P., & Dommergues, J. L. (2009). Sinemurian to lowermost Toarcian ammonites of the Brescian Prealps (Southern Alps, Italy): Preliminary biostratigraphical framework and correlations. Volumina Jurassica, 7, 9–18.
Meister, C., Schirolli, P., & Dommergues, J. L. (2017). Early jurassic (sinemurian to basal Toarcian) ammonites of the Brescian Prealps (Southern Alps, Italy). Rivista Italiana di Paleontologia e Stratigrafia, 123, 79–148.
Mouterde, R. (1967). Le Lias du Portugal. Vue d’ensemble et divisións en zones. – Comunicaçoes Serviços Geológicos de Portugal, 52, 209–226.
Mouterde, R., Dommergues, J. L., & Rocha, R. B. (1983). Atlas des fossiles caractéristiques du lias portugais, II: Carixien. Ciências da Terra (UNL), 7, 187–254.
Mouterde, R., Dommergues, J. L., Meister, C., & Rocha, R. B. (2007). Atlas des fossiles caractéristiques du lias portugais, IIIa: Domérien (Ammonites). Ciências da Terra (UNL), 16, 67–111.
Munnecke, A., Westphal, H., Elrick, M., & Reijmer, J. J. G. (2001). The mineralogical composition of precursor sediments of calcareous rhythmites: A new approach. International Journal of Earth Sciences, 90, 795–812. https://doi.org/10.1007/s005310000137.
Nohl, T., & Munnecke, A. (2019). Reconstructing time and diagenesis of limestone-marl alternations from the selective compaction of colonies of the tabulate coral Halysites. Bulletin of Geosciences, 94(3), 279–298. https://doi.org/10.3140/bull.geosci.1752.
Nohl, T., Wetterich, J., Fobbe, N., & Munnecke, A. (2020). Lithological dependence of aragonite preservation in monospecific gastropod deposits of the Miocene Mainz Basin: Implications for the (dia-)genesis of limestone-marl alternations. Journal of Sedimentary Research, 90, 1500–1509. https://doi.org/10.2110/jsr.2020.057.
Nohl, T., Steinbauer, M. J., Sinnesael, M., & Jarochowska, E. (2021). Detecting initial aragonite and calcite variations in limestone-marl alternations. Sedimentology, 68, 3102–3115. https://doi.org/10.1111/sed.12885.
Oates, M. J. (1978). A revised stratigraphy for the western Scottish Lower Lias. Proceedings of the Yorkshire Geological Society, 42, 143–156. https://doi.org/10.1144/pygs.42.1.143
Osete, M. L., Gómez, J. J., Pavón-Carrasco, F. J., Villalaín, J. J., Palencia-Ortas, A., Ruiz-Martínez, V. C., & Heller, F. (2011). The evolution of Iberia during the jurassic from palaeomagnetic data. Tectonophysics, 502, 105–120. https://doi.org/10.1016/j.tecto.2010.05.025.
Page, K. N. (2003). The lower jurassic of Europe: Its subdivision and correlation. Geological Survey of Denmark and Greenland Bulletin, 1, 23–59. https://doi.org/10.34194/geusb.v1.4646.
Page, K. N. (2009). High resolution ammonite stratigraphy of the Charmouth Mudstone formation (lower jurassic: Sinemurian-Lower Pliensbachian) in south-west Ingland, UK. Volumina Jurassica, 7, 19–29.
Palmer, C. P. (1972). A revisión of the zonal classification of the lower Lias of the Dorset coast of south-west England. Newsletters on Stratigraphy, 2, 45–54. https://doi.org/10.1127/nos/2/1972/45.
Patrulius, D., & Popa, E. (1971). Lower and middle jurassic ammonite zones in the Roumanian Carpathians. Annales Instituti Geologici Publici Hungarici, 54, 131–147.
Phelps, M. C. (1985). A refined ammonite biostratigraphy for the Middle and Upper Carixian (Ibex and Davoei zones, lower jurassic) in North-West Europe and stratigraphical details of the Carixian-Domerian boundary. Geobios, 18, 321–367. https://doi.org/10.1016/S0016-6995(85)80098-X.
Popa, E., & Patrulius, D. (1996). Lower jurassic ammonites in the romanian Carphatians. Mémoires de l’Institut géologique roumain, 36, 53–63.
Price, G. D., Baker, S. J., Van De Velde, J., & Clémence, M. E. (2016). High-resolution carbon cycle and seawater temperature evolution during the early jurassic (sinemurian -Early pliensbachian). Geochemistry Geophysics Geosystems, 17, 3917–3928. https://doi.org/10.1002/2016GC006541.
Quesada, S., Robles, S., & Rosales, I. (2005). Depositional architecture and transgressive-regressive cycles within liassic backstepping carbonate ramps in the Basque-Cantabrian Basin, northern Spain. Journal of the Geological Society London, 162, 531–548. https://doi.org/10.1144/0016-764903-041.
Rakus, M., & Guex, J. (2002). Les ammonites du Jurassique inférieur et moyen de la dorsale tunisienne. Mémoires de Géologie (Lausanne), 39, 1–217.
Ricken, W. (1986). Diagenetic bedding. A model of marl-limestone alternations. Lecture Notes in Earth Sciences, 6, Springer-Verlag, Berlin, 210 p.
Rivas, P. (1972). Estudio paleontológico-estratigráfico del Lias en el sector central de las Cordilleras Béticas. PhD thesis. Granada University. Spain.
Rivas, P. (1981). Zonación del Carixiense en la Zona Subbética: zonación. II Coloquio de Estratigrafía y Paleogeografía del Jurásico de España, Granada. Cuadernos de Geología, Universidad de Granada, 10, 383–388.
Robles, S., Quesada, S., Rosales, I., Aurell, M., & García-Ramos, J. C. (2004). El Jurásico marino de la Cordillera Cantábrica. In J. A. Vera (Ed.), Geología de España (pp. 279–285). Madrid: SGE-IGME.
Rodríguez-Luengo, E., Comas-Rengifo, M. J., & Goy, A. (2012). Caracterización de Matteiceras Wiedenmayer, 1980 (Ammonoidea, Hildoceratidae) en el Pliensbachiense superior de la cordillera Cantábrica. In J-C. Liao, J. A. Gámez Vintaned, J. I. Valenzuela-Ríos & A. García-Forner (Eds.), XXVIII Jornadas de la Sociedad Española de Paleontología, Libro de resúmenes (pp. 187–190). Universitat de Valencia.
Ruhl, M., Hesselbo, S. P., Hinnov, L., Jenkyns, H. C., Xu, W., Riding, J. B., Storm, M., Minisini, D., Ullmann, C. V., & Leng, M. J. (2016). Astronomical constraints on the duration of the early jurassic Pliensbachian Stage and global climatic fluctuations. Earth and Planetary Science Letters, 455, 149–165. https://doi.org/10.1016/j.epsl.2016.08.038.
Schlatter, R. (1977). The Biostratigraphy of the Lower Pliensbachian at the type locality (Pliensbach, Württemberg, SW Germany). Stuttgarter Beiträge zur Naturkunde, 27, 1–29.
Schlatter, R., & Württemberg, S. W. D. (1980).Stuttgarter Beiträge zur Naturkunde, Ser. B, 65,1–261.
Schlatter, R. (1991). Biostratigraphie und Ammonitenfauna des Ober-Lotharingium und Unter-Pliensbachium im Klettgau (Kanton Schaffhausen, Schweiz) und angrenzender Gebiete. Schweizerische Paläontologische Abhandlungen, 113, 1–133.
Schulz, G. (1858). Descripción geológica de la Provincia de Oviedo. Madrid: Ed. José González.
Sciau, J. (2004). Coup d’oeil sur les fossiles des causses. I Du Primaire au Lias moyen (p. 142). 2ème édition). Association Paléontologique des Causses. Millau.
Schöllhorn, I., Adatte, T., van de Schootbrugge, B., Houben, A., Charbonnier, G., Janssen, N., & Föllmi, K. B. (2020). Climate and environmental response to the break-up of Pangea during the early jurassic (Hettangian-Pliensbachian); the Dorset coast (UK) revisited. Global and Planetary Change, 185, 103096. https://doi.org/10.1016/j.gloplacha.2019.103096.
Silva, R. L., & Duarte, L. V. (2015). Organic matter production and preservation in the Lusitanian Basin (Portugal) and pliensbachian climatic hot snaps. Global and Planetary Change, 131, 24–34. https://doi.org/10.1016/j.gloplacha.2015.05.002.
Silva, R. L., Duarte, L. V., & Comas-Rengifo, M. J. (2015). Facies and carbon isotope chemostratigraphy of lower jurassic carbonate deposits, Lusitanian Basin (Portugal): Implications and limitations to the application in sequence stratigraphic studies. In M. Ramkumar (Ed.), Chemostratography. Concepts, techniques, and applications (pp. 341–371). Elsevier. https://doi.org/10.1016/B978-0-12-419968-2.00013-3.
Silva, R. L., Duarte, L. V., Wach, G. D., Ruhl, M., Sadki, D., Gómez, J. J., Hesselbo, S. P., Xu, W., O’Connor, D., Rodrigues, B., & Mendonça Filho, J. G. (2021). An early jurassic (Sinemurian–Toarcian) stratigraphic framework for the occurrence of Organic Matter Preservation Intervals (OMPIs). Earth-Science Reviews, 221, 103780. https://doi.org/10.1016/j.earscirev.2021.103780.
Spath, L. F. (1938). A catalogue of the ammonites of the Liassic family Liparoceratidae in the British Museum (Natural History). The British Museum (Natural History); London.
Stampfli, G. M., & Borel, G. D. (2002). A plate tectonic model for the Paleozoic and Mesozoic constrained by dynamic plate boundaries and restored synthetic oceanic isochrons. Earth and Planetary Science Letters, 196, 17–33. https://doi.org/10.1016/S0012-821X(01)00588-X.
Stampfli, G. M., & Borel, G. D. (2004). The TRANSMED transects in space and time: Constraints on the paleotectonic evolution of the Mediterranean domain. In W. Cavazza, F. M. Roure, W. Spakman, G. M. Stampfli, & P. A. Ziegler, P A. (Eds.), The TRANSMED Atlas—The Mediterranean Region from Crust to Mantle (pp. 53–80). Berlin Heidelberg New York: Springer. and CD-ROM10.1007/978-3-642-18919-7.
Su, C. P., Li, F., Tan, X. C., Gong, Q. L., Zeng, K., Tang, H., Li, M. L., & Wang, X. F. (2020). Recognition of diagenetic contribution to the formation of limestone-marl alternations: A case study from Permian of South China. Marine and Petroleum Geology, 111, 765–785. https://doi.org/10.1016/j.marpetgeo.2019.08.033.
Suárez Vega, L. C. (1974). Estratigrafía del Jurásico en Asturias. Cuadernos de Geología Ibérica, 3, 1–369.
Trueman, A. E. (1919). - the evolution of the Liparoceratidae. Quarterly Journal of the Geological Society of London, 74, 247–298.
Valenzuela, M., García-Ramos, J. C., & Suárez de Centi, C. (1986). The jurassic sedimentation in Asturias (N Spain). Trabajos de Geología, 16, 121–132.
Venturi, F., & Ferri, R. (2001). Ammoniti Liassici dell’Appennino Centrale. Città di Castello: Perugia: Ed. Tibergrah.
Venturi, F., Nannarone, C., & Bilotta, M. (2005). Early pliensbachian ammonites from the Furlo Pass (Marche, Italy): Two new faunas for the middle-western Tethys. Bollettino della Società Paleontológica Italiana, 44, 81–115.
Venturi, F., Nannarone, C., & Bilotta, M. (2007). Ammonites from the early Pliensbachian of the Furlo Pass (Marche, Italy): Biostratigraphic and paleobiogeographic implications. Bollettino della Società Paleontologica Italiana, 46, 1–31.
Vera, J. A. (2001). Evolution of the South Iberian Continental Margin. In: P. A. Ziegler, W. Cavazza, A. H. F. Robertson & S. Crasquin-Soleau (Eds.), Pery-Tethys Memoir 6: Peri-Tethyan Rift/Wrench Basins and Passive Margins. Mémoires du Muséum national d’histoire naturelle, 186, 109–143.
Westphal, H. (2006). Limestone-marl alternations as environmental archives and the role of early diagenesis: A critical review. International Journal of Earth Sciences, 95, 947–961. https://doi.org/10.1007/s00531-006-0084-8.
Westphal, H., Hilgen, F., & Munnecke, A. (2010). An assessment of the suitability of individual rhythmic carbonate successions for astrochronological application. Earth-Science Reviews, 99, 19–30. https://doi.org/10.1016/j.earscirev.2010.02.001.
Westphal, H., Munnecke, A., Böhm, F., & Bornholdt, S. (2008). Limestone-marl alternations in epeiric sea settings: Witnesses of environmental changes, or diagenesis? Geological Association of Canada Special Paper, 48, 389–406.
Wiedenmayer, F. (1980). Die Ammoniten der mediterranen Provinz in Pliensbachian und unteren toarcian aufgrund neuer Untersuchungen im Generoso-Becken (Lombardische Alpen). Mémoires de la Société Helvétique des Sciences Naturelles, 93, 1–197.
Wilmsen, M., Blau, J., Meister, C., Mehdi, M., & Neuweiler, F. (2002). Early jurassic (sinemurian to Toarcian) ammonites from the Central Hight Atlas (Morocco) between Er-Rachidia and Rich. Revue de Paléobiologie, 21, 149–175.
Ziegler, P. A. (1990). Geological Atlas of Western and Central Europe. Shell International Petroleum. Maatschappi B.V. Geological Society Publishing House, Bath, pp. 1–239.
Acknowledgements
This work is a contribution to the project CGL2015-66604-R of the Spanish Ministry of Economy and Competitiveness, and to Research Group 910431 “Mesozoic Biotic Processes” of the Complutense University of Madrid. We are grateful to Carlos Alonso Recio and Gema Martín (Palaeontology Area of the Geological Science Faculty) for their excellent photographic works, to Cormac de Brun and Íñigo Vitón for reviewing the English version, to Dr. Juan Carlos Braga, Associate Editor, and two anonymous reviewers for their helpful comments that much improved the original version of this manuscript.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. Financial support for L. Piñuela and J.C. García-Ramos was provided by Sociedad Pública de Gestión y Promoción Turística y Cultural del Principado de Asturias (Board of Tourism and Culture Management and Promotion of the Principality of Asturias).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
We declare that this manuscript is original, has not been published before and is not currently being considered for publication elsewhere.
Conflict of interest
We know of no conflicts of interest associated with this publication, and there has been no significant financial support for this work that could have influenced its outcome. As Corresponding Author, I confirm that the manuscript has been read and approved for submission by all the named authors.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Comas-Rengifo, M., García-Ramos, J., Goy, A. et al. Stratigraphy and Biochronostratigraphy of the Lower Pliensbachian (Jurassic) from the Asturian basin (Northern Spain). J Iber Geol 49, 73–96 (2023). https://doi.org/10.1007/s41513-023-00209-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41513-023-00209-7