
Abstract
Apricot aphid, Myzus mumecola (Matsumura) (Homoptera: Aphididae), was recorded as a new pest of apricot in Hungary in the spring of 2020. Identification was based on morphological and genetical (mitochondrial COI region) characteristics. M. mumecola most likely arrived in Hungary in the last 2–3 years and has quickly become a widespread pest causing significant damage to young apricot trees. Colony development, damage and differences in susceptibility between cultivars are described. The presence of Plum pox virus in M. mumecola samples was detected, and all isolates belonged to the PPV-D subgroup. Illustrations of the most important diagnostic characters of M. mumecola are provided.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Apricot (Prunus armeniaca L.) is an economically important crop in Europe, especially in the Mediterranean Region (FAOSTAT 2020). Although the presence of several aphid species was recorded in apricot orchards in Europe (Avinent et al. 1993; Mosco et al. 1997; Holman 2009Jeremovic et al. 2016), their most significant economic impact was not their direct damage caused on the yield, but the transmission of Plum pox potyvirus (PPV) (Avinent et al. 1993; Labonne et al. 1994; García et al. 2014). The most common aphid species landing on the foliage of apricot trees in Spain were Aphis gossypii Glover and A. spiraecola (Avinent et al. 1993). On the other hand, from the West Palearctic Region, the Hyalopterus pruni species complex, Myzus persicae Sulzer and Rhopalosiphum nymphaeae L. have been most commonly reported to form large colonies on apricot trees (Blackman and Eastop, 1984; Mosco et al. 1997; Lozier et al. 2008; Rakauskas et al. 2013). In Italy, Panini et al. (2017) recorded the appearance and rapid spread of an aphid species on apricot, which was identified as Myzus mumecola (Hemiptera: Aphididae), a new aphid species for Europe. Myzus mumecola originates from Eastern Asia where its main host plant is Prunus mume Siebold (Blackman and Eastop, 1984). The species has been recorded in Japan on P. ansu Maxim, P. armeniaca and P. mume (Takahashi, 1965; Miyazaki, 1971), in Taiwan (Mondor et al. 2007; Lai 2020) on Prunus sp., in China on P. armeniaca (Zhang et al. 1985), in East Russia on Prunus sp. (Pashchenko 1988), in India on Prunus cornuta Steudel (Basu and Raychaudhuri, 1976) and in the Himalayas on P. armeniaca and P. mume (Chakrabarti et al. 1970; Chakrabarti and Sarkar 2001). Based on the laboratory tests of Kimura et al. (2016), M. mumecola has the ability to transmit Plum pox virus (PPV) on P. mume, albeit it does not belong to the abundant aphid species in the P. mume orchards in Japan.
Materials and methods
Morphological identification
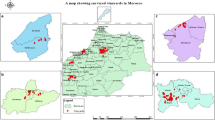
Aphid samples were collected from apricot trees (P. armeniaca) of different ages at six different locations in Hungary in April and May 2020: Győr (lat. 47.699717 N; long. 17.748146 E) and Győrszentiván (lat. 47.676328 N; long. 17.662642 E) in Western Transdanubia, Balatonalmádi (lat. 47.024892 N; long. 18.008070 E) in Central Transdanubia, Budapest Budafok (lat. 47.422926 N; long. 19.032137 E) and Pomáz (lat. 47.652329 N; long. 19.012668 E) in Central Hungary, and Gönc (lat. 48.468253 N; long. 21.282107 E) in Northern Hungary (Supplementary Fig. 1). The studied home garden trees and orchards received no pesticides. The aphid individuals were collected in 1.5-ml Eppendorf tubes with fine painting brush and were preserved in 70 V/V% ethanol for morphological and in 98 V/V % ethanol for molecular identification. From the collected aphids, 12–48 individuals per site were selected for identification. The aphid specimens were mounted after a 15-min soaking in hot (~ 95 °C) 20% KOH solution. The body content was dissolved by distilled water, and the clean cuticle of the aphids was fixed on microscope slides in glycerine jelly. The morphological identification of the aphid samples was performed by stereo- (Leica MZ6) and light (Euromex iScope 1153-PLi) microscopy, using the keys of Basu and Raychaudhuri (1976) and Blackman and Eastop (1984).
Molecular identification
The morphological identification was supported by the sequence analysis of the DNA mitochondrial cytochrome c oxidase subunit 1 gene (COI fragment) barcode (Hebert et al. 2003; Panini et al. 2017). Six aphid individuals (one specimen per site) were sequenced from the samples collected for morphological identification. Total genomic DNA was extracted from a single aphid with DNeasy blood and tissue kit (QIAGEN, Hilden, Germany) according to the manufacturer’s instruction. Amplification of the COI barcode was performed in 20 μl reaction volume containing 20–80 ng DNA, 5 × Phire Reaction Buffer, 0.4 μl of Phire Hot Start II DNA Polymerase (Thermo Scientific, Hungary), 0.2 mM dNTP mix, 3% DMSO, 2.5 μmol of each 5′ and 3′ end primers (LCO 1490: 5′ GGTCAACAAATCATAAAGATATTGG 3′ and HCO 2198: 5′ TAAACTTCAGGGTGACCAAAAAATCA 3′ (Folmer et al. 1994)) and sterile distilled water. PCR was carried out in a Swift MaxPro thermocycler (ESCO Healthcare, Singapore). The cycling parameters were as follows: initial denaturation at 98 °C for 30 s, then 30 cycles of denaturation at 98 °C for 5 s, annealing at 49 °C for 5 s, extension at 72 °C for 15 s and a final extension for 1 min at 72 °C. Amplification was verified on a 1% (w/v) ethidium bromide‐stained agarose gel in 1 × TBE buffer. Fragment size was estimated by comparison with the 1 kb DNA ladder (Fermentas, Waltham, MA, USA). The amplified fragments were purified using ExoSAP-IT Express purification kit (Thermo Scientific, Hungary) for direct sequencing. Sequencing was performed in an automated sequencer ABI PRISM 3100 Genetic Analyser (Applied Biosystems, Foster City, CA, USA). For each fragment, the nucleotide sequences were determined in both directions. Forward and reverse sequences were edited and assembled; alignment and neighbour-joining analysis were conducted in MEGAX (Kumar et al. 2018). DNA sequences were verified using the BLASTN algorithm at NCBI.
Life cycle, damage and susceptibility
The development of the M. mumecola colonies was followed on individual trees in home gardens to assess the presence of different stages and the damage caused on the foliage of apricot trees. Susceptibility of different apricot cultivars to M. mumecola was assessed in five apricot orchards in Hungary in July 2020 (Supplementary Fig. 1, Supplementary Table 1): Orchard 1, Győrszentiván (lat. 47.686939 N, long. 17.793228 E; cultivars ‘Bergeron’, ‘Gönci magyar kajszi’ and ‘Magyar kajszi C235’); Orchard 2, Szentkirályszabadja (lat. 47.070796 N, long. 17.939379 E; cultivars ‘Spring Blush’ and ‘Magyar kajszi C235’); Orchard 3, Érd-Elviramajor (lat. 47.336696 N, long. 18.864777 E; cultivars ‘Summerland’ and mixed seedlings from Greece; Orchard 4, Törökbálint (lat. 47.465428 N, long. 18.863759 E; cultivars ‘Orange Red’ and ‘Magyar kajszi C235’), and Orchard 5 Budapest-Soroksár (lat. 47.395810 N, long. 19.148472 E; cultivars ‘Goldrich’, ‘Orange Red’, ‘Budapest’, ‘Ceglédi bíbor’, ‘Ceglédi kedves’, ‘Korai zamatos’ and ‘Pannónia’). All orchards were planted on cherry plum (Prunus cerasifera Ehrhart) rootstock. Cultivars within each orchard received the same horticultural practices and pest management (Supplementary Table 1). Number of shoots and number of M. mumecola-infested shoots were counted for each sampled tree. The extent of the damage was expressed as percentage of infested shoots on randomly selected trees (n ≥ 9 trees per cultivar) (Supplementary Table 1). Due to the non-normal distribution of the data and lack of homogeneity of variance, for intergroup comparisons the Brunner–Munzel test (two cultivars) or the adjusted rank Welch test followed by Brunner–Munzel test with Bonferroni adjustment (more than two cultivars) were used. As cultivars were grown in non-randomised plots (spatial pseudoreplication), all statistical comparisons were tested at the significance level of 0.0001 and 0.05. Statistical analyses were performed with the software package ROPstat (Vargha et al. 2015).
Detection of PPV viral RNA
For detection of the presence of PPV in the aphids collected from apricot trees at the six aphid collection sites described above (Supplementary Fig. 1), six aphid individuals (one specimen per sampling site) were tested. After releasing the viral RNA template from a single aphid with boiling technique (Kim et al., 2016), a two-step RT-PCR was carried out for the detection of PPV with a PPV-specific primer pair (PP3: 5′ TTATCTCCAGGA(AG)TTGGAGC 3′ and PCI: 5′ TTGAGTCAAATGG(AG)ACAGTTGG 3′(Glasa et al. 2002)) amplifying a 836-bp-long fragment corresponding to the 5′-terminal region of the P3 gene (580 nt), the complete 6K1 (156 nt) and a 100 nt portion from the 3′-terminal part of the CI gene [3′P3–6K1–5′CI]. Amplification was performed with Phire Hot Start (Thermo Scientific, Hungary) polymerase as described above. The cycling parameters were as follows: initial denaturation at 98 °C for 30 s, then 30 cycles of denaturation at 98 °C for 5 s, annealing at 55 °C for 5 s, extension at 72 °C for 15 s and a final extension for 1 min at 72 °C. Amplification was verified on a 1% (w/v) ethidium bromide‐stained agarose gel in 1 × TBE buffer. Fragment size was estimated by comparison with the 1 kb DNA ladder (Fermentas, Waltham, MA, USA).
For PPV subgroup typing, aliquots of the PCI/PP3 PCR products were subjected to CAPS analysis with restriction endonucleases DdeI, EcoRI and EcoRV (Thermo Scientific, Hungary), as suggested by the manufacturer. Based on the resulting digestion pattern, the three most common strains in Hungary (PPV-D, PPV-M, PPV-Rec) can be differentiated (Glasa et al. 2002; Ádám, et al. 2015).
Results
Morphological identification
We confirmed the presence of M. mumecola in Hungary by morphological characteristics. Myzus mumecola appears to be widespread in Hungary, as we could detect it in all of the examined regions of Hungary: Győr (25 May 2020; number of identified specimens: 48) and Győrszentiván (23 April 2020; 24 specimens) in Western Transdanubia, Balatonalmádi (16 May 2020; 24 specimens) in Central Transdanubia, Budafok (2 May 2020; 12 specimens) and Pomáz (25 May 2020; 12 specimens) in Central Hungary, and Gönc (19 May 2020; 12 specimens) in Northern Hungary. The slide-mounted specimens were deposited in the Hungarian Natural History Museum, Budapest. These are the first records of M. mumecola from Central Europe. We propose the common name ‘apricot aphid’ for M. mumecola, referring to the host plant, apricot.
Morphological description of Myzus mumecola
Apterous (wingless) M. mumecola individuals in life are medium-sized (2–2.4 mm) (Panini et al. 2017), pale green in colour with pale cauda and siphunculi. The cauda is short and triangular, and the siphunculi are cylindrical, about 7 times longer than their width (Fig. 1a) (Panini et al. 2017). On the mounted specimens, key morphological characters of M. mumecola are clearly discernible: M. mumecola differs from Phorodon humuli Schrank in the lack of the finger-like processes on the antennal tubercles (Fig. 1b), while from Myzus persicae Sulzer in the divergent antennal tubercles (Fig. 1b), longer hairs of the antennae (Fig. 1c), and the narrowing shape of the siphunculi (Fig. 1d) (Basu and Raychaudhuri 1976; Blackman and Eastop 1984). Other aphid species recorded from apricot differ significantly from M. mumecola in size or colour (Blackman and Eastop 1984).

Morphological characteristics of apterae M. mumecola (a: overview; b: antennal tubercles, c: hairs on antennal segments IV and V, d: siphunculus)
Molecular identification
Identification of M. mumecola was also confirmed by DNA sequence analysis. The COI barcode of six M. mumecola individuals (one specimen per site) was sequenced from the samples collected at six locations in Hungary (Győrszentiván, Győr, Balatonalmádi, Budapest Budafok, Pomáz, Gönc): the sequenced region covered 709 nucleotides. Forward and reverse sequences were aligned, and a consensus sequence was generated for each. All sequences are deposited in the NCBI GenBank under the accession number MT635054-MT635059. For sequence comparison, further M. mumecola (AB738876 and AB738877) and M. persicae (JF883917, HQ971262, GU668755, KR032469, KR039431, KY323048, KP759548, MN320357) COI sequences have been retrieved from NCBI GenBank. For NJ analysis, the sequences were trimmed; hence, finally 644 bases were used in the alignment and cluster analysis. The sequences both for M. mumecola and for M. persicae were uniform within the species, while comparing M. mumecola and M. persicae sequences 42 SNPs were found differentiating the two species. The neighbour-joining analysis (Fig. 2) revealed two independent clades with strong support (100%), one harbouring the M. mumecola individuals and the other the M. persicae individuals.

Neighbour-joining (NJ) tree showing phylogenetic relationships among Myzus mumecola and M. persicae based on mitochondrial COI barcode (644 positions in final data set). The analysis involved 16 nucleotide sequences (six sequences resulting from this study and ten retrieved from the NCBI GenBank database). The evolutionary distances were computed using the maximum composite likelihood method and are in the units of the number of base substitutions per site
Life cycle, damage and susceptibility
Fundatrices were observed on the leaves as early as in the initial stages of leaf development, indicating that M. mumecola probably overwinters as eggs on apricot trees. The following 2–3 apterous generations were found to feed on the backside of the leaves (Fig. 3a) and caused significant malformation on young leaves: developing leaves were distorted, severely curled downwards (perpendicularly to the midrib) forming pseudo-galls as reported by Panini et al. (2017) (Fig. 3b). The apterae of the later spring generations were actively moving on the trees, colonising the tips of the growing shoots in the surrounding of the initial colonies. The growth of the infested shoots was affected, and the shoots became slightly distorted (Fig. 3c). In some cases, we observed delayed flowering (in mid-April) and fructification of the infested shoots. In some cases, aphid individuals were found feeding on the growing fruits. The infested leaves partially discoloured and fell prematurely within a couple of weeks. New shoot growth appeared at the base of the dead shoot tips in early June. The first alatae (winged) forms appeared in mid-May in the pseudo-galls (Fig. 3c), indicating that M. mumecola follows a heterocyclic life cycle in Hungary, although its summer host is still unknown (Panini et al. 2017). Alatae feeding on the winter host (apricot) were found by mid-June.

Damage of M. mumecola on apricot (a M. mumecola colony on the backside of a leaf; b malformed leaves on a growing shoot; c shoots disrupted in development with alateae forms)
In the M. mumecola colonies, ants (Lasius niger L.) and predatory insects (primarily Harmonia axyridis Pallas, Forficula auricularia L. and Episyrphus balteatus De Geer) were observed in high numbers.
The susceptibility of different apricot cultivars to the aphid infestation differed significantly (Table 1). Three susceptibility levels were distinguishable: aphid-tolerant cultivars (0–4% of the shoots were infected) were Greek mixed seedlings, ‘Budapest’, ‘Ceglédi bíbor’, ‘Magyar kajszi C235’, ‘Orange Red’, ‘Ceglédi kedves’ and ‘Pannónia’; moderately sensitive cultivars (10–21%) were ‘Korai zamatos’ ‘Gönci magyar’, ‘Summerland’ and ‘Bergeron’; and the sensitive cultivars (30–34%) were ‘Goldrich’ and ‘Spring Blush’. The aphid infestation on the sensitive cultivars and in most comparisons on the moderately sensitive cultivars differed significantly from that on the cultivars in the aphid-tolerant group (Table 1). There was no significant difference in the number of shoots between most of the cultivars (Supplementary Table 2).
Detection of PPV viral RNA
The RT-PCR carried out with the PP3/PCI PPV-specific primer pair amplified fragments of the expected size (around 836 bp) in the case of all six M. mumecola individuals from six locations of Hungary (Győrszentiván, Győr, Balatonalmádi, Budafok, Pomáz and Gönc). The PCR products could be cleaved by both DdeI and EcoRI, but no restriction site of EcoRV was present in the sequences. Thus, all six PPV isolates detected in aphids belonged to the PPV-D subgroup, and no other strains of PPV from the most common types in Hungary (PPV-D, PPV-M, PPV-Rec) could have been identified.
Discussion
In the past few years, Myzus mumecola became an important pest of apricot in Italy (Panini et al. 2017) and, as we found, in Hungary as well. Its rapid expansion across the apricot-growing regions of Europe is highly probable. Since M. mumecola were not recorded in an extensive faunal survey of the apricot orchards in West Serbia between 2009 and 2011 (Jevremović et al. 2016), the new species had presumably been established in Central Europe in the past few years. As Northern Hungary is one of the northernmost apricot-growing regions of Europe (Bassi 1999; FAOSTAT 2020), it is less likely that continental climate can limit the distribution of the new species (Kaneko 1993). Aphids had been generally uncommon pest of apricot in Central Europe, and therefore insecticide sprays were not required for aphid control (Pénzes et al. 2003). As M. mumecola is now a significant pest of apricot in Central Europe, deeper understanding of its life cycle (including the range of summer hosts and the vector role in the virus and phytoplasma dissemination) and development of new effective control strategies are required.
The morphological characteristics of M. mumecola specimens examined by us matched exactly with the description of Panini et al (2017) and the morphological keys of Takahashi (1965), Basu and Raychaudhuri (1976) and Blackman and Eastop (1984) (Fig. 1). The identity of the species was also confirmed by the COI barcode sequence analysis (Fig. 2), which fully matched with those of the Italian and Japanese samples of M. mumecola (AB738876 and AB738877) proving the reliability of the morphological descriptions in the distinction of the species from other aphid pests of apricot.
The 13 apricot cultivars evaluated varied considerably in their susceptibility to M. mumecola. While most of the cultivars showed low or moderate aphid incidence, ‘Goldrich’ and ‘Spring Blush’ appeared to be particularly susceptible (Table 1). The abundance of aphids on fruit trees is primarily limited by the intensity of shoot growing, quality and quantity of plant soap, and the presence of natural enemies (Dixon, 1973). The susceptibility of different plant cultivars depends on several physiological and genetical factors (Dogimont et al. 2010; Pierson et al. 2011). Some cultivars compensate for the loss caused by aphids (Pierson et al. 2011) or restrain the feeding of aphids in a physical way (Giordanengo et al. 2010), while in other cases the expression of different resistance genes leads to some level of tolerance (Dogimont elt al. 2010). In the case of peach (Prunus persica L.), Pascal et al. (2002) found a single dominant resistance gene in the red leaf rootstock cultivar ‘Rubira’, which is responsible for induced resistance against M. persicae (Sauge et al. 2002). Aphid resistance genes of apricot have not been identified yet.
Our findings, obtained with CAPS analysis of the PP3/PCI amplified PCR product which is used for subgroup typing of PPV isolates (Glasa et al. 2002; Ádám et al. 2015), are in accordance with previously published results where the most prevalent strain of PPV in apricot in Hungary was PPV-D (Sihelská et al. 2017). Although M. mumecola has lower transmission rate (12%) than M. persicae (24.4%) (Kimura et al. 2016), it has the potential to be an important vector of PPV because of its high abundance and high rate of aphids carrying PPV.
References
Avinent L, Hermoso de Mendoza A, Llacer G (1993) Comparison of sampling methods to evaluate aphid populations (Homoptera, Aphidinea) alighting on apricot trees. Agronomie EDP Sci 13:609–613
Ádám J, Palkovics L, Tóbiás I, Almási A (2015) Presence of sharka disease in the North-Hungarian counties. Acta Hortic 1063:55–60. https://doi.org/10.17660/ActaHortic.2015.1063.6
Bassi D (1999) Apricot culture: present and future. Acta Hortic 488:35–40. https://doi.org/10.17660/ActaHortic.1999.488.1
Basu RC, Raychaudhuri DN (1976) Studies on the aphids (Homoptera: Aphididae) from India XXV. The genus Myzus with five new species from Eastern India. Orient Insects 10:93–111. https://doi.org/10.1080/00305316.1976.10432325
Blackman RL, Eastop, VF (1984) Aphids on the world's crops. Chichester, UK, pp 40–42
Chakrabarti S, Gosh AK, Chowdhuri AN (1970) Aphids (Homoptera) of Himachal Pradesh, India-III. Orient Insects 4:447–451. https://doi.org/10.1080/00305316.1970.10433982
Chakrabarti S, Sarkar A (2001) A supplement to the food-plant catalogue of Indian Aphididae (Homoptera). J Aphidol 15:9–62
Dixon AFG (1973) Biology of aphids. Edward Arnold, London
Dogimont C, Bendahmane A, Chovelon V, Boissot N (2010) Host plant resistance to aphids in cultivated crops: genetic and molecular bases, and interactions with aphid populations. CR Biol 333:566–573. https://doi.org/10.1016/j.crvi.2010.04.003
FAOSTAT (Food and Agriculture Organization of the United Nations) (2020) Online database. http://www.fao.org/faostat/en/#data/QC. Accessed 17 June 2020
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotech 3:294–299
García JA, Glasa M, Cambra M, Candresse T (2014) Plum pox virus and sharka: a model potyvirus and a major disease. Mol Plant Pathol 15:226–241. https://doi.org/10.1111/mpp.12083
Giordanengo P, Brunissen L, Rusterucci C, Vincent C, van Bel A, Dinant S, Girousse C, Faucher M, Bonnemain JL (2010) Compatible plant-aphid interactions: how aphids manipulate plant responses. CR Biol 333:516–523. https://doi.org/10.1016/j.crvi.2010.03.007
Glasa M, Marie-Jeanne V, Moury B, Kúdela O, Quiot JB (2002) Molecular variability of the P3–6K1 genomic region among geographically and biologically distinct isolates of Plum pox virus. Adv Virol 147:563–575
Hebert PDN, Ratnasingham S, deWaard JR (2003) Barcoding animal life: cytochrome c oxidase subunit I divergences among closely related species. Proc R Soc Lond B 270:96–99
Holman J (2009) Host plant catalog of aphids. Palaearctic Region. Springer, Berlin
Jevremović D, Paunović SA, Petrović-Obradovic O (2016) Flight dynamics and species composition of aphids landing on plum and apricot leaves in the orchards in Western Serbia. Phytoparasitica 44:501–511. https://doi.org/10.1007/s12600-016-0544-z
Kaneko J (1993) Change in supercooling point of eggs of aphid, Myzus mumecola (Matsumura), through overwintering period. Ann Rep Plant Prot North Jpn (in Japanese, Abstract and Figures in English) 44:167–168
Kim J, Cha DJ, Kwon M, Maharjan R (2016) Potato virus Y (PVY) detection in a single aphid by one-step RT-PCR with boiling technique. Entomol Res 46:278–285. https://doi.org/10.1111/1748-5967.12170
Kimura K, Usugi T, Hoshi H, Kato A, Ono T, Koyano S, Kagiwada S, Nishio T, Tsuda T (2016) Surveys of viruliferous alate aphid of Plum pox virus in Prunus mume orchards in Japan. Plant Dis 100:40–48. https://doi.org/10.1094/PDIS-05-15-0540-RE
Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol 35:1547–1549. https://doi.org/10.1093/molbev/msy096
Labonne G, Lauriaut F, Yvon M, Quiot JB (1994) Aphid transmission of Plum pox potyvirus: analysis of potential vectors in an apricot orchard. Bull. OEPP 24:681–690. https://doi.org/10.1111/j.1365-2338.1994.tb01083.x
Lai K (2020) The National Checklist of Taiwan. Version 1.4. Taiwan Biodiversity Information Facility (TaiBIF). Checklist dataset https://doi.org/https://doi.org/10.15468/auw1kd. Accessed 12 July 2020
Lozier JD, Foottit RG, Miller GL, Mills NJ, Roderick GK (2008) Molecular and morphological evaluation of the aphid genus Hyalopterus Koch (Insecta: Hemiptera: Aphididae), with a description of a new species. Zootaxa 1688:1–19. https://doi.org/10.11646/zootaxa.1688.1.1
Miyazaki M (1971) A revision of the tribe Macrosiphini of Japan (Homoptera: Aphididae, Aphidinae). Insecta Matsumurana 34:1–247
Mondor EB, Tremblay MN, Messing RH (2007) Morphological and ecological traits promoting aphid colonization of the Hawaiian Islands. Biol Invasions 9:87–100. https://doi.org/10.1007/s10530-006-9010-z
Mosco MC, Arduino P, Bullini L, Barbagallo S (1997) Genetic heterogeneity, reproductive isolation and host preferences in mealy aphids of the Hyalopterus pruni complex (Homoptera, Aphidoidea). Mol Ecol 6:667–670. https://doi.org/10.1046/j.1365-294X.1997.00234.x
Panini M, Massimino Cocuzza G, Dradi D, Chiesa O, Mazzoni E (2017) First report of Myzus mumecola (Matsumura, 1917) in Europe. EPPO Bull 47:107–110. https://doi.org/10.1111/epp.12357
Pascal T, Pfeiffer F, Kervella J, Lacroze JP, Sauge MH, Weber WE (2002) Inheritance of green peach aphid resistance in the peach cultivar ‘Rubira.’ Plant Breed 121:459–461. https://doi.org/10.1046/j.1439-0523.2002.734333.x
Pashhchenko NF (1988) Suborder Aphidinea, aphids. In: Lehr PA (ed) Keys to insects of the Far East of the USSR, 2, Nauka, Leningrad (in Russian)
Pénzes B, Glits M, Süle S, Németh M, (2003) A kajszi növényvédelme. In: Pénzes B, Szalay L (eds) Kajszi (Apricot), 1st edn. Mezőgazda Kiadó, Budapest, pp 280–334 (in Hungarian)
Pierson LM, Heng-Moss TM, Hunt TE, Reese J (2011) Physiological responses of resistant and susceptible reproductive stage soybean to soybean aphid (Aphis glycines Matsumura) feeding. Arthropod–Plant Interact 5:49–58. https://doi.org/10.1007/s11829-010-9115-2
Rakauskas R, Havelka J, Zaremba A (2013) Mitochondrial COI and morphological specificity of the mealy aphids (Hyalopterus ssp.) collected from different hosts in Europe (Hemiptera, Aphididae). ZooKeys 319:255–267. https://doi.org/10.3897/zookeys.319.4251
Sauge MH, Lacroze JP, Poëssel JL, Pascal T, Kervella J (2002) Induced resistance by Myzus persicae in the peach cultivar ‘Rubira.’ Entomol Exp Appl 102:29–37. https://doi.org/10.1046/j.1570-7458.2002.00922.x
Sihelská N, Glasa M, Šubr ZW (2017) Host preference of the major strains of Plum pox virus–Opinions based on regional and world-wide sequence data. J Integrat Agric 16:510–515. https://doi.org/10.1016/S2095-3119(16)61356-4
Takahashi R (1965) Myzus of Japan (Aphididae). Mushi 38:43–78
Vargha A, Torma B, Bergman LR (2015) ROPstat: a general statistical package useful for conducting person-oriented analyses. J Person-Orient Res 1:87–98. https://doi.org/10.17505/jpor.2015.09
Zhang G, Liu L, He F, Zhong T (1985) Six aphid species newly recorded from China. Acta Entomol Sin 28:287
Acknowledgements
We would like to thank the growers and garden owners, György Bedey, Valéria Elszeszer, Petra Enzsöl, Katalin Fazekas, Péter Kovács and Csilla Molnár, for their cooperation and help in sample collection.
Funding
This research was supported by the National Research Development and Innovation Office of Hungary (K 129311) and by the Ministry for Innovation and Technology within the framework of the Thematic Excellence Programme 2020- Institutional Excellence Subprogram (TKP2020-IKA-12) for research on plant breeding and plant protection. Open access funding provided by Szent István University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Borbély, C., György, Z., Szathmáry, E. et al. Apricot aphid, Myzus mumecola (Matsumura), a new and important pest of apricot in Hungary. J Plant Dis Prot 128, 781–787 (2021). https://doi.org/10.1007/s41348-021-00436-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41348-021-00436-z