
Abstract
Purpose of Review
Global warming and pollution are among the five major causes of global biodiversity loss, particularly in aquatic invertebrates which are highly diverse but understudied. In this review, we highlight advancements in current environmental studies investigating the interactive effects between warming and contaminants in freshwater and marine invertebrates. We not only focused on temperate regions but also synthesized information on the less studied Arctic/Antarctic and tropical regions.
Recent Findings
In general, the same combination of warming and contaminants may result in either additive or non-additive interactive effects depending on taxa, the response variable, life stage, genotype, exposure level, duration and order of exposure, and the number of exposed generations. For traditional contaminants such as metals and pesticides, combined effects with warming at the individual level were generally synergistic. Growing evidence suggests that multigenerational exposure can shift the interaction between warming and contaminants toward antagonism, while contemporary evolution may change the interaction type.
Summary
Our synthesis highlights the importance of temporal aspects in shaping interaction type, including order of exposure, ontogenetic effects, transgenerational effects, and evolution. The combination of laboratory experiments (to advance mechanistic understanding) and outdoor mesocosm studies or field observations (to increase realism) is needed to obtain comprehensive assessments of interactive effects of warming and pollutants from genes to ecosystems.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Aquatic invertebrates are among the most biologically diverse animal groups and play a crucial role in transferring energy and resources from algae and plants to higher trophic levels such as fish, birds, and mammals [1, 2]. They can also influence biogeochemical cycles, including carbon sequestration [3]. Globally, over 1.3 million invertebrate species have been identified, but many more have gone extinct before even being recorded [4]. Aquatic invertebrates are highly vulnerable to both warming and contaminants, and generally have a limited dispersal capacity to escape exposures to both stressors [5]. For example, the abundance of many crayfish, dragonflies, and damselflies in New South Wales, Australia, is projected to decline from 20% to over 50% by 2085 under warming due to their limited dispersal capacity [6]. Investigations of the combined effects of warming and contaminants have emerged as a major multidisciplinary research theme, bridging ecotoxicology and climatology, limnology, oceanography, ecology, and evolution [7, 8••].
Global warming and anthropogenic contaminants are among the five dominant threats that place ~1 million species at risk of extinction [9]. During the last 15 years, there have been several influential review articles, particularly emphasizing the synergistic effects of warming and contaminants on different biological levels of organization, from physiology to communities [10,11,12,13,14, 15••]. In this review, we discuss emerging research themes at the interface of global warming and traditional contaminants (metals, pesticides, herbicides, biocides), and emerging contaminants (pharmaceuticals, microplastics, and nanoparticles) on aquatic invertebrates focusing on the period 2017–2022 (see Supplementary material S1). The interaction between warming and contaminants is considered synergistic, additive, or antagonistic when their combined effect is greater, equal, or smaller than the sum of the individual effects on a biological response, respectively [16]. In the next sections, we will discuss how contaminant effects on marine and freshwater invertebrates may depend on (i) different aspects of warming, including elevated temperatures, heat waves, and daily temperature fluctuations; (ii) the number of exposed generations (transgenerational effects); (iii) the evolution of increased tolerance; and (iv) argue the importance of studying the interaction between the two stressors in mesocosm experiments. In each of the sections, we highlight understudied aspects to inspire future studies.
Interactive Effects Between Contaminants and Different Aspects of Warming
Interactive Effects of Contaminants and Elevated Mean Temperatures
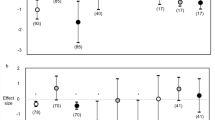
Most experiments tested the interaction of contaminants with a relatively small step-increase in temperature within a range of 10–30 °C; some of these were specifically based on scenarios of the Intergovernmental Panel on Climate Change [17]. These studies are relevant for the reproductive and growing seasons of marine and freshwater invertebrates, particularly in temperate regions. At the individual level, the combined effect of warming and traditional contaminants (pesticides, metals, oil substances) is mainly synergistic within this temperature range (Table 1, Fig. 1, Wald χ2 = 26.94, P < 0.01) and this pattern was not statistically different between marine and freshwater studies (Wald χ2 = 5.06, P = 0.08). Most studies on interactive effects, and all those summarized in Table 1, are laboratory experiments that focused on life history, with behavioural responses understudied. Nevertheless, the latter can be important in linking individual responses to community-level responses. For example, when tested at a higher temperature, the pesticide chlorpyrifos reduced food intake of the predatory larvae of the damselfly Ischnura elegans more, while negative effects of the pesticide on their antipredator behaviour were not temperature-dependent [18]. This suggests the overall pesticide-induced changes in interactions with lower and higher trophic levels can be differentially temperature-dependent [19].

Summary of the proportion of each type of interactive effects between warming and contaminants on marine (A) and freshwater (B) invertebrates, and (C) schematic overview how the combined effects of warming and contaminants depend on the aspects of warming and contaminant type, the intensity of stressors being manipulated, and the level of biological organization studied
Studies listed in Table 1 mainly focused on traditional contaminants. In contrast, information on the combined effects of warming and emerging contaminants of global concern, such as microplastics [20, 21] remains limited to a handful of studies on a few aquatic invertebrates (e.g., bivalves, cladocerans, and corals, Table 2 [22,23,24]). In general, these studies found that warming had a much greater effects on the life history and physiological traits than microplastics, whereby the total effect of the two stressors was mostly equal to the effect of warming alone [22, 23]. Nevertheless, synergistic effects have been observed in the coral Pocillopora verrucosa, with microplastics increasing the susceptibility to bleaching under warming, possibly due to the need to spend more energy on coping with the microplastics [23]. In addition, microplastics can also be a carrier of various contaminants, and therefore, studies on how microplastics interact with these and warming are urgently needed.
Very few experiments have tested the toxicity of contaminants on aquatic invertebrates of the thermally more extreme regions of our planet: Arctic [25] and Antarctic [26•], and tropics [27, 28]. Polar invertebrates are typically stenothermic with a very slow development and long generation time, and limited plasticity. Just a few degrees increase in water temperature can take temperatures beyond the optimal range of these species. For example, the Arctic marine copepods Calanus glacialis and C. hyperboreus have a peak distribution at a seawater temperature of 1–2 °C and the upper range of the thermal niche for both species is around 7 °C [29]. Similar thermal adaptations to low temperatures are found in Antarctic invertebrates, e.g. the upper critical thermal limit is ~2 °C for the Weddell Sea bivalve Limopsis marionensis and ~4 °C for the brachiopod Liothyrella uva [30]. Most Antarctic marine invertebrates cannot survive after a short exposure to temperatures of 5–10 °C (reviewed in [30]) that can be linked to the limited response to thermal stress. For example, the Arctic copepod C. glacialis shows no upregulation of genes encoding heat shock proteins when exposed to temperatures from 0 to 15 °C [31].
Only few exposure studies have tested the interaction of warming and contaminants on polar marine invertebrates at relevant temperatures [25, 26•]. These studies found weak interactive effects which depended on the response variable, species, warming level, and the exposure concentration (Table 1). For instance, the lethal effect of the polycyclic aromatic hydrocarbon pyrene on C. glacialis was only evident at 0 °C, but not at higher temperature (5 °C and 10 °C) [32]. This result suggests that high thermal stress overshadowed the pyrene effects as indicated by a high mortality in the pyrene-free treatment at 10 °C [32]. Pyrene-exposed C. finmarchicus showed stronger decreases in egg production at both lower (0.5 °C) and higher (8 °C) temperatures than at an intermediate temperature (5 °C) [25]. The interactive effects of warming and trace metals on the survival of polar invertebrates are more consistent and generally synergistic (Table 1). For example, the lethal toxicity of copper on the sub-Antarctic marine copepod Harpacticus sp., the isopod Limnoria stephenseni, and the bivalve Gaimardia trapesina increased when the temperature increased from 6 to 8 °C or 10 °C [26•].
In tropical ecosystems, aquatic invertebrates are hyperdiverse [33] and are highly sensitive to contaminants, particularly when the temperature is above 25 °C [34]. Furthermore, tropical aquatic invertebrates occur close to their upper thermal limit and are therefore highly susceptible to warming [35,36,37,38]. A small increase in the water temperature may result in great ecological consequences [35] and this is expected to be more severe in contaminated environments. For example, the tropical copepod Pseudodiaptomus annandalei lives in coastal areas with a seawater temperature of 24–30 °C; an increase of 4 °C in temperature may severely impair its growth, development, and reproduction [39]. Exposure to copper (26.5 μg/L) reduced the survival of P. annandalei males and females with 32% and 62% at 30 °C, and considerably more so at 34 °C (74% and 95%, respectively) [27]. At the sublethal level, Cu-exposed females produced eight times fewer nauplii at 34 °C than at 30 °C [27].
Another study on a tropical species highlighted how exposure duration may result in different single and additive effects of warming and the metal copper on the physiology of the reef-building coral Mussismilia harttii and its photosynthetic symbionts [40]. There was no change in either corals or symbionts in cellular damage (e.g., lipid peroxidation) or total antioxidant capacity after 4 days of exposure to thermal stress. After 12 days of exposure, however, the symbiont had reduced lipid peroxidation under thermal stress, and increased total antioxidant capacity under both stressors [40]. The coral showed an increase in cellular damage under thermal stress or copper exposure, but there was no interaction between the two stressors [40].
Interactive Effects of Contaminants and Heat Extremes
Heat waves (duration ≥ 5 days) and heat spikes (duration < 5 days) can be defined as discrete periods of abnormally high temperatures above the 90th percentile of temperatures recorded in a 30 year timespan at a given locality [41]. Invertebrates typically respond negatively to heat waves [42], including mass coral bleaching (e.g., along ~2,300 km of the Great Barrier Reef in 2017 [43]) and mass mortality of freshwater and coastal marine invertebrates (e.g. Cnidaria, Porifera, Bryozoa, Bivalvia, Annelida, Chlorophyta, Echinodermata in the Mediterranean Sea [44]). Episodes of heat waves occur at a higher frequency and have a longer duration and greater intensity under global warming, particularly in Arctic and tropical ecosystems [45].
The few studies on combined effects of heat waves/spikes and contaminants showed interactive effects with pesticides in temperate freshwater invertebrates [46, 47, 48••], and with metals and oil substances in tropical marine copepods [49, 50]. Life history traits of contaminant-exposed invertebrates are generally severely impacted under heat waves, which can be linked to metabolic depression, and an associated reduced protein synthesis, and to increased oxidative damage [46]. While a heat wave may be lethal in itself [46], contaminant exposure may reduce heat tolerance [51], further increasing the mortality risk under heat stress [46]. Synergistic effects between heat waves/spikes and contaminants have indeed been reported [46], yet a study on water fleas revealed that within a set of six Daphnia magna clones of a single population, the interaction type could vary between a synergism to an antagonism, whereby the more pesticide-tolerant clones showed a stronger synergism [47].
In marine ecosystems, heat waves occur in 25% of the world oceans [52], yet the investigations on the interactive effects of heat waves and contaminants on marine species are very limited. In the tropical marine copepod Centropages velificatus, effects of a short-term exposure (5 days) to pyrene and heat waves on survival, reproduction, and grazing were synergistic, antagonistic, or additive depending on the life history trait measured and the intensity of the stressors [50]. Indeed, at the control temperature (24.5 °C), pyrene exposure only reduced the survival at a concentration of ~300 nM, but under a simulated heat wave, the lethal effect of pyrene already occurred at a concentration of 100 nM, indicating a synergistic effect [50]. There was also a synergistic effect for fecal pellet production, whereby exposure to the heat wave or to 100 nM pyrene alone did not reduce this variable, while it was reduced by 86% when the two stressors were combined [50]. In contrast, the highest pyrene exposure concentration (~300 nM) also caused a strong reduction in pellet production, which was not reduced further under a heat wave, suggesting an antagonistic effect and highlighting the role of pyrene as a dominant stressor in this system [53] and the importance of stressor magnitude in determining interactive effects.
No studies tested for the combined effect of heat waves or heat spikes and emerging contaminants on marine and freshwater invertebrates.
Interactive Effects of Contaminants and Daily Temperature Fluctuations
One limitation in current studies on the interactive effects of warming and contaminants is that experiments have been conducted under constantly elevated temperatures. In nature, however, ambient temperatures fluctuate on a daily basis, which may be more challenging for ectotherms compared to exposure to increased average temperatures [54]. Exposure to daily temperature fluctuations (DTF) is energetically costly [55] and may therefore shape how organisms deal with contaminants [56]. In the majority of studies, daily temperature fluctuations and contaminants reinforced each other’s effects, resulting in synergisms, which has been mostly observed in terms of reduced survival rates (Tables 1, 2). For example, larval survival of C. pipiens mosquitoes under chlorpyrifos exposure was ~15% lower in the presence of 7 and 14 °C DTF than at the constant temperature of 20 °C [57]. A similar pattern has been observed for I. elegans damselflies, where chlorpyrifos exposure decreased larval survival (−25%) and growth rate (−100%), yet only under 5 and 10 °C DTF and not at a constant temperature of 20 °C [58]. Notably, the synergistic effect between warming and a contaminant was stronger under the more realistic global warming scenario in which both higher average temperatures and higher daily temperature fluctuations are combined [58], yet antagonistic interactions between pesticides and daily temperature fluctuations have also been observed. For example, while chlorpyrifos exposure reduced larval survival of C. pipiens mosquitoes with ~40% at a constant temperature of 20 °C, chlorpyrifos did not affect larval survival under 10 °C DTF, which was explained by a faster chlorpyrifos degradation rate under DTF [59]. Although physiological responses are important for mechanistic insights to explain observed patterns in life history traits, they are understudied and mostly show no interaction patterns [56] (Table 1), yet few studies have found that daily temperature fluctuations increased the impact of chlorpyrifos at the physiological level, in terms of increased levels of oxidative damage to lipids [60] and reduced energy budget levels [61].
Only two studies tested the effects of emerging contaminants in the presence of DTF. A study on reef building corals found additive effects of microplastics and DTF [23], whereby the effects of realistic microplastics concentrations were weakly positive while those of DTF were strongly negative [23]. For the freshwater invertebrate D. magna, exposure to microplastics increased fecundity and intrinsic growth rate, but only under 5 °C DTF and not under a constant optimal temperature, likely because of an associated increase in food ingestion [24].
In summary, there is a need for improving our understanding of how aquatic invertebrates, particularly those occurring in extreme thermal environments such as polar and tropical regions cope with contaminants under warming, and especially under realistic daily temperature fluctuations and heatwaves. Investigations considering different stress levels, exposure durations, and temporal aspects of stressors on different life stages are particularly needed.
Multigenerational Effects Shaping the Interaction Type Between Contaminants and Warming
Recent studies on aquatic invertebrates revealed that the interaction effects between warming and contaminants may depend on exposure of the parental generation, highlighting the role of transgenerational effects in shaping the susceptibility of aquatic invertebrates to human-induced rapid environmental change [62]. Parental exposure to warming or contaminants can have two contrasting outcomes that may eventually shape their interaction type. Parental exposure can, through acclimation, alleviate in the offspring the negative effects of single stressors such as warming [63], and contaminants [64]. This was shown in the mosquito C. pipiens, the chlorpyrifos-induced lethal and sublethal effects in the offspring were smaller when parents had also been exposed to chlorpyrifos [64]. Similarly, when the parental generation of the mosquito C. pipiens was exposed to warming, warming no longer reduced diving times in the offspring [63], yet generally, our review indicates that parental exposure to warming or a contaminant increased the vulnerability of offspring to warming [65], and to contaminants such as metals (e.g. copper [49], and lead [66•]) and pesticides (e.g. chlorpyrifos [65]) and this pattern was observed across marine and freshwater invertebrates. This can be explained by poor maternal provisioning and/or the transfer of contaminants or metabolites from mothers to offspring [67].
Such transgenerational shaping of the offspring’s tolerance to single stressors may eventually affect the interaction type in the offspring. In freshwater invertebrates, a series of studies on the mosquito C. pipiens demonstrated evidence for this but at the same time showed it to be case-specific. Two studies showed that the increased sensitivity to the pesticide chlorpyrifos under warming was less pronounced when also the parental generation had been exposed to warming [64, 68]. This could be explained by the observation that only in the latter case warming caused no reduction in the net energy budget [64]. Furthermore, the synergism between warming and the pesticide chlorpyrifos shifted toward an additive effect in offspring whose parents were exposed to both warming and the pesticide because in this condition the pesticide was already more lethal at the lower temperature [65]. In the same species, the interaction type between warming and the pesticide chlorpyifos changed from an antagonism (caused by increased degradation of the pesticide) when parents were not exposed to warming toward an additive pattern when parents experienced warming [63], yet some studies found no effect of the parental exposure history on the interactions between warming and contaminants in the offspring. For example, a heat spike enhanced the toxicity of chlorpyrifos on the survival of C. pipiens in both parental and offspring generations, and the magnitude of the interactive effects remained the same in the offspring [64].
A few studies went a step further and tested for interactive effects between warming and a contaminants after many generations of exposure. These experiments have been limited to species with relatively short generation times, mainly marine copepods and water fleas, mosquitoes, and rotifers. These studies so far indicate that such multigenerational exposure may strengthen as well as weaken the interaction type. In the tropical marine copepods Pseudodiaptomus annandalei and P. incisus the interaction between copper and a heat wave was synergistic, additive, or antagonistic depending on the life history trait and the number of exposed generations [27, 49]. Heat waves typically magnified the lethal and sublethal effects of copper in the parental generation [27, 49]. Interestingly, the interaction between copper and a heat wave on the clutch size, the nauplii and fecal pellet production was synergistic in the parental generation, but became additive in the offspring generations [49]. In the freshwater water flea Moina dubia, the interaction effect between warming and the metal lead was antagonistic, and this antagonistic effect increased gradually across 10 exposed generations and levelled off in the 11th exposed generation [66•].
In summary, transgenerational effects of warming and contaminants tend to shift the interaction type from synergistic in the parental generations toward antagonistic in the offspring generation(s), likely due to the poor maternal provisioning of exposed parents.
Evolution Shaping the Interaction Type Between Contaminants and Warming
While transgenerational effects are reflecting plastic changes, also, evolutionary (genetic) changes may shape the interaction type between contaminants and warming in aquatic invertebrates. Recent studies have shown that aquatic invertebrates can rapidly evolve adaptations to warming (e.g., [69, 70, 71••]) and contaminants (e.g., metal copper [72]). Such evolution of tolerance to a single stressor may eventually shape the interaction type with a second stressor. In marine ecosystems, rapid evolution of the copepod Acartia tonsa to warming and CO2-induced ocean acidification changed the interaction type between both stressors [71••]. Indeed, the synergistic negative effects of warming and ocean acidification on egg production rate and hatching success in the first generation disappeared in generations 3–25 as both traits recovered [71••] caused by rapid genetic adaptation [73]. Furthermore, the combined effects of these stressors on survival and development time shifted from additive (no interaction) toward antagonistic, particularly after 15 generations caused by increased survival in the warming treatment, but not in the selection treatment where both stressors were combined [71••]. The genome-wide analyses showed substantial allelic frequency changes in response to warming (57%), ocean acidification (20%), and their combination (63%) whereby warming was the dominant stressor in driving the interactive effects [74••].
In freshwater ecosystems, two studies in the water flea D. magna showed contrasting effects of how thermal evolution can shape the interaction between warming and zinc oxide nanoparticles. A resurrection study of a natural population showed that the old “ancestral” subpopulation, which were obtained from resting eggs in lake sediments, showed a synergism for intrinsic growth rate, metabolic activity, and energy reserves which disappeared in the recent “derived” subpopulation that evolved a lower sensitivity to the nanoparticles at 24 °C [75]. This indicates that thermal evolution could offset the elevated toxicity of nano-zinc particles under warming. Instead, experimental evolution trials in heated mesocosms showed that nano-zinc particles were more toxic at 20 °C than at 24 °C due to a higher accumulated zinc burden at 20 °C than at 24 °C, and this did not change after thermal evolution [76]. This suggests that the ecological context may critically shape the evolutionary outcome of the stressor interaction type. In an experimental evolution trial with the rotifer Brachionus calyciflorus, animals evolved to both single and combined exposure to contaminants (salt and copper) and a lower temperature [77]. Here, rapid evolution to contaminants and a lower temperature shifted their interactive effect toward a synergism [78••].
From the limited number of evolutionary experiments, no pattern seems to emerge as rapid evolution to warming and contaminants may both enhance antagonisms, and enhance or reduce synergisms.
Mesocosm Studies Testing Higher-Level Effects of the Interaction Between Warming and Contaminants
All studies discussed above are indoor laboratory experiments, and while these are highly relevant for providing detailed and mechanistic insights into the interactive effects of warming and contaminants (see e.g., [46, 79]), their limited environmental realism limits extrapolation of their findings to real-world systems. Only few freshwater studies have empirically attempted to link the combined effects of warming and contaminants from the individual to the ecosystem levels in a single study (see [48••, 80, 81]). Such studies are important for demonstrating how individual trophic level studies are predictive of multitrophic community responses, where biotic interactions such as competition, predation, and parasitism can modify how effects manifest throughout the food web [15••]. The costly nature of manipulating water temperature in semi-natural conditions and challenge of controlling treatments that follow the natural fluctuations of the ambient, environmental temperature are reasons for the rarity of these studies. These manipulations of thermal regimes are more achievable in smaller, closed mesocosm systems mimicking pond systems [80,81,82,83,84]. By contrast, achieving these in open stream mesocosm systems (e.g. [48••, 85, 86]) is considerably more challenging due to the constant, yet varying input of heated water required to maintain the desired temperature difference. Measuring and processing samples across multiple trophic levels (as achieved by Van de Perre et al. [80] in indoor laboratory microcosms, see below) are also costly and time-consuming.
Recent meta-analyses have shown that the combined effects of two stressors are more often antagonistic at the community and ecosystem levels [53, 87]. This might be caused by increased functional redundancy or species acclimating to similar stressors so exposure to additional stress has a less-than-additive effect [53]. There have been two published outdoor aquatic mesocosm studies to investigate the combined effects of raised water temperature and contaminants; one each in pond mesocosms [82] and in flow-through circular stream channels [48••]. The pond mesocosm study found no effects of three simulated heat waves on the effect of a single pulse of the insecticide esfenvalerate on the zooplankton community. However, pesticide effects on the common and sensitive taxon, Daphnia spp., lasted twice as long under warming compared to constant ambient temperatures, likely increased interspecific competition under warming prolonged Daphnia spp. recovery. The second study simulated climate warming and reduced flow velocity and pulsed exposure of the neonicotinoid imidacloprid in flow-through stream mesocosms [48••]. Owing in part to a natural heat wave that occurred during the experiment, only antagonistic interactions between imidacloprid exposure and raised water temperature were observed, where the negative effects of imidacloprid on the macroinvertebrate community were the strongest at ambient temperatures. These results resemble those observed in a laboratory microcosm study, where negative effects of chronic imidacloprid exposure only manifested in mayfly nymphs that had had no prior exposure to simulated heat waves [79]. By contrast, short-term laboratory exposures with freshwater macroinvertebrates to imidacloprid at higher concentrations over a wider temperature gradient had previously shown synergism between these two stressors, demonstrating temperature-enhanced toxicity of imidacloprid [19]. Taken together, these findings suggest that the observed interaction type can be heavily dependent on the level (and environmental realism) of each manipulated stressor.
Two further studies have investigated the combined effects of temperature and contaminants at the community level using indoor microcosm experiments [80, 81]. A first study [80] found that interactions between raised water temperature and zinc on freshwater zooplankton, phytoplankton, and protozoan communities were far more prevalent at the community level than at the species population level. Most of these interactions were antagonistic where the effect of zinc (at the highest concentration applied) was smaller at the higher temperature. A second study [81] observed both synergistic and antagonistic interactions between raised water temperatures and the insecticide lufenuron for different zooplankton taxa. Overall, increased temperature did not affect the sensitivity of the community to the insecticide but it did reduce the time to onset of toxic effects (likely due to increased chemical uptake as a result of raised metabolic rates [11]) and subsequent recovery from low-level insecticide exposure (likely because of increased pesticide degradation [88]).
In future research, there is a need for more studies to address the important question of how contaminant and temperature interactions affect whole ecosystems with multiple trophic levels and higher levels of biological organization in environmentally realistic scenarios [15••]. With continually improving methods of manipulating temperature in semi-natural mesocosm studies (e.g. [83]), such studies will likely become more common. Mesocosms are an ideal tool for assessing the impacts of contaminants at the ecosystem-level, in combination with other climate-related stressors that are not easily manipulated in situ [48••, 89]. By replicating the trophic diversity of natural ecosystems, maintaining environmentally realistic physicochemical conditions and biotic interactions, and allowing the manipulation of toxic substances in controlled, yet realistic environments, mesocosm experiments represent a promising opportunity for providing the empirical data needed to improve our understanding in this field.
Summary Remarks
Global warming and heat extremes continue to be among the most pressing threats for global invertebrate biodiversity [90]. In addition, continued and increased use and release of chemical contaminants in the environment poses another threat. Our review highlights that the interaction type between an increase in mean temperature and contaminants is mainly synergistic but also indicates the type and strength of the interaction between both stressors depends on taxa, the response variable, life stage, genotype, exposure level, duration and order of exposure, and the number of exposed generations (Tables 1, 2). The combined effects of warming and contaminants also need to be assessed under more realistic thermal scenarios of temperature fluctuations, particularly in mesocosms to increase the environmental realism. Linking laboratory experiments and mesocosm studies are essential for understanding field observations of the combined effects of the two stressors. This information will be crucial for managing and mitigating the combined risk of global warming and pollutants in natural systems.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance •• Of major importance
Chew LL, Chong VC, Tanaka K, Sasekumar A. Phytoplankton fuel the energy flow from zooplankton to small nekton in turbid mangrove waters. Mar Ecol Prog Ser. 2012;469:7–24. https://doi.org/10.3354/meps09997.
Falk-Petersen S, Mayzaud P, Kattner G, Sargent J. Lipids and life strategy of Arctic Calanus. Mar Biol Res. 2009;5(1):18–39. https://doi.org/10.1080/17451000802512267. Pii 907389972.
Jonasdottir SH, Visser AW, Richardson K, Heath MR. Seasonal copepod lipid pump promotes carbon sequestration in the deep North Atlantic. Proc Natl Acad Sci USA. 2015;112(39):12122–6. https://doi.org/10.1073/pnas.1512110112.
Eisenhauer N, Bonn A, Guerra CA. Recognizing the quiet extinction of invertebrates. Nat Commun. 2019;10(1):50. https://doi.org/10.1038/s41467-018-07916-1.
Dudgeon D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr Biol. 2019;29(19):R960–7. https://doi.org/10.1016/j.cub.2019.08.002.
Bush A, Hoskins AJ. Does dispersal capacity matter for freshwater biodiversity under climate change? Freshwat Biol. 2017;62(2):382–96. https://doi.org/10.1111/fwb.12874.
Stoks R, Debecker S, Dinh KV, Janssens L. Integrating ecology and evolution in aquatic toxicology: insights from damselflies. Freshw Sci. 2015;34(3):1032–9. https://doi.org/10.1086/682571.
•• Orr JA, Vinebrooke RD, Jackson MC, Kroeker KJ, Kordas RL, Mantyka-Pringle C, et al. Towards a unified study of multiple stressors: divisions and common goals across research disciplines. Proc R Soc B Biol Sci. 1926;2020(287):20200421. This review article used quantitative bibliometric analysis to identify the division between disciplines and linked previously disconnected research communities towards a unified research framework which can be applied to all ecological research.
IPBES. Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science - Policy Platform on Biodiversity and Ecosystem Services. In: Brondizio ES, Settele J, Díaz S, Ngo HT, editors. Intergovernmental science-policy platform on biodiversity and ecosystem services. Bonn, Germany: IPBES; 2019. p. 56.
Sokolova IM, Lannig G. Interactive effects of metal pollution and temperature on metabolism in aquatic ectotherms: implications of global climate change. Clim Res. 2008;37(2–3):181–201. https://doi.org/10.3354/cr00764.
Noyes PD, Lema SC. Forecasting the impacts of chemical pollution and climate change interactions on the health of wildlife. Curr Zool. 2015;61(4):669–89. https://doi.org/10.1093/czoolo/61.4.669.
Noyes PD, McElwee MK, Miller HD, Clark BW, Van Tiem LA, Walcott KC, et al. The toxicology of climate change: environmental contaminants in a warming world. Environ Int. 2009;35(6):971–86. https://doi.org/10.1016/j.envint.2009.02.006.
Holmstrup M, Bindesbol A-M, Oostingh GJ, Duschl A, Scheil V, Koehler H-R, et al. Interactions between effects of environmental chemicals and natural stressors: a review. Sci Total Environ. 2010;408(18):3746–62. https://doi.org/10.1016/j.scitotenv.2009.10.067.
Moe SJ, De Schamphelaere K, Clements WH, Sorensen MT, Van den Brink PJ, Liess M. Combined and interactive effects of global climate change and toxicants on populations and communities. Environ Toxicol Chem. 2013;32(1):49–61. https://doi.org/10.1002/etc.2045.
•• Polazzo F, Roth SK, Hermann M, Mangold-Döring A, Rico A, Sobek A, et al. Combined effects of heatwaves and micropollutants on freshwater ecosystems: towards an integrated assessment of extreme events in multiple stressors research. Glob Change Biol. 2022;28(4):1248–67. https://doi.org/10.1111/gcb.15971. This paper highlights the urgency of integrating extreme events into multiple stressors studies.
Côté IM, Darling ES, Brown CJ. Interactions among ecosystem stressors and their importance in conservation. Proc R Soc B Biol Sci. 1824;2016(283):20152592.
IPCC. Technical Summary. In: Climate Change 2021: The Physical Science Basis. Contribution of working group I to the sixth assessment report of the intergovernmental panel on climate change. Cambridge University Press; 2021.
Van Dinh K, Janssens L, Debecker S, Stoks R. Temperature- and latitude-specific individual growth rates shape the vulnerability of damselfly larvae to a widespread pesticide. J Appl Ecol. 2014;51(4):919–28.
Macaulay SJ, Buchwalter DB, Matthaei CD. Water temperature interacts with the insecticide imidacloprid to alter acute lethal and sublethal toxicity to mayfly larvae. N Z J Mar Freshwat Res. 2020;54(1):115–30. https://doi.org/10.1080/00288330.2019.1614961.
Eerkes-Medrano D, Thompson RC, Aldridge DC. Microplastics in freshwater systems: a review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 2015;75:63–82. https://doi.org/10.1016/j.watres.2015.02.012.
Zhou CY, Bi R, Su CH, Liu WH, Wang TY. The emerging issue of microplastics in marine environment: a bibliometric analysis from 2004 to 2020. Mar Pollut Bull. 2022;179:113712. https://doi.org/10.1016/j.marpolbul.2022.113712.
Bertucci JI, Bellas J. Combined effect of microplastics and global warming factors on early growth and development of the sea urchin (Paracentrotus lividus). Sci Total Environ. 2021;782:146888. https://doi.org/10.1016/j.scitotenv.2021.146888.
Reichert J, Tirpitz V, Anand R, Bach K, Knopp J, Schubert P, et al. Interactive effects of microplastic pollution and heat stress on reef-building corals. Environ Pollut. 2021;290:118010. https://doi.org/10.1016/j.envpol.2021.118010.
Chang M, Zhang C, Li M, Dong J, Li C, Liu J, et al. Warming, temperature fluctuations and thermal evolution change the effects of microplastics at an environmentally relevant concentration. Environ Pollut. 2022;292:118363.
Hjorth M, Nielsen TG. Oil exposure in a warmer Arctic: potential impacts on key zooplankton species. Mar Biol. 2011;158(6):1339–47. https://doi.org/10.1007/s00227-011-1653-3.
• Holan JR, King CK, Proctor AH, Davis AR. Increased sensitivity of subantarctic marine invertebrates to copper under a changing climate - effects of salinity and temperature. Environ Pollut. 2019;249:54–62. https://doi.org/10.1016/j.envpol.2019.02.016. This is a rare study investigating the interactive effects of warming and pollutants on subantarctic invertebrates.
Dinh KV, Nguyen TTQ, Vo T-M-C, Bui BT, Dao T-S, Tran DM, et al. Interactive effects of extreme temperature and a widespread coastal metal contaminant reduce the fitness of a common tropical copepod across generations. Mar Pollut Bull. 2020;159:111509. https://doi.org/10.1016/j.marpolbul.2020.111509.
Lima D, Zacchi FL, Mattos JJ, Flores-Nunes F, Gomes CHADM, de Mello ÁCP, et al. Molecular and cellular effects of temperature in oysters Crassostrea brasiliana exposed to phenanthrene. Chemosphere. 2018;209:307–18. https://doi.org/10.1016/j.chemosphere.2018.06.094.
Strand E, Bagøien E, Edwards M, Broms C, Klevjer T. Spatial distributions and seasonality of four Calanus species in the Northeast Atlantic. Prog Oceanogr. 2020;185:102344. https://doi.org/10.1016/j.pocean.2020.102344.
Pörtner HO, Peck L, Somero G. Thermal limits and adaptation in marine Antarctic ectotherms: an integrative view. Philos Trans R Soc Lond Ser B Biol Sci. 2007;362(1488):2233–58. https://doi.org/10.1098/rstb.2006.1947.
Smolina I, Kollias S, Møller EF, Lindeque P, Sundaram AY, Fernandes JM, et al. Contrasting transcriptome response to thermal stress in two key zooplankton species, Calanus finmarchicus and C. glacialis. Mar Ecol Prog Ser. 2015;534:79–93.
Grenvald JC, Nielsen TG, Hjorth M. Effects of pyrene exposure and temperature on early development of two co-existing Arctic copepods. Ecotoxicology. 2013;22(1):184–98. https://doi.org/10.1007/s10646-012-1016-y.
Barlow J, Franca F, Gardner TA, Hicks CC, Lennox GD, Berenguer E, et al. The future of hyperdiverse tropical ecosystems. Nature. 2018;559(7715):517–26. https://doi.org/10.1038/s41586-018-0301-1.
Heuschele J, Lode T, Konestabo HS, Titelman J, Andersen T, Borgå K. Drivers of copper sensitivity in copepods: a meta-analysis of LC50s. Ecotoxicol Environ Saf. 2022;242:113907. https://doi.org/10.1016/j.ecoenv.2022.113907.
Pinsky ML, Eikeset AM, McCauley DJ, Payne JL, Sunday JM. Greater vulnerability to warming of marine versus terrestrial ectotherms. Nature. 2019;569(7754):108–11. https://doi.org/10.1038/s41586-019-1132-4.
Sasaki MC, Dam HG. Integrating patterns of thermal tolerance and phenotypic plasticity with population genetics to improve understanding of vulnerability to warming in a widespread copepod. Glob Change Biol. 2019;25(12):4147–64. https://doi.org/10.1111/gcb.14811.
Sasaki M, Hedberg S, Richardson K, Dam HG. Complex interactions between local adaptation, phenotypic plasticity and sex affect vulnerability to warming in a widespread marine copepod. R Soc Open Sci. 2019;6(3):182115. https://doi.org/10.1098/rsos.182115.
Sasaki M, Dam HG. Global patterns in copepod thermal tolerance. J Plankton Res. 2021;43(4):598–609. https://doi.org/10.1093/plankt/fbab044.
Doan XN, Vu MTT, Pham HQ, Wisz MS, Nielsen TG, Dinh KV. Extreme temperature impairs growth and productivity in a common tropical marine copepod. Sci Rep. 2019;9:4550. https://doi.org/10.1038/s41598-019-40996-7.
da Silva FJ, Mies M, Paranhos A, Taniguchi S, Güth AZ, Bícego MC, et al. Isolated and combined effects of thermal stress and copper exposure on the trophic behavior and oxidative status of the reef-building coral Mussismilia harttii. Environ Pollut. 2021;268:115892. https://doi.org/10.1016/j.envpol.2020.115892.
Hobday AJ, Alexander LV, Perkins SE, Smale DA, Straub SC, Oliver ECJ, et al. A hierarchical approach to defining marine heatwaves. Prog Oceanogr. 2016;141:227–38. https://doi.org/10.1016/j.pocean.2015.12.014.
Smale DA, Wernberg T, Oliver ECJ, Thomsen M, Harvey BP, Straub SC, et al. Marine heatwaves threaten global biodiversity and the provision of ecosystem services. Nat Clim Chang. 2019;9:306–12. https://doi.org/10.1038/s41558-019-0412-1.
Hughes TP, Kerry JT, Connolly SR, Baird AH, Eakin CM, Heron SF, et al. Ecological memory modifies the cumulative impact of recurrent climate extremes. Nat Clim Chang. 2019;9(1):40–3. https://doi.org/10.1038/s41558-018-0351-2.
Garrabou J, Gómez-Gras D, Medrano A, Cerrano C, Ponti M, Schlegel R, et al. Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Glob Change Biol. 2022. https://doi.org/10.1111/gcb.16301.
Frölicher TL, Fischer EM, Gruber N. Marine heatwaves under global warming. Nature. 2018;560(7718):360–4. https://doi.org/10.1038/s41586-018-0383-9.
Dinh KV, Janssens L, Stoks R. Exposure to a heat wave under food limitation makes an agricultural insecticide lethal: a mechanistic laboratory experiment. Glob Change Biol. 2016;22(10):3361–72. https://doi.org/10.1111/gcb.13415.
Delnat V, Verheyen J, Van Hileghem I, Stoks R. Genetic variation of the interaction type between two stressors in a single population: from antagonism to synergism when combining a heat spike and a pesticide. Environ Pollut. 2022;308:119654. https://doi.org/10.1016/j.envpol.2022.119654.
•• Macaulay SJ, Hageman KJ, Piggott JJ, Juvigny-Khenafou NPD, Matthaei CD. Warming and imidacloprid pulses determine macroinvertebrate community dynamics in experimental streams. Glob Change Biol. 2021;27(21):5469–90. https://doi.org/10.1111/gcb.15856. This is an excellent paper investigating the interactive effects of heatwaves and pesticides on stream macroinvertebrate community.
Dinh KV, Doan KLU, Doan NX, Pham HQ, Le THO, Le M-H, et al. Parental exposures increase the vulnerability of copepod offspring to copper and a simulated marine heatwave. Environ Pollut. 2021;287:117603. https://doi.org/10.1016/j.envpol.2021.117603.
Ruiz LH, Ekumah B, Asiedu DA, Albani G, Acheampong E, Jonasdottir SH, et al. Climate change and oil pollution: a dangerous cocktail for tropical zooplankton. Aquat Toxicol. 2021;231:105718. https://doi.org/10.1016/j.aquatox.2020.105718.
Op de Beeck L, Verheyen J, Stoks R. Integrating both interaction pathways between warming and pesticide exposure on upper thermal tolerance in high- and low-latitude populations of an aquatic insect. Environ Pollut. 2017;224:714–21. https://doi.org/10.1016/j.envpol.2016.11.014.
Oliver ECJ, Benthuysen JA, Bindoff NL, Hobday AJ, Holbrook NJ, Mundy CN, et al. The unprecedented 2015/16 Tasman Sea marine heatwave. Nat Commun. 2017;8(1):16101. https://doi.org/10.1038/ncomms16101.
Jackson MC, Loewen CJG, Vinebrooke RD, Chimimba CT. Net effects of multiple stressors in freshwater ecosystems: a meta-analysis. Glob Change Biol. 2016;22(1):180–9. https://doi.org/10.1111/gcb.13028.
Vasseur DA, DeLong JP, Gilbert B, Greig HS, Harley CDG, McCann KS, et al. Increased temperature variation poses a greater risk to species than climate warming. Proc R Soc B Biol Sci. 2014;281(1779):20132612. https://doi.org/10.1098/rspb.2013.2612.
Colinet H, Sinclair BJ, Vernon P, Renault D. Insects in fluctuating thermal environments. Annu Rev Entomol. 2015;60:123.
Verheyen J, Delnat V, Theys C. Daily temperature fluctuations can magnify the toxicity of pesticides. Curr Opin Insect Sci. 2022;51:100919. https://doi.org/10.1016/j.cois.2022.100919.
Delnat V, Tran TT, Verheyen J, Dinh KV, Janssens L, Stoks R. Temperature variation magnifies chlorpyrifos toxicity differently between larval and adult mosquitoes. Sci Total Environ. 2019;690:1237–44. https://doi.org/10.1016/j.scitotenv.2019.07.030.
Verheyen J, Stoks R. Current and future daily temperature fluctuations make a pesticide more toxic: contrasting effects on life history and physiology. Environ Pollut. 2019;248:209–18. https://doi.org/10.1016/j.envpol.2019.02.022.
Delnat V, Verborgt J, Janssens L, Stoks R. Daily temperature variation lowers the lethal and sublethal impact of a pesticide pulse due to a higher degradation rate. Chemosphere. 2021;263:128114. https://doi.org/10.1016/j.chemosphere.2020.128114.
Verheyen J, Stoks R. Shrinking body size and physiology contribute to geographic variation and the higher toxicity of pesticides in a warming world. Environ Sci Technol. 2019;53(19):11515–23. https://doi.org/10.1021/acs.est.9b03806.
Verheyen J, Stoks R. Negative bioenergetic responses to pesticides in damselfly larvae are more likely when it is hotter and when temperatures fluctuate. Chemosphere. 2020;243:125369. https://doi.org/10.1016/j.chemosphere.2019.125369.
Donelan SC, Hellmann JK, Bell AM, Luttbeg B, Orrock JL, Sheriff MJ, et al. Transgenerational plasticity in human-altered environments. Trends Ecol Evol. 2020;35(2):115–24. https://doi.org/10.1016/j.tree.2019.09.003.
Tran TT, Janssens L, Dinh KV, Stoks R. An adaptive transgenerational effect of warming but not of pesticide exposure determines how a pesticide and warming interact for antipredator behaviour. Environ Pollut. 2019;245:307–15. https://doi.org/10.1016/j.envpol.2018.11.022.
Meng S, Delnat V, Stoks R. Multigenerational effects modify the tolerance of mosquito larvae to chlorpyrifos but not to a heat spike and do not change their synergism. Environ Pollut. 2022;292:118333. https://doi.org/10.1016/j.envpol.2021.118333.
Tran TT, Janssens L, Dinh KV, Stoks R. Transgenerational interactions between pesticide exposure and warming in a vector mosquito. Evol Appl. 2018;11(6):906–17. https://doi.org/10.1111/eva.12605.
• Pham HT, Dinh KV, Nguyen CC, Quoc LB. Changes in the magnitude of the individual and combined effects of contaminants, warming, and predators on tropical cladocerans across 11 generations. Environ Sci Technol. 2020;54(23):15287–95. https://doi.org/10.1021/acs.est.0c05366. This paper highlights the antagonistic effects of warming and pollutants on freshwater invertebrates across multiple exposed generations.
Wang MH, Zhang C, Lee JS. Quantitative shotgun proteomics associates molecular-level cadmium toxicity responses with compromised growth and reproduction in a marine copepod under multigenerational exposure. Environ Sci Technol. 2018;52(3):1612–23. https://doi.org/10.1021/acs.est.8b00149.
Meng S, Tran TT, Delnat V, Stoks R. Transgenerational exposure to warming reduces the sensitivity to a pesticide under warming. Environ Pollut. 2021;284:117217. https://doi.org/10.1016/j.envpol.2021.117217.
Geerts AN, Vanoverbeke J, Vanschoenwinkel B, Van Doorslaer W, Feuchtmayr H, Atkinson D, et al. Rapid evolution of thermal tolerance in the water flea Daphnia. Nat Clim Chang. 2015;5(7):665–8. https://doi.org/10.1038/nclimate2628.
Sasaki MC, Dam HG. Negative relationship between thermal tolerance and plasticity in tolerance emerges during experimental evolution in a widespread marine invertebrate. Evol Appl. 2021;14(8):2114–23. https://doi.org/10.1111/eva.13270.
•• Dam HG, deMayo JA, Park G, Norton L, He X, Finiguerra MB, et al. Rapid, but limited, zooplankton adaptation to simultaneous warming and acidification. Nat Clim Chang. 2021;11(9):780–6. https://doi.org/10.1038/s41558-021-01131-5. This paper highlights the role of rapid evolution in shaping the interactive effects of warming and CO2-induced acidic water on a marine copepod.
Dinh KV, Dinh HT, Pham HT, Selck H, Truong KN. Development of metal adaptation in a tropical marine zooplankton. Sci Rep. 2020;10(1):10212. https://doi.org/10.1038/s41598-020-67096-1.
Brennan RS, deMayo JA, Dam HG, Finiguerra MB, Baumann H, Pespeni MH. Loss of transcriptional plasticity but sustained adaptive capacity after adaptation to global change conditions in a marine copepod. Nat Commun. 2022;13(1):1147. https://doi.org/10.1038/s41467-022-28742-6.
•• Brennan RS, deMayo JA, Dam HG, Finiguerra M, Baumann H, Buffalo V, et al. Experimental evolution reveals the synergistic genomic mechanisms of adaptation to ocean warming and acidification in a marine copepod. Proc Natl Acad Sci USA. 2022;119(38):e2201521119. https://doi.org/10.1073/pnas.2201521119. This paper explored the genetic mechanisms underlying the rapid evolution of a marine copepod to warming and CO2-induced acidic ocean.
Zhang C, Jansen M, De Meester L, Stoks R. Thermal evolution offsets the elevated toxicity of a contaminant under warming: a resurrection study in Daphnia magna. Evol Appl. 2018;11(8):1425–36. https://doi.org/10.1111/eva.12637.
Zhang C, Jansen M, De Meester L, Stoks R. Rapid evolution in response to warming does not affect the toxicity of a pollutant: Insights from experimental evolution in heated mesocosms. Evol Appl. 2019;12(5):977–88. https://doi.org/10.1111/eva.12772.
Luijckx P, Ho EKH, Gasim M, Chen S, Stanic A, Yanchus C, et al. Higher rates of sex evolve during adaptation to more complex environments. Proc Natl Acad Sci USA. 2017;114(3):534–9. https://doi.org/10.1073/pnas.1604072114.
•• Orr JA, Luijckx P, Arnoldi J-F, Jackson AL, Piggott JJ. Rapid evolution generates synergism between multiple stressors: linking theory and an evolution experiment. Glob Change Biol. 2022;28(5):1740–52. https://doi.org/10.1111/gcb.15633. This paper highlights the role of rapid evolution in shaping the interactive effects of warming and pollutant on freshwater invertebrates.
Macaulay SJ, Hageman KJ, Piggott JJ, Matthaei CD. Time-cumulative effects of neonicotinoid exposure, heatwaves and food limitation on stream mayfly nymphs: a multiple-stressor experiment. Sci Total Environ. 2021;754:141941. https://doi.org/10.1016/j.scitotenv.2020.141941.
Van de Perre D, Roessink I, Janssen CR, Smolders E, De Laender F, Van den Brink PJ, et al. The combined and interactive effects of zinc, temperature, and phosphorus on the structure and functioning of a freshwater community. Environ Toxicol Chem. 2018;37(9):2413–27. https://doi.org/10.1002/etc.4201.
Arenas-Sánchez A, López-Heras I, Nozal L, Vighi M, Rico A. Effects of increased temperature, drought, and an insecticide on freshwater zooplankton communities. Environ Toxicol Chem. 2019;38(2):396–411. https://doi.org/10.1002/etc.4304.
Knillmann S, Stampfli NC, Noskov YA, Beketov MA, Liess M. Elevated temperature prolongs long-term effects of a pesticide on Daphnia spp. due to altered competition in zooplankton communities. Glob Change Biol. 2013;19(5):1598–609. https://doi.org/10.1111/gcb.12151.
Hermann M, Jansen R, van de Glind J, Peeters ETHM, Van den Brink PJ. A transportable temperature and heatwave control device (TENTACLE) for laboratory and field simulations of different climate change scenarios in aquatic micro- and mesocosms. HardwareX. 2022;11:e00307. https://doi.org/10.1016/j.ohx.2022.e00307.
Tüzün N, Op de Beeck L, Oliarinony R, Van Dievel M, Stoks R. Warming under seminatural outdoor conditions in the larval stage negatively affects insect flight performance. Biol Lett. 2018;14(5):20180121. https://doi.org/10.1098/rsbl.2018.0121.
Piggott JJ, Salis RK, Lear G, Townsend CR, Matthaei CD. Climate warming and agricultural stressors interact to determine stream periphyton community composition. Glob Change Biol. 2015;21(1):206–22. https://doi.org/10.1111/gcb.12661.
Piggott JJ, Lange K, Townsend CR, Matthaei CD. Multiple stressors in agricultural streams: a mesocosm study of interactions among raised water temperature, sediment addition and nutrient enrichment. PLoS ONE. 2012;7(11):e49873. https://doi.org/10.1371/journal.pone.0049873.
Tekin E, Diamant ES, Cruz-Loya M, Enriquez V, Singh N, Savage VM, et al. Using a newly introduced framework to measure ecological stressor interactions. Ecol Lett. 2020;23(9):1391–403. https://doi.org/10.1111/ele.13533.
Op de Beeck L, Verheyen J, Olsen K, Stoks R. Negative effects of pesticides under global warming can be counteracted by a higher degradation rate and thermal adaptation. J Appl Ecol. 2017;54(6):1847–55. https://doi.org/10.1111/1365-2664.12919.
Stewart RIA, Dossena M, Bohan DA, Jeppesen E, Kordas RL, Ledger ME, et al. Chapter two - Mesocosm experiments as a tool for ecological climate-change research. In: Woodward G, O’Gorman EJ, editors. Adv Ecol Res. Academic Press; 2013. p. 71–181.
Masson-Delmotte V, Zhai P, Pirani A, Connors SL, Péan C, Berger S, et al. Climate change 2021: the physical science basis. In: Contribution of working group I to the sixth assessment report of The Intergovernmental Panel on Climate Change, vol. 2339. 2021.
Delnat V, Tran TT, Janssens L, Stoks R. Daily temperature variation magnifies the toxicity of a mixture consisting of a chemical pesticide and a biopesticide in a vector mosquito. Sci Total Environ. 2019;659:33–40. https://doi.org/10.1016/j.scitotenv.2018.12.332.
•• Verheyen J, Delnat V, Stoks R. Increased daily temperature fluctuations overrule the ability of gradual thermal evolution to offset the increased pesticide toxicity under global warming. Environ Sci Technol. 2019;53(8):4600–8. https://doi.org/10.1021/acs.est.8b07166. This paper highlight the role of daily temperature fluctuations in shaping the interactive effects of warming and pollutants on freshwater invertebrates.
Theys C, Verheyen J, Tüzün N, Stoks R. Higher mean and fluctuating temperatures jointly determine the impact of the pesticide chlorpyrifos on the growth rate and leaf consumption of a freshwater isopod. Chemosphere. 2021;273:128528. https://doi.org/10.1016/j.chemosphere.2020.128528.
de Melo MS, Das K, Gismondi E. Inorganic mercury effects on biomarker gene expressions of a freshwater amphipod at two temperatures. Ecotoxicol Environ Saf. 2021;209:111815. https://doi.org/10.1016/j.ecoenv.2020.111815.
Haque MN, Nam S-E, Kim B-M, Kim K, Rhee J-S. Temperature elevation stage-specifically increases metal toxicity through bioconcentration and impairment of antioxidant defense systems in juvenile and adult marine mysids. Comp Biochem Physiol Part C Toxicol Pharmacol. 2020;237:108831. https://doi.org/10.1016/j.cbpc.2020.108831.
Chen Y, Dong W. Predicted near-future oceanic warming enhances mercury toxicity in marine copepods. Bull Environ Contam Toxicol. 2022;108(5):824–9. https://doi.org/10.1007/s00128-021-03385-3.
Boukadida K, Cachot J, Morin B, Clerandeau C, Banni M. Moderate temperature elevation increase susceptibility of early-life stage of the Mediterranean mussel, Mytilus galloprovincialis to metal-induced genotoxicity. Sci Total Environ. 2019;663:351–60. https://doi.org/10.1016/j.scitotenv.2019.01.215.
Rodríguez BM, Bhuiyan MKA, Freitas R, Conradi M. Mission impossible: reach the carrion in a lithium pollution and marine warming scenario. Environ Res. 2021;199:111332. https://doi.org/10.1016/j.envres.2021.111332.
Nardi A, Mincarelli LF, Benedetti M, Fattorini D, d’Errico G, Regoli F. Indirect effects of climate changes on cadmium bioavailability and biological effects in the Mediterranean mussel Mytilus galloprovincialis. Chemosphere. 2017;169:493–502. https://doi.org/10.1016/j.chemosphere.2016.11.093.
Dinh KV, Olsen MW, Altin D, Vismann B, Nielsen TG. Impact of temperature and pyrene exposure on the functional response of males and females of the copepod Calanus finmarchicus. Environ Sci Pollut Res. 2019;26(28):29327–33. https://doi.org/10.1007/s11356-019-06078-x.
Amid C, Olstedt M, Gunnarsson JS, Le Lan H, Tran Thi Minh H, Van den Brink PJ, et al. Additive effects of the herbicide glyphosate and elevated temperature on the branched coral Acropora formosa in Nha Trang, Vietnam. Environ Sci Pollut Res. 2018;25(14):13360–72. https://doi.org/10.1007/s11356-016-8320-7.
Barbosa M, Inocentes N, Soares AMVM, Oliveira M. Synergy effects of fluoxetine and variability in temperature lead to proportionally greater fitness costs in Daphnia: a multigenerational test. Aquat Toxicol. 2017;193:268–75. https://doi.org/10.1016/j.aquatox.2017.10.017.
Koagouw W, Hazell RJ, Ciocan C. Induction of apoptosis in the gonads of Mytilus edulis by metformin and increased temperature, via regulation of HSP70, CASP8, BCL2 and FAS. Mar Pollut Bull. 2021;173:113011. https://doi.org/10.1016/j.marpolbul.2021.113011.
Lai RWS, Yung MMN, Zhou G-J, He YL, Ng AMC, Djurišić AB, et al. Temperature and salinity jointly drive the toxicity of zinc oxide nanoparticles: a challenge to environmental risk assessment under global climate change. Environ Sci Nano. 2020;7(10):2995–3006.
Asadi Dokht Lish R, Johari SA, Sarkheil M, Yu IJ. On how environmental and experimental conditions affect the results of aquatic nanotoxicology on brine shrimp (Artemia salina): A case of silver nanoparticles toxicity. Environ Pollut. 2019;255:113358. https://doi.org/10.1016/j.envpol.2019.113358.
Funding
Open access funding provided by University of Oslo (incl Oslo University Hospital). K.V.D., K.B., and K.H. received grants from The Nansen Legacy (RCN#276730) and Researcher Project for Young Talents (RCN#325334) of the Research Council of Norway. S.J.M. and M.C.J received funding from UK’s Natural Environment Research Council (NE/V001396/1). R.S. was supported by grants from the Fund for Scientific Research Flanders (FWO) and KU Leuven C1 grant C16/17/002.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Biology and Pollution
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dinh, K.V., Konestabo, H.S., Borgå, K. et al. Interactive Effects of Warming and Pollutants on Marine and Freshwater Invertebrates. Curr Pollution Rep 8, 341–359 (2022). https://doi.org/10.1007/s40726-022-00245-4
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40726-022-00245-4