
Abstract
The study of marine soundscapes is a growing field of research. Recording hardware is becoming more accessible; there are a number of off-the-shelf autonomous recorders that can be deployed for months at a time; software analysis tools exist as shareware; raw or preprocessed recordings are freely and publicly available. However, what is missing are catalogues of commonly recorded sounds. Sounds related to geophysical events (e.g. earthquakes) and weather (e.g. wind and precipitation), to human activities (e.g. ships) and to marine animals (e.g. crustaceans, fish and marine mammals) commonly occur. Marine mammals are distributed throughout Australia’s oceans and significantly contribute to the underwater soundscape. However, due to a lack of concurrent visual and passive acoustic observations, it is often not known which species produces which sounds. To aid in the analysis of Australian and Antarctic marine soundscape recordings, a literature review of the sounds made by marine mammals was undertaken. Frequency, duration and source level measurements are summarised and tabulated. In addition to the literature review, new marine mammal data are presented and include recordings from Australia of Omura’s whales (Balaenoptera omurai), dwarf sperm whales (Kogia sima), common dolphins (Delphinus delphis), short-finned pilot whales (Globicephala macrorhynchus), long-finned pilot whales (G. melas), Fraser’s dolphins (Lagenodelphis hosei), false killer whales (Pseudorca crassidens), striped dolphins (Stenella coeruleoalba) and spinner dolphins (S. longirostris), as well as the whistles and burst-pulse sounds of Australian pygmy killer whales (Feresa attenuata). To date, this is the most comprehensive acoustic summary for marine mammal species in Australian waters.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Marine mammals have evolved to use sound as their primary sensory modality—both actively (sound production) and passively (sound reception). That is because sound travels much better (i.e. with less loss and over longer ranges) underwater than does light. Sound also travels faster and over longer ranges underwater than it does in air. Marine mammals produce sound prolifically for communication [1]. Odontocetes (toothed whales) also emit echolocation clicks during foraging and navigation [2]. While marine mammal vocalisations are produced in the larynx, and echolocation clicks in nasal air sacs, some marine mammals also produce mechanical sounds during behaviours at the surface of the water such as breaching or fluke slapping [3].
Recording and listening to these sounds for investigating the presence, distribution, migration, relative abundance and behaviour of marine mammals has proven to be a powerful tool. While traditional visual detection methods of surveying marine mammals continue to be relevant for many applications, passive acoustic methods are increasingly employed due to their high success rates independent of time of day and weather, and their cost-effectiveness in remote, inaccessible locations where long-term data sets are required [4]. Such long data sets are becoming publicly available. For example, there are many years of recordings from Australia’s Integrated Marine Observing System’s (IMOS) passive acoustic stations located off the WA, SA, VIC and NSW coasts. All of the data can be downloaded or requested from the Australian Ocean Data Network (http://imos.org.au/aodn.html).
Underpinning all applications of passive acoustics is the ability to attribute recorded sounds to their sources, including marine mammals. Few Australian studies have comprised simultaneous visual and passive acoustic surveys, compared to studies off North America or Europe. While the underwater biophony in Australia is rich and complex, specifically in Australia’s tropical and subtropical regions (e.g. [5, 6]), we often do not know what species make what sounds. This is a major problem in particular for marine environmental impact assessments. Australia’s waters and continental shelf are rich in resources ranging from minerals and hydrocarbons to commercial fish. Industries such as fisheries, tourism, oil and gas exploration and production, mining, construction and defence abide by environmental health and safety guidelines which may include underwater soundscape analyses, marine mammal surveying and noise impact assessments.
To aid these studies and assessments, we present a summary of sounds emitted by marine mammals occurring in both inshore and offshore waters around the Australian continent and the Australian Antarctic Territory. Example sound files are available at http://cmst.curtin.edu.au/research/marine-mammal-bioacoustics/. Our article is not intended to be a complete summary of all of the literature published on the sounds made by these species, but rather a guide to the literature that best describes the species-specific sound characteristics. We envisage this overview to be used by readers tasked with analysing passive acoustic recordings from Australia.
Methods
A list of Australian marine mammal species and information on each species’ physical appearance, distribution, ecology and threats can be found on the website of the Australian Government Department of the Environment and Energy (http://www.environment.gov.au/marine/marine-species). Based on this list of species occurring in Australia, a literature search was undertaken of sounds recorded worldwide from each species. While some species have been shown to emit slightly different sounds in different geographic regions, information on the sounds of Australian mammals is typically so limited, that recordings from outside of Australia were included to give a broad overview of the types of sounds these animals emit. Articles that reported simultaneous visual and acoustic detections, rather than articles from autonomous passive acoustic recorders in the absence of visual observers, were preferred. The latter were considered only in cases where the sound-producing species could be identified without doubt based on the sound characteristics and where the study added new information important for passive acoustic monitoring (e.g. source levels). We further focussed on articles that described the spectrographic characteristics of sounds, showed example spectrograms and presented measurements such as bandwidth, duration and source level. In addition to the literature review, we provide spectrograms of vocalisations recorded in Australian waters and tables summarising their spectrographic features.
This article is organised by species, using the taxonomy of the Society for Marine Mammalogy (https://www.marinemammalscience.org/species-information/list-marine-mammal-species-subspecies/). Species distribution within Australia and the Commonwealth marine area is based on the maps and information available at the Department of the Environment and Energy website (http://www.environment.gov.au/cgi-bin/sprat/public/mapcaveat.pl), augmented with geographically farther-reaching information from maps on the Convention on the Conservation of Migratory Species of Wild Animals website (http://www.cms.int) and the Society for Marine Mammalogy website.
Marine Mammal Species of Australia
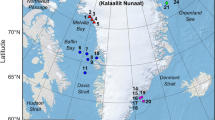
A total of 57 marine mammal species may be found around Australia, including 10 mysticetes (baleen whales), 36 odontocetes (toothed whales), one sirenian (dugong) and 10 pinnipeds (seals and sea lions). These species are listed in Table 1, also indicating whether they are most commonly encountered inshore versus offshore. Many of the species undertake annual migrations. For example, polar feeding mysticetes migrate between their polar feeding grounds in the summer and temperate to tropical breeding grounds in the winter. The “dwarf” mysticete species follow a similar annual migration pattern but shifted towards the equator, with most not reaching polar regions. Some odontocetes migrate, others do not and instead shift their distribution on more local scales typically driven by prey. Some of the pinnipeds stay within the vicinity of their breeding grounds, while others travel over extended distances for food. Indicative ranges of species distribution are given in Fig. 1.
Types of Sounds
In the literature, animal sounds are sometimes given onomatopoeic names (phonetically imitating the sound) and grouped based on what they sound like to the human ear. The same sounds are often given different names by different authors. In this article, we group sounds based on their spectrographic features. However, these can depend on some of the recording and analysis settings (e.g. sampling frequency, filters and analysis window used during Fourier transform). A series of broadband pulses is sometimes called a bout of pulses in the literature, other times an amplitude-modulated (AM) sound, and, if the analysis window is longer than the pulses, the sound looks tonal in spectrograms. To make this even more interesting, some animals are able to produce sounds along a continuum from tonal to pulsed [7,8,9]. As we did not have access to the raw data underlying the various published spectrograms, we based our grouping on published spectral features.
All sounds were grouped into three classes based on their spectrographic features: 1) constant-wave (CW) tones, 2) frequency-modulated (FM) sounds and 3) broadband pulses. CW sounds appear as straight, horizontal lines in spectrograms. FM sounds include upsweeps, downsweeps or sinusoidal contours in spectrograms. Both CW and FM sounds may have harmonic overtones. Pulses are broadband and lack tonal characteristics.
Mysticetes make sounds of all three categories. Pulses may be emitted as fast pulse trains and have often been described as AM sounds (e.g. “moans”). Blue, fin and humpback whales may arrange their sounds into “song” that lasts for hours to days.
Odontocete sounds are typically classified as whistles, burst-pulse sounds and clicks in the literature though not all odontocetes whistle. Whistles are tonal sounds with CW or FM characteristics and may or may not have harmonic overtones. Most studies on whistles have focussed on describing the contour of the fundamental. Burst-pulse sounds are series of rapid pulses, typically reported for some delphinid species. Depending on the settings during Fourier analysis, burst-pulse sounds may appear as FM sounds with multiple overtones and non-harmonic sidebands [10]; however, they were classified as pulses in this article. Clicks are very short (typically <1 ms) and broadband pulses with peak energy at high frequencies (typically tens of kHz). Clicks can be heard as single signals or “slow” repetitive signals with inter-click intervals (ICI) of a few seconds, as in the case of sperm whales, or as rapid trains with millisecond ICI as in the case of odontocete echolocation. The ICI in click trains is typically larger than the inter-pulse interval (IPI) in burst-pulse sounds. The separation into FM sounds and pulses can be confusing. For example, the brief pulses of sei whales are FM downsweeps [11, 12]. And even the very brief clicks of odontocetes can be FM sweeps, like those of many beaked whales [13].

Indicative distribution of a baleen whales and some beaked whales, b, c odontocetes and dugong, d beaked whales and e pinnipeds. The grey circle is around Macquarie Island
Pinnipeds may emit sound in both media, air and water. Most studies have recorded their aerial vocalisations. Some species, such as the Ross seal, produce very similar sounds in both media [14]. The majority of pinniped sounds were classified as pulses in this article (including AM “moans” and “growls”); however, each pulse might be a brief frequency sweep or FM sound as in the case of Weddell seal chugs [15].
Despite these inconsistencies in naming sounds, for each species, we tabulate the sound types, with some notes on common vocalisation names, the frequency range, the duration, the source level (SL) as peak-to-peak sound pressure level (pp), root-mean-square sound pressure level (rms) or sound exposure level (SEL), the location where the sounds were recorded and the references identifying the source of the information. We have tried to keep the localities mentioned in the original literature, but sometimes summarise these to “Mediterranean” or “Northeast Atlantic” when there were several sites, studies or publications.
It is difficult to compile such summary tables from the literature, as each article lists different quantities in different formats. For frequency, duration and source level measurements, sometimes absolute ranges are given, other times means ± standard deviation (even though the measurements were often not Gaussian distributed), or medians and quartiles, or \(5{\mathrm{th}}\) and \(95{\mathrm{th}}\) percentiles, or \(10{\mathrm{th}}\) and \(90{\mathrm{th}}\) percentiles. These quantities and statistics cannot be accurately combined without access to the raw data. In an attempt to provide an overview of the available information, we decided to report the ranges (i.e. the broadest ranges). Therefore, the lowest frequency or duration or level we report is the minimum of all values published (e.g. the lowest percentile, the lowest range or the mean minus the standard deviation), and the highest value we list corresponds to the maximum of all values published (including the highest percentile, the highest range and the mean plus the standard deviation). If only one publication existed, we kept the original metrics as there was no need to combine data from multiple studies. Sometimes, measurements were not given, so we attempted to read data off spectrograms. All quantities, and in particular the observed bandwidth, can strongly be affected by the sound propagation environment and range of source to receiver; however, this is mostly not corrected in the literature.

Duration and fundamental frequency range of tonal (CW and FM) sounds of mysticetes, odontocetes, dugong and pinnipeds under water

Duration and frequency range of mysticete, odontocete, dugong and pinniped underwater pulses and burst-pulse sounds
In the case of whistles, the frequency range reported in this document corresponds to the range in primary frequency fp (i.e. the maximum frequency minus minimum frequency of the fundamental contour) and excludes harmonics. Some whistles are emitted with harmonics, some without. In fact, the same fundamental contour may sometimes be recorded with harmonics and other times without (e.g. in the case of killer whales [16]). This is different for FM sounds produced by mysticetes, where the frequency range reported in the literature mostly includes harmonics. If measurements were reported for only the fundamental frequency and not for harmonics present, we indicate this as “+ harmonics” in the following mysticete table.
In the case of clicks, listing only the range from minimum to maximum reported frequency resulted in all animals apparently producing hugely broadband clicks. Odontocete click emission is directional, and spectra change as a function of angle in both the horizontal and vertical planes, leading to variability in frequency measurements. Furthermore, clicks are commonly bimodal, exhibiting not just one but two peak frequencies, which is missed by studies that report mean or median frequencies. In addition to uncertainty in the angular aspect and range to the animal, the lack of a low-pass anti-aliasing filter during recordings at low sampling frequencies can affect the spectrum. If the click spectrum extends beyond the Nyquist frequency and no anti-aliasing filter is used, the spectral energy “folds” about the Nyquist frequency to lower frequencies. We list multiple frequency measurements for clicks if available: fp, the frequency of maximum spectral power; fc, the centroid frequency that divides the power in a spectrum into two equal parts; and the bandwidth at the \(-\)3 dB or \(-\)10 dB points, or the rms bandwidth, i.e. the standard deviation of the spectrum about fc.
Figures 2, 3 and 4 are an attempt to visualise the frequency ranges and durations of the various animal sounds based on the literature and the authors’ recordings.

Duration and frequency range of odontocete clicks

Southern right whale FM sounds recorded in Fowlers Bay (SA), August 2013 [26] [fs = 12 kHz, NFFT = 1200, 50% overlap, Hanning window]
The Sounds of Mysticetes—Baleen Whales
Eubalaena australis—Southern Right Whale
Southern right whales are found in the southern hemisphere, along the southern coasts of South America, Africa, Australia and New Zealand. They spend summers on their feeding grounds close to Antarctica and migrate to their northern breeding grounds in winter. Their seasonal Australian range (May–November) extends from Hervey Bay (QLD) on the east coast, along the southern coastline, to Exmouth (WA) on the west coast [17]. Close to shore, southern right whales seasonally form distinct groups in about a dozen coastal aggregation areas [18,19,20].
Descriptions of the sounds of southern right whales have been published from the South Atlantic, off Argentina and Brazil [21,22,23,24] and from the South Pacific, around New Zealand [25]. CMST has been recording southern right whales in Fowlers Bay (SA) during the austral winter [26].
All of the populations produce FM sounds, with fundamentals ranging from 30 to 500 Hz, harmonics up to 5 kHz, and durations of 0.2–3.8 s (Fig. 5a; [21,22,23,24,25]). The most frequently reported FM sound is a 0.2–2.7 s upsweep from 30 to 450 Hz plus harmonics (referred to as the “upcall” in the right whale literature; Fig. 5b; [21, 24, 25]). It seems to be the primary contact call, with variability in the call structure potentially coding caller identity [21]. Right whales off Argentina have shifted their upcall frequency upwards by about 10–20 Hz from 1977 to 2000, possibly due to an increase in low-frequency anthropogenic noise [27].
All populations also emit CW sounds, with fundamentals ranging from 30 to 500 Hz, and harmonics up to 3.5 kHz, and durations of 0.2–6 s [21,22,23,24,25]. An exception is a low-frequency CW sound of up to 15.5-s duration, recorded off New Zealand [25]. Pulsed sounds last 0.2–6.3 s with energy in the band 30–3300 Hz [21, 22, 24, 25]. Finally, southern right whales also emit brief, broadband (30–8400 Hz) pulses, called “gunshots” with source levels of 172–187 dB re 1 \(\upmu \)Pa rms @ 1 m, and 0.1–0.4-s duration [21, 22, 25].
Caperea marginata—Pygmy Right Whale
Pygmy right whales have been seen around the southernmost coasts of South America, Africa, Australia [28, 29] and New Zealand suggesting a circumpolar distribution in temperate waters of the southern hemisphere [30]. The rarity of sightings means that very little is known about the distribution of this species. Currently, there is only one published paper on pygmy right whale acoustics, describing an FM downsweep from 200 to 50 Hz, lasting 0.3 s (Fig. 6; [31])

Pygmy right whale downsweep [31] [fs = 12 kHz, NFFT = 1200, 50% overlap, Hanning window]
Balaenoptera acutorostrata—Common Minke Whale
B. acutorostrata is found globally and typically referred to as the common minke whale. There is a North Atlantic subspecies (B.a. acutorostrata), a North Pacific subspecies (B.a. scammoni) and an unnamed subspecies that occurs in the southern hemisphere and that is referred to as the dwarf minke whale. This subspecies is typically sighted and recorded in warm, tropical to subtropical waters, rather than at high latitude. Its range extends from \(11^{\circ }\hbox {S}\) to about \(41^{\circ }\hbox {S}\). These animals are more likely distributed closer to shore [32]. Like other baleen whales, their breeding grounds are likely in lower latitudes (e.g. the Great Barrier Reef), with mature animals migrating there during the winter months.
Common minke whales have been recorded in the east of Canada [33, 34], in the Gulf of Maine [35, 36], in the Caribbean [37, 38], in the North Pacific including Hawaii [39,40,41] and the Mariana Islands [42], in the Great Barrier Reef, Australia [43], and along the Western Australian coast to as far north as \(14^{\circ }\hbox {S}\) [44].
Common minke whales primarily produce pulsed sounds which are arranged into trains that either have a constant pulse repetition rate, or are sped up, or slowed down. The frequency band is 10–800 Hz. Pulses have a duration of 0.04–0.3 s; 1.5–4.5 pulses occur per second, in trains of 10–70-s duration, and with source levels of 159–176 dB re 1 \(\upmu \)Pa rms @ 1 m or 173–189 dB re 1 \(\upmu \)Pa pp @ 1 m [35,36,37]. Very-high-frequency clicks (4–7.5 kHz; 6.75 ± 1.02 clicks/s, 166–173 dB re 1 \(\upmu \)Pa pp @ 1 m) were only reported once from Newfoundland [33].
North Pacific minke whales were first reported to produce the famous “boing” sound. The “boing” is an AM sound, often described as a train of rapid pulses, and which appears in spectrograms (with analysis windows longer than the pulse duration) as a broadband FM sound with many sidebands. The “boing” covers a bandwidth of 1–5 kHz, lasts 1.4–4.2 s and exhibits about 112–118 pulses/s, at a source level of about 150 dB re 1 \(\upmu \)Pa rms @ 1 m [39,40,41]. These “boing” sounds are commonly recorded by CMST on Australia’s Northwest Shelf as well (e.g. [44]; Fig. 7a, b). At the Australian Great Barrier Reef, dwarf minke whales produce the pulsed, metallic-sounding “star-wars call” (50–9400 Hz, 1–3 s, 150–165 dB re 1 \(\upmu \)Pa rms @ 1 m; [43]; Fig. 7c).
FM downsweeps (50–250 Hz; 0.2–0.4 s) were reported from the St Lawrence Estuary, Canada [34], and the Great Barrier Reef, Australia [43].

a Spectrogram of a minke whale “boing” recorded by CMST on Australia’s Northwest Shelf, May 2009 [fs = 6 kHz, NFFT = 600, 50% overlap, Hanning window], and b corresponding waveform. c Spectrogram of the “star-wars call” recorded near the Great Barrier Reef, July 1998 [43] [fs = 44 kHz, NFFT = 2048, 50% overlap, Hanning window]
Balaenoptera bonaerensis—Antarctic Minke Whale, Dark-shoulder Minke Whale
There are two species of minke whales: B. acutorostrata and B. bonaerensis. The latter only occurs in the southern hemisphere from Antarctica to near the equator (\(10^{\circ }\hbox {S}\)) and migrates seasonally as do other baleen species. This species’ Australian range extends along the whole of the eastern and western coastlines (though less is known about their distribution on the western coastline), and its southern range extends to the ice edge well within the Australian Antarctic Territory [45]. Antarctic minke whales are considered an offshore and migratory species. In the winter breeding grounds at low latitudes in other regions, these animals appear to be distributed off the continental shelf edge [46], suggesting a similar winter distribution for Australian waters during the breeding season. Although some individuals might remain in Antarctica over the winter [47].
In Antarctica, FM downsweeps with a fundamental of 60–140 Hz, occurring with and without harmonics, with a duration of 0.2 ± 0.1 s and a source level of 147.3 ± 5.3 dB re 1 \(\upmu \)Pa rms @ 1 m have been recorded [48,49,50,51]. Antarctic mine whales also emit bouts of rapid broadband pulses (which are brief FM downsweeps), sounding like a duck’s quack (named the “bioduck” sound). This unique “bioduck” sound has been recorded in Antarctica and off Australia for decades, yet it was not until Risch et al. [49], that this sound was positively attributed to Antarctic minke whales. The “bioduck” sound contains pulses of 0.1–0.4-s duration, in bouts of 5–12 pulses, with a 1–3-s interval between bouts, and source levels of 140.2 ± 3.6 dB re 1 \(\upmu \)Pa rms @ 1 m [49, 51]. The “bioduck” (Fig. 8) is also commonly heard in the Perth Canyon in the austral winter, where it covers 100–500 Hz in frequency with about 0.2 s/pulse and 5 pulses/train [6] and occurs as far north as Exmouth, WA [44].

“Bioduck” sound recorded by CMST a south of Exmouth (WA), August 2011, and b, c in the Perth Canyon (WA), September 2003 [fs = 10 kHz, NFFT = 128, 50% overlap, Hanning window]
Balaenoptera borealis—Sei Whale
The sei whale is a global species and occurs in both hemispheres. The northern hemisphere subspecies is B.b. borealis, and the southern hemisphere subspecies is B.b. schlegelii. This species migrates once a year between its polar feeding grounds (summer) and tropical breeding grounds (winter). Sei whales tend to stay well offshore [52], though their tendency to be confused with Bryde’s whales has resulted in confusion about their distribution. The extent of occurrence, their geographic habitat, seasonal presence and absolute abundance around Australia are still poorly understood [53].
Acoustic recordings exist from the North Atlantic, off the US east coast [54,55,56] and Nova Scotia, Canada [11, 12], off the Azores [57], from Hawaii [58], south of New Zealand [59] and Antarctica [60]. There are no recordings of sei whales in Australian waters with simultaneous visual species identification.
Sei whales produce 0.5–2-s CW sounds at 200–500 Hz, without or with higher-frequency harmonics, including multi-part frequency stepping tonals [60]. The most frequently reported sounds are simple FM sounds: upsweeps and downsweeps, lasting 0.7–2.2 s, with the fundamental covering a low-frequency band (20–220 Hz; [54,55,56,57,58,59]) or mid-frequency band (200–600 Hz; [60]). These FM sounds have been recorded with and without harmonics. Source levels of FM and CW sounds are 147–183 dB re 1 \(\upmu \)Pa rms @ 1 m [55,56,57, 60]. Very brief (0.03–0.04 s) high-frequency (1.5–3.5 kHz) downsweeps occurring in bouts of 10–20 have been reported from Nova Scotia [11] and also been described as a series of pulses with peak energy at 3 kHz and 7–10 pulses over 0.7 s [12]. Broadband (100–800 Hz) pulsed sounds of 1–3-s duration have also been documented [60].
Balaenoptera edeni—Bryde’s Whale
Bryde’s whales inhabit tropical to subtropical waters in both the northern and the southern hemispheres. Two subspecies have been identified: the larger Bryde’s whale (B.e. brydei) that occurs worldwide in offshore waters and the smaller Eden’s whale (B.e. edeni) that may only occur in the Indo-Pacific. Offshore and coastal forms have been found in South Africa [61] and Japan [62], and similar forms likely exist in Australian waters. Bryde’s whales have been sighted in offshore and inshore waters all around the Australian mainland. The coastal form is considered resident, with no evidence of large-scale movements in Australia [63]. The offshore form may migrate seasonally, as do other baleen whale species, though more data are required.
Bryde’s whales have been recorded in the Gulf of Mexico [64, 65], in the Gulf of California [66,67,68], in the Caribbean [69], off Brazil [70], off Japan [69, 71], in the Eastern Tropical Pacific [69, 72] and off Western Australia.
Bryde’s whales emit 0.3–7-s CW tones at 8–80 Hz with and without higher harmonics [69, 70, 72]. They also make FM sounds, primarily downsweeps (50–1000 Hz, 0.1–5 s, 141–174 dB re 1 \(\upmu \)Pa rms @ 1 m) with and without harmonics [64,65,66, 68,69,70,71], sometimes occurring in sequences of 2–25 downsweeps [65]. FM sounds recorded by CWR in the presence of Bryde’s whales are shown in Fig. 9. Bryde’s whales further make AM sounds, which have been described as “moans” or rapid pulse trains (83–950 Hz, 0.3–51 s, 0.03–0.1 s/pulse, 10–130 pulses/s [64, 67, 70].

Sounds recorded by CWR in the presence of Bryde’s whales off Broome (WA), July 2013 [fs = 48 kHz, NFFT = 12,000, 50% overlap, Hanning window]
Balaenoptera musculus—Blue Whale
Blue whales (B. musculus) come in currently five recognised subspecies: the northern blue whale (B.m. musculus), which is only seen in the northern hemisphere, the Antarctic blue whale (B.m. intermedia), the northern Indian Ocean blue whale (B.m. indica), the pygmy blue whale (B.m. brevicauda) and the Chilean blue whale (unnamed). Based on acoustic analyses of blue whale sounds, size differences and genetics, there could be additional distinct populations or subspecies [73, 74]. Australian waters are home to at least two of these subspecies: the Antarctic blue whale and the pygmy blue whale. The Antarctic blue whale occurs at high latitudes into the Antarctic during the austral summer and migrates to lower latitudes around Australia, southern Africa and South America in the winter [75], although Antarctic blue whale vocalisations have been recorded in Antarctica all year-round, indicating that some animals remain at high latitudes throughout the year [76]. Pygmy blue whales are considered subantarctic, not occurring as far south as Antarctic blue whales. They also migrate annually between higher latitudes in the summer and lower latitudes in the winter and are believed to roam as far north as the northern Indian Ocean and Banda Sea [77]. The Perth Canyon (WA; [78, 79]) and the Bonney upwelling (SA; [80]) are two notable feeding aggregations of pygmy blue whales.
Southern hemisphere blue whales have been acoustically recorded in the southeast Pacific, off Chile [73, 81], in the southwest Pacific, off New Zealand [82, 83], in the Indian Ocean [84, 85], off Antarctica [76, 86, 87] and Australia [88,89,90]. They emit patterned and repetitive sequences of vocalisations, called “song”, lasting up to hours at a time. It appears only males sing, possibly as a form of reproductive display [91]. On a worldwide basis, song units can be CW, FM or pulsed, covering a frequency band of 14–300 Hz. Three- and four-unit phrases are common in blue whale song [73, 81,82,83,84]. Unit precursors in the form of 0.7–1-s pulses with peak energy between 347 and 430 Hz have been recorded in Chilean blue whale song [73, 81] and are similar to precursors recorded from blue whales in the North Pacific [92]. In addition to song, blue whales emit non-patterned sounds with FM sweeps being very common (e.g. 1–3 s, 80–40 Hz downsweeps called “D-calls” [91, 92]).
Pygmy blue whale song has been recorded off WA and along the southern Australian coast [6, 79, 85, 93,94,95,96,97,98]. Song units are FM type, with a fundamental frequency of 15–25 Hz and harmonics up to 100 Hz, although this can be higher for nearby animals. Song units are 15–40 s long, and several units (The classic form is three units.) are arranged into phrases of 60–120-s duration (Fig. 10a, b). Source levels of 168–181 dB re 1 \(\upmu \)Pa rms @ 1 m have been reported. It is interesting to note that the frequency of the song units has been decreasing steadily by about 0.35 ± 0.3 Hz/year over nine years [93] and is still decreasing at the same rate (A. Gavrilov, personal communication 2017). Non-song sounds can be 0.9–4.4 s long, are usually FM and are up to several hundred Hertz in frequency (Fig. 10c; [95]).
Blue whale song around New Zealand differs from the Western Australian pygmy blue whale song [83, 98, 99]. The typical sequence of units comprises two consecutive AM tones with associated harmonics, followed by an upsweep. A weak AM sound sometimes precedes this sequence. This phrase is of the order of 40–60 s, and the fundamental frequency is in the range 15–30 Hz, although harmonics can reach several hundred Hz (Fig. 10d). The New Zealand song type is also heard off the southeast coast of Australia with the dividing line between the New Zealand and Western Australian song types appearing to approximate the longitude of Bass Strait.
It is not always certain which subspecies of blue whale was recorded, except in the case of the “Z-call”, which has been attributed to the Antarctic blue whale (Fig. 10e). The “Z-call” appears as a Z in spectrograms, with the first part being a CW component at about 28 Hz lasting a few seconds, followed by a rapid downsweep to about 18 Hz, which is the last CW component, also lasting a few seconds [85,86,87, 89, 100]. In total, the “Z-call” has a duration of 16–30 s and can occur arranged into song of several hours. It is not necessarily emitted as a complete Z, but also occurs as just the higher tone, or the higher tone plus the rapid downsweep [100]. It is interesting to note that the frequency of the first tonal of the “Z-call” of Antarctic blue whales decreased by 1 Hz in 9 years [88] and is still decreasing at the same rate (A. Gavrilov, personal communication 2017). In addition to the “Z-call”, FM sweeps have been recorded from Antarctic blue whales, at 13–107 Hz, 0.8–7.5-s duration [100].
Blue whales produce some of the loudest biological sounds in the ocean (up to 189 ± 3 dB re 1 \(\upmu \)Pa rms @ 1 m; [81, 85, 86]), at the lowest frequencies, potentially yielding very long sound propagation ranges via ducting in the deep sound channel.

a Pygmy blue whale song units, b song sequence, c non-song sounds and d D-calls recorded by CMST in Geographe Bay (WA), November 2011. e Antarctic blue whale “Z-call” recorded by CMST in Bass Strait, May 2004. f New Zealand song type recorded by CMST in Bass Strait, March 2016. [fs = 12 kHz, NFFT = 8192, 50% overlap, Hanning window]
Balaenoptera omurai—Omura’s Whale
Omura’s whales were only described as a separate species from Bryde’s whales that have a similar appearance, in 2003 [101, 102]. Most confirmed records of the species are from specimens taken during research whaling and strandings. Based on these records, the distribution of the species is thought to be in tropical to subtropical latitudes of the Indo-Pacific region [103]. In Australia, validated specimens have been recorded offshore from the Cocos Islands [101], near Exmouth (WA; [103]) and in the Gulf of St. Vincent (SA; [104]). However, there is no information available on the spatial and temporal distribution, behaviour and abundance of this species in Australia. In fact, except for recent work off northwest Madagascar, there is a complete absence of ecological and behavioural data for the species throughout its distribution [105]. Only one published study to date has attributed vocalisations to Omura’s whale [105]. Low-frequency (15–50 Hz) AM vocalisations with an average duration of 9.2 ± 0.92 s produced every 134–177 s were described from boat-based recordings in the presence of Omura’s whales. Similar sounds (20–60 Hz, 12–15 s, repeated every 160–210 s; Fig. 11) are frequently recorded by CMST and CWR on Australia’s Northwest Shelf and simultaneous visual observations and photographs confirmed the presence of Omura’s whales.

Omura’s whale sound recorded by CMST on Australia’s Northwest Shelf, May 2006 [fs = 10 kHz, NFFT = 5000, 50% overlap, Hanning window]
Balaenoptera physalus—Fin Whale
The fin whale is a global species, inhabiting all the world’s oceans, migrating from the polar regions (feeding grounds) in summer to the equatorial regions (breeding grounds) in winter. However, there are exceptions, and non-migratory fin whales have been reported in the northern hemisphere [106]. There are three recognised subspecies: pygmy fin whales (B.p. patachonica), northern fin whales (B.p. physalus) and southern fin whales (B.p. quoyi). Both pygmy and southern fin whales occur in the southern hemisphere. Fin whale occurrence and migration along the Australian coasts are still poorly understood and known primarily from stranding events and whaling records. Strandings have been reported in WA, SA, VIC and TAS, suggesting that they are present around the southern coasts of the Australian continent. The migration routes and location of breeding grounds are uncertain [107].
Fin whales have been acoustically recorded off north-eastern Canada [108], in the St Lawrence Estuary [109], in the Bering Sea [110], in the North Pacific and North Atlantic [111,112,113], in the Gulf of California [66, 110, 114, 115], in Antarctica [76, 86] and in the Perth Canyon [6].
Fin whales emit characteristic 1-s FM downsweeps from about 30 to 15 Hz, commonly called the “20-Hz signal” [109, 114, 116]. These can be emitted in singlets, doublets or triplets and arranged into “song” lasting many hours, with IPI of 5–26 s [112, 115]. It is believed only males sing in order to attract females [117]. The IPI varies with geographic region, season and behaviour [118]. There sometimes is a higher-frequency component at 90–140 Hz (Fig. 12; [6, 76, 86, 108]). Many simultaneously singing fin whales can significantly raise ambient noise levels around 20 Hz by creating a continuous 20 Hz band in underwater acoustic recordings [6]. Source levels of up to 195 dB re 1 \(\upmu \)Pa rms @ 1 m have been reported [86, 111, 113] and are among the highest source levels of biological sounds underwater.
They also emit higher-frequency FM sounds of up to 310 Hz and up to 5-s duration [109, 110, 115], an AM “rumble” at 10–30 Hz and up to 30-s duration [109], AM “moans” up to 100 Hz, 2-s duration and 159–183 dB re 1 \(\upmu \)Pa rms @ 1 m source level [66], CW tones between 50 and 150 Hz and 0.3–0.7-s duration [109], and 34 and 68 Hz CW units arranged into 2-unit phrases of 3-s duration with 1.6–2.2 phrases/minute [66].

Fin whale song recorded by CMST in the Perth Canyon (WA), May 2003 [fs = 10 kHz, NFFT = 5000, 50% overlap, Hanning window]
Megaptera novaeangliae—Humpback Whale
Humpback whales occur globally as three subspecies: North Atlantic humpback whales (M.n. novaeangliae), North Pacific humpback whales (M.n. kuzira) and southern humpback whales (M.n. australis). All migrate annually between their winter breeding grounds in tropical waters and their summer feeding grounds in polar waters. The Australian migration follows two main routes along the QLD and WA coasts resulting in two different migratory populations [119]. Some animals skip a migration and over-winter at high latitudes [120].
The sounds of humpback whales have been studied extensively in Alaska [121,122,123], off the eastern USA [124], at low latitudes [125,126,127,128,129], off eastern Australia [130,131,132,133,134,135,136] and off Antarctica [137].
Male humpback whales use “song” likely as a means of sexual selection aimed at females or to mediate male–male interactions [128, 138, 139]. A song session can last for hours or days. All males of one population sing approximately the same song at any one time. Song normally changes slowly throughout the season and from season to season, although there can be abrupt song changes between years [140]. Males sing during the winter breeding season and during migration to and from the breeding grounds. Only rarely has singing been heard on the summer feeding grounds [137, 141]. Exceptions where humpback whales remained and sang at high latitudes during winter (breeding season) have been reported from Iceland [142]. Australian humpback whales that over-winter on their summer feeding grounds have not been heard singing [120].
There is a lot of literature on humpback whale song, geographic differences and changes over time (e.g. [121]). In Australia, the song differs between the west coast and the east coast populations of humpback whales; however, a rapid adaptation to the west coast song was observed by the east coast animals in 1 year, perhaps due to a few individuals moving from west to east [140] or due to song exchange on the summer feeding grounds in Antarctica where the Australian populations might converge in summer, and where some singing has been heard during the summer feeding season [137].
Song consists of 0.2–4 s units arranged into phrases, which are arranged into themes [127, 128, 139]. Units can be of CW, FM and AM type, typically with harmonics. The bandwidth of song with overtones can be as broad as from 20 Hz to more than 24 kHz; source levels are 151–189 dB re 1 \(\upmu \)Pa rms @ 1 m [125, 126] (Fig. 13).
Both male and female humpback whales produce social sounds. Social sounds include CW, FM, AM sounds and discrete pulses, as well as sounds from the blowhole, and sounds from breaching or slapping the pectoral or tail fins [3]. Social sounds can also include song units, termed “song-unit social sounds” [131], which are used during social interactions but are not emitted as continuous patterned song. Source levels of all types of social sounds range from 123 to 192 dB re 1 \(\upmu \)Pa rms @ 1 m [122, 130]. A 3-year study off the east coast of Australia found 34 different sound types [131], either as single sounds or in bouts [133]. However, only a small subset of these sound types remain in the repertoire over time [134], meaning that the sound catalogue is highly variable and constantly changing. All three of these studies recorded whales during their southern migration only; therefore, it is possible they produce many more sound types while feeding or on the breeding grounds.
Click trains termed “megapclicks” were recorded with DTAGs attached to two animals, during night-time foraging [143]. Clicks were broadband, with most energy below 2 kHz. The click interval decreased towards the end of a click train, ending in a buzz similar to feeding buzzes in odontocetes [144]. As further proof of these click trains being associated with feeding, click trains were emitted at the bottom of dives and the DTAGs recorded sharp body rolls towards the end of a click train [143]. Paired burst bouts of 0.25-s duration, with peak energy below 1 kHz, occurring in series of 2–120 paired bursts were also recorded during bottom feeding [145].

Examples of humpback whale song recorded by CMST on Australia’s Northwest Shelf, August 2013 [fs = 10 kHz, NFFT = 5000, 50% overlap, Hanning window]
The Sounds of Odontocetes—Toothed Whales
Delphinus delphis—Common Dolphin, Short-beaked Common Dolphin
This is a global species with currently four recognised subspecies: the common dolphin (D.d. delphis), the eastern North Pacific long-beaked common dolphin (D.d. bairdii), the Black Sea common dolphin (D.d. ponticus) and the Indo-Pacific common dolphin (D.d. tropicalis). Altogether, this species can be found in inshore and offshore tropical to temperate waters. It has been sighted all around Australia, albeit rarely in the north [146, 147]. There is some indication of migration and seasonal shifts in distribution [148]. There have been no publications of this species’ sounds in Australia previously. Instead, this species’ vocalisations have been described from New Zealand [149, 150], the Eastern Tropical Pacific [151, 152], the UK [153,154,155], the Mediterranean [156,157,158] and the Atlantic Ocean [158].
Whistles have fundamentals from 3 to 24 kHz and last 0.01–4 s [149,150,151,152,153,154,155,156,157,158,159]. Burst-pulse sounds have been reported at 2–14 kHz, lasting 0.5–0.75 s [157]; however, recordings from Australia appear much more broadband with energy up to 160 kHz (Fig. 14). Clicks peak between 23 and 67 kHz and have a 17–45-kHz bandwidth. They last 50–150 \(\upmu \)s (Dziedzic, 1978 quoted in [2]).

Common dolphin whistles, burst-pulse sounds and clicks; a recorded by CWR off SA, February 2013 [fs = 48 kHz, NFFT = 4800, 50% overlap, Hanning window]; b, c recorded by Sue Mason in Port Phillip Bay (VIC), June 2013 [fs = 384 kHz, NFFT = 19,200, 50% overlap, Hanning window]
Feresa attenuata—Pygmy Killer Whale
Pygmy killer whales are mainly tropical, but can be found in subtropical waters. They occupy offshore and inshore waters globally, though are thought to only come close to shore near oceanic islands. Sightings—without simultaneous recordings—and strandings have occurred off NSW and WA, and they have been sighted off the northeast of Australia [147, 160]. The spectral characteristics of their clicks (10–150 kHz, 25 \(\upmu \)s, 197–223 dB re 1 \(\upmu \)Pa pp @ 1 m) were published from the northern Indian Ocean [161]. Clicks, burst-pulse sounds and whistles were also recorded by CWR at Broken Ridge, Indian Ocean (Fig. 15). Whistles had fundamentals at frequencies above 5 kHz, often peaking around 10 kHz, with energy up to the Nyquist frequency, and lasted 0.3–0.8 s. Burst-pulse sounds had most energy above 3 kHz up to the Nyquist frequency, and commonly lasted 1–3.5 s.

Pygmy killer whale whistles, burst-pulse sounds and clicks recorded by CWR at Broken Ridge, Indian Ocean, November 2015 [fs = 48 kHz, NFFT = 4800, 50% overlap, Hanning window]
Globicephala macrorhynchus—Short-Finned Pilot Whale
Short-finned pilot whales occur globally in tropical to temperate waters both offshore and inshore; however, they are considered a deep-water species. Though thought abundant, only a few strandings have occurred in Australia [147]. Recordings from Australia have not been published yet.
In the Caribbean, Eastern Tropical Pacific, Gulf of Mexico, south-eastern USA, Canary Islands and Bahamas, this species emits whistles with fundamental frequencies ranging from 1 to 20 kHz and 0.1–2.5-s duration [8, 151, 152, 162,163,164,165,166]. Burst-pulse sounds cover a frequency band from 1 to greater than 30 kHz, lasting up to 1.5 s [8, 165]. Short-finned pilot whales exhibit the same graded structure of burst-pulse sounds as false killer whales, characteristic of a smooth burst-pulse to whistle transition [8]. Short-finned pilot whales further make biphonations, i.e. the simultaneous emission of two vocalisations by the same individual [8, 166, 167]. Foraging clicks and buzzes during deep dives have been recorded in the Canary Islands and off Hawaii showing a peak frequency of 8–39 kHz and 0.2–1.2 ms per click [168, 169]. Feeding buzzes last 0.7–6.8 s with 4–10-ms ICI; rasps are shorter (0.2–0.6 s) with longer ICI (11–50 ms) [166]. Finally, this species has been reported to copy tonal sounds of anthropogenic sources (e.g. mid-frequency sonar [170]). Australian recordings were obtained during a Coral Sea cruise by some of the authors (Fig. 16; [171]).

Short-finned pilot whale whistles (a) and clicks (b) recorded in the Coral Sea, July 2009 [171] [fs = 88 kHz, NFFT = 4400, 50% overlap, Hanning window]
Globicephala melas—Long-Finned Pilot Whale
There are two subspecies of long-finned pilot whales: G.m. melas in the North Atlantic and G.m. edwardii circum-Antarctic in the southern hemisphere. Little is known about the distribution of long-finned pilot whales in Australia, except that they prefer the colder water of the southern states and the Antarctic Territory [147].
Recordings from Australia have not been previously published. In the Mediterranean, eastern USA and Canada, and the northwest Atlantic, this species whistles between 1 and 11 kHz (fundamental) lasting 0.1–3 s per whistle [156, 157, 172,173,174]. CW tones have been reported at 1–8 kHz, 0.7–3-s duration off Newfoundland [173]. Burst-pulses cover a broader band (100–22,000 Hz) and last 0.1–2.2 s [157, 175]. Biphonations have been reported as well as a graded structure of burst-pulse to whistle transitions [157, 175]. Rhythmic repeated call sequences are common during social behaviours [176]. Long-finned pilot whales have also been reported to imitate mid-frequency sonar sounds in controlled-exposure experiments [177]. CMST and CWR recorded long-finned pilot whales on the shelf break south of Esperance (WA) and found CW and FM sounds with fundamentals between 1 and 6 kHz lasting 0.5–1.1 s (Fig. 17).

Long-finned pilot whale whistles, burst-pulse sounds and clicks recorded by CMST and CWR south of Esperance (WA), January 2016 [fs = 48 kHz, NFFT = 1200, 50% overlap, Hanning window]
Grampus griseus—Risso’s Dolphin, Grampus
Risso’s dolphins occur globally in tropical to subantarctic waters, both offshore and inshore. They have been reported in all Australian states except TAS and the NT [147]. They are thought to be an abundant species. Fraser Island has the only suspected “resident” population in Australia [178].
Recordings of low-frequency FM sounds with overtones and sidebands, whistles, burst-pulse sounds, clicks and biphonations (whistles and simultaneous burst-pulse sounds) were obtained off Newcastle (NSW; [179]). FM sounds with overtones and sidebands covered a bandwidth of 100–1300 Hz and lasted 4 ± 1.2 s. Such low-frequency sounds have not been reported from other parts of the world. Whistles recorded off Newcastle had fundamental frequencies from 4 to greater than 22 kHz (the recording bandwidth was limited to 22 kHz) and lasted 1–13 s [179]. While the frequency range matched that of whistles recorded off California, Egypt, Gran Canaria, the Azores, Scotland and in the Mediterranean [156, 180, 181], the duration of whistles from Australia was longer. Burst-pulse sounds off NSW covered a frequency band from 1 to above 22 kHz and lasted 2–13 s [179]. Animals recorded in the northern Indian Ocean, off California and off Gran Canaria, as well as captive animals in Hawaii and Taiwan emitted clicks with spectra extending from 10 to 140 kHz, durations of 26–118 \(\upmu \)s, ICI of 147–292 ms, source levels of 171–222 dB re 1 \(\upmu \)P pp @ 1 m, 163–210 dB re 1 \(\upmu \)Pa rms @ 1 m and 147–166 dB re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m, in trains of 2-s duration [181,182,183,184,185]. Off NSW, click trains were longer, up to 24 s [179]. Click trains come in a variety of ICI patterns, e.g. as speed-up, slow-down and constant-ICI trains. Some click trains end in (feeding) buzzes [181]. An example of clicks recorded by CWR in Jervis Bay (NSW) is shown in Fig. 18.

Risso’s dolphin clicks recorded by CWR in Jervis Bay (NSW), February 2013 [fs = 32 kHz, NFFT = 320, 50% overlap, Hanning window]
Lagenodelphis hosei—Fraser’s Dolphin, Sarawak Dolphin
This is a global, tropical to subtropical, offshore species, also occurring along the outer continental shelf and slope in deep water (>1 km deep). Strandings have been reported along Australia’s northern, western and eastern coasts [147]. Recorded acoustically in the Caribbean and the Eastern Tropical Pacific, these animals make whistles with 4–24 kHz fundamentals, lasting 0.06–2 s [186, 187]. Their clicks range to greater than 40 kHz [187]. On Australia’s northwest coast, whistles had fundamentals from 1.5 to 7 kHz and lasted 0.7–3.5 s (Fig. 19).

Fraser’s dolphin whistles and clicks recorded by CWR on Australia’s northwest coast, October 2012 [fs = 32 kHz, NFFT = 1600, 50% overlap, Hanning window]
Lagenorhynchus cruciger—Hourglass Dolphin
The hourglass dolphin is a southern hemisphere, circumpolar species, found from close to the Antarctic ice to mostly \(45^{\circ }\hbox {S}\), but occasionally as far north as \(36^{\circ }\hbox {S}\). They are rarely seen near land and usually sighted in the Antarctic away from pack ice [147]. Their echolocation clicks cover 110–140 kHz, are 79–176 \(\upmu \)s long and have source levels of 190–203 dB re 1 \(\upmu \)Pa pp @ 1 m or 179–193 dB re 1 \(\upmu \)Pa rms @ 1 m [188, 189].
Lagenorhynchus obscurus—Dusky Dolphin
Dusky dolphins can be found off the coasts of South America (Fitzroy’s dolphin, L.o. fitzroyi; and Peruvian/Chilean dusky dolphin, L.o. posidonia), southern Africa (African dusky dolphin, L.o. obscurus), New Zealand (unnamed subspecies) and southern Australia and TAS, as well as some oceanic islands. The subspecific status of the Australian animals is unknown as they are rarely sighted in Australia but are thought to occur across southern Australia [190]. They have only been recorded acoustically in New Zealand. They emit whistles with fundamentals of 7 to greater than 16 kHz, lasting 0.3–0.8 s. Their burst-pulse sounds range from 1 to greater than 16 kHz and last 0.1–0.6 s. Clicks cover 40–110 kHz with bimodal structure and have a source level of 210 dB re 1 \(\upmu \)Pa pp @ 1 m [191, 192].
Lissodelphis peronii—Southern Right Whale Dolphin
The distribution range of this species is subtropical to subantarctic oceans of the southern hemisphere. The range and total population have not been estimated or closely studied, but the species is abundant off the coast of New Zealand, often recorded with other cetaceans, such as the dusky dolphin. In Australia, they are thought to occur off the southern continental shelf [147]. No records of vocalisations have been found in the literature.
Orcaella heinsohni—Australian Snubfin Dolphin
The Australian snubfin dolphin is a relatively “new” species. These animals were previously classified as Orcaella brevirostris, or Irrawaddy dolphin, common in southeast Asia. Yet consistent and statistically significant differences in anatomical skull characteristics, dorsal fin height, colouration and mtDNA supported the classification of the Australian populations as a new species: O. heinsohni [193]. Snubfin dolphins stay in tropical, shallow, protected coastal and estuarine waters, as far south as Broome (WA) and Brisbane (QLD).
Two publications are available on the acoustic repertoire of this species [194, 195]. Whistles have fundamentals ranging from 600 Hz to 13 kHz and last 0.1–0.45 s. CW sounds were not found. Burst-pulse sounds range from 1 to greater than 22 kHz and last 4 ± 2 s. Clicks trains span 300 Hz to greater than 22 kHz, last 0.1–20 s and have 10–46 clicks/s. Buzzes have up to 116 clicks/s [194]. Example whistles, burst-pulses and clicks are shown in Fig. 20, demonstrating energy of burst-pulse sounds and clicks up to 90 kHz when animals are at close range.

Orcinus orca—Killer Whale
Killer whales are a global species, occurring both offshore and inshore, from the equator to the poles [198]. They are the most cosmopolitan and wide-ranging marine mammal. They have been sighted in all Australian states with frequent sightings around TAS, SA, VIC, as well as Ningaloo Reef and the Bremer Canyon in WA [199, 200].
Their sounds have been studied in coastal British Columbia, Canada, and Washington State, USA [201,202,203,204,205,206,207,208,209,210], in California [211], in other areas of the North Pacific, including Russia [212,213,214,215,216], in the North Atlantic off Norway and Iceland [217, 218], in Antarctica [219, 220] and south-western Australia [16]. A geographic comparison of North Atlantic and North Pacific killer whale sound frequencies was recently published [221].
Killer whales produce FM sounds (whistles) of variable duration (0.06–18.3 s) with a fundamental frequency range of 1–30 kHz (Fig. 21; [16, 208,209,210, 219]). Uncommon, low-frequency FM sounds with a fundamental of 50–270 Hz lasting 0.1–2.8 s were reported from Iceland [222]. Killer whales also produce high-frequency FM sounds with a fundamental of 16–75 kHz and source levels of 185–193 dB re 1 \(\upmu \)Pa pp @ 1 m [212, 215, 217]. CW tones are rare (3–5 kHz, 0.37 ± 0.08 s; [208]).

Their burst-pulse sounds have been studied extensively and cover a frequency range of 500 Hz–25 kHz and last 0.1–1.5 s, at source levels of 131–176 dB re 1 \(\upmu \)Pa rms @ 1 m, or 146–158 dB re 1 \(\upmu \)Pa pp @ 1 m [16, 203,204,205,206,207, 211, 213, 214, 219].
Their echolocation clicks span 10–110 kHz, 100–250 \(\upmu \)s, in trains of 3–7 s, at source levels of 107–224 dB re 1 \(\upmu \)Pa pp @ 1 m [201, 202, 211, 218]. In Australia, slow click trains with an ICI of 0.1 s, as well as speed-up trains ending in a “buzz” sound with an ICI <2.5 ms were recorded. Clicks had a negative Gabor waveform and lasted 100 \(\upmu \)s. Peak energy lay between 12 and 24 kHz [16].
Biphonations are common, specifically in the form of lower-frequency burst-pulse sounds together with higher-frequency, independent FM components [203, 213, 214, 220, 223]. Populations along the Canadian British Columbia coast have been studied extensively using passive acoustics. These and other populations exhibit dialects, i.e. slight differences in vocalisations among pods sharing the same habitat [224, 225].
Peponocephala electra—Melon-Headed Whale
Melon-headed whales are offshore animals inhabiting tropical to temperate waters around the globe. They have been sighted at sea along the Australian coast, including WA and specifically the Browse Basin. Stranding records suggest they occur along the entire northern half of Australia [227]. Acoustic recordings from Australia do not exist. Off Hawaii and in the Caribbean, whistles with fundamentals of 890–24,500 Hz and 0.1–1.4-s duration have been recorded [7, 228, 229]. Burst-pulse sounds appear more broadband (0.5–40 kHz) with 0.1-s duration, 2.47-ms IPI, 47 pulses/train, and a source level of about 165 dB re 1 \(\upmu \)Pa rms @ 1 m [228, 229]. The spectro-temporal structure of vocalisations is graded and falls along a continuum of burst-pulses to whistles [7]. Click spectra cover 6–50 kHz with 0.02–0.45 ms per click, 86–150-ms ICI, up to 1200 clicks/s and source levels of 132–140 dB re 1 \(\upmu \)Pa pp @ 1 m [228,229,230,231].
Pseudorca crassidens—False Killer Whale
False killer whales are found worldwide in tropical to temperate waters, mostly offshore but also inshore, in particular when the continental shelf is narrow. They have stranded in all Australian states [232]. Acoustic recordings of wild animals exist from the northern Indian Ocean [182], Eastern Tropical Pacific [151] and Caribbean [180], in addition to recordings of captive individuals.
False killer whales produce whistles at 5–8 kHz (range of the fundamental) of 0.2–0.8-s duration [151, 180]. Their burst-pulse sounds consist of series of rapid pulses of sinusoidal signals usually in a speed-up pulse train. Burst-pulse sounds cover a band of 0.5–22 kHz and last 0.05–1 s [9]. As the IPI decreases and the damping of sinusoidal signals into pulses decreases, burst-pulse sounds gradually turn into tonals with harmonics and sidebands [9]. False killer whales sometimes emit bimodal clicks, which have two frequency peaks, one at about 40 kHz, the other at about 100 kHz [233]. Peaks at lower frequency (20 kHz) have also been reported [169]. Click duration ranges from 18 to 530 \(\upmu \)s [169, 182, 233,234,235,236,237]. Reported source levels are 195–225 dB re 1 \(\upmu \)Pa pp @ 1 m, 190–215 dB re 1 \(\upmu \)Pa rms @ 1 m and 145–168 dB re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m [182, 233]. This species has been shown to imitate anthropogenic tonal sounds; false killer whales increased their whistle rate and copied the spectral features of mid-frequency sonar during controlled-exposure experiments [170]. False killer whale whistles, burst-pulse sounds and clicks were recorded by some of the authors in the Coral Sea, Australia (Fig. 22; [171]).

False killer whale whistles recorded in the Coral Sea, July 2009 [171] [fs = 300 kHz, NFFT = 2000, 50% overlap, Hanning window]
Sousa sahulensis—Australian Humpback Dolphin
This is a coastal species, occurring in shallow water along Australia’s northern coast with resident populations in Moreton Bay (QLD) and Shark Bay (WA) and potentially others [238]. The Australian humpback dolphin is slightly different in morphology from the other humpback dolphins in southern Asia and India, and it was declared a separate species in 2014 [239]. It had previously been classified as Indo-Pacific humpback dolphin (S. chinensis).
Whistles, burst-pulse sounds and clicks have been recorded off QLD and WA (Fig. 23; [195, 240, 241]). Signature whistles, identifying individuals, were also confirmed [242]. Whistles of S. sahulensis from QLD and of S. chinensis in southeast Asia have fundamentals ranging from 1 to 33 kHz with durations of 0.1–9 s; note that durations longer than 3 s were only reported from QLD [195, 240, 242,243,244,245,246,247]. CW tones were reported from S. chinensis in southern China at 500 Hz–22 kHz and 0.03–2-s duration [245]. The differences in whistle characteristics of S. sahulensis and S. chinensis have been detailed [248]. Burst-pulse sounds cover a frequency band of 600 Hz–25 kHz and last 0.1–8 s [195, 240, 247]. Clicks are very broadband with 10 dB bandwidths of \(116\pm 20\) kHz, peaking at 114 kHz [241]. In China, a bimodal spectrum was found peaking at about 100 kHz and 180 kHz [249, 250]. Durations of clicks are 10–100 \(\upmu \)s with ICIs of 0.01–0.25 s [241, 247, 249,250,251]. Source levels of 199 ± 3 dB re 1 \(\upmu \)Pa pp @ 1 m, 189 ± 3 dB re 1 \(\upmu \)Pa rms @ 1 m and 141 ± 3 dB re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m have been reported [241, 250, 251].

Australian humpback dolphin whistles [195] [fs = 44 kHz, NFFT = 2200, 50% overlap, Hanning window]
Stenella attenuata—Spotted Dolphin, Pantropical Spotted Dolphin
Two species of spotted dolphin exist: Stenella attenuata in all the major tropical oceans (Atlantic, Pacific, Indian) and S. frontalis only in the Atlantic Ocean. S. attenuata has two subspecies: the offshore pantropical spotted dolphin (S.a. attenuata) and the coastal pantropical spotted dolphin (S.a. graffmani). In Australia, S. attenuata occupies the northern coasts and is not seen along the Great Australian Bight or Tasmania [147]. There are no recordings from around Australia. The only recordings of S. attenuata globally are from Hawaii [252], the Eastern Tropical Pacific [151, 152] and the Gulf of Mexico [253]. These animals whistle at 2–23 kHz (fundamental), lasting 0.3–1.3 s with a source level of 115–163 dB re 1 \(\upmu \)Pa rms @ 1 m.
Stenella coeruleoalba—Striped Dolphin, Euphrosyne Dolphin
This species exists globally in tropical to temperate offshore waters. Strandings along Australia’s coastline are infrequent but have occurred on the northern, western and eastern coasts [254]. Acoustic recordings exist from the Eastern Tropical Pacific [151, 152], the Azores [255], the Canary Islands [255] and the Mediterranean [255], where these animals whistle with fundamentals of 1–31 kHz and durations of 0.04–3 s. CWR saw striped dolphins in the Great Australian Bight, south of WA, in Dover Canyon and recorded whistles with 4–22 kHz fundamentals lasting 0.3–1 s (Fig. 24).

Striped dolphin whistles recorded by CWR in Dover Canyon (WA), February 2016 [fs = 48 kHz, NFFT = 2400, 50% overlap, Hanning window]
Stenella longirostris—Long-Snouted Spinner Dolphin
Spinner dolphins are found globally in tropical to temperate offshore waters, with some occurrence on the continental shelf in shallow water near islands and reefs. There are currently four recognised subspecies: the central American spinner dolphin (S.l. centroamericana), the eastern spinner dolphin (S.l. orientalis), the dwarf spinner dolphin (S.l. roseiventris) and Gray’s spinner dolphin (S.l. longirostris), which is found in Australian waters. Spinner dolphins have been sighted in the Indian Ocean hundreds of km from the nearest land. They have been sighted off northern, western and eastern Australia with numerous bycatch records from the Arafura and Timor Seas. They are also known to occur in the Great Barrier Reef [147].
There are no published sound recordings from Australia. Their sounds have instead been document from Hawaii [230, 256,257,258,259,260,261,262], the Eastern Tropical Pacific [151, 152], the north-western Atlantic [174], Brazil [263,264,265] and the western Indian Ocean off Madagascar [266].
This species emits CW sounds, but they have consistently been lumped with FM sounds during analysis. Whistles range from 2 to 23 kHz (fundamental) and last from 0.1 to 1.8 s [151, 152, 174, 256,257,258,259, 264, 266]. Examples of whistles recorded by CWR at Scott Reef (WA) are shown in Fig. 25. Burst-pulse sounds span a frequency band of 220 Hz-130 kHz, lasting 0.05–2 s. There can be 30 pulses/train with an IPI of 3.85 ms [259, 263]. Clicks cover 20–130 kHz, last 0.2–0.4 ms with an ICI of 121–235 ms [230, 260].

Spinner dolphin whistles recorded by CWR at Scott Reef (WA), August 2012 [fs = 48 kHz, NFFT = 4800, 50% overlap, Hanning window]
Steno bredanensis—Rough-Toothed Dolphin
Rough-toothed dolphins are a deep-water offshore species inhabiting tropical to temperate seas globally. In Australian waters, they have been sighted off Barrow Island (WA), NT, QLD and NSW suggesting tropical–subtropical distribution [267]. They have been acoustically recorded in the Mediterranean [268], Central and Eastern Tropical Pacific and off Brazil [151, 152, 269,270,271]. They emit FM sounds between 1.6 and 28.2 kHz (fundamental) of 0.03–2.2-s duration. Off Brazil, their FM sounds were very weakly frequency-modulated, with about 34% CW tones and 29% FM upsweeps [269]. Clicks ranging from 10 to 50 kHz, lasting 0.14–0.35 ms with 1.6–354-ms ICI were recorded in the Tropical Pacific [271].
Tursiops aduncus—Indian Ocean Bottlenose Dolphin, Spotted Bottlenose Dolphin
This species occurs in shallow, subtropical coastal and estuarine waters along the Indian Ocean, Indo-Pacific region and the western Pacific Ocean. In Australia, these animals are found in estuarine and costal waters along the west, north and east coasts [272]. Along Australia’s south coast, there might be a newly defined species endemic to SA, the Burrunan dolphin (Tursiops australis) [273, 274]; however, this species is currently not recognised by the Society for Marine Mammalogy.
Acoustic recordings exist from Australia’s east and west coasts, as well as Japan, Tanzania and South Africa. Comparative studies of vocal repertoires have been undertaken by many researchers [275,276,277,278,279]. Most research has focussed on whistles, which range in frequency from 1 to 22 kHz (fundamental) and in duration from 0.1 to 5.9 s (Fig. 26). CW sounds are mostly lumped with FM sounds and their parameters are therefore not always listed separately [280]. T. aduncus have been shown to produce signature whistles [281]. Echolocation clicks cover a frequency band from 45 to 109 kHz with a peak frequency of 124 ± 13 kHz, last 18 ± 6 \(\upmu \)s, with an ICI of 63 ± 45 ms and source levels of 205 ± 7 dB re 1 \(\upmu \)Pa pp @ 1 m, 195 ± 4 dB re 1 \(\upmu \)Pa rms @ 1 m and 146 ± 7 dB re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m [241, 282]. There is no discussion of burst-pulse sounds in the literature. Acoustic recordings of Burrunan dolphins have not been published yet [283].

Tursiops truncatus—Bottlenose Dolphin
There are two subspecies of T. truncatus: the Black Sea bottlenose dolphin (T.t. ponticus) and the common bottlenose dolphin (T.t. truncatus). Common bottlenose dolphins inhabit tropical to temperate waters globally and occur around Australia in coastal as well as offshore waters [147, 272]. Bottlenose dolphins have been acoustically recorded in QLD, Australia [284], New Zealand [285], the Eastern Tropical Pacific [151, 152], Hawaii [230], California [276, 286], the Gulf of Mexico [163, 253, 276, 287], the Caribbean [288], Central America [286, 289], Brazil [270, 290], Uruguay [291], Argentina [276], Namibia [292], the Northwest Atlantic [163, 174], the Northeast Atlantic [282, 293, 294], Scotland and the UK [282, 295, 296], Mediterranean [156, 293, 297,298,299], and Portugal [300]. Many recordings have also been taken of bottlenose dolphins in captivity (e.g. [2, 301,302,303]).
Much of the literature focuses on whistles and geographic differences [151, 152, 156, 163, 174, 253, 270, 276, 286, 289,290,291,292,293, 295,296,297,298, 300, 302]. Biphonic whistles have been reported [299]. CW sounds are typically lumped with whistles and called “flat” whistles. Together, CW and FM sounds range from 300 Hz to 39 kHz in fundamental frequency and from 40 ms to 4 s in duration (Fig. 27), with source levels of 114–163 dB re 1 \(\upmu \)Pa rms @ 1 m. FM sounds at lower frequency have sometimes been described separately and called “low-frequency narrow-band” sounds with a 30 Hz–1 kHz fundamental frequency and 10-ms–8.7-s duration [284, 287, 292, 303].
Bottlenose dolphins develop signature whistles during the first few months of their lives and emit these when meeting conspecifics; they also copy signature whistles of others interpreted as “calling others” [294, 296, 299, 302, 304,305,306,307,308,309,310,311,312,313].
Burst-pulse sounds range from a few kHz to more than 150 kHz, last 0.1–4 s, with 150–1050 pulses/s and source levels of about 195 dB re 1 \(\upmu \)Pa pp @ 1 m [292, 301, 314]. Echolocation clicks range from 20 to 150 kHz, peaking between 50 and 110 kHz. Clicks last 10–80 \(\upmu \)s with 20–180-ms ICI, 11.4 ± 1.6 clicks/train and 1–4 s/train, and source levels of 193–228 dB re 1 \(\upmu \)Pa pp @ 1 m have been reported [2, 230, 282, 291, 314].
The signals emitted by T. truncatus are directional (clicks, burst-pulse sounds and whistles). Directionality increases with frequency. Higher frequencies and amplitudes are emitted forward (i.e. on-axis; [301]). The spectra of extreme off-axis clicks were also published [315]. While click rates are listed in many publications, these can depend on the depth of the animal during a foraging dive [316].

Bottlenose dolphin whistles recorded by CWR off Albany (WA), March 2013 [fs = 200 kHz, NFFT = 10,000, 50% overlap, Hanning window]
Kogia breviceps—Pygmy Sperm Whale
Pygmy sperm whales, a global species, tend to stay in deeper water off the continental shelf ranging from tropical to warm temperate zones [317]. Strandings have been reported in all Australian states, yet sightings at sea are rare. There are no acoustic recordings of pygmy sperm whales in the wild at the time of concurrent visual sightings. Pygmy sperm whales that stranded in Monterey Bay and New Jersey and were then transported to an aquarium produced clicks in captivity between 60 and 200 kHz, with a spectral peak at 125 kHz, 120–600 \(\upmu \)s/click, in trains of 20 clicks/s, including speed-up click trains [318,319,320]. A captive animal that had stranded in Hawaii further produced 0.42-s FM sounds at about 1.4–1.5 kHz [321]. The earliest recordings of pygmy sperm whale sounds were done with a contact microphone in air, while the animal was out of the water [322], likely leading to unrealistically low-frequency content [319]. No recordings from around Australia exist.
Kogia sima—Dwarf Sperm Whale
Dwarf sperm whales are global, offshore animals, likely preferring tropical and warm temperate water [317]. Strandings have been reported in all Australian states. The dwarf sperm whale is an inconspicuous animal that is rarely seen at sea. No records of its acoustic signals were found in the literature. CWR obtained acoustic recordings simultaneously with visual species identification on the Rowley Shoals (WA) (Fig. 28). Whistles had a lot of steps (discontinuities in frequency).

Dwarf sperm whale sounds recorded by CWR on the Rowley Shoals (WA), October 2012: a CW, b FM, c burst-pulse sound and d clicks [fs = 32 kHz, NFFT = 1600 (640 for c), 50% overlap, Hanning window]
Phocoena dioptrica—Spectacled Porpoise
The spectacled porpoise occurs only in the southern hemisphere at subantarctic latitudes. Its distribution is circumpolar [147]. It is rarely seen at sea. Strandings have been reported from the south-eastern coast of South America, and various offshore islands. No records of its vocalisations were found in the literature.
Physeter macrocephalus—Sperm Whale
Sperm whales, a global species, tend to stay along the shelf edge and offshore. They have been sighted off all Australian states. Females and young tend to stay in warmer waters (N of \(45^{\circ }\hbox {S}\)), while males travel to Antarctica and back [323].
Sperm whales have been recorded off the US east coast [324,325,326,327,328], off Nova Scotia [329, 330], around the Bahamas [331,332,333], around Bermuda [334], in the Caribbean [335,336,337,338,339], in the Gulf of Mexico [340,341,342,343], in Alaska [344,345,346,347,348], in the northeast Pacific [349], off Japan [350, 351] around Hawaii [352], at the Galapagos Islands [173, 353, 354], at the Azores [355], off Scotland [356], off Norway [357,358,359,360,361], in the Mediterranean [362,363,364,365,366], in the South Pacific [367], in New Zealand [368,369,370,371,372] and Antarctica [373].
The only sounds that sperm whales produce are clicks, first reported in 1957 [374]. Apart from a geographic variation in repertoire, group-specific dialects have been found in interacting groups with overlapping geographic range [367, 375]. It has been suggested that clicks serve communication, individual identification and echolocation purposes and may be used to debilitate prey [144, 326, 336, 342, 354, 357, 370, 376,377,378,379,380,381].
Sperm whale clicks are now commonly categorised into five classes according to their temporal pattern [330]:
-
1.
“Squeals” with up to 1600 clicks/s [382],
-
2.
“Creaks” with up to 220 clicks/s,
-
3.
“Usual clicks” with an ICI of about 0.5–1 s, and
-
4.
“Slow clicks” (sometimes called “single clicks”) with a long ICI of 3–8 s.
-
5.
Socialising groups emit stereotyped click sequences (patterns) called “codas” over periods lasting up to several hours [328].
Clicks have a bandwidth of 100 Hz–30 kHz, with peak energy around 15 kHz. Click duration is 0.2–3 ms. Each click can consist of a handful of pulses, and the time between pulses correlates with the size of the sperm whale (head) [383]. Source levels of usual clicks are 186 ± 0.9 dB re 1 \(\upmu \)Pa rms @ 1 m, 170 ± 0.7 dB re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m and 200–205 dB re 1 \(\upmu \)Pa pp @ 1 m [346], with extremes of up to 196 dB re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m and 236 dB re 1 \(\upmu \)Pa rms @ 1 m [358] and are the loudest levels reported for a biological source. Source levels of creaks have been reported as 166–205 dB re 1 \(\upmu \)Pa rms @ 1 m, 180 ± 6 dB re 1 \(\upmu \)Pa pp @ 1 m and 145–161 dB re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m [346, 357]. Source levels of slow clicks are 175–190 dB re 1 \(\upmu \)Pa rms @ 1 m and 156–166 re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m [357]. The click rate recorded from a group of sperm whales has been reported to increase with the number of animals in the group at 1.22 clicks/s/animal, depending on behaviour [384]. Two neonate sperm whales in rehabilitation produced clicks of low directionality, longer duration and lower frequency than did adults in the wild [385].
It is interesting to note that with such high click repetition rates of up to 1600 clicks/s, depending on sampling frequency and Fourier parameters, squeals will appear as tonal (CW and FM) sounds with sidebands in spectrograms and have been classified as typical odontocete burst-pulse sounds [382]. Squeals were only recorded during social behaviour and hence might serve a social communication role.
Click emission is directional (directivity index 27 dB, [358, 359]), i.e. the angle of measurement from the head affects the click spectrum. Computed source levels are higher on-axis than off-axis. Furthermore, usual clicks appear mono-pulsed on-axis (with 40 dB more energy in the main pulse than following pulses) and multi-pulsed off-axis [358, 386, 387]. With the general uncertainty about the orientation of the whale at the time of recording, differences in published source levels and spectral characteristics of usual clicks can be explained. Example sperm whale clicks recorded by CMST are shown in Fig. 29.

Sperm whale clicks recorded by CMST on Australia’s Northwest Shelf, April 2011 [fs = 192 kHz, NFFT = 768, 50% overlap, Hanning window]
Berardius arnuxii—Arnoux’s Beaked Whale
Arnoux’s beaked whale is circumpolar in the southern hemisphere, occurring in cold waters from the ice edge to its northernmost stranding reported at \(34^{\circ }\hbox {S}\). A small number of strandings have occurred in SA, WA, TAS and the subantarctic. There have been possible sightings near shore in SA and NSW and confirmed sightings in the Australian Antarctic Territory [388].
Dolphin-like clicks and squeaks from individuals with their heads exposed from the water were reported, but recordings were not taken at the time [389]. Click trains (12–19 kHz, 1.2 s/train, 25 clicks/train), burst-pulse sounds (3–11 kHz, 0.53 s), AM sounds (1–8.5 kHz, 0.77 s) and whistles (4.3–5.2 kHz plus higher-frequency harmonics, 0.65 s) were recorded off Antarctica (Fig. 30; [390]).

Arnoux’s beaked whale sounds: a, b whistles and c burst-pulse sounds [390] [fs = 192 kHz, NFFT = 768, 50% overlap, Hanning window]
Hyperoodon planifrons—Southern Bottlenose Whale
The southern bottlenose whale occurs circumpolar in deep water, with recorded sightings and strandings south of \(30^{\circ }\hbox {S}\). Strandings in Australia have occurred in WA, SA, VIC, TAS and NSW [147]. Click trains with peak energy at 18 kHz and click repetition rates of 17–200 clicks/s were attributed to this species based on nearby visuals in the Southern Ocean [391, 392].
Indopacetus pacificus—Longman’s Beaked Whale
Longman’s beaked whales have been reported from the Indo-Pacific region [393, 394]. Sightings and strandings in Australia are rare with only one record in QLD. This species is expected to occur in the warmer waters of Australia. In Hawaii, clicks (10–60 kHz, 90–340 \(\upmu \)s, 0.1–0.9-s ICI), an FM upsweep click (10–60 kHz, 100–500 \(\upmu \)s, 0.3–0.4-s ICI) and burst-pulse sounds (9–25 kHz, 0.5-s duration, 240 pulses/s) were recorded [13, 395].
Mesoplodon bowdoini—Andrew’s Beaked Whale
Distribution of Andrew’s beaked whales is thought to be circumpolar, though that is unconfirmed. Strandings have occurred off Australia in WA, VIC, TAS and NSW, as well as off New Zealand, Macquarie Island, the Falkland Islands, Argentina, Uruguay and Tristan da Cunha [396]. No reports of vocalisations by this species have been found in the literature.
Mesoplodon densirostris—Blainville’s Beaked Whale, Dense-beaked Whale
Blainville’s beaked whales are oceanic animals, occurring globally in tropical to temperate waters. They are very rarely seen at sea, yet strandings have been reported in all Australian states [147]. These animals embark on deep foraging dives of up to 45-min duration to 1250 m depth followed by a sequence of short and shallow resting dives [397], hence spending little time at the surface where they could otherwise be easily observed.
Blainville’s beaked whales have been recorded off the Canary Islands [398,399,400,401], in the Bahamas [402, 403], off Hawaii [13, 404] and off the Northern Mariana Islands [13].
They emit high-frequency FM clicks, where each click of 0.3–1 ms sweeps upwards from about 24 to 51 kHz [13]. During foraging, FM echolocation clicks are produced in long trains, including speed-up click trains (0.2–0.4-s ICI, 15–60 s/train), terminating in buzzes (400 clicks/buzz, 3 s/buzz) indicating a prey-capture attempt. Source levels are 200–220 dB re 1 \(\upmu \)Pa pp @ 1 m [399,400,401, 403, 405, 406]. Blainville’s beaked whales mostly do not echolocate during shallow dives or during deep descents and ascents, clicking vividly only at deep foraging depths. They were reported silent for 80% of time spent in water shallower than 170 m [398].
Blainville’s beaked whales also emit tonal CW and FM sounds (8–18 kHz fundamental, 0.2–2-s duration), as well as AM or rapidly pulsed sounds (2–80 kHz, 0.1–0.6 s) with source levels of 123–149 dB re 1 \(\upmu \)Pa rms @ 1 m, likely for communication [398, 402, 404]. There are no recordings from Australia with positive species identification.
Mesoplodon ginkgodens—Ginkgo-Toothed Beaked Whale, Ginkgo-toothed Whale, Ginkgo Beaked Whale
Ginkgo-toothed whales are rarely seen at sea; their distribution can only be inferred from strandings that have occurred along the tropical to temperate coasts of the Indian and Pacific Oceans. There have been a few stranding events in Australia, in southern NSW and VIC [407]. No records of sounds produced by Ginkgo-toothed beaked whales were found in the literature.
Mesoplodon grayi—Gray’s Beaked Whale
Gray’s beaked whales are distributed around the southern hemisphere in temperature waters with most sightings reported from south of \(30^{\circ }\hbox {S}\). Many stranding records have occurred along the coastlines of WA, SA, VIC, TAS and NSW as well as New Zealand, South Africa, Argentina, Chile and Peru [408]. Waters between New Zealand’s south island and the Chatham Islands are considered to be of high abundance. No reports of vocalisations by this species were found in the literature.
Mesoplodon hectori—Hector’s Beaked Whale
This species has a potentially circumpolar distribution in cool temperate waters of the southern hemisphere between \(35^{\circ }\) and \(55^{\circ }\hbox {S}\). There are no records from the northern hemisphere. Sightings are rare (with a small number recorded in WA, SA and TAS; [147]), and there are no reports of vocalisations.
Mesoplodon layardii—Strap-Toothed Beaked Whale
Strap-toothed beaked whales are distributed in cool temperate waters of the southern hemisphere between \(30^{\circ }\hbox {S}\) and the Antarctic Convergence. They may occur south of \(38^{\circ }\hbox {S}\) year-round moving north of \(38^{\circ }\hbox {S}\) seasonally, and they tend to occur in deep waters off the continental shelf [408]. This species is the most commonly stranded beaked whale species in Australia with stranding events recorded in WA, SA, VIC, TAS, NSW, QLD, Macquarie Island and Heard Island. There is only one mention of vocalisations from this species. This came from a stranded individual that reportedly “squealed” when attempts were made to drag it back to the ocean, but no acoustic characteristics were specified [409].
Mesoplodon mirus—True’s Beaked Whale
In the southern hemisphere, this species has stranded in the southern Indian Ocean, in South Africa, New Zealand, WA, VIC and TAS (though in very low numbers). In the northern hemisphere, records only exist from the North Atlantic. The species has not been recorded in the North Pacific, South Atlantic or northern Indian Ocean and appears to avoid tropical waters [408]. No reports of vocalisations of this species were available.
Tasmacetus shepherdi—Shepherd’s Beaked Whale
Few sightings and strandings of Shepherd’s beaked whale have been reported, with some located off New Zealand, southeast Australia (TAS and VIC), Argentina and islands around Tristan da Cunha. The species is thought to prefer subantarctic and temperate deep oceanic waters [408, 410]. No records of vocalisations were found.
Ziphius cavirostris—Cuvier’s Beaked Whale, Goose-Beaked Whale
Cuvier’s beaked whales occur in all the world’s oceans except for high-latitude polar waters, and they have stranded on all Australian coasts. Globally, they are considered to be one of the most abundant beaked whale species with the most extensive range [147, 411].
They are an oceanic species and have been acoustically recorded in the Mediterranean and Ligurian Seas [399, 412, 413] as well as various locations across the North Pacific [13]. They hold the marine mammal record for deep diving down to 3 km depth over 140 minutes [414]. They echolocate only when foraging at depth (>475 m; [399]). Their clicks are FM upsweeps from 13 to 64 kHz over 0.2–1.6 ms with an ICI of 0.1–0.5 s and source levels of 214 dB re 1 \(\upmu \)Pa pp @ 1 m or 164 dB re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m [13, 399, 413]. Click trains last 16–45 s [412]. Buzzes are rapid clicks (up to 250 clicks/s) and are heard only at the end of foraging click trains, marking the prey-capture event [399]. There are no confirmed recordings from Australia.
The Sounds of Sirenians—Sea Cows
Dugong dugon—Dugong
Sirenians are found in shallow, inshore waters of tropical and subtropical climate worldwide. The dugong is the only species of Sirenia found in Australian waters. Distributed across the northern, tropical, inshore waters (up to 40 m depth) from Shark Bay (WA), along Australia’s northern coast, to Moreton Bay (QLD), dugongs are also occasional visitors to NSW coastal and estuarine waters [415, 416].
Dugong sounds (Fig. 31) have been recorded on both east and west coasts of Australia. They make FM sounds with 2–4.7 kHz fundamentals, lasting 0.1–2.2 s with source levels of 131–161 dB re 1 \(\upmu \)Pa rms @ 1 m, 145–180 dB re 1 \(\upmu \)Pa pp @ 1 m and 110–146 dB re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m [417,418,419]. Their pulsed or AM sounds cover 300 Hz–22 kHz, last 0.1–1.2 s and have source levels of 129–163 dB re 1 \(\upmu \)Pa rms @ 1 m, 148–172 dB re 1 \(\upmu \)Pa pp @ 1 m and 110–135 dB re 1 \(\upmu \hbox {Pa}^{2}\)s @ 1 m [417, 418]. Broadband feeding (mastication) sounds ranging from 1 to over 22 kHz were recorded in Thailand [420].

Dugong sounds: a) whistle and b) burst-pulse sound [418] [fs = 48 kHz, NFFT = 512, 50% overlap, Hanning window]
The Sounds of Pinnipeds—Seals and Sea Lions
Arctocephalus forsteri—New Zealand Fur Seal
New Zealand fur seals, also known as long-nosed fur seals or South Australian fur seals, are found in subantarctic waters. They occur around both the south and north islands of New Zealand (up to 72 m depth [421]) and the coastal waters of SA and WA, with small populations in VIC, NSW and TAS [422, 423].
New Zealand fur seals have not been recorded underwater. In-air vocalisations are mostly of pulsed nature, and even FM sounds are typically amplitude-modulated (Fig. 32). Sounds have been classified as pup-attraction calls produced by females (burst-pulse sounds, 100–8000 Hz, 0.8–2.2 s), female-attraction calls produced by pups (burst-pulse sounds, 140–4600 Hz, 0.5–2.0 s), as well as guttural threats, growls preceding guttural threats, barks and a full-threat call, all of which were produced by males (pulsed sounds, 85–7800 Hz, 0.1–2 s) [424,425,426].
New Zealand fur seals breed sympatrically and hybridise with other fur seals (A. gazella and A. tropicalis) at subantarctic islands such as Macquarie Island. These hybrid fur seals produce vocalisations that have mixed traits and are intermediate to their parental species’ calls [425,426,427].

New Zealand fur seal sounds in air: a, b FM, c burst-pulse sounds recorded by UNSW on the Otago Peninsula [fs = 44 kHz, NFFT = 4410, 50% overlap, Hanning window]
Arctocephalus gazella—Antarctic Fur Seal
Antarctic fur seals are found in subantarctic waters. The majority of the Antarctic fur seal population is located on South Georgia Island. Other smaller populations are found on islands off South America and in the southern Indian Ocean. They are seen as vagrants in the Antarctic and along the coastlines of Australia, Argentina, Brazil, Chile and South Africa [428].
Antarctic fur seal calls have only been recorded in air. Sounds are of FM or pulsed nature (Fig. 33). Barks (100–7700 Hz) last 0.1–1.1 s/pulse and 5.6 s/bout. Guttural threat sounds cover a similar frequency band (85–6000 Hz) and last 0.5–4 s. AM sounds such as growls, pup-attraction calls and female-attraction calls cover 100–9000 Hz and last 0.2–2.1 s [424,425,426, 429]. The latter two call types are individually distinctive [426, 430].
The Antarctic fur seal breeds sympatrically and hybridises with other fur seals (A. forsteri and A. tropicalis) at subantarctic islands such as Macquarie Island. These hybrids produce vocalisations that are intermediate to their parental species’ vocalisations [425, 427].

Antarctic fur seal sounds in air recorded by UNSW in South Georgia in 2008: a, b FM, c burst-pulses [fs = 44 kHz, NFFT = 2205 (a, b), NFFT = 882 (c), 50% overlap, Hanning window]
Arctocephalus pusillus doriferus—Australian Fur Seal
There are two species of A. pusillus: the Cape fur seal (A.p. pusillus) and the Australian fur seal (A.p. doriferus). Australian fur seals are found in coastal waters of eastern SA, VIC, TAS and southern NSW. Breeding colonies exist at islands of Bass Strait, between TAS and VIC [431,432,433].
There is no record of Australian fur seal calls under water. In air, these animals mostly produce mostly pulsed vocalisations (Fig. 34). Male and female barks have most energy between 100 and 2400 Hz with each pulse of the bark lasting 0.1–0.2 s. Male and female guttural threat pulses are longer (0.14–0.3 s) but span a similar frequency band (40–3500 Hz). Burst-pulse sounds, including AM growls, pup-attraction and female-attraction calls, cover 100–5500 Hz in frequency and 0.2–1.8 s in duration [424, 434, 435]. Australian fur seal pups produce individually distinct calls to attract their mothers (female-attraction call) throughout the maternal dependence period [436].

Australian fur seal barks in air recorded by Joy Tripovich in VIC [fs = 22 kHz, NFFT = 1100, 50% overlap, Hanning window]
Arctocephalus tropicalis—Subantarctic Fur Seal
Subantarctic fur seals are widely distributed throughout the southern hemisphere and found in the southern waters of the Indian, Pacific and Atlantic Oceans. They breed on subantarctic islands such as Macquarie Island. Vagrants have been recorded on the coasts of Antarctica, Australia (from WA to QLD), New Zealand, southern South America and southern Africa [437, 438].
There are no records of subantarctic fur seals vocalising under water. On land, they produce pulsed calls: barks (120–3020 Hz, 0.07–0.2 s/pulse, 5–8 s/bout), guttural challenge calls (250–1930 Hz, 0.13–1.56 s) and burst-pulse sounds including pup-attraction calls, female-attraction calls and full-threat calls (100–8000 Hz, 0.2–3.5 s) [425, 426, 429, 439,440,441]. Individual identity has been demonstrated in mothers and pups [439, 440]. On Macquarie Island, subantarctic fur seals breed sympatrically and hybridise with other fur seals (A. forsteri and A. gazella). The vocalisations of hybrids have mixed traits from both their parental species [425, 427].
Neophoca cinerea—Australian Sea Lion
Australian sea lions are found in shallow waters along the southern and south-western coasts of Australia. They are endemic to Australia, distributed from the Houtman Abrolhos Islands (WA) to The Pages Island (SA; [442]), in waters up to 105 m depth [443]. The species has been known to venture to the southern coast of eastern Australia. Sixty-six Australian sea lion colonies have been recorded (38 in SA, 28 in WA). The four largest colonies are located on The Pages Island, at Seal Bay on Kangaroo Island and at Dangerous Reef [444].
There are no recordings of Australian sea lion sounds underwater. Their in-air sounds were first described in the early 1970s [445, 446]. In air, on their haul-out locations, males produce loud barks consisting of pulses with peak energy between 350 and 2600 Hz, lasting 0.02–0.1 s [441, 447, 448] (Fig. 35). Bark series that show accelerated rhythmicity and higher formants elicited stronger responses from other males [449]. A bleating burst-pulse sound (400–4000 Hz, 0.5 s) was also reported [441]. Mothers and pups produce FM sounds peaking between 400 and 3400 Hz, lasting 0.55–1.4 s [450]. Vocalisations of females and males are individually distinctive [441, 450,451,452,453]. Acoustic mother-offspring recognition is well developed in pinnipeds, with Australian sea lions being one example. Geographic variation along the southern WA and SA coasts was investigated by Ahonen et al. [448].

Australian sea lion barks in air recorded by Joy Tripovich in SA [fs = 22 kHz, NFFT = 1100, 50% overlap, Hanning window]

Leopard seal underwater burst-pulse sounds recorded by UNSW in the Davis Sea (Eastern Antarctica) in October 1996 [fs = 11 kHz, NFFT = 2000, 50% overlap, Hanning window]
Hydrurga leptonyx—Leopard Seal
Leopard seals are found in the Antarctic pack ice and on subantarctic islands. They are difficult to survey in the Antarctic, because they are widely dispersed at low densities throughout the Antarctic pack ice [454,455,456,457,458]. Vagrants are sighted in the austral winter on southern continents as far north as the coasts of QLD and WA [459].
Leopard seals produce vocalisations underwater particularly during their breeding season in the austral summer [455, 460]. In-air vocalisations have not been reported. Most of the underwater vocalisations consist of rapid pulse trains, commonly referred to as trills that are typically given in two parts (double trills; Fig. 36). Trills cover a frequency range of 150 Hz to 6 kHz and last 2–8 s with source levels of 156–175 dB re 1 \(\upmu \)Pa rms @ 1 m [459, 461,462,463]. These seals also produce lower-frequency burst-pulse sounds including AM growls at 50 Hz–6 kHz lasting 0.5–4.3 s [459, 462]. Ultrasonic (64–160 kHz) pulse trains consisting of rapid (9–40 ms) downsweeps were recorded in captivity and are the only account of such high-frequency sounds in pinnipeds [464]. Leopard seal vocalisations have individual [465] and age-related [466] differences, as well as geographic variation [462, 467, 468].
Leptonychotes weddellii—Weddell Seal
The Weddell seal is the world’s most southerly breeding mammal. It breeds on the fast ice around Antarctica where it occurs right up to the shoreline of the Antarctic continent. It also occurs offshore in the pack ice zone north to the seasonally shifting limits of the Antarctic Convergence. Vagrants have been recorded in South America, New Zealand and southern Australia [469,470,471].
Weddell seals produce a wide array of both in-air and underwater vocalisations. The majority of calling occurs during the breeding period, with vocalisation numbers dropping afterwards [472, 473]. Weddell seal calls were first described crudely phonetically by Weddell in 1825, then by Lindsey [474] and by Schevill and Watkins [475]. Different studies have used different descriptors and terminology making comparisons difficult. Weddell seals underwater make CW sounds at frequencies ranging from 100 to 1600 Hz, lasting 1–90 s [15, 476,477,478]. They make a variety of FM sounds, commonly downsweeps, between 100 Hz and 14 kHz, lasting 0.2–67 s, with source levels of 153–193 dB re 1 \(\upmu \)Pa rms @ 1 m (Fig. 37; [15, 476,477,478,479]). Pulse trains and AM sounds cover a band from 400 Hz to 15 kHz and last 1–54 s with source levels of 171–188 dB re 1 \(\upmu \)Pa rms @ 1 m [15, 476,477,478,479]. Pulsed sounds have distinct rhythmic patterns and can be emitted in constant, increasing as well as decreasing IPI [480]. In-air, FM calls between 100 and 500 Hz, lasting 0.25–2 s, commonly of convex spectrographic shape, have been recorded of mothers and pups [481, 482]. Individual [481,482,483] and geographic [477, 479, 484] variability of aerial and underwater vocalisations has been noted.

Weddell seal underwater FM sounds recorded by UNSW in the Davis Sea (Eastern Antarctica) in October 1996 [fs = 11 kHz, NFFT = 200 (a, b), NFFT = 500 (c), 50% overlap, Hanning window]
Lobodon carcinophaga—Crabeater Seal
Crabeater seals are found year-round in Antarctic circumpolar pack ice up to 528 m depth [485], and migrants are found as far north as Australia, South Africa and New Zealand.
Crabeater seal underwater vocalisations are mostly pulsed and include moans and groans (Fig. 38). There are lower-frequency calls with peak energy around a few hundred Hertz and higher-frequency calls with peak energy above 1 kHz. Together, these sounds cover a frequency range from 10 Hz to 6 kHz and last about 2 s with a range of 0.1–6 s [461, 486, 487]. Whistles have also been reported at 900–1200 Hz lasting about 1 s [486].

Crabeater seal underwater burst-pulse sound recorded by UNSW in the Davis Sea (Eastern Antarctica) in October 1996 [fs = 11 kHz, NFFT = 550, 50% overlap, Hanning window]
Mirounga leonina—Southern Elephant Seal
Elephant seals are found in subpolar waters worldwide. There are two species of elephant seal, but the southern elephant seal (M. leonina) is the only species in the southern hemisphere. Southern elephant seals are distributed across subantarctic waters north of the pack ice up to 1500 m depth [488] from Macquarie Island in Australia to the tip of the Antarctic continent [489].
To date, underwater vocalisations have not been recorded. Their aerial vocalisations were recorded on Sea Lion Island, one of the Falkland Islands [490,491,492]. Sounds consisted of broadband (10 Hz to greater than 4 kHz) pulses emitted in bouts with variable IPI (Fig. 39). Most energy lay between 200 and 800 Hz, and vocalisations lasted between 1 and 50 s with source levels of 103–120 dB re 20 \(\upmu \)Pa rms @ 1 m.

Southern elephant seal aerial pulsed call recorded by UNSW in the Western Antarctic Peninsula in 2008 [fs = 44 kHz, NFFT = 900, 50% overlap, Hanning window]
Ommatophoca rossii—Ross Seal
Ross seals are found in circumpolar pack ice about Antarctica. They are distributed in the Antarctic pack ice during the moult and breeding season and migrate north of the pack ice after moulting where they establish a pelagic lifestyle through the year [493]. Vagrant animals have been seen along the coastlines of the southern hemisphere including South Australia.
Ross seals vocalise both in air and under water with first recordings described by Ray [494]. Their vocalisations include pulses that are very brief downsweeps from 1000 to 100 Hz, lasting 0.05–0.1 s/pulse with 5–12 pulses in sequence—both in air and under water. They also produce U-shaped FM calls in both media, lasting 1–1.5 s. The frequency range in air is 100–800 Hz and higher (1000–4500 Hz) under water (Fig. 40). Underwater tones are amplitude-modulated creating sidebands at 650 Hz spacing [14].

Ross seal underwater siren recorded by UNSW in the Davis Sea (Eastern Antarctica) in October 1996 [fs = 22 kHz, NFFT = 440, 50% overlap, Hanning window]
Discussion
We intend this article to be used by researchers tasked with acoustically identifying species recorded in Australian waters. The first step should be to determine which species could have been present at the time of recording. A good starting place is the Department of the Environment and Energy website, which gives distribution and migration patterns for most species and was one of the main resources used for this study. Figure 1 summarises the geographic regions where the various species might be encountered. In addition, Table 1 indicates which species mostly stay offshore versus inshore. Figures 2, 3 and 4 can then be used to compare the bandwidth and duration of the signals recorded to the summary data for each species. More details on the sounds emitted by the resulting candidate species can be found in Tables 2, 3, 4 and Sects. 5 through 8, potentially leading to positive species identification.
There is an increasing amount of literature on the sounds of marine mammals in the world’s oceans. As hardware and software tools have advanced, the certainty of measured quantities has also improved. For example, in the case of echolocation clicks, sampling frequencies are now high enough to instil confidence that the full spectrum was captured in many recordings. In addition, the use of hydrophone arrays allows the determination of range and sometimes angular aspect to the animal so that researchers know whether spectra were recorded on- versus off-axis. What is missing though is a unified approach to analysing and reporting data, or the development of standards, so that results from different studies can be combined or compared [495].
Perhaps related to this is a lack of agreement on how to classify animal sounds. The biggest hurdle in completing this manuscript was the need to summarise information from hundreds of publications and the corresponding dilemma of whether to group two sounds as “the same” or to separate them. Onomatopoeic names have different connotations and imply different features for different people. Spectrographic representations are affected by various recording and analysis settings. The effect of sound propagation on altering received call structure from that transmitted is almost always ignored but hardly ever negligible. Rather than classifying sounds by a variety of characteristics that are recorded by specific gear at some range and angle from the animal, it might be less arbitrary to group them according to how they are produced. This requires more research into actual sound production, an understanding of resonances and filters and sound propagation through animal tissues. Until such time, it would at least be helpful to have consensus about minimum standards for the recording, analysis and reporting of animal sounds.
Symbols and Abbreviations
Sound types | CW | Constant-wave, flat tonal sound |
FM | Frequency-modulated | |
AM | Amplitude-modulated | |
Pulses | Broadband sounds; typically brief; rapid series of pulses are often classified as AM sounds | |
Sound measurements | fp | Frequency of fundamental contour of whistles; frequency of maximum spectral power in clicks |
fc | Centroid frequency that divides the power in a spectrum into two equal parts | |
bw3 | \(-\)3 dB bandwidth | |
bw10 | \(-\)10 dB bandwidth | |
bwrms | rms bandwidth, i.e. the standard deviation of the spectrum about the centroid frequency | |
SL | Source level | |
pp | Peak-to-peak | |
rms | Root-mean-square | |
IPI | Inter-pulse interval | |
ICI | Inter-click interval | |
Recording and analysis parameters | fs | Sampling frequency |
NFFT | Number of Fourier components | |
Specific sounds | PAC | Pup-attraction call made by female pinnipeds |
FAC | Female-attraction call made by pinniped pups | |
Geographic references | N | North |
S | South | |
E | East | |
W | West | |
WA | Western Australia | |
SA | South Australia | |
VIC | Victoria | |
TAS | Tasmania | |
NSW | New South Wales | |
QLD | Queensland | |
NT | Northern Territory | |
Isl. | Island | |
Institutions | CMST | Centre for Marine Science & Technology, Curtin University |
CWR | Centre for Whale Research | |
UNSW | University of New South Wales |
Summary Tables of Marine Mammal Sounds
References
Tyack, P.L., Clark, C.W.: Communication and acoustic behavior of dolphins and whales. In: Au, W.W.L., Popper, A.N., Fay, R.R. (eds.) Hearing by Whales and Dolphins, pp. 156–224. Springer, New York (2000)
Au, W.W.L.: The Sonar of Dolphins. Springer, New York (1993)
Dunlop, R.A., Cato, D.H., Noad, M.J.: Non-song acoustic communication in migrating humpback whales (Megaptera novaeangliae). Mar. Mamm. Sci. 24(3), 613–629 (2008). doi:10.1111/j.1748-7692.2008.00208.x
Erbe, C.: Underwater passive acoustic monitoring & noise impacts on marine fauna—a workshop report. Acoust. Aust. 41(1), 113–119 (2013)
Erbe, C., McCauley, R., Gavrilov, A., Madhusudhana, S., Verma, A.: The underwater soundscape around Australia. In: Proceedings of Acoustics 2016, 9–11 November 2016, Brisbane, Australia (2016)
Erbe, C., Verma, A., McCauley, R., Gavrilov, A., Parnum, I.: The marine soundscape of the Perth Canyon. Prog. Oceanogr. 137, 38–51 (2015). doi:10.1016/j.pocean.2015.05.015
Kaplan, M.B., Mooney, T.A., Sayigh, L.S., Baird, R.W.: Repeated call types in Hawaiian melon-headed whales (Peponocephala electra). J. Acoust. Soc. Am. 136(3), 1394–1401 (2014). doi:10.1121/1.4892759
Sayigh, L., Quick, N., Hastie, G., Tyack, P.: Repeated call types in short-finned pilot whales, Globicephala macrorhynchus. Mar. Mamm. Sci. 29(2), 312–324 (2013). doi:10.1111/j.1748-7692.2012.00577.x
Murray, S.O., Mercado, E., Roitblat, H.L.: Characterizing the graded structure of false killer whale (Pseudorca crassidens) vocalizations. J. Acoust. Soc. Am. 104(3), 1679–1688 (1998). doi:10.1121/1.424380
Watkins, W.A.: The harmonic interval: fact or artifact in spectral analysis of pulse trains. In: Tavolga, W.N. (ed.) Marine Bio-Acoustics, vol. 2, pp. 15–43. Pergamon Press, New York (1967)
Knowlton, A.R., Clark, C.W., Kraus, S.D.: Sounds recorded in the presence of sei whales, Balaenoptera borealis. J. Acoust. Soc. Am. 89(4, Pt. 2), 1968–1968 (1991). doi:10.1121/1.2029710
Thompson, T.J., Winn, H.E., Perkins, P.J.: Mysticete sounds. In: Winn, H.E., Olla, B.L. (eds.) Behavior of Marine Animals, vol. 3: cetaceans, pp. 403–431. Plenum Press, New York (1979)
Baumann-Pickering, S., McDonald, M.A., Simonis, A.E., Solsona Berga, A., Merkens, K.P.B., Oleson, E.M., Roch, M.A., Wiggins, S.M., Rankin, S., Yack, T.M., Hildebrand, J.A.: Species-specific beaked whale echolocation signals. J. Acoust. Soc. Am. 134(3), 2293–2301 (2013). doi:10.1121/1.4817832
Watkins, W.A., Ray, G.C.: In air and underwater sounds of the Ross seal, Ommatophoca rossi. J. Acoust. Soc. Am. 77(4), 1598–1600 (1985). doi:10.1121/1.392003
Terhune, J.M.: Weddell seals do not lengthen calls in response to conspecific masking. Bioacoustics 25(1), 75–88 (2016). doi:10.1080/09524622.2015.1089791
Wellard, R., Erbe, C., Fouda, L., Blewitt, M.: Vocalisations of killer whales (Orcinus orca) in the Bremer Canyon, Western Australia. PLoS ONE 10(9), e0136535 (2015). doi:10.1371/journal.pone.0136535
Bannister, J.L., Kemper, C.M., Warneke, R.M.: The Action Plan for Australian Cetaceans. Australian Nature Conservation Agency, Canberra (1996)
Department of Sustainability: Environment, Water, Population and Communities: Conservation Management Plan for the Southern RightWhale, Canberra, ACT, Australia (2012)
Bannister, J.L.: Annual Reports on Aerial Surveys of Southern Right Whales on the Southern Australian Coastline. Western Australian Museum, Perth, WA (1979–2005)
Burnell, S.R.: The Population Biology of Southern Right Wales in Southern Australian Waters. Ph.D. Thesis, Department of Veterinary Anatomy and Pathology, Faculty of Veterinary Science, University of Sydney (1999)
Clark, C.W.: The acoustic repertoire of the southern right whale, a quantitative analysis. Anim. Behav. 30(4), 1060–1071 (1982). doi:10.1016/S0003-3472(82)80196-6
Cummings, W.C., Fish, J.F., Thompson, P.O.: Sound production and other behavior of southern right whales, Eubalaena glacialis. Trans. San Diego Soc. Nat. Hist. 17(1), 1–13 (1972)
Payne, R., Payne, K.: Underwater sounds of southern right whales. Zoologica 56(4), 159–165 (1972)
Dombroski, J.R.G., Parks, S.E., Groch, K.R., Flores, P.A.C., Sousa-Lima, R.S.: Vocalizations produced by southern right whale (Eubalaena australis) mother-calf pairs in a calving ground off Brazil. J. Acoust. Soc. Am. 140(3), 1850–1857 (2016). doi:10.1121/1.4962231
Webster, T.A., Dawson, S.M., Rayment, W.J., Parks, S.E., Van Parijs, S.M.: Quantitative analysis of the acoustic repertoire of southern right whales in New Zealand. J. Acoust. Soc. Am. 140(1), 322–333 (2016). doi:10.1121/1.4955066
Ward, R., Charlton, C., Guggenheimer, S.: Southern Right Whale (Eubalaena australis) acoustics at Fowlers Bay, South Australia. Centre for Marine Science and Technology, Curtin University, Perth (2014)
Parks, S.E., Clark, C.W., Tyack, P.L.: Short- and long-term changes in right whale calling behavior: the potential effects of noise on acoustic communication. J. Acoust. Soc. Am. 122(6), 3725–3731 (2007). doi:10.1121/1.2799904
Gill, P.C., Kemper, C.M., Talbot, M., Lyons, S.A.: Large group of pygmy right whales seen in a shelf upwelling region off Victoria, Australia. Mar. Mamm. Sci. 24(4), 962–968 (2008). doi:10.1111/j.1748-7692.2008.00220.x
Kemper, C.M.: Distribution of the pygmy right whale, Caperea marginata, in the Australasian region. Mar. Mamm. Sci. 18(1), 99–111 (2002). doi:10.1111/j.1748-7692.2002.tb01021.x
Reilly, S.B., Bannister, J.L., Best, P.B., Brown, M., Brownell, Jr. R.L., Butterworth, D.S., Clapham, P.J., Cooke, J., Donovan, G.P., Urbán, J., Zerbini, A.N.: Caperea marginata. The IUCN Red List of Threatened Species, e.T3778A10071743 (2008). doi:10.2305/IUCN.UK.2008.RLTS.T3778A10071743.en
Dawbin, W.H., Cato, D.H.: Sounds of a pygmy right whale (Caperea marginata). Mar. Mamm. Sci. 8(3), 213–219 (1992). doi:10.1111/j.1748-7692.1992.tb00405.x
Department of the Environment: Balaenoptera acutorostrata unnamed subsp. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Beamish, P., Mitchell, E.: Short pulse length audio frequency sounds recorded in the presence of a minke whale (Balaenoptera acutorostrata). Deep Sea Res. 20(4), 375–386 (1973). doi:10.1016/0011-7471(73)90060-0
Edds-Walton, P.L.: Vocalizations of minke whales Balaenoptera acutorostrata in the St. Lawrence estuary. Bioacoustics 11(1), 31–50 (2000)
Risch, D., Clark, C.W., Dugan, P.J., Popescu, M., Siebert, U., Van Parijs, S.M.: Minke whale acoustic behavior and multi-year seasonal and diel vocalization patterns in Massachusetts Bay, USA. Mar. Ecol. Prog. Ser. 489, 279–295 (2013). doi:10.3354/meps10426
Risch, D., Siebert, U., Van Parijs, S.M.: Individual calling behaviour and movements of North Atlantic minke whales (Balaenoptera acutorostrata). Behaviour 151(9), 1335–1360 (2014). doi:10.1163/1568539X-00003187
Mellinger, D.K., Carson, C.D., Clark, C.W.: Characteristics of minke whale (Balaenoptera acutorostrata) pulse trains recorded near Puerto Rico. Mar. Mamm. Sci. 16(4), 739–756 (2000). doi:10.1111/j.1748-7692.2000.tb00969.x
Winn, H.E., Perkins, P.J.: Distribution and sounds of the minke whale, with a review of mysticete sound. Cetology 19, 1–12 (1976)
Rankin, S., Barlow, J.: Source of the North Pacific ’boing’ sound attributed to minke whales. J. Acoust. Soc. Am. 118(5), 3346–3351 (2005). doi:10.1121/1.2046747
Thompson, P.W.F.: A long term study of low frequency sounds from several species of whales off Oahu, Hawaii. Cetology 45, 1–19 (1982)
Oswald, J.N., Au, W.W.L., Duennebier, F.: Minke whale (Balaenoptera acutorostrata) boings detected at the Station ALOHA Cabled Observatory. J. Acoust. Soc. Am. 129(5), 3353–3360 (2011). doi:10.1121/1.3575555
Norris, T.F., Dunleavy, K.J., Yack, T.M., Ferguson, E.L.: Estimation of minke whale abundance from an acoustic line transect survey of the Mariana Islands. Mar. Mamm. Sci. (2017). doi:10.1111/mms.12397
Gedamke, J., Costa, D.P., Dunstan, A.: Localization and visual verification of a complex minke whale vocalization. J. Acoust. Soc. Am. 109(6), 3038–3047 (2001). doi:10.1121/1.1371763
McCauley, R.D.: Woodside Kimberley Sea Noise Logger Program, September 2006 to June 2010: Whales, Fish and Man-Made Noise. Centre for Marine Science & Technology, Curtin University, Perth (2011)
Department of the Environment: Balaenoptera bonaerensis. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Zerbini, A.N., Secchi, E.R., Siciliano, S., Simões-Lopes, P.C.: A review of the occurrence and distribution of whales of the genus Balaenoptera along the Brazilian coast. Rep. Int. Whal. Comm. 47, 407–417 (1997)
Thiele, D., Gill, P.C.: Cetacean observations during a winter voyage into Antarctic sea ice south of Australia. Antarct. Sci. 11(1), 48–53 (2004). doi:10.1017/S0954102099000073
Leatherwood, S., Thomas, J.A., Awbrey, F.T.: Minke whales off northwestern Ross Island. Antarctic 16, 154–156 (1981)
Risch, D., Gales, N.J., Gedamke, J., Kindermann, L., Nowacek, D.P., Read, A.J., Siebert, U., Van Opzeeland, I.C., Van Parijs, S.M., Friedlaender, A.S.: Mysterious bio-duck sound attributed to the Antarctic minke whale (Balaenoptera bonaerensis). Biol. Lett. 10(4), 20140175 (2014). doi:10.1098/rsbl.2014.0175
Schevill, W.E., Watkins, W.A.: Intense low-frequency sounds from an Antarctic minke whale, Balaenoptera acutorostrata. Breviora 388, 8 (1972)
Dominello, T., Širović, A.: Seasonality of Antarctic minke whale (Balaenoptera bonaerensis) calls off the western Antarctic Peninsula. Mar. Mamm. Sci. 32(3), 826–838 (2016). doi:10.1111/mms.12302
Miller, B.S., Kelly, N., Double, M.C., Childerhouse, S.J., Laverick, S., Gales, N.: Cruise report on SORP 2012 blue whale voyages: development of acoustic methods. (Report No. SC/64/SH11), 16 pp. International Whaling Commission (2012)
Department of the Environment: Balaenoptera borealis. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Baumgartner, M.F., Van Parijs, S.M., Wenzel, F.W., Tremblay, C.J., Esch, H.C., Warde, A.M.: Low frequency vocalizations attributed to sei whales (Balaenoptera borealis). J. Acoust. Soc. Am. 124(2), 1339–1349 (2008). doi:10.1121/1.2945155
Newhall, A., Lin, Y.-T., Lynch, J., Baumgartner, M., Gawarkiewicz, G.: Long distance passive localization of vocalizing sei whales using an acoustic normal mode approach. J. Acoust. Soc. Am. 131(2), 1814–1825 (2012). doi:10.1121/1.3666015
Wang, D., Huang, W., Garcia, H., Ratilal, P.: Vocalization source level distributions and pulse compression gains of diverse baleen whale species in the Gulf of Maine. Rem. Sens. 8(11), 881 (2016). doi:10.3390/rs8110881
Romagosa, M., Boisseau, O., Cucknell, A.-C., Moscrop, A., McLanaghan, R.: Source level estimates for sei whale (Balaenoptera borealis) vocalizations off the Azores. J. Acoust. Soc. Am. (2015). doi:10.1121/1.4930900
Rankin, S., Barlow, J.: Vocalizations of the sei whale Balaenoptera borealis off the Hawaiian Islands. Bioacoustics 16, 137–145 (2007)
Calderan, S., Miller, B., Collins, K., Ensor, P., Double, M., Leaper, R., Barlow, J.: Low-frequency vocalizations of sei whales (Balaenoptera borealis) in the Southern Ocean. J. Acoust. Soc. Am. 136(6), 418–423 (2014). doi:10.1121/1.4902422
McDonald, M.A., Hildebrand, J.A., Wiggins, S.M., Thiele, D., Glasgow, D., Moore, S.E.: Sei whale sounds recorded in the Antarctic. J. Acoust. Soc. Am. 118(6), 3941–3945 (2005). doi:10.1121/1.2130944
Best, P.: Two allopatric forms of Bryde’s whale off South Africa. Rep. Int. Whal. Comm. 1, 10–38 (1977)
Kawamura, A., Satake, Y.: Preliminary report on the geographical distribution of the Bryde’s whale in the North Pacific with special reference to the structure of filtering apparatus. Sci. Rep. Whales Res. Inst. 28, 1–35 (1976)
Bannister, J., Kemper, C.M., Warneke, R.M.: The Action Plan for Australian Cetaceans. Australian Nature Conservation Agency, Canberra (1996)
Rice, A.N., Palmer, K.J., Tielens, J.T., Muirhead, C.A., Clark, C.W.: Potential Bryde’s whale (Balaenoptera edeni) calls recorded in the northern Gulf of Mexico. J. Acoust. Soc. Am. 135(5), 3066–3076 (2014). doi:10.1121/1.4870057
Širović, A., Bassett, H.R., Johnson, S.C., Wiggins, S.M., Hildebrand, J.A.: Bryde’s whale calls recorded in the Gulf of Mexico. Mar. Mamm. Sci. 30(1), 399–409 (2014). doi:10.1111/mms.12036
Cummings, W.C., Thompson, P.O., Ha, S.J.: Sounds from Bryde, Balaenoptera edeni, and finback, B. physalus, whales in the Gulf of California. Fish. Bull. 84(2), 359–370 (1986)
Edds, P.L., Odell, D.K., Tershy, B.R.: Vocalizations of a captive juvenile and free-ranging adult-calf pairs of Bryde’s whales, Balaenoptera edeni. Mar. Mamm. Sci. 9(3), 269–284 (1993)
Viloria-Gómora, L., Romero-Vivas, E., Urbán, R.J.: Calls of Bryde’s whale (Balaenoptera edeni) recorded in the Gulf of California. J. Acoust. Soc. Am. 138(5), 2722 (2015). doi:10.1121/1.4932032
Oleson, E.M., Barlow, J., Gordon, J., Rankin, S., Hildebrand, J.A.: Low frequency calls of Bryde’s whales. Mar. Mamm. Sci. 19, 160–172 (2003)
Figueiredo, L.D., Simao, S.M.: Bryde’s Whale (Balaenoptera edeni) vocalizations from Southeast Brazil. Aquat. Mamm. 40(3), 225–231 (2014). doi:10.1578/AM.40.3.2014.225
Chiu, P.-H.: Studies on the population and acoustic signals of Bryde’s whales (Balaenoptera edeni) in Tosa Bay. National Sun Yat-sen University, Japan (2009)
Heimlich, S.L., Mellinger, D.K., Nieukirk, S.L., Fox, C.G.: Types, distribution, and seasonal occurrence of sounds attributed to Bryde’s whales (Balaenoptera edeni) recorded in the eastern tropical Pacific, 1999–2001. J. Acoust. Soc. Am. 118(3), 1830–1837 (2005). doi:10.1121/1.1992674
Buchan, S.J., Hucke-Gaete, R., Rendell, L., Stafford, K.M.: A new song recorded for blue whales in the Corcovado Gulf, Southern Chile, and an acoustic link to the Eastern Tropical Pacific. Endanger. Species Res. 23, 241–252 (2014). doi:10.3354/esr00566
Samaran, F., Stafford, K.M., Branch, T.A., Gedamke, J., Royer, J.-Y., Dziak, R.P., Guinet, C.: Seasonal and geographic variation of Southern Blue Whale subspecies in the Indian Ocean. PLoS ONE 8(8), e71561 (2013). doi:10.1371/journal.pone.0071561
Branch, T.A., Stafford, K.M., Palacios, D.M., Allison, C., Bannister, J.L., Burton, C.L.K., Cabrera, E., Carlson, C.A., Galletti Vernazzani, B., Gill, P.C., Hucke-Gaete, R., Jenner, K.C.S., Jenner, M.-N.M., Matsuoka, K., Mikhalev, Y.A., Miyashita, T., Morrice, M.G., Nishiwaki, S., Sturrock, V.J., Tormosov, D., Anderson, R.C., Baker, A.N., Best, P.B., Borsa, P., Brownell Jr., R.L., Childerhouse, S., Findlay, K.P., Gerrodette, T., Ilangakoon, A.D., Joergensen, M., Kahn, B., Ljungblad, D.K., Maughan, B., McCauley, R.D., McKay, S., Norris, T.F.: Oman Whale and Dolphin Research Group, Rankin, S., Samaran, F., Thiele, D., van Waerebeek, K., Warneke, R.M.: Past and present distribution, densities and movements of blue whales Balaenoptera musculus in the Southern Hemisphere and northern Indian Ocean. Mamm. Rev. 37(2), 116–175 (2007). doi:10.1111/j.1365-2907.2007.00106.x
Sirovic, A., Hildebrand, J.A., Wiggins, S.M., Thiele, D.: Blue and fin whale acoustic presence around Antarctica during 2003 and 2004. Mar. Mamm. Sci. 25(1), 125–136 (2009). doi:10.1111/j.1748-7692.2008.00239.x
Double, M.C., Andrews-Goff, V., Jenner, K.C.S., Jenner, M.-N., Laverick, S.M., Branch, T.A., Gales, N.J.: Migratory movements of Pygmy Blue Whales (Balaenoptera musculus brevicauda) between Australia and Indonesia as revealed by satellite telemetry. PLoS ONE 9(4), e93578 (2014). doi:10.1371/journal.pone.0093578
Rennie, S., Hanson, C.E., McCauley, R.D., Pattiaratchi, C., Burton, C., Bannister, J., Jenner, C., Jenner, M.N.: Physical properties and processes in the Perth Canyon, Western Australia: links to water column production and seasonal pygmy blue whale abundance. J. Mar. Syst. 77(1–2), 21–44 (2009). doi:10.1016/j.jmarsys.2008.11.008
McCauley, R.D., Jenner, C., J.L., B., Burton, C.L.K., Cato, D.H., Duncan, A.: Blue whale calling in the Rottnest trench—2000, Western Australia. Centre for Marine Science & Technology, Curtin University, Perth, WA, Australia (2001)
Gill, P.C., Morrice, M.G., Page, B., Pirzl, R., Levings, A.H., Coyne, M.: Blue whale habitat selection and within-season distribution in a regional upwelling system off southern Australia. Mar. Ecol. Prog. Ser. 421, 243–263 (2011). doi:10.3354/meps08914
Cummings, W.C., Thompson, P.O.: Underwater sounds from the blue whale, Balaenoptera musculus. J. Acoust. Soc. Am. 50(4 (pt. 2)), 1193–1198 (1971). doi:10.1121/1.1912752
Kibblewhite, A.C., Denham, R.N., Barnes, D.J.: Unusual low-frequency signals observed in New Zealand waters. J. Acoust. Soc. Am. 41(3), 644–655 (1967)
McDonald, M.A., Mesnick, S.L., Hildebrand, J.A.: Biogeographic characterisation of blue whale song worldwide: using song to identify populations. J. Cetacean Res. Manag. 8(1), 55–65 (2006)
Alling, A., Dorsey, E.M., Gordon, J.C.D.: Blue whales Balaenoptera musculus off the northeast coast of Sri Lanka. UNEP Mar. Mamm. Tech. Rep. 3, 247–258 (1991)
Samaran, F., Guinet, C., Adam, O., Motsch, J.F., Cansi, Y.: Source level estimation of two blue whale subspecies in southwestern Indian Ocean. J. Acoust. Soc. Am. 127(6), 3800–3808 (2010). doi:10.1121/1.3409479
Sirovic, A., Hildebrand, J.A., Wiggins, S.M.: Blue and fin whale call source levels and propagation range in the Southern Ocean. J. Acoust. Soc. Am. 122(2), 1208–1215 (2007). doi:10.1121/1.2749452
Sirovic, A., Hildebrand, J.A., Wiggins, S.M., McDonald, M.A., Moore, S.E., Thiele, D.: Seasonality of blue and fin whale calls and the influence of sea lee in the Western Antarctic Peninsula. Deep Sea Res. Pt II Top. Stud. Oceanogr. 51(17–19), 2327–2344 (2004). doi:10.1016/j.dsr2.2004.08.005
Gavrilov, A., McCauley, R., Gedamke, J.: Steady inter and intra-annual decrease in the vocalization frequency of Antarctic blue whales. J. Acoust. Soc. Am. 131(6), 4476–4480 (2012). doi:10.1121/1.4707425
Stafford, K.M., Bohnenstiehl, D.R., Tolstoy, M., Chapp, E., Mellinger, D.K., Moore, S.E.: Antarctic-type blue whale calls recorded at low latitudes in the Indian and eastern Pacific Oceans. Deep Sea Res. Pt I Oceanogr. Res. Pap. 51(10), 1337–1346 (2004). doi:10.1016/j.dsr.2004.05.007
Balcazar, N.E., Klinck, H., Nieukirk, S.L., Mellinger, D.K., Klinck, K., Dziak, R.P., Rogers, T.L.: Using calls as an indicator for Antarctic blue whale occurrence and distribution across the southwest Pacific and southeast Indian Oceans. Mar. Mamm. Sci. 33(1), 172–186 (2016). doi:10.1111/mms.12373
Oleson, E.M., Calambokidis, J., Burgess, W.C., McDonald, M.A., LeDuc, C.A., Hildebrand, J.A.: Behavioral context of call production by eastern North Pacific blue whales. Mar. Ecol. Prog. Ser. 330, 269–284 (2007)
McDonald, M.A., Calambokidis, J., Teranishi, A.M., Hildebrand, J.A.: The acoustic calls of blue whales off California with gender data. J. Acoust. Soc. Am. 109(4), 1728–1735 (2001). doi:10.1121/1.1353593
Gavrilov, A., McCauley, R., Salgado-Kent, C., Tripovich, J., Burton, C.: Vocal characteristics of pygmy blue whales and their change over time. J. Acoust. Soc. Am. 130(6), 3651–3660 (2011). doi:10.1121/1.3651817
McCauley, R.D., Jenner, C., Bannister, J.L., Cato, D.H., Duncan, A.J.: Blue whale calling in the Rottnest trench, Western Australia, and low frequency sea noise. In: Annual Conference of the Australian Acoustical Society, Joondalup, Western Australia, 15–17 Nov. 2000. Australian Acoustical Society (2000)
Recalde-Salas, A., Salgado Kent, C.P., Parsons, M.J.G., Marley, S.A., McCauley, R.D.: Non-song vocalizations of pygmy blue whales in Geographe Bay, Western Australia. J. Acoust. Soc. Am. 135(5), EL213–EL218 (2014). doi:10.1121/1.4871581
McCauley, R.D., Jenner, C.K.: Migratory patterns and estimated population size of pygmy blue whales (Balaenoptera musculus brevicauda) traversing the Western Australian coast based on passive acoustics. In: 62nd International Whaling Commission Annual Meeting, Agadir, Morocco 2010. International Whaling Commission (2010)
Tripovich, J.S., Klinck, H., Nieukirk, S.L., Adams, T., Mellinger, D.K., Balcazar, N.E., Klinck, K., Hall, E.J.S., Rogers, T.L.: Temporal segregation of the Australian and Antarctic blue whale call types (Balaenoptera musculus spp.). J. Mammal. 96(3), 603–610 (2015). doi:10.1093/jmammal/gyv065
Balcazar, N.E., Tripovich, J.S., Klinck, H., Nieukirk, S.L., Mellinger, D.K., Dziak, R.P., Rogers, T.L.: Calls reveal population structure of blue whales across the southeast Indian Ocean and southwest Pacific Ocean. J. Mammol. 96(6), 1184–1193 (2015). doi:10.1093/jmammal/gyv126
Miller, B.S., Collins, K., Barlow, J., Calderan, S., Leaper, R., Mark, M., Ensor, P., Olson, P.A., Olavarria, C., Double, M.C.: Blue whale vocalizations recorded around New Zealand: 1964–2013. J. Acoust. Soc. Am. 135(3), 1616–1623 (2014). doi:10.1121/1.4863647
Rankin, S., Ljungblad, D., Clark, C.W., Kato, H.: Vocalisations of Antarctic blue whales, Balaenoptera musculus intermedia, recorded during the 2001/2002 and 2002/2003 IWC/SOWER circumpolar cruises, Area V, Antarctica. J. Cetacean Res. Manag. 7(1), 13–20 (2005)
Wada, S., Oishi, M., Yamada, T.K.: A newly discovered species of living baleen whale. Nature 426(6964), 278–281 (2003). doi:10.1038/nature02103
Sasaki, T., Nikaido, M., Wada, S., Yamada, T.K., Cao, Y., Hasegawa, M., Okada, N.: Balaenoptera omurai is a newly discovered baleen whale that represents an ancient evolutionary lineage. Mol. Phylogenet. Evol. 41(1), 40–52 (2006). doi:10.1016/j.ympev.2006.03.032
Ottewell, K., Coughran, D., Gall, M., Irvine, L., Byrne, M.: A Recent Stranding of Omura’s Whale (Balaenoptera omurai) in Western Australia. Aquat. Mamm. 42(2), 193–197 (2016). doi:10.1578/AM.42.2.2016.193
Yamada, T.K., Kemper, C.M., Wada, S., Oishi, M.: Omura’s whale (Balaenoptera omurai). In: Dyck, S.V., Strahan, R. (eds.) The Mammals of Australia, pp. 801–802. New Holland Publishers, Sydney (2008)
Cerchio, S., Andrianantenaina, B., Lindsay, A., Rekdahl, M., Andrianarivelo, N., Rasoloarijao, T.: Omura’s whales (Balaenoptera omurai) off northwest Madagascar: ecology, behaviour and conservation needs. R. Soc. Open Sci. (2015). doi:10.1098/rsos.150301
Mizroch, S.A., Rice, D.W., Zwiefelhofer, D., Waite, J., Perryman, W.L.: Distribution and movements of fin whales in the North Pacific Ocean. Mamm. Rev. 39(3), 193–227 (2009). doi:10.1111/j.1365-2907.2009.00147.x
Department of the Environment: Balaenoptera physalus. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Simon, M., Stafford, K.M., Beedholm, K., Lee, C.M., Madsen, P.T.: Singing behavior of fin whales in the Davis Strait with implications for mating, migration and foraging. J. Acoust. Soc. Am. 128(5), 3200–3210 (2010). doi:10.1121/1.3495946
Edds, P.L.: Characteristics of finback Balaenoptera physalus vocalizations in the St. Lawrence Estuary. Bioacoustics 1, 131–149 (1988)
Sirovic, A., Williams, L., Kerosky, S., Wiggins, S., Hildebrand, J.: Temporal separation of two fin whale call types across the eastern North Pacific. Mar. Biol. (Berlin) 160(1), 47–57 (2013)
Charif, R.A., Mellinger, D.K., Dunsmore, K.J., Fristrup, K.M., Clark, C.W.: Estimated source levels of fin whale (Balaenoptera physalus) vocalizations: adjustments for surface interference. Mar. Mamm. Sci. 18(1), 81–98 (2002). doi:10.1111/j.1748-7692.2002.tb01020.x
Watkins, W.A., Tyack, P., Moore, K.E., Bird, J.E.: The 20-Hz signals of finback whales (Balaenoptera physalus). J. Acoust. Soc. Am. 82(6), 1901–1912 (1987). doi:10.1121/1.395685
Weirathmueller, M.J., Wilcock, W.S.D., Soule, D.C.: Source levels of fin whale 20 Hz pulses measured in the Northeast Pacific Ocean. J. Acoust. Soc. Am. 133(2), 741–749 (2013). doi:10.1121/1.4773277
Goldbogen, J.A., Stimpert, A.K., Deruiter, S.L., Calambokidis, J., Friedlaender, A.S., Schorr, G.S., Moretti, D.J., Tyack, P.L., Southall, B.L.: Using accelerometers to determine the calling behavior of tagged baleen whales. J. Exp. Biol. 217(14), 2449–2455 (2014). doi:10.1242/jeb.103259
Thompson, P.O., Findley, L.T., Vidal, O.: 20-Hz pulses and other vocalizations of fin whales, Balaenoptera physalus, in the Gulf of California, Mexico. J. Acoust. Soc. Am. 92(6), 3051–3057 (1992). doi:10.1121/1.404201
Schevill, W.E., Watkins, W.A., Backus, R.H.: The 20- cycle signals and Balaenoptera (fin whales). Mar. Bio-Acoust. 45, 147–152 (1964)
Croll, D.A., Clark, C.W., Acevedo, A., Tershy, B., Flores, S., Gedamke, J., Urban, J.: Only male fin whales sing loud songs. Nature 417(20), 809 (2002). doi:10.1038/417809a
Morano, J., Salisbury, D., Rice, A., Conklin, K., Falk, K., Clark, C.: Seasonal and geographical patterns of fin whale song in the western North Atlantic Ocean. J. Acoust. Soc. Am. 132(2), 1207–1212 (2012). doi:10.1121/1.4730890
Chittleborough, R.: Dynamics of two populations of the humpback whale, Megaptera novaeangliae (Borowski). Mar. Freshw. Res. 16(1), 33–128 (1965). doi:10.1071/MF9650033
Van Opzeeland, I., Van Parijs, S., Kindermann, L., Burkhardt, E., Boebel, O.: Calling in the cold: pervasive acoustic presence of humpback whales (Megaptera novaeangliae) in Antarctic coastal waters. PLoS ONE 8(9), e73007 (2013). doi:10.1371/journal.pone.0073007
Cerchio, S., Dahlheim, M.: Variation in feeding vocalizations of humpback whales (Megaptera novaeangliae) from Southeast Alaska. Bioacoustics 11(4), 277–295 (2001). doi:10.1080/09524622.2001.9753468
Thompson, P.O., Cummings, W.C., Ha, S.J.: Sounds, source levels, and associated behavior of humpback whales, southeast Alaska. J. Acoust. Soc. Am. 80(3), 735–740 (1986). doi:10.1121/1.393947
Fournet, M.E., Szabo, A., Mellinger, D.K.: Repertoire and classification of non-song calls in Southeast Alaskan humpback whales (Megaptera novaeangliae). J. Acoust. Soc. Am. 137(1), 1–10 (2015). doi:10.1121/1.4904504
Stimpert, A.K., Au, W.W.L., Parks, S.E., Hurst, T., Wiley, D.N.: Common humpback whale (Megaptera novaeangliae) sound types for passive acoustic monitoring. J. Acoust. Soc. Am. 129(1), 476–482 (2011). doi:10.1121/1.3504708
Au, W.W.L., James, D., Andrews, K.: High-frequency harmonics and source level of humpback whale songs. J. Acoust. Soc. Am. 110(5), 2770 (2001). doi:10.1121/1.4777702
Au, W.W.L., Pack, A.A., Lammers, M.O., Herman, L.M., Deakos, M.H., Andrews, K.: Acoustic properties of humpback whale songs. J. Acoust. Soc. Am. 120(2), 1103–1110 (2006). doi:10.1121/1.2211547
Payne, K., Payne, R.: Large scale changes over 19 years in songs of humpback whales in Bermuda. Z. Tierpsychol. 6, 89–114 (1985)
Payne, R.S., McVay, S.: Songs of humpback whales. Science 173(3997), 585–597 (1971). doi:10.1126/science.173.3997.585
Zoidis, A.M., Smultea, M.A., Frankel, A.S., Hopkins, J.L., Day, A., McFarland, A.S., Whitt, A.D., Fertl, D.: Vocalizations produced by humpback whale (Megaptera novaeangliae) calves recorded in Hawaii. J. Acoust. Soc. Am. 123(3), 1737–1746 (2008). doi:10.1121/1.2836750
Dunlop, R.A., Cato, D.H., Noad, M.J., Stokes, D.M.: Source levels of social sounds in migrating humpback whales (Megaptera novaeangliae). J. Acoust. Soc. Am. 134(1), 706–714 (2013). doi:10.1121/1.4807828
Dunlop, R.A., Noad, M.J., Cato, D.H., Stokes, D.: The social vocalization repertoire of east Australian migrating humpback whales (Megaptera novaeangliae). J. Acoust. Soc. Am. 122(5), 2893–2905 (2007). doi:10.1121/1.2783115
Garland, E., Noad, M., Goldizen, A., Lilley, M., Rekdahl, M., Garrigue, C., Constantine, R., Hauser, N.D., Poole, M., Robbins, J.: Quantifying humpback whale song sequences to understand the dynamics of song exchange at the ocean basin scale. J. Acoust. Soc. Am. 133(1), 560–569 (2013). doi:10.1121/1.4770232
Rekdahl, M.L., Dunlop, R.A., Goldizen, A.W., Garland, E.C., Biassoni, N., Miller, P., Noad, M.J.: Non-song social call bouts of migrating humpback whales. J. Acoust. Soc. Am. 137(6), 3042–3053 (2015). doi:10.1121/1.4921280
Rekdahl, M.L., Dunlop, R.A., Noad, M.J., Goldizen, A.W.: Temporal stability and change in the social call repertoire of migrating humpback whales. J. Acoust. Soc. Am. 133(3), 1785–1795 (2013). doi:10.1121/1.4789941
Cato, D.H.: Songs of humpback whales: the Australian perspective. Mem. Qld. Mus. 30(2), 277–290 (1991)
Helweg, D.A., Cato, D.H., Jenkins, P.F., Claire, G., McCauley, R.D.: Geographic variation in South Pacific Humpback Whale songs. Behaviour 135(1), 1–27 (1998)
Garland, E.C., Gedamke, J., Rekdahl, M.L., Noad, M.J., Garrigue, C., Gales, N.: Humpback whale song on the Southern Ocean feeding grounds: implications for cultural transmission. PLoS ONE 8(11), e79422 (2013). doi:10.1371/journal.pone.0079422
Darling, J.D., Jones, M.E., Nicklin, C.P.: Humpback whale songs: do they organize males during the breeding season? Behaviour 143(9), 1051–1101 (2006)
Winn, H.E., Winn, L.K.: The song of the humpback whale Megaptera novaeangliae in the West Indies. Mar. Biol. 47, 97–114 (1978)
Noad, M.J., Cato, D.H., Bryden, M.M., Jenner, M.N., Jenner, K.C.S.: Cultural revolution in whale songs. Nature 408, 537 (2000)
Vu, E., Risch, D., Clark, C., Gaylord, S., Hatch, L., Thompson, M., Wiley, D., Van Parijs, S.: Humpback whale song occurs extensively on feeding grounds in the western North Atlantic Ocean. Aquat. Biol. 14(2), 175–183 (2011)
Magnusdottir, E.E., Rasmussen, M.H., Lammers, M.O., Svavarsson, J.: Humpback whale songs during winter in subarctic waters. Polar Biol. 37(3), 427–433 (2014)
Stimpert, A.K., Wiley, D.N., Au, W.W.L., Johnson, M.P., Arsenault, R.: ’Megapclicks’: acoustic click trains and buzzes produced during night-time foraging of humpback whales (Megaptera novaeangliae). Biol. Lett. 3(5), 467–470 (2007). doi:10.1098/rsbl.2007.0281
Miller, P.J.O., Johnson, M.P., Tyack, P.L.: Sperm whale behaviour indicates the use of echolocation click buzzes “creaks” in prey capture. Proc. R. Soc. Lond. B 271, 2239–2247 (2004)
Parks, S.E., Cusano, D.A., Stimpert, A.K., Weinrich, M.T., Friedlaender, A.S., Wiley, D.N.: Evidence for acoustic communication among bottom foraging humpback whales. Sci. Rep. 4, 7508 (2014). doi:10.1038/srep07508
Jefferson, T.A., Waerebeek, K.V.: The taxonomic status of the nominal dolphin species Delphinus tropicalis van Bree, 1971. Mar. Mamm. Sci. 18(4), 787–818 (2002). doi:10.1111/j.1748-7692.2002.tb01074.x
Ross, G.J.B.: Review of the Conservation Status of Australia’s Smaller Whales and Dolphins. Department of the Environment and Heritage, Canberra (2006)
Neumann, D.R.: Seasonal movements of short-beaked common dolphins (Delphinus delphis) in the north-western Bay of Plenty, New Zealand: influence of sea surface temperature and El Niño/La Niña. New Zeal. J. Mar. Freshw. 35(2), 371–374 (2001). doi:10.1080/00288330.2001.9517007
Petrella, V.: Whistle Characteristics of Common Dolphins (Delphinus sp.) in the Hauraki Gulf, New Zealand. Ph.D. Thesis, Department of Applied Biology, University of Napes, Italy (2009)
Petrella, V., Martinez, E., Anderson, M., Stockin, K.: Whistle characteristics of common dolphins (Delphinus sp.) in the Hauraki Gulf, New Zealand. Mar. Mamm. Sci. 28(3), 479–496 (2011). doi:10.1111/j.1748-7692.2011.00499.x
Oswald, J.N., Barlow, J., Norris, T.F.: Acoustic identification of nine delphinid species in the eastern tropical Pacific Ocean. Mar. Mamm. Sci. 19(1), 20–37 (2003). doi:10.1111/j.1748-7692.2003.tb01090.x
Oswald, J.N., Rankin, S., Barlow, J., Lammers, M.O.: A tool for real-time acoustic species identification of delphinid whistles. J. Acoust. Soc. Am. 122(1), 587–595 (2007). doi:10.1121/1.2743157
Ansmann, I.C.: The whistle repertoire and acoustic behavior of short-beaked common dolphins, Delphinus delphis, around the British Isles, with applications for acoustic surveying. M.Sc. Thesis, School of Biological Sciences, Bangor University, Wales, UK (2005)
Ansmann, I.C., Goold, J.C., Evans, P.G.H., Simmonds, M., Keith, S.G.: Variation in the whistle characteristics of short-beaked common dolphins, Delphinus delphis, at two locations around the British Isles. J. Mar. Biol. Assoc. UK 87(1), 19–26 (2007). doi:10.1017/S0025315407054963
Scullion, A.: Short-Beaked Common Dolphin, Delphinus delphis Whistles: Whistle Density, a Reliable form of Measuring Group Size? M.Sc. Thesis, Bangor University (2004)
Gannier, A., Fuchs, S., Quebre, P., Oswald, J.N.: Performance of a contour-based classification method for whistles of Mediterranean delphinids. Appl. Acoust. 71(11), 1063–1069 (2010). doi:10.1016/j.apacoust.2010.05.019
Busnel, R.-G., Dziedzic, A.: Acoustic signals of the pilot whale Globicephala melaena and of the porpoises Delphinus delphis and Phocoena phocoena. In: Norris, K.S. (ed.) Whales, Dolphins, and Porpoises, pp. 607–646. University of California Press, Berkeley (1966)
Papale, E., Azzolin, M., Cascao, I., Gannier, A., Lammers, M.O., Martin, V.M., Oswald, J., Perez-Gil, M., Prieto, R., Silva, M.A., Giacoma, C.: Macro- and micro-geographic variation of short-beaked common dolphin’s whistles in the Mediterranean Sea and Atlantic Ocean. Ethol. Ecol. Evol. 26, 392–404 (2014). doi:10.1080/03949370.2013.851122
Moore, S.E., Ridgway, S.H.: Whistles produced by common dolphins from the Southern California Bight. Aquat. Mamm. 21(1), 55–63 (1995)
Bryden, M.M.: Observations on a Pygmy Killer Whale, Feresa attenuata, stranded on the East Coast of Australia. Wildl. Res. 3(1), 21–28 (1976). doi:10.1071/WR9760021
Madsen, P.T., Kerr, I., Payne, R.: Source parameter estimates of echolocation clicks from wild pygmy killer whales (Feresa attenuata) (L). J. Acoust. Soc. Am. 116(4), 1909–1912 (2004). doi:10.1121/1.1788726
Rendell, L.E., Gordon, J.C.D.: Vocal response of long-finned pilot whales (Globicephala melas) to military sonar in the Ligurian Sea. Mar. Mamm. Sci. 15(1), 198–204 (1999)
Baron, S.C., Martinez, A., Garrison, L.P., Keith, E.O.: Differences in acoustic signals from Delphinids in the western North Atlantic and northern Gulf of Mexico. Mar. Mamm. Sci. 24(1), 42–56 (2008). doi:10.1111/j.1748-7692.2007.00168.x
Jensen, F., Perez, J., Hojnson, M., Soto, N., Madsen, P.: Calling under pressure: short-finned pilot whales make social calls during deep foraging dives. Proc. R. Soc. Lond. Ser. B Biol. Sci. 278(1721), 3017–3025 (2011)
Scheer, M.: Call vocalizations recorded among short-finned pilot whales (Globicephala macrorhynchus) off Tenerife, Canary Islands. Aquat. Mamm. 39(3), 306–313 (2013)
Pérez, J.M., Jensen, F.H., Rojano-Doñate, L., Aguilar de Soto, N.: Different modes of acoustic communication in deep-diving short-finned pilot whales (Globicephala macrorhynchus). Mar. Mamm. Sci. 33(1), 59–79 (2017). doi:10.1111/mms.12344
Caldwell, M.C., Caldwell, D.K.: Simultaneous but different narrow-band sound emissions by a captive eastern Pacific pilot whale, Globicephala scammoni. Mammalia 33(3), 505–508 (1969)
Aguilar de Soto, N., Johnson, M.P., Madsen, P.T., Diaz, F., Dominguez, I., Brito, A.E., Tyack, P.: Cheetahs of the deep sea: deep foraging sprints in short-finned pilot whales off Tenerife (Canary Islands). J. Anim. Ecol. 77, 936–947 (2008). doi:10.1111/j.1365-2656.2008.01393.x
Baumann-Pickering, S., Simonis, A.E., Oleson, E.M., Baird, R.W., Roch, M.A., Wiggins, S.M.: False killer whale and short-finned pilot whale acoustic identification. Endanger. Species Res. 28(2), 97–108 (2015). doi:10.3354/esr00685
DeRuiter, S.L., Boyd, I.L., Claridge, D.E., Clark, C.W., Gagnon, C., Southall, B.L., Tyack, P.L.: Delphinid whistle production and call matching during playback of simulated military sonar. Mar. Mamm. Sci. 29(2), E46–E59 (2012). doi:10.1111/j.1748-7692.2012.00587.x
Cato, D.H., Savage, M., Dunlop, R.A., Parnum, I., Blewitt, M., Gibbs, S., Donnelly, D., Cleary, J., McCauley, R.D.: Acoustic surveying for beaked whales in the Coral Sea as a mitigation measure for naval exercises. In: OCEANS 2010 IEEE-Sydney 2010, pp. 1–4. IEEE
Taruski, A.G.: The whistle repertoire of the North Atlantic pilot whale (Globicephala melaena) and its relationship to behavior and environment. In: Winn, H.E., Olla, B.L. (eds.) Behavior of Marine Animals, vol. 3: cetaceans, pp. 345–368. Plenum Press, New York (1979)
Weilgart, L.S., Whitehead, H.: Vocalizations of the North Atlantic pilot whale (Globicephala melas) as related to behavioral contexts. Behav. Ecol. Sociobiol. 26, 399–402 (1990)
Steiner, W.W.: Species-specific differences in pure tonal whistle vocalizations of five western North Atlantic dolphin species. Behav. Ecol. Sociobiol. 9, 241–246 (1981)
Nemiroff, L., Whitehead, H.: Structural characteristics of pulsed calls of long-finned pilot whales Globicephala melas. Bioacoustics 19, 67–92 (2009)
Zwamborn, E.M.J., Whitehead, H.: Repeated call sequences and behavioural context in long-finned pilot whales off Cape Breton, Nova Scotia, Canada. Bioacoustics 26(2), 169–183 (2016). doi:10.1080/09524622.2016.1233457
Alves, A., Antunes, R., Tyack, P.L., Miller, P.J.O., Lam, F.-P.A., Kvadsheim, P.H.: Vocal matching of naval sonar signals by long-finned pilot whales (Globicephala melas). Mar. Mamm. Sci. 30(3), 1248–1257 (2014). doi:10.1111/mms.12099
Corkeron, P.J., Bryden, M.M.: Sightings of Risso’s Dolphin, Grampus griseus (Cetacea: Delphinidae), off Fraser Island, Queensland. Aust. Mammal. 15, 129–130 (1991)
Corkeron, P.J., Van Parijs, S.M.: Vocalizations of eastern Australian Risso’s dolphins, Grampus griseus. Can. J. Zool. 79, 160–164 (2001). doi:10.1139/z00-180
Rendell, L.E., Matthews, J.N., Gill, A., Gordon, J.C.D., Macdonald, D.W.: Quantitative analysis of tonal calls from five odontocete species, examining interspecific and intraspecific variation. J. Zool. 249, 403–410 (1999)
Neves, S.: Acoustic Behaviour of Risso’s dolphins, Grampus griseus, in the Canary Islands, Spain. Ph.D. University of St Andrews (2013)
Madsen, P.T., Kerr, I., Payne, R.: Echolocation clicks of two free-ranging, oceanic delphinids with different food preferences: false killer whales Pseudorca crassidens and Risso’s dolphins Grampus griseus. J. Exp. Biol. 207(11), 1811–1823 (2004)
Philips, J.D., Nachtigall, P.E., Au, W.W.L., Pawloski, J.L., Roitblat, H.L.: Echolocation in the Risso’s dolphin, Grampus griseus. J. Acoust. Soc. Am. 113(1), 605–616 (2003). doi:10.1121/1.1527964
Soldevilla, M.S., Henderson, E.E., Campbell, G.S., Wiggins, S.M., Hildebrand, J.A., Roch, M.A.: Classification of Risso’s and Pacific white-sided dolphins using spectral properties of echolocation clicks. J. Acoust. Soc. Am. 124(1), 609–624 (2008). doi:10.1121/1.2932059
Smith, A.B., Kloepper, L.N., Yang, W.-C., Huang, W.-H., Jen, I.-F., Rideout, B.P., Nachtigall, P.E.: Transmission beam characteristics of a Risso’s dolphin (Grampus griseus). J. Acoust. Soc. Am. 139(1), 53–62 (2016). doi:10.1121/1.4937752
Oswald, J.N., Rankin, S., Barlow, J.: First description of whistles of Pacific Fraser’s dolphins Lagenodelphis hosei. Bioacoustics 16(2), 99–111 (2007)
Watkins, W., Daher, M., Fristrup, K., Notarbartolo Di Sciara, G.: Fishing and acoustic behavior of Fraser’s dolphin (Lagenodelphis hosei) near Dominica, Southeast Caribbean. Caribb. J. Sci. 30(1–2), 76–82 (1994)
Kyhn, L.A., Tougaard, J., Jensen, F., Wahlberg, M., Stone, G., Yoshinaga, A., Beedholm, K., Madsen, P.T.: Feeding at a high pitch: source parameters of narrow band, high-frequency clicks from echolocating off-shore hourglass dolphins and coastal Hector’s dolphins. J. Acoust. Soc. Am. 125(3), 1783–1791 (2009). doi:10.1121/1.3075600
Tougaard, J., Kyhn, L.A.: Echolocation sounds of hourglass dolphins (Lagenorhynchus cruciger) are similar to the narrow band high-frequency echolocation sounds of the dolphin genus Cephalorhynchus. Mar. Mamm. Sci. 26(1), 239–245 (2010). doi:10.1111/j.1748-7692.2009.00307.x
Gill, P.C., Ross, G.J.B., Dawbin, W.H., Wapstra, H.: Confirmed sightings of dusky dolphins (Lagenorhynchus obscurus) in southern Australian waters. Mar. Mamm. Sci. 16(2), 452–459 (2000). doi:10.1111/j.1748-7692.2000.tb00937.x
Au, W.W.L., Würsig, B.: Echolocation signals of dusky dolphins (Lagenorhynchus obscurus) in Kaikoura, New Zealand. J. Acoust. Soc. Am. 115(5), 2307–2313 (2004). doi:10.1121/1.1690082
Vaughn-Hirshorn, R., Hodge, K., Würsig, B., Sappenfield, R., Lammers, M., Dudzinski, K.: Characterizing dusky dolphin sounds from Argentina and New Zealand. J. Acoust. Soc. Am. 132(1), 498–506 (2012). doi:10.1121/1.4728191
Beasley, I., Robertson, K.M., Arnold, P.: Description of a new dolphin, the Australian snubfin dolphin Orcaella Heinsohni Sp. N. (Cetacea, Delphinidae). Mar. Mamm. Sci. 21(3), 365–400 (2005). doi:10.1111/j.1748-7692.2005.tb01239.x
Van Parijs, S.M., Parra, G.J., Corkeron, P.J.: Sounds produced by Australian Irrawaddy dolphins, Orcaella brevirostris. J. Acoust. Soc. Am. 108(4), 1938–1940 (2000). doi:10.1121/1.1289667
Berg Soto, A., Marsh, H., Everingham, Y., Smith, J.N., Parra, G.J., Noad, M.: Discriminating between the vocalizations of Indo-Pacific humpback and Australian snubfin dolphins in Queensland, Australia. J. Acoust. Soc. Am. 136(2), 930–938 (2014). doi:10.1121/1.4884772
Brown, A.M., Smith, J., Kent, C.S., Marley, S., Allen, S.J., Thiele, D., Bejder, L., Erbe, C., Chabanne, D.: Relative Abundance, Population Genetic Structure and Passive Acoustic Monitoring of Australian Snubfin and Humpback Dolphins in Regions Within the Kimberley. Western Australian Marine Science Institution, Perth (2017)
Marley, S.A., Salgado Kent, C.P., Erbe, C., Thiele, D.: A tale of two soundscapes: comparing the acoustic characteristics of urban versus pristine coastal dolphin habitats in Western Australia. Acoust. Aust. (2017). doi:10.1007/s40857-017-0106-7
Culik, B.: Review of Small Cetaceans: Distribution, Behaviour, Migration and Threats. Convention on Migratory Species (CMS), Bonn (2002)
Wellard, R., Lightbody, K., Fouda, L., Blewitt, M., Riggs, D., Erbe, C.: Killer whale (Orcinus orca) predation on beaked whales (Mesoplodon spp.) in the Bremer Sub-Basin, Western Australia. PLoS ONE 11(12), e0166670 (2016). doi:10.1371/journal.pone.0166670
Department of the Environment: Orcinus orca. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Au, W.W.L., Ford, J.K.B., Horne, J.K., Allman, K.A.N.: Echolocation signals of free-ranging killer whales (Orcinus orca) and modelling of foraging for chinook salmon (Oncorhynchus tshawytscha). J. Acoust. Soc. Am. 115(2), 901–909 (2004). doi:10.1121/1.1642628
Barrett-Lennard, L.G., Ford, J.K.B., Heise, K.A.: The mixed blessing of echolocation: differences in sonar use by fish-eating and mammal-eating killer whales. Anim. Behav. 51(3), 553–565 (1996). doi:10.1006/anbe.1996.0059
Ford, J.K.B.: A catalogue of underwater calls produced by killer whales (Orcinus orca) in British Columbia. Can. Data Rep. Fish. Aquat. Sci. 633, 1–165 (1987)
Ford, J.K.B.: Acoustic behaviour of resident killer whales (Orcinus orca) off Vancouver Island, British-Columbia. Can. J. Zool. 67(3), 727–745 (1989)
Ford, J.K.B.: Vocal traditions among resident killer whales (Orcinus orca) in coastal waters of British Columbia. Can. J. Zool. 69(6), 1454–1483 (1991)
Holt, M., Noren, D., Emmons, C.: Effects of noise levels and call types on the source levels of killer whale calls. J. Acoust. Soc. Am. 130(5), 3100–3106 (2011). doi:10.1121/1.3641446
Miller, P.J.O.: Diversity in sound pressure levels and estimated active space of resident killer whale vocalizations. J. Comp. Psychol. A 192, 449–459 (2006). doi:10.1007/s00359-005-0085-2
Riesch, R., Ford, J.K.B., Thomsen, F.: Stability and group specificity of stereotyped whistles in resident killer whales, Orcinus orca, off British Columbia. Anim. Behav. 71, 79–91 (2006). doi:10.1016/j.anbehav.2005.03.026
Riesch, R., Ford, J.K.B., Thomsen, F.: Whistle sequences in wild killer whales (Orcinus orca). J. Acoust. Soc. Am. 124(3), 1822–1829 (2008). doi:10.1121/1.2956467
Thomsen, F., Franck, D., Ford, J.K.B.: Characteristics of whistles from the acoustic repertoire of resident killer whales (Orcinus orca) off Vancouver Island, British Columbia. J. Acoust. Soc. Am. 109(3), 1240–1246 (2001). doi:10.1121/1.1349537
Gassmann, M., Henderson, E.E., Wiggins, S.M., Roch, M.A., Hildebrand, J.A.: Offshore killer whale tracking using multiple hydrophone arrays. J. Acoust. Soc. Am. 134(5), 3513–3521 (2013). doi:10.1121/1.4824162
Filatova, O., Ford, J., Matkin, C., Barrett-Lennard, L., Burdin, A., Hoyt, E.: Ultrasonic whistles of killer whales (Orcinus orca) recorded in the North Pacific (L). J. Acoust. Soc. Am. 132(6), 3618–3621 (2012). doi:10.1121/1.4764874
Filatova, O.A., Fedutin, I.D., Burdin, A.M., Hoyt, E.: The structure of the discrete call repertoire of killer whales Orcinus orca from southeast Kamchatka. Bioacoustics 16(3), 261–280 (2007)
Filatova, O.A., Fedutin, I.D., Nagaylik, M.M., Burdin, A.M., Hoyt, E.: Usage of monophonic and biphonic calls by free-ranging resident killer whales (Orcinus orca) in Kamchatka, Russian Far East. Acta Ethol. 12(1), 37–44 (2009). doi:10.1007/s10211-009-0056-7
Simonis, A., Baumann-Pickering, S., Oleson, E., Melcon, M., Gassmann, M., Wiggins, S., Hildebrand, J.: High-frequency modulated signals of killer whales (Orcinus orca) in the North Pacific. J. Acoust. Soc. Am. 131(4), EL295–EL301 (2012). doi:10.1121/1.3690963
Filatova, O., Deecke, V., Ford, J., Matkin, C., Barrett-Lennard, L., Guzeev, M., Burdin, A., Hoyt, E.: Call diversity in the North Pacific killer whale populations: implications for dialect evolution and population history. Anim. Behav. 83(3), 595–603 (2012)
Samarra, F.I.P., Deecke, V.B., Vinding, K., Rasmussen, M.H., Swift, R.J., Miller, P.J.O.: Killer whales (Orcinus orca) produce ultrasonic whistles. J. Acoust. Soc. Am. 128, EL205–EL210 (2010). doi:10.1121/1.3462235
Simon, M., Wahlberg, M., Miller, L.A.: Echolocation clicks from killer whales (Orcinus orca) feeding on herring (Clupea harengus) (L). J. Acoust. Soc. Am. 121(2), 749–752 (2007). doi:10.1121/1.2404922
Richlen, M.F., Thomas, J.A.: Acoustic behavior of Antarctic killer whales (Orcinus orca) recorded near the ice edge of McMurdo Sound, Antarctica. Aquat. Mamm. 34(4), 448–457 (2008)
Schall, E., Van Opzeeland, I.: Calls produced by Ecotype C killer whales (Orcinus orca) off the Eckstroem Iceshelf, Antarctica. Aquat. Mamm. 43(2), 117–126 (2017). doi:10.1578/AM.43.2.2017.117
Filatova, O.A., Miller, P.J.O., Yurk, H., Samarra, F.I.P., Hoyt, E., Ford, J.K.B., Matkin, C.O., Barrett-Lennard, L.G.: Killer whale call frequency is similar across the oceans, but varies across sympatric ecotypes. J. Acoust. Soc. Am. 138, 251–257 (2015). doi:10.1121/1.4922704
Samarra, F.I.P., Deecke, V.B., Miller, P.J.O.: Low-frequency signals produced by Northeast Atlantic killer whales (Orcinus orca). J. Acoust. Soc. Am. 139(3), 1149–1157 (2016). doi:10.1121/1.4943555
Miller, P.J.O.: Mixed-directionality of killer whale stereotyped calls: a direction of movement cue? Behav. Ecol. Sociobiol. 52(3), 262–270 (2002). doi:10.1007/s00265-002-0508-9
Ford, J., Fisher, H.D.: Killer whale (Orcinus orca) dialects as an indicator of stocks in British Columbia. Rep. Int. Whal. Comm. 32, 671–679 (1982)
Strager, H.: Pod-specific call repertoires and compound calls of killer whales, Orcinus orca Linnaeus, 1758, in the waters of northern Norway. Can. J. Zool. 73(6), 1037–1047 (1995)
Wellard, R., Erbe, C., Fouda, L., Blewitt, M.: Data from: vocalisations of killer whales (Orcinus orca) in the Bremer Canyon. In: Dryad Data Repository, Western Australia (2015)
Bryden, M.M., Dawbin, W.H., Heinsohn, G.E., Brown, D.H.: Melon-Headed Whale, Peponocephala electra, on the East Coast of Australia. J. Mammal. 58(2), 180–187 (1977). doi:10.2307/1379575
Frankel, A.S., Yin, S.: A description of sounds recorded from melon-headed whales (Peponocephala electra) off Hawai’i. J. Acoust. Soc. Am. 127(5), 3248–3255 (2010). doi:10.1121/1.3365259
Watkins, W.A., Daher, M.A., Samuels, A., Gannon, D.P.: Observations of Peponocephala electra, the melon-headed whale, in the southeastern Caribbean. Caribb. J. Sci. 33, 34–40 (1997)
Baumann-Pickering, S., Wiggins, S.M., Hildebrand, J.A., Roch, M.A., Schnitzler, H.U.: Discriminating features of echolocation clicks of melon-headed whales (Peponocephala electra), bottlenose dolphins (Tursiops truncatus), and Gray’s spinner dolphins (Stenella longirostris longirostris). J. Acoust. Soc. Am. 128(4), 2212–2224 (2010). doi:10.1121/1.3479549
Baumann-Pickering, S., Roch, M.A., Wiggins, S.M., Schnitzler, H.-U., Hildebrand, J.A.: Acoustic behavior of melon-headed whales varies on a diel cycle. Behav. Ecol. Sociobiol. 69(9), 1553–1563 (2015). doi:10.1007/s00265-015-1967-0
Department of the Environment: Pseudorca crassidens. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Au, W.W.L., Pawloski, J.L., Nachtigall, P.E., Blonz, M., Gisiner, R.C.: Echolocation signals and transmission beam pattern of a false killer whale (Pseudorca crassidens). J. Acoust. Soc. Am. 98(1), 51–59 (1995). doi:10.1121/1.413643
Brill, R.L., Pawloski, J.L., Helweg, D.A., Au, W.W., Moore, P.W.B.: Target detection, shape discrimination, and signal characteristics of an echolocating false killer whale (Pseudorca crassidens). J. Acoust. Soc. Am. 92(3), 1324–1330 (1992). doi:10.1121/1.403926
Kamminga, C., Van Velden, J.G.: Investigations on Cetacean Sonar VIII, sonar signals of Pseudorca crassidens in comparison with Tursiops truncatus. Aquat. Mamm. 13(2), 43–49 (1987)
Kloepper, L., Nachtigall, P., Quintos, C., Vlachos, S.: Single-lobed frequency-dependent beam shape in an echolocating false killer whale (Pseudorca crassidens). J. Acoust. Soc. Am. 131(1), 577–581 (2012). doi:10.1121/1.3664076
Thomas, J.A., Turl, C.W.: Echolocation characteristics and range detection threshold of a false killer whale (Pseudorca crassidens). In: Thomas, J.A., Kastelein, R.A. (eds.) Sensory Abilities of Cetaceans, pp. 321–333. De Spil Publishers, New York (1990)
Parra, G.J., Corkeron, P.J., Marsh, H.: The Indo-Pacific humpback dolphin, Sousa chinensis (Osbeck, 1765), in Australian waters: a summary of current knowledge. Aquat. Mamm. 30(1), 197–206 (2004). doi:10.1578/AM.30.1.2004.197
Jefferson, T.A., Rosenbaum, H.C.: Taxonomic revision of the humpback dolphins (Sousa spp.), and description of a new species from Australia. Mar. Mamm. Sci. 30(4), 1494–1541 (2014). doi:10.1111/mms.12152
Van Parijs, S.M., Corkeron, P.J.: Vocalizations and behaviour of Pacific humpback dolphins Sousa chinensis. Ethology 107(8), 701–716 (2001)
de Freitas, M., Jensen, F.H., Tyne, J., Bejder, L., Madsen, P.T.: Echolocation parameters of Australian humpback dolphins (Sousa sahulensis) and Indo-Pacific bottlenose dolphins (Tursiops aduncus) in the wild. J. Acoust. Soc. Am. 137(6), 3033–3043 (2015). doi:10.1121/1.4921277
Van Parijs, S.M., Corkeron, P.J.: Evidence for signature whistle production by a Pacific humpback dolphin, Sousa chinensis. Mar. Mamm. Sci. 17(4), 944–949 (2001). doi:10.1111/j.1748-7692.2001.tb01308.x
Xu, X., Zhang, L., Wei, C.: Whistles of Indo-Pacific humpback dolphins (Sousa chinensis). AIP Conf. Proc. 1495(1), 556–562 (2012). doi:10.1063/1.4765955
Schultz, K.W., Corkeron, P.J.: Interspecific differences in whistles produced by inshore dolphins in Moreton Bay, Queensland, Australia. Can. J. Zool. 72(6), 1061–1068 (1994)
Wang, Z., Fang, L., Shi, W., Wang, K., Wang, D.: Whistle characteristics of free-ranging Indo-Pacific humpback dolphins (Sousa chinensis) in Sanniang Bay, China. J. Acoust. Soc. Am. 133(4), 2479–2489 (2013). doi:10.1121/1.4794390
Seekings, P.J., Yeo, K.P., Chen, Z.P., Nanayakkara, S.C., Tan, J., Tay, P., Taylor, E.: Classification of a large collection of whistles from Indo-Pacific Humpback Dolphins (Sousa chinensis). In: IEEE Oceans 2010 Conference, Sydney, Australia, 24–27 May 2010
Sims, P., Vaughn, R., Hung, S., Würsig, B.: Sounds of Indo-Pacific humpback dolphins (Sousa chinensis) in West Hong Kong: a preliminary description. J. Acoust. Soc. Am. 131(1), EL48–EL53 (2012). doi:10.1121/1.3663281
Hoffman, J.M.: Regional Differences in the Whistles of Australasian Humpback Dolphins (Genus Sousa). M.Sc. Thesis, Trent University (2015)
Goold, J.C., Jefferson, T.A.: A note on clicks recorded from free-ranging Indo-Pacific humpback dolphins, Sousa chinensis. Aquat. Mamm. 30(1), 175–178 (2004)
Fang, L., Li, S., Wang, K., Wang, Z., Shi, W., Wang, D.: Echolocation signals of free-ranging Indo-Pacific humpback dolphins (Sousa chinensis) in Sanniang Bay, China. J. Acoust. Soc. Am. 138, 1346–1352 (2015). doi:10.1121/1.4929492
Kimura, S., Akamatsu, T., Fang, L., Wang, Z., Wang, K., Wang, D., Yoda, K.: Apparent source level of free-ranging humpback dolphin, Sousa chinensis, in the South China Sea. J. Mar. Biol. Assoc. UK 96(4), 845–851 (2016). doi:10.1017/S0025315414000071
Silva, T.L., Mooney, T.A., Sayigh, L.S., Tyack, P.L., Baird, R.W., Oswald, J.N.: Whistle characteristics and daytime dive behavior in pantropical spotted dolphins (Stenella attenuata) in Hawai‘i measured using digital acoustic recording tags (DTAGs). J. Acoust. Soc. Am. 140(1), 421–429 (2016). doi:10.1121/1.4955081
Frankel, A.S., Zeddies, D., Simard, P., Mann, D.: Whistle source levels of free-ranging bottlenose dolphins and Atlantic spotted dolphins in the Gulf of Mexico. J. Acoust. Soc. Am. 135(3), 1624–1631 (2014). doi:10.1121/1.4863304
Department of the Environment: Stenella coeruleoalba. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Papale, E., Azzolin, M., Cascão, I., Gannier, A., Lammers, M.O., Martin, V.M., Oswald, J., Perez-Gil, M., Prieto, R., Silva, M.A., Giacoma, C.: Geographic variability in the acoustic parameters of striped dolphin’s (Stenella coeruleoalba) whistles. J. Acoust. Soc. Am. 133(2), 1126–1134 (2013). doi:10.1121/1.4774274
Bazua-Duran, C., Au, W.W.L.: The whistles of Hawaiian spinner dolphins. J. Acoust. Soc. Am. 112(6), 3064–3072 (2002). doi:10.1121/1.1508785
Bazua-Duran, C., Au, W.W.L.: Geographic variations in the whistles of spinner dolphins (Stenella longirostris). J. Acoust. Soc. Am. 116(6), 3757–3769 (2004). doi:10.1121/1.4779792
Lammers, M.O., Au, W.W.L.: Directionality in the whistles of Hawaiian spinner dolphins (Stenella longirostris): a signal feature to cue direction of movement? Mar. Mamm. Sci. 19(2), 249–264 (2003)
Lammers, M.O., Au, W.W.L., Herzing, D.L.: The broadband social acoustic signaling behavior of spinner and spotted dolphins. J. Acoust. Soc. Am. 114(3), 1629–1639 (2003). doi:10.1121/1.1596173
Lammers, M.O., Schotten, M., Au, W.W.L.: The spatial context of free-ranging Hawaiian spinner dolphins (Stenella longirostris) producing acoustic signals. J. Acoust. Soc. Am. 119(2), 1244–1250 (2006). doi:10.1121/1.2151804
Benoit-Bird, K.J., Au, W.W.L.: Phonation behavior of cooperatively foraging spinner dolphins. J. Acoust. Soc. Am. 125(1), 539–546 (2009). doi:10.1121/1.2967477
Heenehan, H.L., Tyne, J.A., Bejder, L., Van Parijs, S.M., Johnston, D.W.: Passive acoustic monitoring of coastally associated Hawaiian spinner dolphins, Stenella longirostris, ground-truthed through visual surveys. J. Acoust. Soc. Am. 140(1), 206–215 (2016). doi:10.1121/1.4955094
Rossi-Santos, M.R., Da Silva, J.M., Silva, F.L., Monteiro-Filho, E.L.A.: Descriptive parameters of pulsed calls for the spinner dolphin, Stenella longirostris, in the Fernando de Noronha Archipelago, Brazil. J. Mar. Biol. Assoc. UK 88(6), 1093–1097 (2008). doi:10.1017/S0025315408000404
Camargo, F.S., Rollo, M.M., Giampaoli, V., Bellini, C.: Whistle variability in South Atlantic spinner dolphins from the Fernando de Noronha Archipelago off Brazil. J. Acoust. Soc. Am. 120(6), 4071–4079 (2006). doi:10.1121/1.2359704
Moron, J.R., Amorim, T.O.S., Sucunza, F., Castro, F.R.D., Rossi-Santos, M., Andriolo, A.: Spinner dolphin whistle in the Southwest Atlantic Ocean: is there a geographic variation? J. Acoust. Soc. Am. 138(4), 2495–2498 (2015). doi:10.1121/1.4931900
Bonato, M., Papale, E., Pingitore, G., Ricca, S., Attoumane, A., Ouledi, A., Giacoma, C.: Whistle characteristics of the spinner dolphin population in the Comoros Archipelago. J. Acoust. Soc. Am. 138(5), 3262–3271 (2015). doi:10.1121/1.4935518
Department of the Environment: Steno bredanensis. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Watkins, W.A., Tyack, P.L., Moore, K.E., Notarbartolo di Sciara, G.: Steno bredanensis in the Mediterranean Sea (Note). Mar. Mamm. Sci. 3(1), 78–82 (1987)
de Lima, I.M.S., Andrade, L.G., Carvalho, R.R., Lailson-Brito, J., Azeyedo, A.F.: Characteristics of whistles from rough-toothed dolphins (Steno bredanensis) in Rio de Janeiro coast, southeastern Brazil. J. Acoust. Soc. Am. 131(5), 4173–4181 (2012). doi:10.1121/1.3701878
Lima, I.M.S., Andrade, L.G., Bittencourt, L., Bisi, T.L., Flach Jr., L., Lailson-Brito, J., Azevedo, A.F.: Whistle comparison of four delphinid species in Southeastern Brazil. J. Acoust. Soc. Am. 139(5), 124–127 (2016). doi:10.1121/1.4947310
Rankin, S., Oswald, J.N., Simonis, A.E., Barlow, J.: Vocalizations of the rough-toothed dolphin, Steno bredanensis, in the Pacific Ocean. Mar. Mamm. Sci. 31(4), 1538–1548 (2015). doi:10.1111/mms.12226
Hale, P.T., Barreto, A.S., Ross, G.J.B.: Comparative morphology and distribution of the aduncus and truncatus forms of bottlenose dolphin Tursiops in the Indian and Western Pacific Oceans. Aquat. Mamm. 26(2), 101–110 (2000)
Charlton-Robb, K., Gershwin, L., Thompson, R., Austin, J., Owen, K., McKenchie, S.: A new dolphin species, the Burrunan dolphin Tursiops australis sp. nov., endemic to southern Australian coastal waters. PLoS ONE 6(9), e24047 (2011). doi:10.1371/journal.pone.0024047
Möller, L.M., Bilgmann, K., Charlton-Robb, K., Beheregaray, L.: Multi-gene evidence for a new bottlenose dolphin species in southern Australia. Mol. Phylogenet. Evol. 49(2), 674–681 (2008). doi:10.1016/j.ympev.2008.08.011
Morisaka, T., Shinohara, M., Nakahara, F., Akamatsu, T.: Geographic variations in the whistles among three Indo-Pacific bottlenose dolphin Tursiops aduncus populations in Japan. Fish. Sci. 71, 568–576 (2005)
Ding, W., Würsig, B., Evans, W.E.: Whistles of bottlenose dolphins: comparisons among populations. Aquat. Mamm. 21(1), 65–77 (1995)
Gridley, T., Berggren, P., Cockcroft, V., Janik, V.: Whistle vocalizations of Indo-Pacific bottlenose dolphins (Tursiops aduncus) inhabiting the south-west Indian Ocean. J. Acoust. Soc. Am. 132(6), 4032–4040 (2012). doi:10.1121/1.4763990
Hawkins, E.R.: Geographic variations in the whistles of bottlenose dolphins (Tursiops aduncus) along the east and west coasts of Australia. J. Acoust. Soc. Am. 128(2), 924–935 (2010). doi:10.1121/1.3459837
Ward, R., Parnum, I., Erbe, C., Salgado-Kent, C.P.: Whistle characteristics of Indo-Pacific bottlenose dolphins (Tursiops aduncus) in the Fremantle Inner Harbour, Western Australia. Acoust. Aust. 44(1), 159–169 (2016). doi:10.1007/s40857-015-0041-4
Hawkins, E.R., Gartside, D.F.: Patterns of whistles emitted by wild Indo-Pacific bottlenose dolphins (Tursiops aduncus) during a provisioning program. Aquat. Mamm. 35(2), 171–186 (2009)
Gridley, T., Cockcroft, V.G., Hawkings, E.R., Blewitt, M.L., Morisaka, T., Janik, V.M.: Signature whistles in free-ranging populations of Indo-Pacific bottlenose dolphins. Mar. Mamm. Sci. 30(2), 512–527 (2014). doi:10.1111/mms.12054
Wahlberg, M., Jensen, F., Soto, N., Beedholm, K., Bejder, L., Oliveira, C., Rasmussen, M., Simon, M., Villadsgaard, A., Madsen, P.: Source parameters of echolocation clicks from wild bottlenose dolphins (Tursiops aduncus and Tursiops truncatus). J. Acoust. Soc. Am. 130(4), 2263–2274 (2011). doi:10.1121/1.3624822
Wellard, R., Charlton-Robb, K., Erbe, C., Wong, B.: Whistle characteristics of newly defined species, the Burrunan dolphin (Tursiops australis), in coastal Victorian waters in Australia. J. Acoust. Soc. Am. 140(4), 3416–3416 (2016). doi:10.1121/1.4970980
Schultz, K.W., Cato, D.H., Corkeron, P.J., Bryden, M.M.: Low-frequency narrow-band sounds produced by bottle-nosed dolphins. Mar. Mamm. Sci. 11(4), 503–509 (1995)
Boisseau, O.: Quantifying the acoustic repertoire of a population: the vocalizations of free ranging bottlenose dolphins in Fjordland, New Zealand. J. Acoust. Soc. Am. 117(4), 2318–2329 (2005). doi:10.1121/1.1861692
Campbell, G.S.: Quantitative comparison of bottlenose dolphin (Tursiops spp.) whistles from three geographic regions. M.Sc. Thesis, San Diego State University (2004)
Simard, P., Lace, N., Gowans, S., Quintana-Rizzo, E., Kuczaj, S., Wells, R., Mann, D.: Low frequency narrow-band calls in bottlenose dolphins (Tursiops truncatus): signal properties, function, and conservation implications. J. Acoust. Soc. Am. 130(5), 3068–3076 (2011). doi:10.1121/1.3641442
Herzing, D.L.: Synchronous and rhythmic vocalizations and correlated underwater behavior of free-ranging Atlantic spotted dolphins (Stenella frontalis) and bottlenose dolphins (Tursiops truncatus) in the Bahamas. Anim. Behav. Cogn. 2(1), 14–29 (2015). doi:10.12966/abc.02.02.2015
May-Collado, L.J., Wartzok, D.: A comparison of bottlenose dolphin whistles in the Atlantic Ocean: factors promoting whistle variation. J. Mammal. 89, 1229–1240 (2008)
Azevedo, A.F., Oliveira, A.M., Rosa, L.D., Lailson-Brito, J.: Characteristics of whistles from resident bottlenose dolphins (Tursiops truncatus) in southern Brazil. J. Acoust. Soc. Am. 121, 2978–2983 (2007). doi:10.1121/1.2713726
Tellechea, J.S., Bouvier, D., Cambon-Tait, D., Norbis, W.: Passive acoustic monitoring of bottlenose dolphins (Tursiops truncatus) on the Uruguayan Coast: vocal characteristics and seasonal cycles. Aquat. Mamm. 40(2), 173–184 (2014)
Gridley, T., Nastasi, A., Kriesell, H.J., Elwen, S.H.: The acoustic repertoire of wild common bottlenose dolphins (Tursiops truncatus) in Walvis Bay, Namibia. Bioacoust. J. 24(2), 153–174 (2015). doi:10.1080/09524622.2015.1014851
Papale, E., Azzolin, M., Cascao, I., Gannier, A., Lammers, M.O., Martin, V.M., Oswald, J., Perez-Gil, M., Prieto, R., Silva, M.A., Giacoma, C.: Acoustic divergence between bottlenose dolphin whistles from the Central-Eastern North Atlantic and Mediterranean Sea. Acta Ethol. 17(3), 155–165 (2014). doi:10.1007/s10211-013-0172-2
Hiley, H.M., Perry, S., Hartley, S., King, S.L.: What’s occurring? Ultrasonic signature whistle use in Welsh bottlenose dolphins (Tursiops truncatus). Bioacoustics 26(1), 25–35 (2017). doi:10.1080/09524622.2016.1174885
Janik, V., King, S., Sayigh, L., Wells, R.: Identifying signature whistles from recordings of groups of unrestrained bottlenose dolphins (Tursiops truncatus). Mar. Mamm. Sci. 29(1), 109–122 (2013). doi:10.1111/j.1748-7692.2011.00549.x
Quick, N., Janik, V.: Bottlenose dolphins exchange signature whistles when meeting at sea. Proc. R. Soc. Lond. Ser. B Biol. Sci. 279(1738), 2539–2545 (2012)
Díaz López, B.: Whistle characteristics in free-ranging bottlenose dolphins (Tursiops truncatus) in the Mediterranean Sea: influence of behaviour. Mammal. Biol. 76(2), 180–189 (2011). doi:10.1016/j.mambio.2010.06.006
Rako Gospić, N., Picciulin, M.: Changes in whistle structure of resident bottlenose dolphins in relation to underwater noise and boat traffic. Mar. Pollute Bull. 105(1), 193–198 (2016). doi:10.1016/j.marpolbul.2016.02.030
Papale, E., Buffa, G., Filiciotto, F., Maccarrone, V., Mazzola, S., Ceraulo, M., Giacoma, C., Buscaino, G.: Biphonic calls as signature whistles in a free-ranging bottlenose dolphin. Bioacoustics 24(3), 223–231 (2015). doi:10.1080/09524622.2015.1041158
Dos Santos, M.E., Louro, S., Couchinho, M., Brito, C.: Whistles of bottlenose dolphins (Tursiops truncatus) in the Sado Estuary, Portugal: characteristics, production rates, and long-term contour stability. Aquat. Mamm. 31(4), 453–462 (2005)
Branstetter, B.K., Moore, P.W., Finneran, J.J., Tormey, M.N., Aihara, H.: Directional properties of bottlenose dolphin (Tursiops truncatus) clicks, burst-pulse, and whistle sounds. J. Acoust. Soc. Am. 131(2), 1613–1621 (2012). doi:10.1121/1.3676694
Nakahara, F., Miyazaki, N.: Vocal exchanges of signature whistles in bottlenose dolphins (Tursiops truncatus). J. Ethol. 29(2), 309–320 (2011). doi:10.1007/s10164-010-0259-4
van der Woude, S.E.: Bottlenose dolphins (Tursiops truncatus) moan as low in frequency as baleen whales. J. Acoust. Soc. Am. 126(3), 1552–1562 (2009). doi:10.1121/1.3177272
Caldwell, M.C., Caldwell, D.K.: Individualized whistle contours in bottlenosed dolphins (Tursiops truncatus). Nature 207(4995), 434–435 (1965). doi:10.1038/207434a0
Cook, M.L.H., Sayigh, L.S., Blum, J.E., Wells, R.S.: Signature-whistle production in undisturbed free-ranging bottlenose dolphins (Tursiops truncatus). Proc. R. Soc. Lond. Ser. B Biol. Sci. 271(1543), 1043–1049 (2004). doi:10.1098/rspb.2003.2610
Janik, V.M., Sayigh, L.S.: Communication in bottlenose dolphins: 50 years of signature whistle research. J. Comp. Physiol. A 199(6), 479–489 (2013)
King, S.L., Harley, H.E., Janik, V.M.: The role of signature whistle matching in bottlenose dolphins, Tursiops truncatus. Anim. Behav. 96, 79–86 (2014). doi:10.1016/j.anbehav.2014.07.019
Kriesell, H.J., Elwen, S.H., Nastasi, A., Gridley, T.: Identification and characteristics of signature whistles in wild bottlenose dolphins (Tursiops truncatus) from Namibia. PLoS ONE 9(9), e106317 (2014). doi:10.1371/journal.pone.0106317
Sayigh, L.S., Esch, H.C., Wells, R.S., Janik, V.M.: Facts about signature whistles of bottlenose dolphins, Tursiops truncatus. Anim. Behav. 74, 1631–1642 (2007)
Sayigh, L.S., Tyack, P.L., Wells, R.S., Scott, M.D.: Signature whistles of free-ranging bottlenose dolphins Tursiops truncatus: stability and mother-offspring comparisons. Behav. Ecol. Sociobiol. 26, 247–260 (1990)
Smolker, R.A., Mann, J., Smuts, B.B.: Use of signature whistles during separations and reunions by wild bottlenose dolphin mothers and infants. Behav. Ecol. Sociobiol. 33, 393–402 (1993)
Kuczaj II, S.A., Eskelinen, H.C., Jones, B.L., Borger-Turner, J.L.: Gotta go, mom’s calling: Dolphin (Tursiops truncatus) mothers use individually distinctive acoustic signals to call their calves. Anim. Behav. Cogn. 2(1), 88–95 (2015). doi:10.12966/abc.02.07.2015
King, S.L., Guarino, E., Donegan, K., Hecksher, J., Jaakkola, K.: Further insights into postpartum signature whistle use in bottlenose dolphins (Tursiops truncatus). Mar. Mamm. Sci. 32(4), 1458–1469 (2016). doi:10.1111/mms.12317
Luís, A.R., Couchinho, M.N., Santos, M.E.d.: A quantitative analysis of pulsed signals emitted by wild bottlenose dolphins. PLoS ONE 11(7) (2016). doi:10.1371/journal.pone.0157781
Au, W., Branstetter, B., Moore, P., Finneran, J.: Dolphin biosonar signals measured at extreme off-axis angles: insights to sound propagation in the head. J. Acoust. Soc. Am. 132(2), 1199–1206 (2012). doi:10.1121/1.4730901
Simard, P., Hibbard, A.L., McCallister, K.A., Frankel, A.S., Zeddies, D.G., Sisson, G.M., Gowans, S., Forys, E.A., Mann, D.A.: Depth dependent variation of the echolocation pulse rate of bottlenose dolphins (Tursiops truncatus). J. Acoust. Soc. Am. 127(1), 568–578 (2010). doi:10.1121/1.3257202
McAlpine, D.F.: Pygmy and dwarf sperm whales: Kogia breviceps and K. sima. In: Würsig, B., Thewissen, J.G.M. (eds.) Encyclopedia of Marine Mammals, pp. 936–938. Academic Press, London (2009)
Marten, K.: Ultrasound analysis of pygmy sperm whale (Kogia breviceps) and Hubbs’ beaked whale (Mesoplodon carlhubbsi) clicks. Aquat. Mamm. 26(1), 45–48 (2000)
Ridgway, S.H., Carder, D.A.: Assessing hearing and sound production in cetaceans not available for behavioral audiograms: experiences with sperm, pygmy sperm, and gray whales. Aquat. Mamm. 27(3), 267–276 (2001)
Madsen, P.T., Carder, D.A., Bedholm, K., Ridgway, S.H.: Porpoise clicks from a sperm whale nose—convergent evolution of 130 kHz pulses in toothed whale sonars? Bioacoustics 15(2), 195–206 (2005)
Thomas, J.A., Moore, P.W.B., Nachtigall, P.E., Gilmartin, W.G.: A new sound from a stranded pygmy sperm whale. Aquat. Mamm. 16(1), 28–30 (1990)
Caldwell, D.K., Prescott, J.H., Caldwell, M.C.: Production of pulsed sounds by the pygmy sperm whale, Kogia breviceps. Bull. South. Calif. Acad. Sci. 65(4), 245–248 (1966)
Department of the Environment: Physeter macrocephalus. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Backus, R.H., Schevill, W.E.: Physeter clicks. In: Norris, K.S. (ed.) Whales, Dolphins, and Porpoises, pp. 510–528. University of California Press, Berkeley (1966)
Tran, D.D., Huang, W., Bohn, A.C., Wang, D., Gong, Z., Makris, N.C.: Using a coherent hydrophone array for observing sperm whale range, classification, and shallow-water dive profiles. J. Acoust. Soc. Am. 135(6), 3352–3363 (2014). doi:10.1121/1.4874601
Watkins, W.A.: Acoustic behavior of sperm whales. Oceanus 20(2), 50–58 (1977)
Watkins, W.A.: Acoustics and the behavior of sperm whales. In: Busnel, R.-G., Fish, J.F. (eds.) Animal Sonar Systems, pp. 283–290. Plenum Press, New York (1980). doi:10.1121/1.381678
Watkins, W.A., Schevill, W.E.: Sperm whale codas. J. Acoust. Soc. Am. 62(6), 1485–1490 (1977)
Levenson, C.: Source level and bistatic target strength of the sperm whale (Physeter catodon) measured from an oceanographic aircraft. J. Acoust. Soc. Am. 55(5), 1100–1103 (1974). doi:10.1121/1.1914660
Mullins, J., Whitehead, H., Weilgart, L.S.: Behavior and vocalizations of two single sperm whales, Physeter macrocephalus, off Nova Scotia. Can. J. Fish. Aquat. Sci. 45(10), 1736–1743 (1988)
Morrissey, R.P., Ward, J., DiMarzio, N., Jarvis, S., Moretti, D.J.: Passive acoustic detection and localization of sperm whales (Physeter macrocephalus) in the tongue of the ocean. Appl. Acoust. 67(11–12), 1091–1105 (2006). doi:10.1016/j.apacoust.2006.05.014
Nosal, E.M., Frazer, L.N.: Sperm whale three-dimensional track, swim orientation, beam pattern, and click levels observed on bottom-mounted hydrophones. J. Acoust. Soc. Am. 122(4), 1969–1978 (2007). doi:10.1121/1.2775423
Ward, J., Thomas, L., Jarvis, S., Di Marzio, N., Moretti, D., Marques, T., Dunn, C., Iaridge, D., Hartvig, E., Tyack, P.: Passive acoustic density estimation of sperm whales in the Tongue of the Ocean, Bahamas. Mar. Mamm. Sci. 28(4), E444–E455 (2012). doi:10.1111/j.1748-7692.2011.00560.x
Dunn, L.J.L.: Sperm whale acoustic characteristic measurements from the Asweps aircraft. Naval Oceanographic Office, Informal Report(69-14), 9 (1969)
Moore, K.E., Watkins, W.A., Tyack, P.L.: Pattern similarity in shared codas from sperm whales (Physeter catodon). Mar. Mamm. Sci. 9(1), 1–9 (1993)
Schulz, T.M., Whitehead, H., Gero, S., Rendell, L.: Individual vocal production in a sperm whale (Physeter macrocephalus) social unit. Mar. Mamm. Sci. 27(1), 149–166 (2011). doi:10.1111/j.1748-7692.2010.00399.x
Smultea, M.A., Holst, M., Koski, W.R., Roi, S.S., Sayegh, A.J., Fossati, C., Goldstein, H.H., Beland, J.A., Maclean, S., Yin, S.: Visual-acoustic survey of cetaceans during a seismic study in the Southeast Caribbean Sea, April–June 2004. Caribb. J. Sci. 47(2–3), 273–283 (2013)
Watkins, W.A., Moore, K.E., Tyack, P.: Sperm whale acoustic behaviors in the Southeast Caribbean. Cetology 49, 1–15 (1985)
Whitehead, H., Weilgart, L.: Patterns of visually observable behavior and vocalizations in groups of female sperm whales. Behaviour 118(3/4), 275–296 (1991)
Ackleh, A., Ioup, G., Ioup, J., Ma, B.: Assessing the Deepwater Horizon oil spill impact on marine mammal population through acoustics: endangered sperm whales. J. Acoust. Soc. Am. 131(3), 2306–2314 (2012). doi:10.1121/1.3682042
Antunes, R., Rendell, L., Gordon, J.: Measuring inter-pulse intervals in sperm whale clicks: consistency of automatic estimation methods. J. Acoust. Soc. Am. 127(5), 3239–3247 (2010). doi:10.1121/1.3327509
Antunes, R., Schulz, T., Gero, S., Whitehead, H., Gordon, J., Rendell, L.: Individually distinctive acoustic features in sperm whale codas. Anim. Behav. 81(4), 723–730 (2011). doi:10.1016/j.anbehav.2010.12.019
Thode, A., Mellinger, D.K., Stienessen, S., Martinez, A., Mullin, K.: Depth-dependent acoustic features of diving sperm whales (Physeter macrocephalus) in the Gulf of Mexico. J. Acoust. Soc. Am. 112(1), 308–321 (2002). doi:10.1121/1.1482077
Mathias, D., Thode, A., Straley, J., Calambokidis, J., Schorr, G., Folkert, K.: Acoustic and diving behavior of sperm whales (Physeter macrocephalus) during natural and depredation foraging in the Gulf of Alaska. J. Acoust. Soc. Am. 132(1), 518–532 (2012). doi:10.1121/1.4726005
Mathias, D., Thode, A., Straley, J., Folkert, K.: Relationship between sperm whale (Physeter macrocephalus) click structure and size derived from videocamera images of a depredating whale (sperm whale prey acquisition). J. Acoust. Soc. Am. 125(5), 3444–3453 (2009). doi:10.1121/1.3097758
Mathias, D., Thode, A.M., Straley, J., Andrews, R.D.: Acoustic tracking of sperm whales in the Gulf of Alaska using a two-element vertical array and tags. J. Acoust. Soc. Am. 134(3), 2446–2461 (2013). doi:10.1121/1.4816565
Mellinger, D.K., Stafford, K.M., Fox, C.G.: Seasonal occurrence of sperm whale (Physeter macrocephalus) sounds in the Gulf of Alaska, 1999–2001. Mar. Mamm. Sci. 20(1), 48–62 (2004). doi:10.1111/j.1748-7692.2004.tb01140.x
Thode, A.M., Wild, L., Mathias, D., Straley, J., Lunsford, C.: A comparison of acoustic and visual metrics of sperm whale longline depredation. J. Acoust. Soc. Am. 135(5), 3086–3100 (2014). doi:10.1121/1.4869853
Barlow, J., Taylor, B.L.: Estimates of sperm whale abundance in the northeastern temperate Pacific from a combined acoustic and visual survey. Mar. Mamm. Sci. 21(3), 429–445 (2005). doi:10.1111/j.1748-7692.2005.tb01242.x
Hirotsu, R., Yanagisawa, M., Ura, T., Sakata, M., Sugimatsu, H., Kojima, J., Bahl, R.: Localization of sperm whales in a group using clicks received at two separated short baseline arrays. J. Acoust. Soc. Am. 127(1), 133–147 (2010). doi:10.1121/1.3268593
Masao, A., Kourogi, A., Aoki, K., Yoshioka, M., Kori, K.: Differences in sperm whale codas between two waters off Japan: possible geographic separation of vocal clans. J. Mammal. 95(1), 169–175 (2014). doi:10.1644/13-MAMM-A-172
Au, W.W.L., Giorli, G., Chen, J., Copeland, A., Lammers, M.O., Richlen, M., Jarvis, S., Morrissey, R., Moretti, D.: Presence and seasonal variation of deep diving foraging odontocetes around Kauai, Hawaii using remote autonomous acoustic recorders. J. Acoust. Soc. Am. 135(1), 521–530 (2014). doi:10.1121/1.4836575
Weilgart, L., Whitehead, H.: Coda communication by sperm whales (Physeter macrocephalus) off the Galapagos Islands. Can. J. Zool. 71, 744–752 (1993). doi:10.1139/z93-098
Weilgart, L.S., Whitehead, H.: Distinctive vocalizations from mature male sperm whales (Physeter macrocephalus). Can. J. Zool. 66, 1931–1937 (1988). doi:10.1139/z88-282
Goold, J.C., Jones, S.E.: Time and frequency domain characteristics of sperm whale clicks. J. Acoust. Soc. Am. 98(3), 1279–1291 (1995). doi:10.1121/1.413465
Goold, J.C.: Behavioural and acoustic observations of sperm whales in Scapa Flow, Orkney Islands. J. Mar. Biol. Assoc. UK 79(3), 541–550 (1999)
Madsen, P.T., Wahlberg, M., Møhl, B.: Male sperm whale (Physeter macrocephalus) acoustics in a high-latitude habitat: implications for echolocation and communication. Behav. Ecol. Sociobiol. 53(1), 31–41 (2002). doi:10.1007/s00265-002-0548-1
Møhl, B., Wahlberg, M., Madsen, P.T., Heerfordt, A., Lund, A.: The monopulsed nature of sperm whale clicks. J. Acoust. Soc. Am. 114(2), 1143–1154 (2003). doi:10.1121/1.1586258
Møhl, B., Wahlberg, M., Madsen, P.T., Miller, L.A., Surlykke, A.: Sperm whale clicks: directionality and source level revisited. J. Acoust. Soc. Am. 107(1), 638–648 (2000). doi:10.1121/1.428329
Teloni, V., Mark, J.P., Patrick, M.J.O., Peter, M.T.: Shallow food for deep divers: dynamic foraging behavior of male sperm whales in a high latitude habitat. J. Exp. Mar. Biol. Ecol. 354(1), 119–131 (2008). doi:10.1016/j.jembe.2007.10.010
Wahlberg, M.: The acoustic behaviour of diving sperm whales observed with a hydrophone array. J. Exp. Mar. Biol. Ecol. 281(1–2), 53–62 (2002)
Lewis, T., Gillespie, D., Matthews, L.J., Danbolt, M., Leaper, R., McLanaghan, R., Moscrop, A.: Sperm whale abundance estimates from acoustic surveys of the Ionian Sea and Straits of Sicily in 2003. J. Mar. Biol. Assoc. UK 87(1), 353–357 (2007). doi:10.1017/S0025315407054896
Pavan, G., Hayward, T.J., Borsani, J.F., Priano, M., Manghi, M., Fossati, C., Gordon, J.: Time patterns of sperm whale codas recorded in the Mediterranean Sea 1985–1996. J. Acoust. Soc. Am. 107(6), 3487–3495 (2000). doi:10.1121/1.429419
Teloni, V.: Patterns of sound production in diving sperm whales in the northwestern Mediterranean. Mar. Mamm. Sci. 21(3), 446–457 (2005). doi:10.1111/j.1748-7692.2005.tb01243.x
Zaugg, S., van der Schaar, M., Houegnigan, L., Gervaise, C., Andre, M.: Real-time acoustic classification of sperm whale clicks and shipping impulses from deep-sea observatories. Appl. Acoust. 71(11), 1011–1019 (2010). doi:10.1016/j.apacoust.2010.05.005
Zimmer, W.M.X., Tyack, P.L., Johnson, M.P., Madsen, P.T.: Three-dimensional beam pattern of regular sperm whale clicks confirms bent-horn hypothesis. J. Acoust. Soc. Am. 117(3), 1473–1485 (2005). doi:10.1121/1.1828501
Weilgart, L., Whitehead, H.: Group-specific dialects and geographical variation in coda repertoire in South Pacific sperm whales. Behav. Ecol. Sociobiol. 40(5), 277–285 (1997)
Douglas, L.A., Dawson, S.M., Jaquet, N.: Click rates and silences of sperm whales at Kaikoura. New Zealand. J. Acoust. Soc. Am. 118(1), 523–529 (2005). doi:10.1121/1.1937283
Growcott, A., Miller, B., Sirguey, P., Slooten, E., Dawson, S.: Measuring body length of male sperm whales from their clicks: the relationship between inter-pulse intervals and photogrammetrically measured lengths. J. Acoust. Soc. Am. 130(1), 568–573 (2011). doi:10.1121/1.3578455
Jaquet, N., Dawson, S., Douglas, L.: Vocal behaviour of male sperm whales: why do they click? J. Acoust. Soc. Am. 109(5), 2254–2259 (2001). doi:10.1121/1.1360718
Miller, B., Dawson, S., Vennell, R.: Underwater behavior of sperm whales off Kaikoura, New Zealand, as revealed by a three-dimensional hydrophone array. J. Acoust. Soc. Am. 134(4), 2690–2700 (2013). doi:10.1121/1.4818896
Miller, B.S., Growcott, A., Slooten, E., Dawson, S.M.: Acoustically derived growth rates of sperm whales (Physeter macrocephalus) in Kaikoura, New Zealand. J. Acoust. Soc. Am. 134(3), 2438–2445 (2013). doi:10.1121/1.4816564
Gedamke, J., Robinson, S.M.: Acoustic survey for marine mammal occurrence and distribution off East Antarctica (30–80\(^{\circ }\)E) in January-February 2006. Deep Sea Res. Pt II Top. Stud. Oceanogr. 57(9–10), 968–981 (2010). doi:10.1016/j.dsr2.2008.10.042
Worthington, L.V., Schevill, W.E.: Underwater sounds heard from sperm whales. Nature (London) 180(4580), 291 (1957). doi:10.1038/180291a0
Rendell, L.E., Whitehead, H.: Vocal clans in sperm whales (Physeter macrocephalus). Proc. R. Soc. Lond. Ser. B Biol. Sci. 270(1512), 225–231 (2003). doi:10.1098/rspb.2002.2239
Fristrup, K.M., Harbison, G.R.: How do sperm whales catch squids? Mar. Mamm. Sci. 18(1), 42–54 (2002). doi:10.1111/j.1748-7692.2002.tb01017.x
Oliveira, C., Wahlberg, M., Johnson, M., Miller, P.J.O., Madsen, P.T.: The function of male sperm whale slow clicks in a high latitude habitat: communication, echolocation, or prey debilitation? J. Acoust. Soc. Am. 133(5), 3135–3144 (2013). doi:10.1121/1.4795798
Whitehead, H.: Sperm whale. In: Perrin, W.F., Würsig, B., Thjewissen, J.G.M. (eds.) Encyclopedia of Marine Mammals, pp. 1165–1172. Academic Press, New York (2002)
Fais, A., Johnson, M., Wilson, M., Soto, N.A., Madsen, P.T.: Sperm whale predator-prey interactions involve chasing and buzzing, but no acoustic stunning. Sci. Rep. (2016). doi:10.1038/srep28562
Gero, S., Whitehead, H., Rendell, L.: Individual, unit and vocal clan level identity cues in sperm whale codas. Open Sci. 3(1), 150372 (2016). doi:10.1098/rsos.150372
Oliveira, C., Wahlberg, M., Silva, M.A., Johnson, M., Antunes, R., Wisniewska, D.M., Fais, A., Gonçalves, J., Madsen, P.T.: Sperm whale codas may encode individuality as well as clan identity. J. Acoust. Soc. Am. 139(5), 2860–2869 (2016). doi:10.1121/1.4949478
Weir, C.R., Frantzis, A., Alexiadou, P., Goold, J.C.: The burst-pulse nature of ’squeal’ sounds emitted by sperm whales (Physeter macrocephalus). J. Mar. Biol. Assoc. UK 87(1), 39–46 (2007). doi:10.1017/S0025315407054549
Rhinelander, M.Q., Dawson, S.M.: Measuring sperm whales from their clicks: stability of interpulse intervals and validation that they indicate whale length. J. Acoust. Soc. Am. 115(4), 1826–1831 (2004). doi:10.1121/1.1689346
Whitehead, H., Weilgart, L.: Click rates from sperm whales. J. Acoust. Soc. Am. 87(4), 1798–1806 (1990). doi:10.1121/1.399376
Madsen, P.T., Carder, D.A., Au, W.W.L., Nachtigall, P.E., Møhl, B., Ridgway, S.H.: Sound production in neonate sperm whales (L). J. Acoust. Soc. Am. 113(6), 2988–2991 (2003). doi:10.1121/1.1572137
Schulz, T.M., Whitehead, H., Rendell, L.: Off-axis effects on the multi-pulse structure of sperm whale coda clicks. J. Acoust. Soc. Am. 125(3), 1768–1773 (2009). doi:10.1121/1.3075598
Zimmer, W.M.X., Madsen, P.T., Teloni, V., Johnson, M.P., Tyack, P.L.: Off-axis effects on the multipulse structure of sperm whale usual clicks with implications for sound production. J. Acoust. Soc. Am. 118(5), 3337–3345 (2005). doi:10.1121/1.2082707
Department of the Environment: Berardius arnuxii http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Hobson, R.P., Martin, A.R.: Behaviour and dive times of Arnoux’s beaked whales, Berardius arnuxii, at narrow leads in fast ice. Can. J. Zool. 74, 388–393 (1996)
Rogers, T.L., Brown, S.M.: Acoustic observations of Arnoux’s beaked whale (Berardius arnuxii) off Kemp Land, Antarctica (Note). Mar. Mamm. Sci. 15(1), 192–198 (1999)
Leaper, R., Gillespie, D., Papastavrou, V.: Results of passive acoustic surveys for odontocetes in the Southern Ocean. J. Cetacean Res. Manag. 2(3), 187–196 (2000)
Leaper, R., Scheidat, M.: An acoustic survey for cetaceans in the Southern Ocean Sanctuary conducted from the German Government research vessel Polarstern. Rep. Int. Whal. Comm. 48, 431–437 (1998)
Anderson, R.C., Clark, R., Madsen, P.T., Johnson, C.: Observations of Longman’s beaked whale (Indopacetus pacificus) in the Western Indian Ocean. Aquat. Mamm. 32(2), 223 (2006). doi:10.1578/AM.32.2.2006.223
Dalebout, M.L., Baker, C.S., Anderson, R.C., Best, P.B., Cockcroft, V.G., Hinsz, H.L., Peddemors, V., Pitman, R.L.: Appearance, distribution and genetic distinctiveness of Longman’s beaked whale, Indopacetus pacificus. Mar. Mamm. Sci. 19(3), 421–461 (2003). doi:10.1111/j.1748-7692.2003.tb01314.x
Rankin, S., Baumann-Pickering, S., Yack, T., Barlow, J.: Description of sounds recorded from Longman’s beaked whale, Indopacetus pacificus. J. Acoust. Soc. Am. 130(5), EL339–EL344 (2011)
Baker, A.N.: Status, relationships, and distribution of Mesoplodon bowdoini Andrews, 1908 (Cetacea: Ziphiidae). Mar. Mamm. Sci. 17(3), 473–493 (2001). doi:10.1111/j.1748-7692.2001.tb00999.x
Tyack, P.L., Johnson, M.P., Zimmer, W.M.X., de Soto, N.A., Madsen, P.T.: Acoustic behavior of beaked whales, with implications for acoustic monitoring. In: Oceans-IEEE, pp. 509–514 (2006)
Aguilar de Soto, N., Madsen, P., Tyack, P., Arranz, P., Marrero, J., Fais, A., Revelli, E., Johnson, M.: No shallow talk: cryptic strategy in the vocal communication of Blainville’s beaked whales. Mar. Mamm. Sci. 28(2), E75–E92 (2012). doi:10.1111/j.1748-7692.2011.00495.x
Johnson, M., Madsen, P.T., Zimmer, W.M.X., de Soto, N.A., Tyack, P.L.: Beaked whales echolocate on prey. Proc. R. Soc. Lond. Ser. B Biol. Sci. 271, S383–S386 (2004). doi:10.1098/rsbl.2004.0208
Johnson, M., Madsen, P.T., Zimmer, W.M.X., de Soto, N.A., Tyack, P.L.: Foraging Blainville’s beaked whales (Mesoplodon densirostris) produce distinct click types matched to different phases of echolocation. J. Exp. Biol. 209(24), 5038–5050 (2006). doi:10.1242/Jeb.02596
Madsen, P.T., Aguilar de Soto, N., Arranz, P., Johnson, M.: Echolocation in Blainville’s beaked whales (Mesoplodon densirostris). J. Comp. Physiol. A 199(6), 451–469 (2013)
Dunn, C., Hickmott, L., Talbot, D., Boyd, I., Rendell, L.: Mid-frequency broadband sounds of Blainville’s beaked whales. Bioacoustics 22(2), 153–163 (2013)
Shaffer, J.W., Moretti, D., Jarvis, S., Tyack, P., Johnson, M.: Effective beam pattern of the Blainville’s beaked whale (Mesoplodon densirostris) and implications for passive acoustic monitoring. J. Acoust. Soc. Am. 133(3), 1770–1784 (2013). doi:10.1121/1.4776177
Rankin, S., Barlow, J.: Sounds recorded in the presence of Blainville’s beaked whales, Mesoplodon densirostris, near Hawai’i (L). J. Acoust. Soc. Am. 122(1), 42–45 (2007). doi:10.1121/1.2743159
Madsen, P.T., Johnson, M., de Soto, N.A., Zimmer, W.M.X., Tyack, P.: Biosonar performance of foraging beaked whales (Mesoplodon densirostris). J. Exp. Biol. 208(2), 181–194 (2005). doi:10.1242/Jeb.01327
Johnson, M., Hickmott, L.S., Aguilar Soto, N., Madsen, P.T.: Echolocation behaviour adapted to prey in foraging Blainville’s beaked whale (Mesoplodon densirostris). Proc. R. Soc. B Biol. Sci. 275, 133–139 (2008). doi:10.1098/rspb.2007.1190
Department of the Environment: Mesoplodon ginkgodens. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Macleod, C.D., Perrin, W.F., Pitman, R., Barlow, J., Ballance, L., Gerrodette, T., Joyce, G., Palka, D.L.: Known and inferred distributions of beaked whale species (Cetacea: Ziphiidae). J. Cetacean Res. Manag. 7(3), 271 (2006)
Chit, A.M., Smith, B.D., Tun, M.T.: An extreme extra-limital record of a strap-toothed beaked whale Mesoplodon layardii from the northern Indian Ocean in Myanmar. Mar. Biodivers. Rec. 5, 1–3 (2012). doi:10.1017/S1755267211001060
Pitman, R.L., Van Helden, A.L., Best, P.B., Pym, A.: Shepherd’s beaked whale (Tasmacetus shepherdi): information on appearance and biology based on strandings and at-sea observations. Mar. Mamm. Sci. 22(3), 744–755 (2006). doi:10.1111/j.1748-7692.2006.00066.x
Dalebout, M.L., Robertson, K.M., Frantzis, A., Engelhaupt, D.A.N., Mignucci-Giannoni, A.A., Rosario-Delestre, R.J., Baker, C.S.: Worldwide structure of mtDNA diversity among Cuvier’s beaked whales (Ziphius cavirostris): implications for threatened populations. Mol. Ecol. 14(11), 3353–3371 (2005). doi:10.1111/j.1365-294X.2005.02676.x
Frantzis, A., Goold, J.C., Skarsoulis, E.K., Taroudakis, M.I., Kandia, V.: Clicks from Cuvier’s beaked whales, Ziphius cavirostris. J. Acoust. Soc. Am. 112(1), 34–37 (2002). doi:10.1121/1.1479149
Zimmer, W.M.X., Johnson, M.P., Madsen, P.T., Tyack, P.L.: Echolocation clicks of free-ranging Cuvier’s beaked whales (Ziphius cavirostris). J. Acoust. Soc. Am. 117(6), 3919–3927 (2005). doi:10.1121/1.1910225
Schorr, G.S., Falcone, E.A., Moretti, D.J., Andrews, R.D.: First long-term behavioral records from Cuvier’s beaked whales (Ziphius cavirostris) reveal record-breaking dives. PLoS ONE 9(3), e92633 (2014). doi:10.1371/journal.pone.0092633
Department of the Environment: Dugong dugon. http://www.environment.gov.au/sprat (2017). Accessed 16 Mar 2017
Allen, S.G., Marsh, H., Hodgson, A.J.: Occurrence and conservation of the dugong (Sirenia: Dugongidae) in New South Wales. Proc. Linn. Soc. NSW 125, 211–216 (2004)
Anderson, P.K., Barclay, R.M.R.: Acoustic signals of solitary dugongs: physical characteristics and behavioural correlates. J. Mammal. 76(4), 1226–1237 (1995). doi:10.2307/1382616
Parsons, M.J.G., Holley, D., McCauley, R.D.: Source levels of dugong (Dugong dugon) vocalizations recorded in Shark Bay. J. Acoust. Soc. Am. 134(3), 2582–2588 (2013). doi:10.1121/1.4816583
Ichikawa, K., Arai, N., Akamatsu, T., Shinke, T., Hara, T., Adulyanukosol, K.: Acoustical analyses on the calls of dugong. Paper presented at the 4th SEASTAR 2000 Workshop, Bangkok, Thailand
Tsutsumi, C., Ichikawa, K., Arai, N., Akamatsu, T., Shinke, T., Hara, T., Adulyanukosol, K.: Feeding behavior of wild dugongs monitored by a passive acoustical method. J. Acoust. Soc. Am. 120(3), 1356–1360 (2006). doi:10.1121/1.2221529
Harcourt, R.G., Bradshaw, C.J., Dickson, K., Davis, L.S.: Foraging ecology of a generalist predator, the female New Zealand fur seal. Mar. Ecol. Prog. Ser. 227, 11–24 (2002)
Abbott, I.: The past and present distribution and status of sea lions and fur seals in Western Australia. Rec. West. Aust. Mus. 7(4), 375–390 (1979)
Shaughnessy, P.D., Gales, N., Dennis, T., Goldsworthy, S.: Distribution and abundance of New Zealand fur seals, Arctocephalus forsteri, in South Australia and Western Australia. Wildl. Res. 21(6), 667–695 (1994)
Stirling, I., Warneke, R.M.: Implications of a comparison of the airborne vocalizations and some aspects of the behaviour of the two Australian fur seals, Arctocephalus spp. on the evolution and present taxonomy of the genus. Aust. J. Zool. 19(3), 227–241 (1971). doi:10.1071/ZO9710227
Page, B., Goldsworthy, S.D., Hindell, M.A.: Vocal traits of hybrid fur seals: intermediate to their parental species. Anim. Behav. 61(5), 959–967 (2001). doi:10.1006/anbe.2000.1663
Page, B., Goldsworthy, S.D., Hindell, M.A.: Individual vocal traits of mother and pup fur seals. Bioacoustics 13(2), 121–143 (2002). doi:10.1080/09524622.2002.9753491
Page, B., Goldsworthy, S.D., Hindell, M.A., Mckenzie, J.: Interspecific differences in male vocalizations of three sympatric fur seals (Arctocephalus spp.). J. Zool. 258(1), 49–56 (2002). doi:10.1017/S095283690200119X
Hofmeyr, G.J.G.: Arctocephalus gazella. The IUCN Red List of Threatened Species e.T2058A66993062 (2016). doi:10.2305/IUCN.UK.2016-1.RLTS.T2058A66993062.en
St Clair Hill, M., Ferguson, J.W.H., Bester, M.N., Kerley, G.I.H.: Preliminary comparison of calls of the hybridizing fur seals Arctocephalus tropicalis and A. gazella. Afr. Zool. 36(1), 45–53 (2001). doi:10.1080/15627020.2001.11657113
Aubin, T., Jouventin, P., Charrier, I.: Mother vocal recognition in Antarctic fur seal Arctocephalus gazella pups: a two-step process. PLoS ONE 10(9), e0134513 (2015). doi:10.1371/journal.pone.0134513
Kirkwood, R., Pemberton, D., Gales, R., Hoskins, A.J., Mitchell, T., Shaughnessy, P.D., Arnould, J.P.: Continued population recovery by Australian fur seals. Mar. Freshw. Res. 61(6), 695–701 (2010)
Arnould, J.P., Hindell, M.A.: Dive behaviour, foraging locations, and maternal-attendance patterns of Australian fur seals (Arctocephalus pusillus doriferus). Can. J. Zool. 79(1), 35–48 (2001)
Kirkwood, R., Gales, R., Terauds, A., Arnould, J.P., Pemberton, D., Shaughnessy, P.D., Mitchell, A.T., Gibbens, J.: Pup production and population trends of the Australian fur seal (Arctocephalus pusillus doriferus). Mar. Mamm. Sci. 21(2), 260–282 (2005). doi:10.1111/j.1748-7692.2005.tb01227.x
Tripovich, J.S., Canfield, R., Rogers, T.L., Arnould, J.P.Y.: Characterization of Australian fur seal vocalizations during the breeding season. Mar. Mamm. Sci. 24(4), 913–928 (2008). doi:10.1111/j.1748-7692.2008.00229.x
Tripovich, J.S., Rogers, T.L., Arnould, J.P.Y.: Species-specific characteristics and individual variation of the bark call produced by male Australian fur seals, Arctocephalus pusillus doriferus. Bioacoustics 15(1), 79–96 (2005). doi:10.1080/09524622.2005.9753539
Tripovich, J., Canfield, R., Rogers, T., Arnould, J.P.: Individual variation of the female attraction call produced by Australian fur seal pups throughout the maternal dependence period. Bioacoustics 18(3), 259–276 (2009). doi:10.1080/09524622.2009.9753605
Gales, N., Coughran, D., Queale, L.: Records of subantarctic fur seals Arctocephalus tropicalis in Australia. Aust. Mammal. 15, 135–138 (1992)
Hofmeyr, G.J.G.: Arctocephalus tropicalis. The IUCN Red List of Threatened Species e.T2062A45224547 (2015). doi:10.2305/IUCN.UK.2015-4.RLTS.T2062A45224547.en
Charrier, I., Mathevon, N., Jouventin, P.: Individuality in the voice of fur seal females: an analysis study of the pup attraction call in Arctocephalus tropicalis. Mar. Mamm. Sci. 19(1), 161–172 (2003). doi:10.1111/j.1748-7692.2003.tb01099.x
Charrier, I., Mathevon, N., Jouventin, P.: How does a fur seal mother recognize the voice of her pup? An experimental study of Arctocephalus tropicalis. J. Exp. Biol. 205(5), 603–612 (2002)
Gwilliam, J., Charrier, I., Harcourt, R.G.: Vocal identity and species recognition in male Australian sea lions, Neophoca cinerea. J. Exp. Biol. 211(14), 2288 (2008). doi:10.1242/jeb.013185
Gales, N., Cheal, A., Pobar, G., Williamson, P.: Breeding biology and movements of Australian sea-lions, Neophoca cinerea, off the west coast of Western Australia. Wildl. Res. 19(4), 405–415 (1992). doi:10.1071/WR9920405
Costa, D.P., Gales, N.J.: Energetics of a benthic diver: seasonal foraging ecology of the Australian sea lion, Neophoca cinerea. Ecol. Monogr. 73(1), 27–43 (2003). doi:10.1890/0012-9615(2003)073[0027:EOABDS]2.0.CO;2
Shaughnessy, P.D., Dennis, T., Seager, P.: Status of Australian sea lions, Neophoca cinerea, and New Zealand fur seals, Arctocephalus forsteri, on Eyre Peninsula and the far west coast of South Australia. Wildl. Res. 32(1), 85–101 (2005). doi:10.1071/WR03068
Stirling, I.: Observations on the Australian sea lion, Neophoca cinerea (Peron). Aust. J. Zool. 20(3), 271–279 (1972). doi:10.1071/ZO9720271
Marlow, B.: The comparative behaviour of the Australasian sea lions Neophoca cinerea and Phocarctos hookeri (Pinnipedia: Otariidae). Mammalia 39(2), 159–230 (1975). doi:10.1515/mamm.1975.39.2.159
Attard, M.R.G., Pitcher, B.J., Charrier, I., Ahonen, H., Harcourt, R.G.: Vocal discrimination in mate guarding male Australian sea lions: familiarity breeds contempt. Ethology 116(8), 704–712 (2010). doi:10.1111/j.1439-0310.2010.01786.x
Ahonen, H., Stow, A.J., Harcourt, R.G., Charrier, I.: Adult male Australian sea lion barking calls reveal clear geographical variations. Anim. Behav. 97, 229–239 (2014). doi:10.1016/j.anbehav.2014.09.010
Charrier, I., Ahonen, H., Harcourt, R.G.: What makes an Australian sea lion (Neophoca cinerea) male’s bark threatening? J. Comp. Psychol. 125(4), 385 (2011). doi:10.1037/a0024513
Charrier, I., Harcourt, R.G.: Individual vocal identity in mother and pup Australian sea lions (Neophoca cinerea). J. Mammal. 87(5), 929–938 (2006). doi:10.1644/05-MAMM-A-344R3.1
Charrier, I., Pitcher, B.J., Harcourt, R.G.: Vocal recognition of mothers by Australian sea lion pups: individual signature and environmental constraints. Anim. Behav. 78(5), 1127–1134 (2009). doi:10.1016/j.anbehav.2009.07.032
Pitcher, B.J., Ahonen, H., Harcourt, R.G., Charrier, I.: Delayed onset of vocal recognition in Australian sea lion pups (Neophoca cinerea). Naturwissenschaften 96(8), 901–909 (2009). doi:10.1007/s00114-009-0546-5
Pitcher, B., Harcourt, R., Charrier, I.: Individual identity encoding and environmental constraints in vocal recognition of pups by Australian sea lion mothers. Anim. Behav. 83(3), 681–690 (2012). doi:10.1016/j.anbehav.2011.12.012
Southwell, C., Paxton, C.G., Borchers, D., Boveng, P., Rogers, T., William, K.: Uncommon or cryptic? Challenges in estimating leopard seal abundance by conventional but state-of-the-art methods. Deep Sea Res. Pt I Oceanogr. Res. Pap. 55(4), 519–531 (2008)
Rogers, T.L., Ciaglia, M.B., Klinck, H., Southwell, C.: Density can be misleading for low-density species: benefits of passive acoustic monitoring. PLoS ONE 8(1), e52542 (2013). doi:10.1371/journal.pone.0052542
Forcada, J., Trathan, P.N., Boveng, P.L., Boyd, I.L., Burns, J.M., Costa, D.P., Fedak, M., Rogers, T.L., Southwell, C.J.: Responses of Antarctic pack-ice seals to environmental change and increasing krill fishing. Biol. Conserv. 149(1), 40–50 (2012). doi:10.1016/j.biocon.2012.02.002
Rogers, T., Bryden, M.: Density and haul-out behavior of leopard seals (Hydrurga leptonyx) in Prydz Bay, Antarctica. Mar. Mamm. Sci. 13(2), 293–302 (1997). doi:10.1111/j.1748-7692.1997.tb00632.x
Meade, J., Ciaglia, M.B., Slip, D.J., Negrete, J., Márquez, M.E.I., Mennucci, J., Rogers, T.L.: Spatial patterns in activity of leopard seals Hydrurga leptonyx in relation to sea ice. Mar. Ecol. Prog. Ser. 521, 265–275 (2015). doi:10.3354/meps11120
Rogers, T.L., Cato, D.H., Bryden, M.M.: Behavioral significance of underwater vocalizations of captive leopard seals, Hydrurga leptonyx. Mar. Mamm. Sci. 12(3), 414–427 (1996). doi:10.1111/j.1748-7692.1996.tb00593.x
Southwell, C., Kerry, K., Ensor, P., Woehler, E.J., Rogers, T.: The timing of pupping by pack-ice seals in East Antarctica. Polar Biol. 26(10), 648–652 (2003). doi:10.1007/s00300-003-0534-8
Stirling, I., Siniff, D.B.: Underwater vocalizations of leopard seals (Hydrurga leptonyx) and crabeater seals (Lobodon carcinophagus) near the South Shetland Islands, Antarctica. Can. J. Zool. 57(6), 1244–1248 (1979). doi:10.1139/z79-160
Rogers, T.L., Cato, D.H., Bryden, M.M.: Underwater vocal repertoire of the leopard seal, Hydrurga leptonyx, in Prydz Bay, Antarctica. In: Kastelein, R.A., Thomas, J.A., Nachtigal, P.E. (eds.) Sensory Abilities of Aquatic Animals. DeSpil Publishers, Amsterdam (1995)
Rogers, T.L.: Source levels of the underwater calls of a male leopard seal. J. Acoust. Soc. Am. (2014). doi:10.1121/1.4895685
Awbrey, F.T., Thomas, J.A., Evans, W.E.: Ultrasonic underwater sounds from a captive leopard seal (Hydrurga leptonyx). In: Thomas, J.A., Moss, C.F., Vater, M. (eds.) Echolocation in Bats and Dolphins. University of Chicago Press, Chicago (2002)
Rogers, T.L., Cato, D.H.: Individual variation in the acoustic behaviour of the adult male leopard seal, Hydrurga leptonyx. Behaviour 139(10), 1267–1286 (2002). doi:10.1163/156853902321104154
Rogers, T.L.: Age-related differences in the acoustic characteristics of male leopard seals, Hydrurga leptonyx. J. Acoust. Soc. Am. 122(1), 596–605 (2007). doi:10.1121/1.2736976
Thomas, J.A., Golladay, C.L.: Geographic variation in leopard seal (Hydrurga leptonyx) underwater vocalizations. In: Kastelein, R.A., Thomas, J.A., Nachtigall, P.E. (eds.) Sensory Systems of Aquatic Mammals. De Spil Publishers, Woerden (1995)
Kreiss, C.M., Boebel, O., Bornemann, H., Kindermann, L., Klinck, H., Klinck, K., Plötz, J., Rogers, T.L., Opzeeland, I.C.V.: Call characteristics of high-double trill leopard seal (Hydrurga leptonyx) vocalizations from three Antarctic locations. Polarforschung 83(2), 63–71 (2013)
Hückstädt, L.: Leptonychotes weddellii. The IUCN Red List of Threatened Species, e.T11696A45226713 (2015). doi:10.2305/IUCN.UK.2015-4.RLTS.T11696A45226713.en
Rice, D.W.: Marine Mammals of the World-Systematics and Distribution. The Society for Marine Mammalogy, Lawrence (1998)
Stirling, I.: Ecology of the Weddell seal in McMurdo Sound, Antarctica. Ecology 50(4), 573–586 (1969). doi:10.2307/1936247
Doiron, E., Rouget, P., Terhune, J.: Proportional underwater call type usage by Weddell seals (Leptonychotes weddellii) in breeding and nonbreeding situations. Can. J. Zool. 90(2), 237–247 (2012). doi:10.1139/z11-131
Green, K., Burton, H.R.: Annual and diurnal variations in the underwater vocalizations of Weddell seals. Polar Biol. 8(3), 161–164 (1988). doi:10.1007/BF00443447
Lindsey, A.A.: The Weddell seal in the Bay of Whales, Antarctica. J. Mammal. 18(2), 127–144 (1937). doi:10.2307/1374459
Schevill, W.E., Watkins, W.A.: Underwater calls of Leptonychotes (Weddell seal). Zoologica 50(1), 45–46 (1965)
Thomas, J.A., Kuechle, V.B.: Quantitative analysis of Weddell seal (Leptonychotes weddelli) underwater vocalizations at McMurdo Sound, Antarctica. J. Acoust. Soc. Am. 72(6), 1730–1738 (1982). doi:10.1121/1.388667
Pahl, B.C., Terhune, J.M., Burton, H.R.: Repertoire and geographic variation in underwater vocalisations of Weddell Seals (Leptonychotes weddellii, Pinnipedia: Phocidae) at the Vestfold Hills, Antarctica. Aust. J. Zool. 45(2), 171–187 (1997). doi:10.1071/ZO95044
Terhune, J.M., Dell’Apa, A.: Stereotyped calling patterns of a male Weddell seal (Leptonychotes weddellii). Aquat. Mamm. 32(2), 175 (2006). doi:10.1578/AM.32.2.2006.175
Morrice, M.G., Burton, H.R., Green, K.: Microgeographic variation and songs in the underwater vocalisation repertoire of the Weddell seal (Leptonychotes weddellii). Polar Biol. 14(7), 441–446 (1994). doi:10.1007/BF00239046
Moors, H.B., Terhune, J.M.: Repetition patterns in Weddell seal (Leptonychotes weddellii) underwater multiple element calls. J. Acoust. Soc. Am. 116(2), 1261–1270 (2004). doi:10.1121/1.1763956
Collins, K.T., Rogers, T.L., Terhune, J.M., McGreevy, P.D., Wheatley, K.E., Harcourt, R.G.: Individual variation of in-air female ’Pup Contact’ calls in Weddell Seals, Leptonychotes weddellii. Behaviour 142(2), 167–189 (2005). doi:10.1163/1568539053627668
Collins, K.T., Terhune, J.M., Rogers, T.L., Wheatley, K.E., Harcourt, R.G.: Vocal individuality of in-air Weddell seal (Leptonychotes weddellii) pup “primary” calls. Mar. Mamm. Sci. 22(4), 933–951 (2006). doi:10.1111/j.1748-7692.2006.00074.x
Van Opzeeland, I., Van Parijs, S., Frickenhaus, S., Kreiss, C., Boebel, O.: Individual variation in pup vocalizations and absence of behavioral signs of maternal vocal recognition in Weddell seals (Leptonychotes weddellii). Mar. Mamm. Sci. 28(2), E158–E172 (2012). doi:10.1111/j.1748-7692.2011.00505.x
Collins, K.T., Terhune, J.M.: Geographic variation of Weddell seal (Leptonychotes weddellii) airborne mother-pup vocalisations. Polar Biol. 30(11), 1373–1380 (2007). doi:10.1007/s00300-007-0297-8
Nordøy, E.S., Folkow, L., Blix, A.S.: Distribution and diving behaviour of crabeater seals (Lobodon carcinophagus) off Queen Maud Land. Polar Biol. 15(4), 261–268 (1995). doi:10.1007/BF00239846
McCreery, L., Thomas, J.A.: Acoustic analysis of underwater vocalizations from crabeater seals (Lobodon carcinophagus): not so monotonous. Aquat. Mamm. 35(4), 490 (2009). doi:10.1578/AM.35.4.2009.490
Klinck, H., Mellinger, D.K., Klinck, K., Hager, J., Kindermann, L., Boebel, O.: Long-range underwater vocalizations of the crabeater seal (Lobodon carcinophaga). J. Acoust. Soc. Am. 128(1), 474–479 (2010). doi:10.1121/1.3442362
McConnell, B., Chambers, C., Fedak, M.: Foraging ecology of southern elephant seals in relation to the bathymetry and productivity of the Southern Ocean. Antarct. Sci. 4(04), 393–398 (1992). doi:10.1017/S0954102092000580
Mcmahon, C.R., Burton, H.R., Bester, M.N.: A demographic comparison of two southern elephant seal populations. J. Anim. Ecol. 72(1), 61–74 (2003). doi:10.1046/j.1365-2656.2003.00685.x
Sanvito, S., Galimberti, F.: Bioacoustics of southern elephant seals. II. Individual and geographical variation in male aggressive vocalisations. Bioacoustics 10(4), 287–307 (2000). doi:10.1080/09524622.2000.9753439
Sanvito, S., Galimberti, F.: Source level of male vocalisations in the genus Mirounga: repeatability and correlates. Bioacoustics 14(1), 47–59 (2003). doi:10.1080/09524622.2003.9753512
Sanvito, S., Galimberti, F., Miller, E.H.: Development of aggressive vocalizations in male southern elephant seals (Mirounga leonina): maturation or learning? Behaviour 145(2), 137–170 (2008). doi:10.1163/156853907783244729
Blix, A.S., Nordøy, E.S.: Ross seal (Ommatophoca rossii) annual distribution, diving behaviour, breeding and moulting, off Queen Maud Land, Antarctica. Polar Biol. 30(11), 1449–1458 (2007). doi:10.1007/s00300-007-0306-y
Ray, C.: Population ecology of Antarctic seals. In: Holdgate, M.W. (ed.) Antarctic Ecology, vol. 1, pp. 398–414. Academic Press, London (1970)
Erbe, C., Ainslie, M.A., de Jong, C.A.F., Racca, R., Stocker, M.: The need for protocols and standards in research on underwater noise impacts on marine life. In: Popper, A.N., Hawkins, A. (eds.) The Effects of Noise on Aquatic Life II. Advances in Experimental Medicine and Biology, vol. 875, pp. 1265–1271. Springer, New York (2016)
Dunlop, R.A.: Changes in vocal parameters with social context in humpback whales: considering the effect of bystanders. Behav. Ecol. Sociobiol. 70(6), 857–870 (2016). doi:10.1007/s00265-016-2108-0
Au, W.W.L., Lammers, M.O., Yin, S.: Acoustics of dusky dolphins (Lagenorhynchus obscurus). In: Würsig, B., Würsig, M. (eds.) The dusky dolphin: Master acrobats off different shores. Elsevier, Amsterdam (2010)
Azzolin, M., Papale, E., Lammers, M.O., Gannier, A., Giacoma, C.: Geographic variation of whistles of the striped dolphin (Stenella coeruleoalba) within the Mediterranean Sea. J. Acoust. Soc. Am. 134(1), 694–705 (2013). doi:10.1121/1.4808329
Norris, K.S., Evans, W.E.: Directionality of echolocation clicks in the rough-tooth porpoise, Steno bredanensis (Lesson). In: Tavolga, W.N. (ed.) Marine Bio-Acoustics, vol. 2, pp. 305–316. Pergamon Press, New York (1967)
Au, W., Branstetter, B., Moore, P., Finneran, J.: The biosonar field around an Atlantic bottlenose dolphin (Tursiops truncatus). J. Acoust. Soc. Am. 131(1), 569–576 (2012). doi:10.1121/1.3662077
Marley, S.A., Erbe, C., Salgado Kent, C.P.: Underwater recordings of the whistles of bottlenose dolphins in Fremantle Inner Harbour, Western Australia. Sci. Data 4, 170126 (2017). doi:10.1038/sdata.2017.126
Acknowledgements
The following people have kindly donated recordings for spectrograms: Doug Cato (pygmy right whales), Jason Gedamke (dwarf minke whales), Sarah Marley (snubfin dolphins), Sue Mason (common dolphins), Angela Recalde-Salas (pygmy blue whales and humpback whales), Joshua Smith (humpback dolphins), Joy Tripovich (Australian fur seals, Australian sea lions), Rhianne Ward (southern right whales and Indian Ocean bottlenose dolphins), Rebecca Wellard (killer whales). We also wish to thank Carrie Skorcz for extracting WAV files from CWR recordings. The Australian Government Department of Defence provided logistical assistance and L3 Technologies and Sonartech Atlas provided technical assistance for much of the CWR recordings.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Erbe, C., Dunlop, R., Jenner, K.C.S. et al. Review of Underwater and In-Air Sounds Emitted by Australian and Antarctic Marine Mammals. Acoust Aust 45, 179–241 (2017). https://doi.org/10.1007/s40857-017-0101-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40857-017-0101-z