
Abstract
This paper proposes and analyses a nonlinear mathematical model to study the impact of anthropogenic activities on forest biomass and forest-dependent wildlife populations using a system of differential equations. It is assumed that the growth of forest biomass, forest-dependent wildlife populations, and the human population follow logistic equations. The effect of forest biomass depletion on the survival of forest-dependent wildlife populations is investigated by introducing a function that denotes the dependence on forest biomass. The system’s behaviour near all ecologically acceptable equilibria is studied, and to confirm the analytical conclusions, a numerical simulation is performed. The model analysis shows that as forest biomass declines due to an increase in human population and its associated activities, the population of wildlife species also declines, and if no measures are taken, both forest biomass and the wildlife population may become extinct.










Similar content being viewed by others
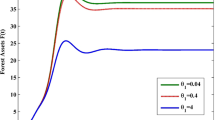
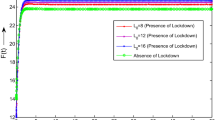

Availability of data and materials
Data used for numerical simulations can be obtained from the cited references and within the manuscript. The MATLAB codes can be provided upon request from the corresponding author.
References
Bologna M, Aquino G (2020) Deforestation and world population sustainability: a quantitative analysis. Sci Rep 10(1):1–9. https://doi.org/10.1038/s41598-020-63657-6
MNRT (2015) National forest resources monitoring and assessment of Tanzania mainland (naforma). Technical report, Tanzania Forest Services Agency, Dar es Salaam,Tanzania
FAO, UNEP (2020) The State of the World’s Forests 2020 (SOFO): forests, biodiversity and people. Technical report, FAO and UNEP, Rome, Italy. https://doi.org/10.4060/ca8642en
Kideghesho JR (2015) Realities on deforestation in Tanzania: trends, drivers, implications and the way Forward. In: Zlatic M (ed) Precious forests. IntechOpen, Rijeka. Chap. 2. https://doi.org/10.5772/61002
Kaeslin E, Williamson D (2010) Forests, people and wildlife: challenges for a common future. Unasylva 61(236):3–10
Bwalya SM (2011) Household dependence on forest income in selected rural communities in Zambia. Zamb Soc Sci J 2(1):67–86
Goshu MD, Endalew MF (2022) Mathematical modeling on conservation of depleted forestry resources. Nat Resour Model 35:1–21
Jyotsna K, Tandon A (2017) A mathematical model to study the impact of mining activities and pollution on forest resources and wildlife population. J Biol Syst 25(02):207–230. https://doi.org/10.1142/S0218339017500115
Pratama MAA, Zikkah RN, Anggriani N, Supriatna AK (2020) A mathematical model to study the effects of population pressure on two-patch forest resources. In: AIP conference proceedings, vol 2264, pp 1–9. AIP Publishing LLC, Bali, Indonesia. https://doi.org/10.1063/5.0023844
Lata K, Misra AK, Shukla JB (2018) Modeling the effect of deforestation caused by human population pressure on wildlife species. Nonlinear Anal Model Control 23(3):303–320. https://doi.org/10.15388/NA.2018.3.2
Lata K, Misra AK (2017) Modeling the effect of economic efforts to control population pressure and conserve forestry resources. Nonlinear Anal Model Control 22(4):473–488. https://doi.org/10.15388/NA.2017.4.4
Wu W, Li Y, Hu Y, Chang Y, Xiong Z (2019) Anthropogenic effect on forest landscape pattern and Cervidae habitats in northeastern china. J Geog Sci 29(7):1098–1112. https://doi.org/10.1007/s11442-019-1647-5
Jyotsna K, Tandon A (2017) A mathematical model studying the survival of forest resource-dependent wildlife population in the presence of population pressure-induced mining activities. Nat Resour Model 30(4):12139. https://doi.org/10.1111/nrm.12139
Sinha SK, Pal J, Jyotsna K (2022) A dynamical study on the adverse effects of industrial activities in the forest and wildlife region through modelling. Model Earth Syst Environ 1–13
Fanuel IM, Kajunguri D, Moyo F (2023) Modelling the impact of human population and its associated pressure on forest biomass and forest–dependent wildlife population. J Appl Math. https://doi.org/10.1155/2023/4826313
Pathak R, Verma V, Agarwal M (2021) Impact of human activities on forest resources and wildlife population. Comput Ecol Softw 11(2):83–99
Mekonen S (2020) Coexistence between human and wildlife: the nature, causes and mitigations of human wildlife conflict around bale mountains national park, southeast ethiopia. BMC Ecol 20(51):1–9
Matandiko K (2023) In tanzania, wild animals kill one person every 52 hours. The Citizens. Available at: https://shorturl.at/jzABV (Accessed: May 16th, 2023)
Freedman HI (1980) Deterministic mathematical models in population ecology, vol 57. Marcel Dekker Incorporated, New York
La Salle J (1961) Stability by Liapunov’s direct method with applications. Academic Press, Cambridge
Gümüş ÖA (2014) Global and local stability analysis in a nonlinear discrete-time population model. Adv Differ Equ 2014(1):1–9. https://doi.org/10.1186/1687-1847-2014-299
Seydel R (2009) Practical bifurcation and stability analysis, vol 5. Springer, New York
Castillo-Chavez C, Song B (2004) Dynamical models of tuberculosis and their applications. Math Biosci Eng 1(2):361–404
Freedman HI, Ruan SG (1995) Uniform persistence in functional differential equations. J Differ Equ 115(1):173–192
Misra AK, Lata K, Shukla JB (2014) Effects of population and population pressure on forest resources and their conservation: a modeling study. Environ Dev Sustain 16(2):361–374. https://doi.org/10.1007/s10668-013-9481-x
Panja P (2020) Is the forest biomass a key regulator of global warming? A mathematical modelling study. Geol Ecol Landsc 6(1):66–74. https://doi.org/10.1080/24749508.2020.1752021
Misra AK, Lata K (2015) A mathematical model to achieve sustainable forest management. Int J Model Simul Sci Comput 6(4):1550040. https://doi.org/10.1142/S1793962315500403
Chaudhary M, Dhar J, Misra OP (2015) A mathematical model for the conservation of forestry biomass with an alternative resource for industrialization: a modified leslie gower interaction. Model Earth Syst Environ 1(4):1–10. https://doi.org/10.1007/s40808-015-0056-8
Chen F (2005) On a nonlinear nonautonomous predator-prey model with diffusion and distributed delay. J Comput Appl Math 180(1):33–49. https://doi.org/10.1016/j.cam.2004.10.001
Acknowledgements
The authors thank the Nelson Mandela African Institution of Science and Technology (NM-AIST) for providing good learning facilities and working conditions.
Funding
No funds, grants, or other support was received.
Author information
Authors and Affiliations
Contributions
The study conception, design, material preparation, and analysis were contributions made by all authors. Ibrahim Fanuel drafted the initial manuscript, and all authors provided feedback on previous versions. The final manuscript was read and approved by all authors.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no competing interests to declare that are relevant to the content of this article.
Appendices
Appendix A: Proof of Lemma 1
Proof
Following Chaudhary et al. [28] and Chen [29], the proof is as follows. Consider the first equation of the system (1). When \(B \rightarrow L\) the rest of the variables become zero, this gives,
Further, from equations two, three, and four of the system, respectively, we get Eqs. (A1), (A2) and (A3).
\(\square \)
Appendix B: Proof of Theorem 3
Proof
To prove Theorem (3), we start by linearising the system (1), about \(E^*\) by using the following transformations:
\(B=B^*+b\), \(W=W^*+w\), \(N=N^*+n\), \(H=H^*+h\), where b, w, n and h are small perturbations around the equilibrium \(E^*\). The following linearised system is obtained:
Following Lata [10], we considered the positive definite function (B5),
where \(k_1\), \(k_2\) and \(k_3\) are positive constants. Upon differentiating function (B5) with respect to t along the solutions of linearized system (B4), the following is obtained:
Eq. (B6) can be arranged as follows:
Choosing \( k_1=\dfrac{1}{\eta }, \) \( k_2=\dfrac{1}{\lambda N^*},\) and \(k_3=1\) which are all positive constants, Eq. (B7) is written as the sum of quadratic expressions
When \(\dfrac{{\text {d}}V}{{\text {d}}t}<0\), the model system (1) becomes locally asymptotically stable in the given region of attraction. Therefore, \(\dfrac{{\text {d}}V}{{\text {d}}t}\) is negative definite if and only if conditions (18)–(22) hold. \(\square \)
Appendix C: Proof of Theorem 4
Proof
We consider the following positive definite function [8] to prove the theorem
where \(l_1\), \(l_2\) and \(l_3\) are positive constants. It can be easily checked that, \(U(B^*, W^*, N^*, H^*)=0\) and \(U(B, W, N, H)>0\) for all other positive values of \(\left\{ B, W, N, H\right\} -\left\{ B^*, W^*, N^*,\right. \)\(\left. H^* \right\} \).
On differentiating (C9) with respect to "t", the following is obtained:
Substituting the values of \(\dfrac{{\text {d}}B}{{\text {d}}t}\), \(\dfrac{{\text {d}}W}{{\text {d}}t}\), \(\dfrac{{\text {d}}N}{{\text {d}}t}\) and \(\dfrac{{\text {d}}H}{{\text {d}}t}\) from the system (1), the following equation is obtained;
where,
Utilizing mean value theorem, we have
such that \(0<K'(B)\le \lambda _1.\) Choosing \( l_1=\dfrac{1}{\eta }\), \(l_2=\dfrac{1}{\lambda }\) and \(l_3= 1\), consequently, \(\dfrac{{\text {d}}U}{{\text {d}}t}\) is reduced to:
Where,
When \(\dfrac{{\text {d}}U}{{\text {d}}t}<0\), the model system (1) becomes globally asymptotically stable in the region of attraction \(E^*\). Therefore, \(\dfrac{{\text {d}}U}{{\text {d}}t}\) is negative if and only if conditions (23)–(27) hold. Theorem 4 follows. \(\square \)
Proof
Following Chen [29], we begin by defining the following condition: The model system (1) is said to be uniformly persistent if there are positive constants \(\varOmega _1\) and \(\varOmega _2\) such that each of the positive solutions \(\left( B, W, N, H \right) \) of the system with positive initial conditions \(\left( B_0, W_0, N_0, H_0 \right) \) satisfies Eq. (C11).
where \(\mathcal {X}(t)=\left( B, W, N, H \right) \).
From Lemma (1), we define \(\varOmega _2\) as
It follows that \(\limsup _{t\rightarrow \infty }\mathcal {X}(t)\le \varOmega _2.\)
This is also true for any sufficiently small \(\epsilon _i>0\), there exists a \(T_i>0\), such that
Now, taking \(\epsilon =\max \left\{ \epsilon _1, \epsilon _2, \epsilon _3, \epsilon _4\right\} \) and \(T=\max \left\{ T_1,\right. \)\(\left. T_2, T_3, T_4 \right\} \) it can be concluded that, for \(\epsilon >0\), there exists \(T>0\) such that for all \(t\ge T\), the following holds;
Thus, the first equation of the model system (1), for all \(t\ge T\), can be written as
Further, we have
which is true for every \(\epsilon >0\), hence,
such that \(s>\alpha K(L)+\beta _1N_m.\)
From the second model equation, we have
It follows
which is true for every \(\epsilon >0\), therefore,
where \(r(B_l)>\nu _1N_m+\nu _2H_m\).
Similarly, the third and fourth equations of the system (1), respectively, yield
Hence, it follows
where \(\theta >\sigma W_l.\)
Therefore, it follows
It is worth noting that \(\varOmega _1=\min \left\{ B_l, W_l, N_l, H_l \right\} .\) Theorem 5 follows. \(\square \)
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Fanuel, I.M., Mirau, S., Kajunguri, D. et al. Mathematical model to study the impact of anthropogenic activities on forest biomass and forest-dependent wildlife population. Int. J. Dynam. Control 12, 1314–1331 (2024). https://doi.org/10.1007/s40435-023-01265-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40435-023-01265-8