
Abstract
Introduction
Vitamins A and D are able to modulate innate and adaptive immune responses and may therefore influence the development and the course of allergic diseases.
Materials and methods
This article reviews the current evidence for the experimental effects of vitamins A and D in vivo in animal models and on immune cells in vitro, and discusses their translational implication. A systematic literature search over the last 10 years was performed using MEDLINE and PubMed databases.
Results
Deficiencies of vitamin A or vitamin D in mouse models of allergic asthma seem to exacerbate allergic symptoms along with enhanced lung inflammation and Th2 cytokine production. In contrast, supplementation regimes especially with vitamin D were able to attenuate symptoms in therapeutic mouse models. The active metabolites retinoic acid (RA) and 1,25-dihydroxyvitamin D3 (VD3) induced tolerogenic dendritic cells (DCs) and up-regulated T‑regulatory cells in the allergic sensitization phase, which likely contributes to tolerance induction. Additionally, RA and VD3 maintained the stability of eosinophils and mast cells in the effector phase, thereby reducing allergic mediator release. Thus, both active vitamin metabolites RA and VD3 are able to influence allergic immune responses at several immunological sites.
Conclusion
Animal studies predict that vitamin A and D may also be attractive players in the control of allergy in humans. Whether these experimental observations can be translated to the human situation remains open, as results from clinical trials are controversial.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Vitamins and their metabolites regulate tissue growth, differentiation and embryonic development and have an important control function in the immune homeostasis by influencing both innate and adaptive immune responses. Vitamins A and D are distinct from other vitamins as their metabolites, retinoic acid (RA) and 1,25-dihydroxyvitamin D3 (VD3), are synthesized from precursors by different body tissues and they exert their effects on target cells by binding to nuclear hormone receptors.
Vitamin A
is essential for pre- and postnatal development, eyesight and reproduction, but it also has an important role in the maintenance of the immune system [1]. Vitamin A is taken up in form of retinol or retinyl esters from animal-derived food sources (e. g. fish, liver, milk, eggs) or in form of precursors such as carotenoids from plant food (e. g. carrots, red peppers, lettuce). Vitamin A is stored mainly in the liver. In liver cells, retinol is oxidized to retinal by alcohol dehydrogenases and metabolized to its main active metabolite RA [2]. In the target tissue, the two isoforms all-trans-RA and 9‑cis-RA bind to nuclear retinoic acid receptors [3] and/or retinoid X receptors (RXR), regulating over 500 retinoid-responsive genes [2, 4]. Formation of RA is catalysed by enzymes called retinal dehydrogenases (RALDH), which are expressed in epithelial cells and in immune cells such as macrophages and dendritic cells (DCs) [1]. Recent work has revealed that these enzymes are present not only in gut-associated cells but also in the lung, which has prominent extrahepatic stores of vitamin A [3].
Vitamin D
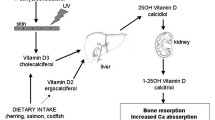
promotes calcium absorption in the gut and helps maintain calcium and phosphate levels to promote bone health [5]. Apart from having neuromuscular and cardiovascular effects it is known to regulate immune cell functions [6]. Vitamin D is acquired via the diet (especially contained in fatty fish) and large amounts are synthesized in the skin from 7‑dehydrocholesterol after exposure to UVB light. After hydroxylation in the liver, the resulting 25-hydroxyvitamin D3 is further hydroxylated in the kidneys by the enzyme 1‑α-hydroxylase resulting in VD3, which is the physiologically active metabolite. VD3 in the target tissue binds to the nuclear vitamin D receptor (VDR), which heterodimerizes with nuclear receptors of the retinoic X receptor family, and binds to VD3 response elements in the promoters of VD3-responsive genes [6]. Both rate-limiting enzymes 1‑α-hydroxylase and the vitamin D receptor are expressed in many tissues, including the lungs, colon, skin, lymph nodes and cells of the immune system. Macrophages, DCs and T and B cells are therefore able to produce VD3 locally, which can act on immune cells in an autocrine or paracrine manner by binding to the vitamin D receptor [7, 8].
The immunomodulatory role of vitamin A/RA and VD3
The immunomodulatory potential of RA and VD3 was investigated since long [9] and recently their general influence on immune cells was reviewed in detail [1, 6]. Here we review reports on the innate and adaptive immune cells which are key players in allergic diseases.
Vitamin A/RA
B cells
The major influence on RA signalling in B cells concerns immunoglobulin class switching [1]. RA from intestinal DCs promotes generation of IgA+ producing B cells and potentiates memory B cell differentiation [1]. Consequently, vitamin A deficiency leads to a severe decrease in intestinal and serum IgA levels [10].
APCs, T cells/Treg cells
On the T cellular level RA influences the homing of both CD4+ and CD8+ T cells by enhancing the expression of the gut-homing molecules α4ß7 and CCR9, promoting preferential migration to the gut-associated lymphoid tissue (GALT) [11]. Most importantly, local production of RA by mucosal DCs together with TGF-ß can induce naive T cells to differentiate into FOXP3+ regulatory T cells (Tregs), thereby maintaining immune tolerance [12]. This phenomenon is not restricted to the gut-associated lymphoid tissue. Alveolar macrophages in the lung were found to promote the induction of FOXP3+ Tregs, which then again mediate respiratory tolerance through the production of RA and TGF-ß [13]. A reciprocal regulatory effect of RA is seen in the suppression of Th17 and Th1 cell differentiation [14]. While under steady-state conditions RA maintains tolerance at mucosal surfaces inducing tolerogenic DCs and Tregs, RA can also induce inflammatory DCs and differentially affect T cell responses in already polarized T cells during infection or autoimmunity [1, 14].
ILCs
In innate lymphocytes RA promotes the generation of ILC3s and antagonizes ILC2 function, highlighting the multiple functions of RA in immune cells [15].
Vitamin D/VD3
APCs
VD3 has a stimulatory effect on monocytes and macrophages, enhancing chemotaxis and activating the cathelicidin antimicrobial peptide [16]. VD3 can also inhibit the differentiation and maturation of DCs, leading to a tolerogenic state with lower antigen presentation but increased production of IL-10 [17]. As DCs express 1‑α-hydroxylase they can acquire high local concentrations of VD3. This notion is supported by studies in VDR/1-α-hydroxylase knockout mice, which show a large increase in mature DCs [17].
B cells
In B cells, which express 1‑α-hydroxylase and VDR upon activation, VD3 has an antiproliferative effect [18]. This effect is mediated by T helper cells but recent studies have shown that VD3 directly inhibits IgG secretion and the generation of memory and plasma cells, and promotes the apoptosis of B cells [8, 19].
T cells/Treg cells
VD3 inhibits the expression of the Th1 (IFN-y, TNF-α), Th9 (IL-9), Th17 (IL-17) and Th22 (IL-22) cytokines in T cells, but has been described to up-regulate Th2 (IL-4, IL-10) cytokines and to induce FOXP3+ regulatory T cells [8, 20]. A recent study on human CD4+ T cells showed a significant increase in the frequency of FOXP3+ regulatory T cells after stimulation with physiologically relevant levels of VD3 in combination with TGF-ß [21]. Interestingly, Th2 cell differentiation can also be inhibited by VD3 [20].
ILCs
In innate lymphoid cells VD3 seems to inhibit ILC2 activation and the expression of gut homing integrin [22].
Taken together, both active vitamin A and D metabolites seem to have a major influence on effector Th1/Th2/Th17 cell differentiation and regulatory T cell induction, which could also have implications for allergic disease development.
Correlation of vitamin A/D deficiency with allergic immune responses in vivo
In recent decades animal models have significantly contributed to the understanding of pathophysiologic mechanisms of allergic diseases such as asthma, anaphylaxis or food allergy [23]. Here we will concentrate on animal models of vitamin deficiency or supplementation that operate with and reflect on vitamin A or vitamin D contents that are taken up via the diet.
In neonate mice vitamin A deficiency (VAD) resulted in impairment of oral tolerance induction [24] with insufficient Treg cell activation by DCs in mesenteric lymph nodes as possible underlying mechanism [25]. Interestingly, feeding of a single dose of vitamin A in adult mice could amplify the tolerogenic properties of mesenteric lymph node DCs [26]. Several studies reported that VAD in adult mice leads to increased airway hyperresponsiveness, lung inflammation and Th2 cytokine production ([27, 28]; Table 1). These mice were also prone to develop skin allergy upon oral antigen administration due to strong, IL-13 dependent IgE response [29]. Earlier observations on VAD found a dose-dependent alteration of asthma symptoms, with low-level VAD decreasing airway inflammation and hyperresponsiveness while high-level dietary vitamin A increased disease severity ([30, 31]; Table 1). The resulting hypothesis that excessive vitamin A intake in industrialized countries induces a Th2 bias and atopy prevalence is still very much under discussion [32].
Vitamin D deficiency in neonate mice seems to contribute to allergic disease severity with high eosinophilia and airway remodelling, which could be improved by vitamin D supplementation ([33]; Table 1). Similarly, supplementation of vitamin D reduced airway hyperresponsiveness and lung inflammation in mouse models of allergic asthma [34, 35]. In this respect, it was reported that eosinophils in vitamin D deficient mice seem to produce higher levels of mediator substances and are prone to spontaneous mediator release [36]. Effects could be demonstrated not only in asthma-related studies, but also in models of food allergy. In this experimental set-up vitamin D deficiency led to exacerbation of symptoms, possibly mediated by increased expression of IL-4 in mesenteric lymph nodes [37].
In allergen-specific immunotherapy (AIT), the only curative treatment for type I allergies, supplementation with 25-hydroxyvitamin D3 (inactive precursor of VD3) was able to enhance the beneficial effects in sensitized, vitamin D deficient mice [38]. This could have major implications for AIT in human allergic patients who often show a prevalence for vitamin D deficiency [39], especially in northern latitudes during wintertime which is exactly the time when pollen-specific AIT is initiated. Thus, a recent study demonstrated enhanced efficacy of sublingual AIT (SLIT) in combination with vitamin D supplementation in grass pollen allergic children [40].
In vivo treatment with RA or VD3 as reference to underlying mechanisms of immunomodulation
There are a number of studies that focus on the in vivo administration of the major vitamin metabolites RA and VD3 and the effects of such treatment on immune cells of the innate as well as adaptive immune system.
Repeated antigen challenge in the presence of 9‑cis-RA reduced specific IgE responses and increased specific IgA responses in mice, probably via B cell derived IL-10 [41]. At the same time, it was reported that the IgE-repressive activity of RA was mediated mainly through the RA receptor alpha (RARα) by down-regulating IgE class-switching recombination [42]. Interestingly, by using lecithin:retinol acetyltransferase-deficient mice that are more susceptible to vitamin A deficiency [43] elevated serum levels of mouse mast cell protease-1 together with significantly reduced numbers of intestinal Treg cells were also found.
DCs at the borderline between the innate and adaptive immune system play a major role in tolerance induction and their phenotypic differentiation can be regulated by vitamin metabolites [44]. In this respect, DCs when differentiated under RA stimulation were able to prevent anaphylactic responses to oral peanut allergen challenge in mice [45]. The RA-generated DCs in this model displayed a tolerogenic mature phenotype expressing IL-10, TGF-ß and IL-27 [45]. Overall, in vivo administration of RA in murine models of allergic rhinitis or asthma was able to attenuate airway inflammation and hyperreactivity due to induction of FOXP3+ regulatory T cells and inhibition of Th2 and Th17 responses ([46, 47]; Table 1).
Concerning VD3, intraperitoneal administration at the time of airway antigen challenge led to reduced airway inflammation and remodeling in murine models of OVA-induced asthma ([48, 49]; Table 1). The protective role of VD3 in the lung of treated mice may be caused by inhibition of NF-kB activation, but also suppression of TGFß/SMAD signalling pathways was reported. Together with activation of the Nrf2/HO-1 pathway VD3 may be able to protect mice from antigen-induced oxidative injury and could be effective in the control of asthma [49].
Another recent report showed that VD3 could prevent the conversion of CD8+ T cells to IL-13 producing pathogenic effector cells in the lungs of antigen challenged mice ([50]; Table 1). The authors argue that vitamin D could therefore be beneficial for asthmatics, but restricted to a subpopulation of steroid-refractory asthmatics with increased numbers of CD8+IL-13+ T cells [50].
In mouse—and human—mast cells, key proinflammatory effector cells of allergic response, VD3 was able to inhibit mediator release in a vitamin D receptor-mediated manner [51]. These data are in line with a recent publication from Liu et al. [52]. They reported that VD3 is important to maintain the stability of mast cell by increasing vitamin D receptor expression together with inhibition of IgE-sensitized mast cell activation. In this respect, VD3 seems to interfere with two major key components of mast cell activation, the adaptor protein Myd88 and the FcεR1ß subunit of the high-affinity IgE receptor [52].
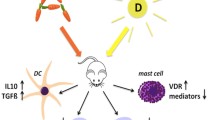
Thus, both active vitamin metabolites RA and VD3 are able to influence allergic immune responses at several immunological sites (Fig. 1).

Vitamin A and vitamin D impact immune cells relevant in the control of allergy. In mouse models of allergic disease vitamin A and D, and their major metabolites retinoic acid (RA) and vitamin D 3 (VD3), down-regulate Th2 and IgE immune responses. Vitamin metabolite-treated dendritic cells (DCs) are prone to adapt a tolerogenic phenotype, leading to up-regulation of T‑regulatory cells (Tregs). Both vitamins also modulate T and B cell responses as well as mediator release from mast cells and eosinophils. (Copyrights from ©motifolio.com, and Fotolia.com©Anastasia Anisenko)
Can data from in vivo animal studies be correlated to the human situation?
The findings from in vivo animal studies suggest that vitamin A and D seem to have a major impact on immune cells as well as on the development, and in the case of vitamin D even on the treatment of allergic Th2-dominated diseases. The question remains whether these findings can be translated to the human situation, especially concerning the dosage of vitamin supplementation in connection with adequate vitamin status [6, 32]. Nonetheless, in some of the studies conducted in mice the doses of vitamins were comparable to those applied in humans [26, 38].
Studies in neonate mice predict that there could exist a “window of opportunity” for prevention of allergic diseases by vitamins A and D. This is reflected in human studies concentrating on the maternal vitamin A or D status that can be of importance for reducing the development of allergic diseases in children [53,54,55]. On the other hand, application of vitamin A as aerosol to asthmatic children was of no benefit [56], and no association between serum carotenoids and risk of asthma in young children could be established in a recently published longitudinal study [57]. Therefore, primary randomized controlled trials of the use of vitamin A to prevent or treat asthma that could elucidate the role of this vitamin in the human allergic disease are still missing.
Earlier studies reported that the vitamin D status in human allergic patients shows a nonlinear relationship with immune parameters relevant for allergic and asthmatic disease (e. g. serum IgE levels) [58]. Nevertheless, vitamin D supplementation trials in asthmatic and/or pollen allergic children gave positive results with regard to asthma exacerbations and improvement of immunotherapy [40, 59], similarly seen in case reports from adult asthmatics [60].
Concluding remarks
The manifold impact of vitamin A and D on immune cell responses and profound evidence from animal studies provide hope that these molecules can help control allergy. More randomized controlled human studies are needed to underpin the potential of these vitamins and their metabolites in prevention or therapy of Th2-dominant allergic diseases including asthma. We propose that more emphasis should be put on the determination of the vitamin status of allergic patients and, especially for vitamin D, on prophylaxis in early life or during pregnancy.
Abbreviations
- AIT:
-
Allergen-specific immunotherapy
- APC:
-
Antigen-presenting cell
- DC:
-
Dendritic cell
- GALT:
-
Gut-associated lymphoid tissue
- HDM:
-
House dust mite
- IL:
-
Interleukin
- ILC:
-
Innate lymphoid cell
- OVA:
-
Ovalbumin
- RA:
-
Retinoic acid
- Treg:
-
T-regulatory cell
- VA:
-
Vitamin A
- VAD:
-
Vitamin A deficiency
- VDR:
-
Vitamin D receptor
- VD3:
-
1,25-dihydroxyvitamin D3
References
Larange A, Cheroutre H. Retinoic acid and retinoic acid receptors as pleiotropic modulators of the immune system. Annu Rev Immunol. 2016;34:369–94.
Li Y, Wongsiriroj N, Blaner WS. The multifaceted nature of retinoid transport and metabolism. Hepatobiliary Surg Nutr. 2014;3:126–39.
Rudraraju R, Jones BG, Surman SL, Sealy RE, Thomas PG, Hurwitz JL. Respiratory tract epithelial cells express retinaldehyde dehydrogenase ALDH1A and enhance IgA production by stimulated B cells in the presence of vitamin A. PLoS ONE. 2014;9:e86554.
Balmer JE, Blomhoff R. Gene expression regulation by retinoic acid. J Lipid Res. 2002;43:1773–808.
Gunton JE, Girgis CM, Baldock PA, Lips P. Bone muscle interactions and vitamin D. Bone. 2015;80:89–94.
Colotta F, Jansson B, Bonelli F. Modulation of inflammatory and immune responses by vitamin D. J Autoimmun. 2017;85:78–97. https://doi.org/10.1016/j.jaut.2017.07.007.
Hansdottir S, Monick MM. Vitamin D effects on lung immunity and respiratory diseases. Vitam Horm. 2011;86:217–37.
Yawn J, Lawrence LA, Carroll WW, Mulligan JK. Vitamin D for the treatment of respiratory diseases: is it the end or just the beginning? J Steroid Biochem Mol Biol. 2015;148:326–37.
Mora JR, Iwata M, von Andrian UH. Vitamin effects on the immune system: vitamins A and D take centre stage. Nat Rev Immunol. 2008;8:685–98.
Maruya M, Suzuki K, Fujimoto H, Miyajima M, Kanagawa O, Wakayama T, et al. Vitamin A‑dependent transcriptional activation of the nuclear factor of activated T cells c1 (NFATc1) is critical for the development and survival of B1 cells. Proc Natl Acad Sci USA. 2011;108:722–7.
Kang SG, Park J, Cho JY, Ulrich B, Kim CH. Complementary roles of retinoic acid and TGF-beta1 in coordinated expression of mucosal integrins by T cells. Mucosal Immunol. 2011;4:66–82.
Liu ZM, Wang KP, Ma J, Guo Zheng S. The role of all-trans retinoic acid in the biology of Foxp3+ regulatory T cells. Cell Mol Immunol. 2015;12:553–7.
Coleman MM, Ruane D, Moran B, Dunne PJ, Keane J, Mills KH. Alveolar macrophages contribute to respiratory tolerance by inducing FoxP3 expression in naive T cells. Am J Respir Cell Mol Biol. 2013;48:773–80.
Raverdeau M, Mills KH. Modulation of T cell and innate immune responses by retinoic acid. J Immunol. 2014;192:2953–8.
Kim MH, Taparowsky EJ, Kim CH. Retinoic acid differentially regulates the migration of innate lymphoid cell subsets to the gut. Immunity. 2015;43:107–19.
Lowry MB, Guo C, Borregaard N, Gombart AF. Regulation of the human cathelicidin antimicrobial peptide gene by 1alpha,25-dihydroxyvitamin D3 in primary immune cells. J Steroid Biochem Mol Biol. 2014;143:183–91.
Bscheider M, Butcher EC. Vitamin D immunoregulation through dendritic cells. Immunology. 2016;148:227–36.
Cantorna MT, Snyder L, Lin YD, Yang L. Vitamin D and 1,25(OH)2D regulation of T cells. Nutrients. 2015;7:3011–21.
Chen S, Sims GP, Chen XX, Gu YY, Chen S, Lipsky PE. Modulatory effects of 1,25-dihydroxyvitamin D3 on human B cell differentiation. J Immunol. 2007;179:1634–47.
Pfeffer PE, Mann EH, Hornsby E, Chambers ES, Chen YH, Rice L, et al. Vitamin D influences asthmatic pathology through its action on diverse immunological pathways. Ann Am Thorac Soc. 2014;11(Suppl 5):S314–S21.
Chambers ES, Suwannasaen D, Mann EH, Urry Z, Richards DF, Lertmemongkolchai G, et al. 1alpha,25-dihydroxyvitamin D3 in combination with transforming growth factor-beta increases the frequency of Foxp3(+) regulatory T cells through preferential expansion and usage of interleukin-2. Immunology. 2014;143:52–60.
Ruiter B, Patil SU, Shreffler WG. Vitamins A and D have antagonistic effects on expression of effector cytokines and gut-homing integrin in human innate lymphoid cells. Clin Exp Allergy. 2015;45:1214–25.
Jensen-Jarolim E, Pali-Scholl I, Roth-Walter F. Outstanding animal studies in allergy I. From asthma to food allergy and anaphylaxis. Curr Opin Allergy Clin Immunol. 2017;17:169–79.
Turfkruyer M, Rekima A, Macchiaverni P, Le Bourhis L, Muncan V, van den Brink GR, et al. Oral tolerance is inefficient in neonatal mice due to a physiological vitamin A deficiency. Mucosal Immunol. 2016;9:479–91.
Nakamoto A, Shuto E, Tsutsumi R, Nakamoto M, Nii Y, Sakai T. Vitamin A deficiency impairs induction of oral tolerance in mice. J Nutr Sci Vitaminol (Tokyo). 2015;61:147–53.
Novotny Nunez I, Barrios BE, Maccio-Maretto L, Correa SG. Migratory capacity and function of dendritic cells from mesenteric afferent lymph nodes after feeding a single dose of vitamin A. J Nutr Biochem. 2017;49:110–6.
Chen F, Marquez H, Kim YK, Qian J, Shao F, Fine A, et al. Prenatal retinoid deficiency leads to airway hyperresponsiveness in adult mice. J Clin Invest. 2014;124:801–11.
Cui W, Zhang P, Gu J, Tian Y, Gao X, Liu Y, et al. Vitamin A deficiency promotes inflammation by induction of type 2 cytokines in experimental ovalbumin-induced asthma murine model. Inflammation. 2016;39:1798–804.
Yokota-Nakatsuma A, Takeuchi H, Ohoka Y, Kato C, Song SY, Hoshino T, et al. Retinoic acid prevents mesenteric lymph node dendritic cells from inducing IL-13-producing inflammatory Th2 cells. Mucosal Immunol. 2014;7:786–801.
Schuster GU, Kenyon NJ, Stephensen CB. Vitamin A deficiency decreases and high dietary vitamin A increases disease severity in the mouse model of asthma. J Immunol. 2008;180:1834–42.
Matheu V, Berggard K, Barrios Y, Barrios Y, Arnau MR, Zubeldia JM, et al. Impact on allergic immune response after treatment with vitamin A. Nutr Metab (Lond). 2009;6:44.
Ruhl R. Non-pro-vitamin A and pro-vitamin A carotenoids in atopy development. Int Arch Allergy Immunol. 2013;161:99–115.
Vasiliou JE, Lui S, Walker SA, Chohan V, Xystrakis E, Bush A, et al. Vitamin D deficiency induces Th2 skewing and eosinophilia in neonatal allergic airways disease. Allergy. 2014;69:1380–9.
Agrawal T, Gupta GK, Agrawal DK. Vitamin D supplementation reduces airway hyperresponsiveness and allergic airway inflammation in a murine model. Clin Exp Allergy. 2013;43:672–83.
Fischer KD, Hall SC, Agrawal DK. Vitamin D supplementation reduces induction of epithelial-mesenchymal transition in allergen sensitized and challenged mice. PLoS ONE. 2016;11:e149180.
Lu H, Xie RD, Lin R, Zhang C, Xiao XJ, Li LJ, et al. Vitamin D‑deficiency induces eosinophil spontaneous activation. Cell Immunol. 2017;322:56–63.
Matsui T, Yamashita H, Saneyasu KI, Tanaka H, Ito K, Inagaki N. Vitamin D deficiency exacerbates sensitization and allergic diarrhea in a murine food allergy model. Allergol Int. 2017; https://doi.org/10.1016/j.alit.2017.08.010.
Heine G, Tabeling C, Hartmann B, Gonzalez Calera CR, Kuhl AA, Lindner J, et al. 25-hydroxvitamin D3 promotes the long-term effect of specific immunotherapy in a murine allergy model. J Immunol. 2014;193:1017–23.
Mirzakhani H, Al-Garawi A, Weiss ST, Litonjua AA. Vitamin D and the development of allergic disease: how important is it? Clin Exp Allergy. 2015;45:114–25.
Jerzynska J, Stelmach W, Rychlik B, Lechanska J, Podlecka D, Stelmach I. The clinical effect of vitamin D supplementation combined with grass-specific sublingual immunotherapy in children with allergic rhinitis. Allergy Asthma Proc. 2016;37:105–14.
Heine G, Hollstein T, Treptow S, Radbruch A, Worm M. 9‑cis retinoic acid modulates the type I allergic immune response. J Allergy Clin Immunol. 2018; 141:650–8.
Seo GY, Lee JM, Jang YS, Kang SG, Yoon SI, Ko HJ, et al. Mechanism underlying the suppressor activity of retinoic acid on IL4-induced IgE synthesis and its physiological implication. Cell Immunol. 2017;322:49–55.
Liu L, Gudas LJ. Disruption of the lecithin:retinol acyltransferase gene makes mice more susceptible to vitamin A deficiency. J Biol Chem. 2005;280(48):40226–34.
Czarnewski P, Das S, Parigi SM, Villablanca EJ. Retinoic acid and its role in modulating intestinal innate immunity. Nutrients. 2017; https://doi.org/10.3390/nu9010068.
Dawicki W, Li C, Town J, Zhang X, Gordon JR. Therapeutic reversal of food allergen sensitivity by mature retinoic acid-differentiated dendritic cell induction of LAG3+CD49b-Foxp3-regulatory T cells. J Allergy Clin Immunol. 2017;139:1608–1620.e3.
Wu J, Zhang Y, Liu Q, Zhong W, Xia Z. All-trans retinoic acid attenuates airway inflammation by inhibiting Th2 and Th17 response in experimental allergic asthma. Bmc Immunol. 2013;14:28.
Son HL, Park HR, Park YJ, Kim SW. Effect of retinoic acid in a mouse model of allergic rhinitis. Allergy Asthma Immunol Res. 2015;7:590–8.
Lai G, Wu C, Hong J, Song Y. 1,25-Dihydroxyvitamin D(3) (1,25-(OH)(2)D(3)) attenuates airway remodeling in a murine model of chronic asthma. J Asthma. 2013;50:133–40.
Wang Z, Zhang H, Sun X, Ren L. The protective role of vitamin D3 in a murine model of asthma via the suppression of TGF-beta/Smad signaling and activation of the Nrf2/HO-1 pathway. Mol Med Rep. 2016;14:2389–96.
Schedel M, Jia Y, Michel S, Takeda K, Domenico J, Joetham A, et al. 1,25D3 prevents CD8(+)Tc2 skewing and asthma development through VDR binding changes to the Cyp11a1 promoter. Nat Commun. 2016;7:10213.
Yip KH, Kolesnikoff N, Yu C, Hauschild N, Taing H, Biggs L, et al. Mechanisms of vitamin D(3) metabolite repression of IgE-dependent mast cell activation. J Allergy Clin Immunol. 2014;133:1356–64. e1-14.
Liu ZQ, Li XX, Qiu SQ, Yu Y, Li MG, Yang LT, et al. Vitamin D contributes to mast cell stabilization. Allergy. 2017;72:1184–92.
Maslova E, Hansen S, Strom M, Halldorsson TI, Olsen SF. Maternal intake of vitamins A, E and K in pregnancy and child allergic disease: a longitudinal study from the Danish National Birth Cohort. Br J Nutr. 2014;111:1096–108.
Litonjua AA, Carey VJ, Laranjo N, Harshfield BJ, McElrath TF, O’Connor GT, et al. Effect of prenatal supplementation with vitamin D on asthma or recurrent wheezing in offspring by age 3 years: the VDAART Randomized Clinical Trial. JAMA. 2016;315:362–70.
Pali-Scholl I, Namazy J, Jensen-Jarolim E. Allergic diseases and asthma in pregnancy, a secondary publication. World Allergy Organ J. 2017;10:10.
Reifen R, Berkovich Z, Mandelberg A. Vitamin A supplementation via aerosol spray in asthmatic children. Pediatr Allergy Immunol. 2015;26:578–9.
Hamalainen N, Nwaru BI, Erlund I, Takkinen HM, Ahonen S, Toppari J, et al. Serum carotenoid and tocopherol concentrations and risk of asthma in childhood: a nested case-control study. Clin Exp Allergy. 2017;47:401–9.
Hypponen E, Berry DJ, Wjst M, Power C. Serum 25-hydroxyvitamin D and IgE—a significant but nonlinear relationship. Allergy. 2009;64:613–20.
Majak P, Olszowiec-Chlebna M, Smejda K, Stelmach I. Vitamin D supplementation in children may prevent asthma exacerbation triggered by acute respiratory infection. J Allergy Clin Immunol. 2011;127:1294–6.
Sennekamp J. Bronchitis and asthma in the setting of vitamin D deficiency. Allergo J Int. 2014;23:113–4.
Funding
This work was supported by grant SFB F4606-B28 of the Austrian Science Fund FWF to EJJ.
Funding
Open access funding provided by Austrian Science Fund (FWF).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
E. Jensen-Jarolim is inventor on EP 2894478 A1, owned by Biomedical International R+D GmbH, Vienna, Austria. K. Hufnagl declares that she has no competing interests.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Hufnagl, K., Jensen-Jarolim, E. Vitamin A and D in allergy: from experimental animal models and cellular studies to human disease. Allergo J Int 27, 72–78 (2018). https://doi.org/10.1007/s40629-018-0054-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40629-018-0054-2