
Abstract
Purpose of the review
The abuse of opioids induces many terrible problems in human health and social stability. For opioid-dependent individuals, withdrawal memory can be reactivated by context, which is then associated with extremely unpleasant physical and emotional feelings during opioid withdrawal. The reactivation of withdrawal memory is considered one of the most important reasons for opioid relapse, and it also allows for memory modulation based on the reconsolidation phenomenon. However, studies exploring withdrawal memory modulation during the reconsolidation window are lacking. By summarizing the previous findings about the reactivation of negative emotional memories, we are going to suggest potential neural regions and systems for modulating opioid withdrawal memory.
Recent findings
Here, we first present the role of memory reactivation in its modification, discuss how the hippocampus participates in memory reactivation, and discuss the importance of noradrenergic signaling in the hippocampus for memory reactivation. Then, we review the engagement of other limbic regions receiving noradrenergic signaling in memory reactivation. We suggest that noradrenergic signaling targeting hippocampus neurons might play a potential role in strengthening the disruptive effect of withdrawal memory extinction by facilitating the degree of memory reactivation.
Summary
This review will contribute to a better understanding of the mechanisms underlying reactivation-dependent memory malleability and will provide new therapeutic avenues for treating opioid use disorders.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Opioids comprise heroin, natural and semisynthetic opioids (such as morphine, codeine, hydrocodone, and oxycodone), and synthetic opioids other than methadone (such as fentanyl, fentanyl analogs, and tramadol) [1]. Chronic use of any of the above drugs could prompt opioid use disorder (OUD), which causes clinically serious suffering or impairment [2]. In the World Drug Report 2021 [3], an estimated 62 million people or 1.2% of the global population used opioids for non-medical purposes in 2019, which was nearly double over the past decades. Among that, almost 13 million people died, which accounted for 70% of the total deaths from drug use disorder. In particular, since the coronavirus disease (COVID-19) pandemic swept the world, opioid overdose deaths have increased observably. OUD is characterized by a high rate of relapse. In a large-scale survey, over 80% of those who completed opioid withdrawal relapsed within one year [4]. The transition from recreational to compulsive drug use has been proposed to entail a neuroadaptive process that shifts motivational processing from seeking positive reinforcement to avoiding the aversive effects of withdrawal [5]. Opioid users become physically dependent and are subject to aversive withdrawal symptoms (such as nausea, vomiting, and diarrhea) when drug levels fall too low or when the drugs are stopped abruptly [6]. Thus, withdrawing patients have intense opioid cravings associated with the feeling of being unwell [7,8,9]. Although opioid withdrawal’s duration varies depending on the specific opioid being used, withdrawal response in most types of opioids only lasts several days. For example, symptoms of heroin withdrawal may last for 7–14 days [10]. However, conditioned withdrawal triggers opioid relapses that occur after the duration of withdrawal symptoms [11]. That is, one of the most important reasons for relapses is the retention of the opioid withdrawal memory induced by the conditioned context previously associated with withdrawal symptoms [12]. Therapeutic approaches to mediating withdrawal memory may be valuable in preventing opioid relapse.
Previous studies on drug abuse have considered approaches to weakening or removing maladaptive reward memories. Cue-exposure treatment in extinction was demonstrated as an important part of reward-related memory modulation [11, 13, 14]. Unfortunately, persistent maladaptive memories that maintain drug seeking and are resistant to extinction are a trademark of addiction. Additionally, extinction learning has several important limitations, the most important of which is the contextual specificity of extinction learning [15]. Despite the certain success of cue-exposure therapy, the long-term efficacy of this treatment remains extremely dubious because extinction learning might not thoroughly delete the previous memory traces but rather creates new learning that inhibits the original memory’s activation and thus is subject to relapse even after long periods of lessening [16,17,18]. Afterward, the reconsolidation phenomenon during the memory process was observed, and the disruption of memory reconsolidation after retrieval has received attention for its therapeutic potential [19]. Modification of memory after retrieval supports the hypothesis that reconsolidation is a true, specific process that maintains, strengthens, and possibly updates memory [20]. Many studies have shown that the memory reactivation–extinction pattern outperforms simple exposure to the context or cue in preventing addiction memory [13, 14, 21, 22]. Hence, whether memory has been activated or not and the degree of activation may be important factors in modulating the original memory. Nevertheless, studies regarding aversive withdrawal memory based on reconsolidation theory are lacking. Revealing the brain circuits handling the activation of opioid withdrawal memory may contribute to developing behavioral and neural regulation therapies for modifying pathological memory in OUD.
The hippocampus is one of the earliest attended brain regions in memory studies [23] and has demonstrated a crucial role in memory reactivation [24, 25]. Previous reports have shown that noradrenergic agents significantly affect memory modulation, which may be achieved by modulating the memory reactivation process [26,27,28]. Locus coeruleus (LC) is a major resource for noradrenergic neurotransmitters, projecting to the hippocampus [29, 30]. Meanwhile, LC is among the brain regions first studied for its response to opioid withdrawal [31, 32]. Here, we review hippocampus involvement in memory reactivation and suggest a potential role of noradrenergic signaling in the hippocampus and LC-hippocampus circuits for disrupting withdrawal memory based on the reactivation–extinction pattern. In particular, following this review’s aim, studies focusing on negative emotion-related memory are mainly discussed, and research on drug reward-associated memory is also mentioned.
Memory is labile under active conditions
Memory includes the acquisition, consolidation, and retrieval phases [33]. From an evolutionary perspective, it is highly functional to remember the most important events in life [34]. However, the putative indelibility of emotional memory can also be harmful and maladaptive, such as in drug abuse and post-stress traumatic disorder [35, 36]. In 2000, the reactivation–extinction pattern was proposed as a new approach targeting memory reconsolidation in a fear memory study [37]. This has been interpreted as proving that reactivated memories re-enter a state of lability and that the pulp will be re-stabilized through a protein-dependent process [37, 38]. Reconsolidation has a time window, and only if the memory is altered in this interval should the memory modification be successful. Much evidence has identified that the reconsolidation window is not fixed. For instance, some studies have shown that it was within six hours after memory reactivation [39, 40], while others have found that memory modification was not impaired beyond three hours after reactivation [41]. The difference may be related to the strength of the original memory [42], memory type [43], and the reactivation degree [44]. Anyhow, memories can be disrupted by amnesic treatments delivered shortly after their reactivation. The opportunity to eliminate pathological memories through pharmacological and behavioral treatment during the reconsolidation window has also been considered in the research field of drug abuse, especially focusing on drug-associated reward memory [21, 39, 41].
Previous research has revealed that a procedure that utilizes memory reactivation to make extinction more effective disrupts maladaptive memories, but the limitations of reactivation–extinction patterns have also recently been noticed for memory modulation. First, the preventing effect of the paradigm is selective to conditioned stimuli (CS) [39, 45]. Whether the memory could be efficaciously activated depends on the degree of similarity between the exposed environment during the memory retrieval phase and CS in the original memory acquisition. Second, the time elapsed since initial learning may be another important factor in context–exposure-induced memory reactivation. In some cases, the inhibition of response by the reactivation–extinction manipulation only exhibits a short duration after the memory acquisition [46]. Some researchers have attempted to find other reliable ways to trigger memory activation to avoid these limitations. Unconditioned stimuli (UCSs) are another possibility to drive memory reactivation, such as a low dose of drugs for addiction-relative reward memory and a weak shock for fear memory [47]. Compared to CS, UCS seems more effective for reactivating memory. A study on drug reward memory showed that extinction after UCS exposure impaired the reinstatement of cocaine-seeking behavior and disrupted non-extinguished CS-induced cocaine-seeking behavior [47]. A similar result was found in the extinction of fear condition memory [48]. There is a possibility that UCS could induce more alterations in intracellular molecules, which implies that UCS could trigger a more unstable state of original memory than CS [48]. However, its neuronal mechanism remains unclear. Furthermore, this approach may paradoxically accelerate the reacquisition of the previously extinguished conditioned response or retard the memory’s extinction [21, 47]. Ethical issues still seem to face UCS usage in memory reactivation.
Collectively, although there are some uncertainties regarding CS in memory modulation, it has been generally accepted that memory is relatively labile when it is in an active state. Maximizing memory reactivation would contribute to the efficiency of the reactivation–extinction approach in modifying pathological memory. Since studies on the modulation of opioid withdrawal memory depending on reconsolidation theory are unexplored, understanding the neuronal mechanisms underlying negative emotion-associated memory reactivation is necessary.
The function of the hippocampus in memory reactivation
The hippocampus is often mentioned in studies for opioid withdrawal memory [49,50,51] and other types of aversive memory reactivation [52,53,54,55,56] (Table 1). Comprehensive presentation of the function of this brain region in memory reactivation comes from fear memory studies [24, 57]. The hippocampus cytoarchitectonically includes the dentate gyrus (DG) and the Cornu Ammonis subfields (CA1-3). Among them, DG, CA1, and CA3 have been demonstrated to be involved in the memory reactivation process [57,58,59,60,61]. In 1904, the term “memory engram” was first used to describe the memory representations by Richard Semon [62]. The hippocampus is mainly involved in studies of memory engrams [63, 64]. A landmark study identified a small subpopulation of granule cells in the DG but not in CA1 of the hippocampus as contextual memory-engram cells, and optogenetic stimulation of these cells is sufficient to activate behavioral retrieval of a context-dependent fear memory formed by foot shocks’ delivery [65]. In another recent study, a subset of memory retrieval-induced neurons in the DG became reactivated during extinction, and the degree of fear reduction was positively correlated with this reactivation after extinction training [57]. These findings further contribute to the understanding of the effect of reactivation–extinction patterns in memory modulation. Unlike DG, the hippocampal area CA3 is suggested to be of minimal importance for contextual memory reactivation [66]. Although it remains unknown whether the engram neurons in the hippocampal CA1 are also reactivated during extinction learning, the previous report that membrane excitability was increased in hippocampal CA1 neurons immediately after the retrieval of contextual fear memory may imply that CA1 is also involved in memory re-writing when the reactivation–extinction training is conducted [58]. It is possible that memory-associated information may flow through the DG-CA3-CA1 neural circuit during memory reactivation [67].
In particular, the induction of the immediate-early gene Arc in the DG was positively correlated with a higher aversion score in morphine-dependent animals [76], implying that the hippocampal DG participates in withdrawal memory reactivation. Recent studies have also revealed the role of hippocampal CA1 in withdrawal memory reactivation with some pharmacological and immunostaining methods [51]. However, how these hippocampal subregions engage in this process still requires further studies with advanced techniques in the neuroscience field.
The role of noradrenergic signaling in the hippocampus for memory reactivation
Several studies have demonstrated the critical and specific role of noradrenergic signaling in the hippocampus for reactivation in different types of memory (Table 2). By behavioral pharmacological manipulation, the causal relationship between noradrenergic signaling in the hippocampus and memory reactivation has been revealed. For example, the infusion of a β-adrenergic receptor antagonist into the hippocampal DG of rats shortly before testing impaired appetitive spatial reference memory expression [87]. Systemic administration or dorsal hippocampal infusion of a β-adrenergic receptor antagonist shortly before testing blocks the expression of cocaine place preference in rats [88]. Additionally, propranolol blocks membrane hyperexcitability in hippocampal CA1 neurons induced by fear memory retrieval [58], suggesting that aversive memory reactivation is also facilitated by β-receptor excitation in CA1. Furthermore, context exposure could evoke noradrenaline release in the hippocampus, stimulating β-adrenergic receptors and thereby causing an increase in intracellular cyclic adenosine monophosphate (cAMP), which then stimulates the activation of protein kinase A (PKA) [58]. PKA activation may increase neuronal excitability by phosphorylation ion channels and by increasing glutamatergic NMDAR-mediated calcium influx [58].
In the hippocampus, β-adrenergic receptors are mainly expressed by DG granular and CA1 pyramidal neurons [29, 30]. The LC is connected to the hippocampus via strong noradrenergic fiber projections [107]. Some studies have revealed that the LC plays a potential role in functional modulation of the hippocampus. Supporting evidence includes that the LC’s stimulation in the rats induced long-term potentiation in the hippocampus [108], that increased levels of hippocampal noradrenaline facilitated the synaptic delivery of glutamatergic AMPA receptors necessary for long-term potentiation [109], and that electrical activation of the LC induces noradrenergic release in the hippocampus, especially in the DG subregion [110]. Besides direct binding to β-adrenergic receptors on hippocampal neurons [111], astrocytes in the hippocampus may also participate in the process of memory retrieval modulated by noradrenergic signals. This is because β-adrenergic receptor distributions in hippocampal astrocytes provide the fundamentals for engaging astrocytes [112]. Previous evidence has shown the relationship between neurons and astrocytes in the hippocampus and that repeated stimulations to astrocytes cause CA1 pyramidal neurons to firing synchronously through the NR1/NR2B subunits of glutamatergic NMDA receptors [113]. Given that a crucial role of NR2B has been demonstrated in memory reactivation [114, 115], astrocytes in the hippocampus might be involved in the memory reactivation modulated by β-adrenergic signaling.
All noradrenergic neurons of the LC intensely expressed immediate-early gene c-fos mRNA during naloxone-precipitated morphine withdrawal [116]. Indeed, the LC and noradrenergic systems in this brain region have attracted much attention in opioid dependence studies since those early years. The withdrawal response has been attributed, at least in part, to elevated activity in the noradrenergic cells of LC [31, 32]. However, a total lesion of noradrenergic neurons of the LC did not alter naloxone-precipitated morphine withdrawal [117], challenging the idea that this structure is vital in producing somatic signs [118, 119]. The LC may be involved in retrieving aversive learned associations, contrasting with the common idea of excluding the LC from the motivational component of withdrawal [116, 117].
Other neurotransmitters and brain regions involved in LC-modulated memory reactivation
The noradrenergic signal and the dopaminergic system are involved in LC-modulated memory reactivation [120]. The hippocampus may be the downstream area of this function of the LC [121]. Although the ventral tegmental area (VTA) is the main brain source for dopaminergic neurotransmitters but not the LC, the primary input from VTA is to the ventral subregion of the hippocampus with only minimal input to the dorsal hippocampus, and none is observed for the stratum radiatum of the dorsal hippocampus [122,123,124]. Given that activation of D1 dopaminergic receptors in proximity to CA3-CA1 synapses in the stratum radiatum of the dorsal hippocampus is required for hippocampal-dependent learning and memory [125, 126], noradrenergic fibers from the LC may be the primary source of dopamine release in the dorsal hippocampus [107, 127]. Therefore, combining the use of dopaminergic and noradrenergic receptor blockades or activation agents in a particular time window, behavioral treatment may develop a helpful approach against pathological memory.
In addition to the hippocampus, the amygdala is another important region in memory encoding and retrieval [97, 128,129,130]. Both the amygdala and the hippocampus are directly innervated by the LC [131, 132]. Noradrenergic modulation of memory likely occurs via projections of the LC to the hippocampus and the amygdala [133] (Table 2). The hippocampus and amygdala may control different aspects of memory. Case studies on lesions in humans suggest a double separation, with hippocampus lesions affecting declarative retrieval of fear conditioning and amygdala lesions affecting only arousal response [134]. Some studies have suggested that the connections between the LC and the amygdala are associated with arousal or aversive responses, while LC projections to the hippocampus are more relevant for the improved encoding of memories [135, 136]. This indication is supported by a previous study in which noradrenergic transmitter release increased in the amygdala during naloxone-precipitated withdrawal [137].
The cerebral cortex is another important brain region receiving the projection from LC [138]. There are few studies showing the involvement of the LC-prefrontal cortex circuit in the retrieval of reward memory or recognition [100, 102]. Direct evidence is lacking for the modulatory function of noradrenergic signaling in the prefrontal cortex in memory reactivation. However, some studies have shown that the prefrontal cortex plays a crucial role in fear memory retrieval [12, 139]. A new perspective suggests a pattern of hippocampal–neocortical interactions in memory retrieval [140]. In addition, the entorhinal cortex, which is closely located by the hippocampus, might be important in memory modulation by connecting a CA1 or a DG subregion of the hippocampus [141].
Conclusions
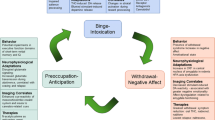
Although it is controversial whether the extinction learning process changes the original memory or establishes a new memory that links to the existing trace [22, 45, 142, 143], the above analysis and discussion suggest that noradrenergic signaling from LC (targeting the hippocampus and other limbic regions) may play a vital role in memory reactivation, providing a basis for the subsequent extinction in the reconsolidation window. Besides noradrenaline-related agents, selective neural stimulation is another choice for reducing pathological memory. Recently, non-invasive brain stimulations, such as transcranial magnetic stimulation (TMS), transcranial direct current stimulation (tDCS), and transcranial alternating current stimulation (tACS), have been applied to intervene in many types of neuropsychiatric disorders [144,145,146,147]. Although these techniques remain unsuitable for targeting deep brain regions, some applicable approaches, such as temporal interference (TI), have been developed recently [148, 149]. An animal study reveals that tDCS modulates excitability in a polarity-specific manner and selectively affects subregions of the hippocampus [150]. Additionally, indirect stimulation of LC seems to work by activating the vagus-nucleus tractus solitarii (NTS)-LC by TMS [151]. Moreover, LC can be activated by behavioral approaches, such as novelty exposure [152], which is based on LC’s physiological function of alertness and arousal [153, 154] (Fig. 1).

Potential frame work for withdrawal memory modulation. Withdrawal memory can be reactivated and become labile when the individual is exposed to the context which is associated with the withdrawal feelings. This labile state can last around 6 hours since the memory is reactivated, which is also called reconsolidation window. The reactivation–extinction pattern is proposed as an approach targeting memory reconsolidation. Maximizing memory reactivation will contribute to the efficiency of the reactivation–extinction approach in modifying pathological memory, such as withdrawal memory. We suggest that noradrenergic signaling from LC (specifically targeting the hippocampus) may play a vital role in facilitating memory reactivation, providing a basis for the subsequent extinction in the reconsolidation window. Optogenetics and chemogenetics are used to stimulate neural pathway directly in basic studies, while they are not suitable for the application in the human brain manipulation so far. The approaches of non-invasion brain stimulation, such as TMS, tDCS, tACS, and TI, have potential possibility for the indirect activation of LC and its projections. LC, locus coeruleus; Hippo, hippocampus; NTS, nucleus tractus solitarii; NA, noradrenaline; DA, dopamine; TMS, transcranial alternating current stimulation; tDCS, transcranial direct current stimulation; tACS, transcranial alternating current stimulation; TI, temporal interference
Theoretically, the idea of the reactivation–extinction paradigm has many advantages since the treatments may affect only the reactivated memory and not others, even closely related memories [155]. We raise the hypothesis that through pharmacological, behavioral, and even direct neural manipulations of the LC-hippocampal circuit, more engram cells are in an active state, and the original memory is more sensitive to modification in the extinction phase. However, noradrenaline from the LC is also involved in modulating the encoding and consolidation of hippocampus-based memory [138]. Thus, clarifying the mechanisms of the LC-hippocampal circuit underlying different memory phases may contribute to choosing the correct time window to conduct the manipulation. In this review, we only summarized the findings regarding the role of hippocampus in the retrieval of the memory, particularly in aversive memory and fear memory. This is a limitation that we did not cover other types or stages of memories, in which hippocampus has been demonstrated its involvements [156,157,158]. These need to be paid attentions in our further investigation and idea organization. In addition, there may be distinct mechanisms between normal memory and pathological memories. Under normal conditions, the reconsolidation state after memory retrieval may act to update and maintain memories. In contrast, under altered conditions due to acute or chronic drug use, stress, or genetic predisposition, reconsolidation may enhance memories, contributing to persistent drug-related memories [159]. More studies are required to dissect how and why some memories become abnormally strong, and others do not, which may be important to avoid the wrong treatments used in memory modulation.
References
Hedegaard H, Miniño AM, Warner M. Centers for Disease Control and Prevention. In: National vital statistics system: mordality data; 2020. https://wwwcdcgov/nchs/products/databriefs/db394htm.
Dydyk AM, Jain NK, Gupta M. Opioid Use Disorder. StatPearls. Treasure Island: StatPearls Publishing Copyright © 2020, StatPearls Publishing LLC; 2020.
https://www.unodc.org/unodc/en/data-and-analysis/wdr2021.html. World Drug Report. 2021.
Hunt WA, Barnett LW, Branch LG. Relapse rates in addiction programs. J Clin Psychol. 1971;27(4):455–6. https://doi.org/10.1002/1097-4679(197110)27:4<455::aid-jclp2270270412>3.0.co;2-r.
Pantazis CB, Gonzalez LA, Tunstall BJ, Carmack SA, Koob GF, Vendruscolo LF. Cues conditioned to withdrawal and negative reinforcement: neglected but key motivational elements driving opioid addiction. Sci Adv. 2021;7(15). https://doi.org/10.1126/sciadv.abf0364.
Hoffman RJ, Sharma AN. Chapter 183 - Withdrawal Syndromes. In: Zaoutis LB, Chiang VW, editors. Comprehensive Pediatric Hospital Medicine. Philadelphia: Mosby; 2007. p. 1140–3.
Wikler A. Recent progress in research on the neurophysiologic basis of morphine addiction. Am J Psychiatry. 1948;105(5):329–38. https://doi.org/10.1176/ajp.105.5.329.
Wikler A. Dynamics of drug dependence. Implications of a conditioning theory for research and treatment. Arch Gen Psychiatry. 1973;28(5):611–6. https://doi.org/10.1001/archpsyc.1973.01750350005001.
Wikler A. A theory of opioid dependence. NIDA Res Monogr. 1980;30:174–8.
O'Connor PG, Fiellin DA. Pharmacologic treatment of heroin-dependent patients. Ann Intern Med. 2000;133(1):40–54. https://doi.org/10.7326/0003-4819-133-1-200007040-00008.
Powell J, Gray JA, Bradley BP, Kasvikis Y, Strang J, Barratt L, et al. The effects of exposure to drug-related cues in detoxified opiate addicts: a theoretical review and some new data. Addict Behav. 1990;15(4):339–54. https://doi.org/10.1016/0306-4603(90)90044-x.
Song J, Shao D, Guo X, Zhao Y, Cui D, Ma Q, et al. Crucial role of feedback signals from prelimbic cortex to basolateral amygdala in the retrieval of morphine withdrawal memory. Sci Adv. 2019;5(2):eaat3210. https://doi.org/10.1126/sciadv.aat3210.
Childress AR, Hole AV, Ehrman RN, Robbins SJ, McLellan AT, O’Brien CP. Cue reactivity and cue reactivity interventions in drug dependence. NIDA Res Monogr. 1993;137:73–95.
Conklin CA, Tiffany ST. Applying extinction research and theory to cue-exposure addiction treatments. Addiction. 2002;97(2):155–67. https://doi.org/10.1046/j.1360-0443.2002.00014.x.
Bouton ME. Context, ambiguity, and unlearning: sources of relapse after behavioral extinction. Biol Psychiatry. 2002;52(10):976–86. https://doi.org/10.1016/s0006-3223(02)01546-9.
Conklin CA, Tiffany ST. Cue-exposure treatment: time for change. Addiction. 2002;97(9):1219–21. https://doi.org/10.1046/j.1360-0443.2002.00205.x.
Foa EB. Prolonged exposure therapy: past, present, and future. Depress Anxiety. 2011;28(12):1043–7. https://doi.org/10.1002/da.20907.
Myers KM, Carlezon WA Jr. D-cycloserine effects on extinction of conditioned responses to drug-related cues. Biol Psychiatry. 2012;71(11):947–55. https://doi.org/10.1016/j.biopsych.2012.02.030.
Tronson NC, Taylor JR. Addiction: a drug-induced disorder of memory reconsolidation. Curr Opin Neurobiol. 2013;23(4):573–80. https://doi.org/10.1016/j.conb.2013.01.022.
Kida S. Function and mechanisms of memory destabilization and reconsolidation after retrieval. Proc Jpn Acad Ser B Phys Biol Sci. 2020;96(3):95–106. https://doi.org/10.2183/pjab.96.008.
Sartor GC, Aston-Jones G. Post-retrieval extinction attenuates cocaine memories. Neuropsychopharmacology. 2014;39(5):1059–65. https://doi.org/10.1038/npp.2013.323.
Monfils MH, Cowansage KK, Klann E, LeDoux JE. Extinction-reconsolidation boundaries: key to persistent attenuation of fear memories. Science. 2009;324(5929):951–5. https://doi.org/10.1126/science.1167975.
Milner B, Penfield W. The effect of hippocampal lesions on recent memory. Trans Am Neurol Assoc. 1955;(80th Meeting):42–8.
Iwasaki S, Ikegaya Y. Contextual Fear memory retrieval is vulnerable to hippocampal noise. Cereb Cortex. 2021;31(2):785–94. https://doi.org/10.1093/cercor/bhaa257.
Molitor RJ, Sherrill KR, Morton NW, Miller AA, Preston AR. Memory reactivation during learning simultaneously promotes dentate Gyrus/CA (2,3) pattern differentiation and CA (1) memory integration. J Neurosci. 2021;41(4):726–38. https://doi.org/10.1523/jneurosci.0394-20.2020.
Schwabe L, Römer S, Richter S, Dockendorf S, Bilak B, Schächinger H. Stress effects on declarative memory retrieval are blocked by a beta-adrenoceptor antagonist in humans. Psychoneuroendocrinology. 2009;34(3):446–54. https://doi.org/10.1016/j.psyneuen.2008.10.009.
Sara SJ. The locus coeruleus and noradrenergic modulation of cognition. Nat Rev Neurosci. 2009;10(3):211–23. https://doi.org/10.1038/nrn2573.
Devauges V, Sara SJ. Memory retrieval enhancement by locus coeruleus stimulation: evidence for mediation by beta-receptors. Behav Brain Res. 1991;43(1):93–7. https://doi.org/10.1016/s0166-4328(05)80056-7.
Nicholas AP, Pieribone V, Hokfelt T. Distributions of mRNAs for alpha-2 adrenergic receptor subtypes in rat brain: an in situ hybridization study. J Comp Neurol. 1993;328(4):575–94. https://doi.org/10.1002/cne.903280409.
Milner TA, Shah P, Pierce JP. beta-adrenergic receptors primarily are located on the dendrites of granule cells and interneurons but also are found on astrocytes and a few presynaptic profiles in the rat dentate gyrus. Synapse. 2000;36(3):178–93. https://doi.org/10.1002/(sici)1098-2396(20000601)36:3<178::Aid-syn3>3.0.Co;2-6.
Rasmussen K, Beitner-Johnson DB, Krystal JH, Aghajanian GK, Nestler EJ. Opiate withdrawal and the rat locus coeruleus: behavioral, electrophysiological, and biochemical correlates. J Neurosci. 1990;10(7):2308–17. https://doi.org/10.1523/jneurosci.10-07-02308.1990.
Streel E, Dan B, Campanella S, Meyvaert A, Hanak C, Pelc I, et al. A pharmacological modulation of opiate withdrawal using an up-/down-regulation of the noradrenergic system in opiate-dependent rats. Int J Neuropsychopharmacol. 2006;9(5):621–6. https://doi.org/10.1017/s1461145705006024.
McGaugh JL. Memory--a century of consolidation. Science. 2000;287(5451):248–51. https://doi.org/10.1126/science.287.5451.248.
Baldi E, Bucherelli C. Brain sites involved in fear memory reconsolidation and extinction of rodents. Neurosci Biobehav Rev. 2015;53:160–90. https://doi.org/10.1016/j.neubiorev.2015.04.003.
Maddox SA, Hartmann J, Ross RA, Ressler KJ. Deconstructing the Gestalt: Mechanisms of Fear, Threat, and Trauma Memory Encoding. Neuron. 2019;102(1):60–74. https://doi.org/10.1016/j.neuron.2019.03.017.
Taujanskaitė U, Cahill EN, Milton AL. Targeting drug memory reconsolidation: a neural analysis. Curr Opin Pharmacol. 2021;56:7–12. https://doi.org/10.1016/j.coph.2020.08.007.
Nader K, Schafe GE, Le Doux JE. Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval. Nature. 2000;406(6797):722–6. https://doi.org/10.1038/35021052.
Nader K, Hardt O. A single standard for memory: the case for reconsolidation. Nat Rev Neurosci. 2009;10(3):224–34. https://doi.org/10.1038/nrn2590.
Xue YX, Luo YX, Wu P, Shi HS, Xue LF, Chen C, et al. A memory retrieval-extinction procedure to prevent drug craving and relapse. Science. 2012;336(6078):241–5. https://doi.org/10.1126/science.1215070.
Xue YX, Deng JH, Chen YY, Zhang LB, Wu P, Huang GD, et al. Effect of Selective Inhibition of Reactivated Nicotine-Associated Memories With Propranolol on Nicotine Craving. JAMA Psychiatry. 2017;74(3):224–32. https://doi.org/10.1001/jamapsychiatry.2016.3907.
Ma X, Zhang JJ, Yu LC. Post-retrieval extinction training enhances or hinders the extinction of morphine-induced conditioned place preference in rats dependent on the retrieval-extinction interval. Psychopharmacology (Berl). 2012;221(1):19–26. https://doi.org/10.1007/s00213-011-2545-4.
Suzuki A, Josselyn SA, Frankland PW, Masushige S, Silva AJ, Kida S. Memory reconsolidation and extinction have distinct temporal and biochemical signatures. J Neurosci. 2004;24(20):4787–95. https://doi.org/10.1523/JNEUROSCI.5491-03.2004.
Muravieva EV, Alberini CM. Limited efficacy of propranolol on the reconsolidation of fear memories. Learn Mem. 2010;17(6):306–13. https://doi.org/10.1101/lm.1794710.
Lee JL, Milton AL, Everitt BJ. Reconsolidation and extinction of conditioned fear: inhibition and potentiation. J Neurosci. 2006;26(39):10051–6. https://doi.org/10.1523/jneurosci.2466-06.2006.
Schiller D, Monfils MH, Raio CM, Johnson DC, Ledoux JE, Phelps EA. Preventing the return of fear in humans using reconsolidation update mechanisms. Nature. 2010;463(7277):49–53. https://doi.org/10.1038/nature08637.
Clem RL, Huganir RL. Calcium-permeable AMPA receptor dynamics mediate fear memory erasure. Science. 2010;330(6007):1108–12. https://doi.org/10.1126/science.1195298.
Luo YX, Xue YX, Liu JF, Shi HS, Jian M, Han Y, et al. A novel UCS memory retrieval-extinction procedure to inhibit relapse to drug seeking. Nat Commun. 2015;6:7675. https://doi.org/10.1038/ncomms8675.
Liu J, Zhao L, Xue Y, Shi J, Suo L, Luo Y, et al. An unconditioned stimulus retrieval extinction procedure to prevent the return of fear memory. Biol Psychiatry. 2014;76(11):895–901. https://doi.org/10.1016/j.biopsych.2014.03.027.
García-Pérez D, Ferenczi S, Kovács KJ, Laorden ML, Milanés MV, Núñez C. Glucocorticoid homeostasis in the dentate gyrus is essential for opiate withdrawal-associated memories. Mol Neurobiol. 2017;54(8):6523–41. https://doi.org/10.1007/s12035-016-0186-7.
Valero E, Gomez-Milanes I, Almela P, Ribeiro Do Couto B, Laorden ML, Milanes MV, et al. The involvement of CRF1 receptor within the basolateral amygdala and dentate gyrus in the naloxone-induced conditioned place aversion in morphine-dependent mice. Prog Neuropsychopharmacol Biol Psychiatry. 2018;84(Pt A):102–14. https://doi.org/10.1016/j.pnpbp.2018.01.018.
Ma Q, Fu Y, Cao Z, Shao D, Song J, Sheng H, et al. A conditioning-strengthened circuit from CA1 of dorsal hippocampus to basolateral amygdala participates in morphine-withdrawal memory retrieval. Front Neurosci. 2020;14:646. https://doi.org/10.3389/fnins.2020.00646.
Pereyra M, de Landeta AB, Dalto JF, Katche C, Medina JH. AMPA receptor expression requirement during long-term memory retrieval and its association with mTORC1 signaling. Mol Neurobiol. 2021;58(4):1711–22. https://doi.org/10.1007/s12035-020-02215-7.
Szapiro G, Barros DM, Ardenghi P, Vianna MR, Choi H, Silva T, et al. Facilitation and inhibition of retrieval in two aversive tasks in rats by intrahippocampal infusion of agonists of specific glutamate metabotropic receptor subtypes. Psychopharmacology (Berl). 2001;156(4):397–401. https://doi.org/10.1007/s002130100755.
Bonini JS, Bevilaqua LR, Zinn CG, Kerr DS, Medina JH, Izquierdo I, et al. Angiotensin II disrupts inhibitory avoidance memory retrieval. Horm Behav. 2006;50(2):308–13. https://doi.org/10.1016/j.yhbeh.2006.03.016.
Hsiung MH, Huang WL, Kan LY, Chen LH, Hu SS. The facilitating effect of MK-801 on inhibitory avoidance memory via mTOR signaling in the mouse hippocampus. Behav Brain Res. 2020;389:112630. https://doi.org/10.1016/j.bbr.2020.112630.
De Oliveira AL, Genro BP, Diehl F, Quillfeldt JA. Differential role of the hippocampal endocannabinoid system in the memory consolidation and retrieval mechanisms. Neurobiol Learn Mem. 2008;90(1):1–9. https://doi.org/10.1016/j.nlm.2008.01.009.
Khalaf O, Resch S, Dixsaut L, Gorden V, Glauser L, Graff J. Reactivation of recall-induced neurons contributes to remote fear memory attenuation. Science. 2018;360(6394):1239–42. https://doi.org/10.1126/science.aas9875.
Lim CS, Kim JI, Kwak C, Lee J, Jang EH, Oh J, et al. beta-Adrenergic signaling is required for the induction of a labile state during memory reconsolidation. Brain Res Bull. 2018;141:50–7. https://doi.org/10.1016/j.brainresbull.2018.04.011.
Nakazawa Y, Pevzner A, Tanaka KZ, Wiltgen BJ. Memory retrieval along the proximodistal axis of CA1. Hippocampus. 2016;26(9):1140–8. https://doi.org/10.1002/hipo.22596.
Khajehpour L, Rezayof A, Zarrindast MR. Involvement of dorsal hippocampal nicotinic receptors in the effect of morphine on memory retrieval in passive avoidance task. Eur J Pharmacol. 2008;584(2-3):343–51. https://doi.org/10.1016/j.ejphar.2008.02.030.
Flasbeck V, Atucha E, Nakamura NH, Yoshida M, Sauvage MM. Spatial information is preferentially processed by the distal part of CA3: Implication for memory retrieval. Behav Brain Res. 2018;354:31–8. https://doi.org/10.1016/j.bbr.2018.07.023.
Josselyn SA, Tonegawa S. Memory engrams: recalling the past and imagining the future. Science. 2020;367(6473). https://doi.org/10.1126/science.aaw4325.
Hirsh R. The hippocampus and contextual retrieval of information from memory: a theory. Behav Biol. 1974;12(4):421–44. https://doi.org/10.1016/s0091-6773(74)92231-7.
Squire LR, Clark RE, Knowlton BJ. Retrograde amnesia. Hippocampus. 2001;11(1):50–5. https://doi.org/10.1002/1098-1063(2001)11:1<50::aid-hipo1019>3.0.co;2-g.
Ramirez S, Liu X, Lin PA, Suh J, Pignatelli M, Redondo RL, et al. Creating a false memory in the hippocampus. Science. 2013;341(6144):387–91. https://doi.org/10.1126/science.1239073.
Denny CA, Kheirbek MA, Alba EL, Tanaka KF, Brachman RA, Laughman KB, et al. Hippocampal memory traces are differentially modulated by experience, time, and adult neurogenesis. Neuron. 2014;83(1):189–201. https://doi.org/10.1016/j.neuron.2014.05.018.
Murchison CF, Zhang XY, Zhang WP, Ouyang M, Lee A, Thomas SA. A distinct role for norepinephrine in memory retrieval. Cell. 2004;117(1):131–43. https://doi.org/10.1016/s0092-8674(04)00259-4.
Zamorano C, Fernandez-Albert J, Storm DR, Carne X, Sindreu C. Memory Retrieval Re-Activates Erk1/2 Signaling in the Same Set of CA1 Neurons Recruited During Conditioning. Neuroscience. 2018;370:101–11. https://doi.org/10.1016/j.neuroscience.2017.03.034.
Rashid H, Ahmed T. Muscarinic activity in hippocampus and entorhinal cortex is crucial for spatial and fear memory retrieval. Pharmacol Rep. 2019;71(3):449–56. https://doi.org/10.1016/j.pharep.2019.02.004.
Wilmot JH, Puhger K, Wiltgen BJ. Acute Disruption of the Dorsal Hippocampus Impairs the Encoding and Retrieval of Trace Fear Memories. Front Behav Neurosci. 2019;13:116. https://doi.org/10.3389/fnbeh.2019.00116.
Hunsaker MR, Kesner RP. Dissociations across the dorsal-ventral axis of CA3 and CA1 for encoding and retrieval of contextual and auditory-cued fear. Neurobiol Learn Mem. 2008;89(1):61–9. https://doi.org/10.1016/j.nlm.2007.08.016.
Bernier BE, Lacagnina AF, Ayoub A, Shue F, Zemelman BV, Krasne FB, et al. Dentate Gyrus Contributes to Retrieval as well as Encoding: Evidence from Context Fear Conditioning, Recall, and Extinction. J Neurosci. 2017;37(26):6359–71. https://doi.org/10.1523/JNEUROSCI.3029-16.2017.
Pignatelli M, Ryan TJ, Roy DS, Lovett C, Smith LM, Muralidhar S, et al. Engram Cell Excitability State Determines the Efficacy of Memory Retrieval. Neuron. 2019;101(2):274–84 e5. https://doi.org/10.1016/j.neuron.2018.11.029.
Perusini JN, Cajigas SA, Cohensedgh O, Lim SC, Pavlova IP, Donaldson ZR, et al. Optogenetic stimulation of dentate gyrus engrams restores memory in Alzheimer's disease mice. Hippocampus. 2017;27(10):1110–22. https://doi.org/10.1002/hipo.22756.
Cho HY, Kim M, Han JH. Specific disruption of contextual memory recall by sparse additional activity in the dentate gyrus. Neurobiol Learn Mem. 2017;145:190–8. https://doi.org/10.1016/j.nlm.2017.10.006.
Garcia-Perez D, Milanes MV. Role of glucocorticoids on noradrenergic and dopaminergic neurotransmission within the basolateral amygdala and dentate gyrus during morphine withdrawal place aversion. Addict Biol. 2019. https://doi.org/10.1111/adb.12728.
Colon LM, Poulos AM. Contextual processing elicits sex differences in dorsal hippocampus activation following footshock and context fear retrieval. Behav Brain Res. 2020;393:112771. https://doi.org/10.1016/j.bbr.2020.112771.
Keiser AA, Turnbull LM, Darian MA, Feldman DE, Song I, Tronson NC. Sex Differences in Context Fear Generalization and Recruitment of Hippocampus and Amygdala during Retrieval. Neuropsychopharmacology. 2017;42(2):397–407. https://doi.org/10.1038/npp.2016.174.
Brito I, Britto LRG, Ferrari EAM. Retrieval of contextual aversive memory and induction of Zenk expression in the hippocampus of pigeons. Brain Res Bull. 2019;153:341–9. https://doi.org/10.1016/j.brainresbull.2019.09.013.
Couto-Pereira NS, Lampert C, Vieira ADS, Lazzaretti C, Kincheski GC, Espejo PJ, et al. Resilience and vulnerability to trauma: early life interventions modulate aversive memory reconsolidation in the dorsal hippocampus. Front Mol Neurosci. 2019;12:134. https://doi.org/10.3389/fnmol.2019.00134.
Popik B, Crestani AP, Silva MO, Quillfeldt JA, de Oliveira Alvares L. Calpain modulates fear memory consolidation, retrieval and reconsolidation in the hippocampus. Neurobiol Learn Mem. 2018;151:53–8. https://doi.org/10.1016/j.nlm.2018.04.002.
Corcoran KA, Desmond TJ, Frey KA, Maren S. Hippocampal inactivation disrupts the acquisition and contextual encoding of fear extinction. J Neurosci. 2005;25(39):8978–87. https://doi.org/10.1523/jneurosci.2246-05.2005.
Gobira PH, Lima IV, Batista LA, de Oliveira AC, Resstel LB, Wotjak CT, et al. N-arachidonoyl-serotonin, a dual FAAH and TRPV1 blocker, inhibits the retrieval of contextual fear memory: Role of the cannabinoid CB1 receptor in the dorsal hippocampus. J Psychopharmacol. 2017;31(6):750–6. https://doi.org/10.1177/0269881117691567.
Jimenez JC, Berry JE, Lim SC, Ong SK, Kheirbek MA, Hen R. Contextual fear memory retrieval by correlated ensembles of ventral CA1 neurons. Nat Commun. 2020;11(1):3492. https://doi.org/10.1038/s41467-020-17270-w.
Xu C, Krabbe S, Gründemann J, Botta P, Fadok JP, Osakada F, et al. Distinct hippocampal pathways mediate dissociable roles of context in memory retrieval. Cell. 2016;167(4):961–72.e16. https://doi.org/10.1016/j.cell.2016.09.051.
Wang D, Huang Z, Ren L, Liu J, Wang X, Yu T, et al. Amygdalar and hippocampal beta rhythm synchrony during human fear memory retrieval. Acta Neurochir (Wien). 2020;162(10):2499–507. https://doi.org/10.1007/s00701-020-04276-y.
Korz V, Frey JU. Hormonal and monoamine signaling during reinforcement of hippocampal long-term potentiation and memory retrieval. Learn Mem. 2007;14(3):160–6. https://doi.org/10.1101/lm.459807.
Otis JM, Fitzgerald MK, Mueller D. Inhibition of hippocampal beta-adrenergic receptors impairs retrieval but not reconsolidation of cocaine-associated memory and prevents subsequent reinstatement. Neuropsychopharmacology. 2014;39(2):303–10. https://doi.org/10.1038/npp.2013.187.
Qi XL, Zhu B, Zhang XH, Li BM. Are beta-adrenergic receptors in the hippocampal CA1 region required for retrieval of contextual fear memory? Biochem Biophys Res Commun. 2008;368(2):186–91. https://doi.org/10.1016/j.bbrc.2008.01.007.
Murchison CF, Schutsky K, Jin SH, Thomas SA. Norepinephrine and ß1-adrenergic signaling facilitate activation of hippocampal CA1 pyramidal neurons during contextual memory retrieval. Neuroscience. 2011;181:109–16. https://doi.org/10.1016/j.neuroscience.2011.02.049.
Schutsky K, Ouyang M, Thomas SA. Xamoterol impairs hippocampus-dependent emotional memory retrieval via Gi/o-coupled β2-adrenergic signaling. Learn Mem. 2011;18(9):598–604. https://doi.org/10.1101/lm.2302811.
Schutsky K, Ouyang M, Castelino CB, Zhang L, Thomas SA. Stress and glucocorticoids impair memory retrieval via β2-adrenergic, Gi/o-coupled suppression of cAMP signaling. J Neurosci. 2011;31(40):14172–81. https://doi.org/10.1523/jneurosci.2122-11.2011.
Atsak P, Hauer D, Campolongo P, Schelling G, McGaugh JL, Roozendaal B. Glucocorticoids interact with the hippocampal endocannabinoid system in impairing retrieval of contextual fear memory. Proc Natl Acad Sci U S A. 2012;109(9):3504–9. https://doi.org/10.1073/pnas.1200742109.
Zarrindast MR, Mashayekhi M, Rezayof A, Ahmadi S. β-Adrenoceptors in the dorsal hippocampus are involved in ethanol-induced state-dependent retrieval in mice. Neurobiol Learn Mem. 2013;100:12–7. https://doi.org/10.1016/j.nlm.2012.11.004.
Roozendaal B, Hahn EL, Nathan SV, de Quervain DJ, McGaugh JL. Glucocorticoid effects on memory retrieval require concurrent noradrenergic activity in the hippocampus and basolateral amygdala. J Neurosci. 2004;24(37):8161–9. https://doi.org/10.1523/jneurosci.2574-04.2004.
Strange BA, Dolan RJ. Beta-adrenergic modulation of emotional memory-evoked human amygdala and hippocampal responses. Proc Natl Acad Sci U S A. 2004;101(31):11454–8. https://doi.org/10.1073/pnas.0404282101.
Fukabori R, Iguchi Y, Kato S, Takahashi K, Eifuku S, Tsuji S, et al. Enhanced retrieval of taste associative memory by chemogenetic activation of locus coeruleus norepinephrine neurons. J Neurosci. 2020;40(43):8367–85. https://doi.org/10.1523/jneurosci.1720-20.2020.
Sterpenich V, D'Argembeau A, Desseilles M, Balteau E, Albouy G, Vandewalle G, et al. The locus ceruleus is involved in the successful retrieval of emotional memories in humans. J Neurosci. 2006;26(28):7416–23. https://doi.org/10.1523/JNEUROSCI.1001-06.2006.
Sara SJ, Devauges V. Idazoxan, an alpha-2 antagonist, facilitates memory retrieval in the rat. Behav Neural Biol. 1989;51(3):401–11. https://doi.org/10.1016/s0163-1047(89)91039-x.
Sampaio TB, Marques NF, Binder LB, Tasca CI, Prediger RD. Role of prefrontal cortex on recognition memory deficits in rats following 6-OHDA-induced locus coeruleus lesion. Oxid Med Cell Longev. 2020;2020:8324565. https://doi.org/10.1155/2020/8324565.
Shelkar GP, Gakare SG, Chakraborty S, Dravid SM, Ugale RR. Interactions of nitric oxide with α2 -adrenoceptors within the locus coeruleus underlie the facilitation of inhibitory avoidance memory by agmatine. Br J Pharmacol. 2016;173(17):2589–99. https://doi.org/10.1111/bph.13531.
Reyes-López J, Nuñez-Jaramillo L, Morán-Guel E, Miranda MI. Differential effects of beta-adrenergic receptor blockade in the medial prefrontal cortex during aversive and incidental taste memory formation. Neuroscience. 2010;169(1):195–202. https://doi.org/10.1016/j.neuroscience.2010.04.054.
Osorio-Gómez D, Guzmán-Ramos K, Bermúdez-Rattoni F. Memory trace reactivation and behavioral response during retrieval are differentially modulated by amygdalar glutamate receptors activity: interaction between amygdala and insular cortex. Learn Mem. 2017;24(1):14–23. https://doi.org/10.1101/lm.042895.116.
Barros DM, e Souza TM, De David T, Choi H, Aguzzoli A, Madche C, et al. Simultaneous modulation of retrieval by dopaminergic D(1), beta-noradrenergic, serotonergic-1A and cholinergic muscarinic receptors in cortical structures of the rat. Behav Brain Res. 2001;124(1):1–7. https://doi.org/10.1016/s0166-4328(01)00208-x.
Osorio-Gómez D, Guzmán-Ramos K, Bermúdez-Rattoni F. Differential involvement of glutamatergic and catecholaminergic activity within the amygdala during taste aversion retrieval on memory expression and updating. Behav Brain Res. 2016;307:120–5. https://doi.org/10.1016/j.bbr.2016.03.038.
Bernardi RE, Ryabinin AE, Berger SP, Lattal KM. Post-retrieval disruption of a cocaine conditioned place preference by systemic and intrabasolateral amygdala beta2- and alpha1-adrenergic antagonists. Learn Mem. 2009;16(12):777–89. https://doi.org/10.1101/lm.1648509.
Smith CC, Greene RW. CNS dopamine transmission mediated by noradrenergic innervation. J Neurosci. 2012;32(18):6072–80. https://doi.org/10.1523/jneurosci.6486-11.2012.
Klukowski G, Harley CW. Locus coeruleus activation induces perforant path-evoked population spike potentiation in the dentate gyrus of awake rat. Exp Brain Res. 1994;102(1):165–70. https://doi.org/10.1007/bf00232449.
Hu H, Real E, Takamiya K, Kang MG, Ledoux J, Huganir RL, et al. Emotion enhances learning via norepinephrine regulation of AMPA-receptor trafficking. Cell. 2007;131(1):160–73. https://doi.org/10.1016/j.cell.2007.09.017.
Yavich L, Jakala P, Tanila H. Noradrenaline overflow in mouse dentate gyrus following locus coeruleus and natural stimulation: real-time monitoring by in vivo voltammetry. J Neurochem. 2005;95(3):641–50. https://doi.org/10.1111/j.1471-4159.2005.03390.x.
Murchison CF, Schutsky K, Jin SH, Thomas SA. Norepinephrine and ss (1)-adrenergic signaling facilitate activation of hippocampal CA1 pyramidal neurons during contextual memory retrieval. Neuroscience. 2011;181:109–16. https://doi.org/10.1016/j.neuroscience.2011.02.049.
Zhu Y, Kimelberg HK. Cellular expression of P2Y and beta-AR receptor mRNAs and proteins in freshly isolated astrocytes and tissue sections from the CA1 region of P8-12 rat hippocampus. Brain Res Dev Brain Res. 2004;148(1):77–87. https://doi.org/10.1016/j.devbrainres.2003.10.014.
Fellin T, Pascual O, Gobbo S, Pozzan T, Haydon PG, Carmignoto G. Neuronal synchrony mediated by astrocytic glutamate through activation of extrasynaptic NMDA receptors. Neuron. 2004;42(5):729–43. https://doi.org/10.1016/j.neuron.2004.08.011.
Zhuo M. Plasticity of NMDA receptor NR2B subunit in memory and chronic pain. Mol Brain. 2009;2(4). https://doi.org/10.1186/1756-6606-2-4.
Zhang XH, Wu LJ, Gong B, Ren M, Li BM, Zhuo M. Induction- and conditioning-protocol dependent involvement of NR2B-containing NMDA receptors in synaptic potentiation and contextual fear memory in the hippocampal CA1 region of rats. Mol Brain. 2008;1(9). https://doi.org/10.1186/1756-6606-1-9.
Frenois F, Cador M, Caille S, Stinus L, Le Moine C. Neural correlates of the motivational and somatic components of naloxone-precipitated morphine withdrawal. Eur J Neurosci. 2002;16(7):1377–89. https://doi.org/10.1046/j.1460-9568.2002.02187.x.
Caillé S, Espejo EF, Reneric JP, Cador M, Koob GF, Stinus L. Total neurochemical lesion of noradrenergic neurons of the locus ceruleus does not alter either naloxone-precipitated or spontaneous opiate withdrawal nor does it influence ability of clonidine to reverse opiate withdrawal. J Pharmacol Exp Ther. 1999;290(2):881–92.
Aghajanian GK. Feedback regulation of central monoaminergic neurons: evidence from single cell recording studies. Essays Neurochem Neuropharmacol. 1978;3:1–32.
Maldonado R. Participation of noradrenergic pathways in the expression of opiate withdrawal: biochemical and pharmacological evidence. Neurosci Biobehav Rev. 1997;21(1):91–104. https://doi.org/10.1016/0149-7634(95)00061-5.
Menezes J, Alves N, Borges S, Roehrs R, de Carvalho MJ, Furini CR, et al. Facilitation of fear extinction by novelty depends on dopamine acting on D1-subtype dopamine receptors in hippocampus. Proc Natl Acad Sci U S A. 2015;112(13):E1652–8. https://doi.org/10.1073/pnas.1502295112.
Liu JF, Yang C, Deng JH, Yan W, Wang HM, Luo YX, et al. Role of hippocampal beta-adrenergic and glucocorticoid receptors in the novelty-induced enhancement of fear extinction. J Neurosci. 2015;35(21):8308–21. https://doi.org/10.1523/jneurosci.0005-15.2015.
Swanson LW. The projections of the ventral tegmental area and adjacent regions: a combined fluorescent retrograde tracer and immunofluorescence study in the rat. Brain Res Bull. 1982;9(1-6):321–53. https://doi.org/10.1016/0361-9230(82)90145-9.
Gasbarri A, Packard MG, Campana E, Pacitti C. Anterograde and retrograde tracing of projections from the ventral tegmental area to the hippocampal formation in the rat. Brain Res Bull. 1994;33(4):445–52. https://doi.org/10.1016/0361-9230(94)90288-7.
Gasbarri A, Sulli A, Packard MG. The dopaminergic mesencephalic projections to the hippocampal formation in the rat. Prog Neuropsychopharmacol Biol Psychiatry. 1997;21(1):1–22. https://doi.org/10.1016/s0278-5846(96)00157-1.
O'Carroll CM, Martin SJ, Sandin J, Frenguelli B, Morris RG. Dopaminergic modulation of the persistence of one-trial hippocampus-dependent memory. Learn Mem. 2006;13(6):760–9. https://doi.org/10.1101/lm.321006.
Bethus I, Tse D, Morris RG. Dopamine and memory: modulation of the persistence of memory for novel hippocampal NMDA receptor-dependent paired associates. J Neurosci. 2010;30(5):1610–8. https://doi.org/10.1523/jneurosci.2721-09.2010.
Kempadoo KA, Mosharov EV, Choi SJ, Sulzer D, Kandel ER. Dopamine release from the locus coeruleus to the dorsal hippocampus promotes spatial learning and memory. Proc Natl Acad Sci U S A. 2016;113(51):14835–40. https://doi.org/10.1073/pnas.1616515114.
Reijmers LG, Perkins BL, Matsuo N, Mayford M. Localization of a stable neural correlate of associative memory. Science. 2007;317(5842):1230–3. https://doi.org/10.1126/science.1143839.
Herry C, Ciocchi S, Senn V, Demmou L, Müller C, Lüthi A. Switching on and off fear by distinct neuronal circuits. Nature. 2008;454(7204):600–6. https://doi.org/10.1038/nature07166.
Herry C, Johansen JP. Encoding of fear learning and memory in distributed neuronal circuits. Nat Neurosci. 2014;17(12):1644–54. https://doi.org/10.1038/nn.3869.
Mason ST, Fibiger HC. Regional topography within noradrenergic locus coeruleus as revealed by retrograde transport of horseradish peroxidase. J Comp Neurol. 1979;187(4):703–24. https://doi.org/10.1002/cne.901870405.
Loughlin SE, Foote SL, Grzanna R. Efferent projections of nucleus locus coeruleus: morphologic subpopulations have different efferent targets. Neuroscience. 1986;18(2):307–19. https://doi.org/10.1016/0306-4522(86)90156-9.
Berridge CW, Waterhouse BD. The locus coeruleus-noradrenergic system: modulation of behavioral state and state-dependent cognitive processes. Brain Res Brain Res Rev. 2003;42(1):33–84. https://doi.org/10.1016/s0165-0173(03)00143-7.
Bechara A, Tranel D, Damasio H, Adolphs R, Rockland C, Damasio AR. Double dissociation of conditioning and declarative knowledge relative to the amygdala and hippocampus in humans. Science. 1995;269(5227):1115–8. https://doi.org/10.1126/science.7652558.
Van Bockstaele EJ, Colago EE, Aicher S. Light and electron microscopic evidence for topographic and monosynaptic projections from neurons in the ventral medulla to noradrenergic dendrites in the rat locus coeruleus. Brain Res. 1998;784(1-2):123–38. https://doi.org/10.1016/s0006-8993(97)01250-x.
Van Bockstaele EJ, Chan J, Pickel VM. Input from central nucleus of the amygdala efferents to pericoerulear dendrites, some of which contain tyrosine hydroxylase immunoreactivity. J Neurosci Res. 1996;45(3):289–302. https://doi.org/10.1002/(sici)1097-4547(19960801)45:3<289::Aid-jnr11>3.0.Co;2-#.
Ivanov A, Aston-Jones G. Local opiate withdrawal in locus coeruleus neurons in vitro. J Neurophysiol. 2001;85(6):2388–97. https://doi.org/10.1152/jn.2001.85.6.2388.
Hansen N. The longevity of hippocampus-dependent memory is orchestrated by the locus coeruleus-noradrenergic system. Neural Plast. 2017;2017:2727602. https://doi.org/10.1155/2017/2727602.
DeNardo LA, Liu CD, Allen WE, Adams EL, Friedmann D, Fu L, et al. Temporal evolution of cortical ensembles promoting remote memory retrieval. Nat Neurosci. 2019;22(3):460–9. https://doi.org/10.1038/s41593-018-0318-7.
Barry DN, Maguire EA. Remote memory and the hippocampus: a constructive critique. Trends Cogn Sci. 2019;23(2):128–42. https://doi.org/10.1016/j.tics.2018.11.005.
Neher T, Cheng S, Wiskott L. Memory storage fidelity in the hippocampal circuit: the role of subregions and input statistics. Plos Comput Biol. 2015;11(5):e1004250. https://doi.org/10.1371/journal.pcbi.1004250.
Bouton ME. Context and behavioral processes in extinction. Learn Mem. 2004;11(5):485–94. https://doi.org/10.1101/lm.78804.
Myers KM, Davis M. Mechanisms of fear extinction. Mol Psychiatry. 2007;12(2):120–50. https://doi.org/10.1038/sj.mp.4001939.
Tsuboyama M, Kaye HL, Rotenberg A. Review of transcranial magnetic stimulation in epilepsy. Clin Ther. 2020;42(7):1155–68. https://doi.org/10.1016/j.clinthera.2020.05.016.
Khedr EM, Lefaucheur JP, Hasan AM, Osama K. Are there differences in cortical excitability between akinetic-rigid and tremor-dominant subtypes of Parkinson's disease? Neurophysiol Clin. 2021. https://doi.org/10.1016/j.neucli.2021.08.002.
Adams TG, Cisler JM, Kelmendi B, George JR, Kichuk SA, Averill CL, et al. Transcranial direct current stimulation targeting the medial prefrontal cortex modulates functional connectivity and enhances safety learning in obsessive-compulsive disorder: Results from two pilot studies. Depress Anxiety. 2021. https://doi.org/10.1002/da.23212.
Bréchet L, Michel CM, Schacter DL, Pascual-Leone A. Improving autobiographical memory in Alzheimer's disease by transcranial alternating current stimulation. Curr Opin Behav Sci. 2021;40:64–71. https://doi.org/10.1016/j.cobeha.2021.01.003.
Grossman N, Bono D, Dedic N, Kodandaramaiah SB, Rudenko A, Suk HJ, et al. Noninvasive deep brain stimulation via temporally interfering electric fields. Cell. 2017;169(6):1029–41.e16. https://doi.org/10.1016/j.cell.2017.05.024.
Higgins C, Liu Y, Vidaurre D, Kurth-Nelson Z, Dolan R, Behrens T, et al. Replay bursts in humans coincide with activation of the default mode and parietal alpha networks. Neuron. 2021;109(5):882-93.e7. https://doi.org/10.1016/j.neuron.2020.12.007.
Rohan JG, Miklasevich MK, McInturf SM, Bechmann NA, Moore RJ, Hatcher-Solis C, et al. Polarity and subfield specific effects of transcranial direct current stimulation on hippocampal plasticity. Neurobiol Learn Mem. 2020;167:107126. https://doi.org/10.1016/j.nlm.2019.107126.
Alqahtani F, Imran I, Pervaiz H, Ashraf W, Perveen N, Rasool MF, et al. Non-pharmacological interventions for intractable epilepsy. Saudi Pharm J. 2020;28(8):951–62. https://doi.org/10.1016/j.jsps.2020.06.016.
Takeuchi T, Duszkiewicz AJ, Sonneborn A, Spooner PA, Yamasaki M, Watanabe M, et al. Locus coeruleus and dopaminergic consolidation of everyday memory. Nature. 2016;537(7620):357–62. https://doi.org/10.1038/nature19325.
Matthews KL, Chen CP, Esiri MM, Keene J, Minger SL, Francis PT. Noradrenergic changes, aggressive behavior, and cognition in patients with dementia. Biol Psychiatry. 2002;51(5):407–16. https://doi.org/10.1016/s0006-3223(01)01235-5.
Chai GS, Wang YY, Yasheng A, Zhao P. Beta 2-adrenergic receptor activation enhances neurogenesis in Alzheimer's disease mice. Neural Regen Res. 2016;11(10):1617–24. https://doi.org/10.4103/1673-5374.193241.
Debiec J, Doyère V, Nader K, Ledoux JE. Directly reactivated, but not indirectly reactivated, memories undergo reconsolidation in the amygdala. Proc Natl Acad Sci U S A. 2006;103(9):3428–33. https://doi.org/10.1073/pnas.0507168103.
Assar N, Mahmoudi D, Farhoudian A, Farhadi MH, Fatahi Z, Haghparast A. D1- and D2-like dopamine receptors in the CA1 region of the hippocampus are involved in the acquisition and reinstatement of morphine-induced conditioned place preference. Behav Brain Res. 2016;312:394–404. https://doi.org/10.1016/j.bbr.2016.06.061.
Zhang JJ, Han J, Sui N. Okadaic acid blocks the effects of 5-aza-2-deoxycytidine on consolidation, acquisition and retrieval of morphine-induced place preference in rats. Neuropharmacology. 2014;86:282–93. https://doi.org/10.1016/j.neuropharm.2014.08.005.
Katebi N, Farahimanesh S, Fatahi Z, Zarrabian S, Haghparast A. Involvement of D1- and D2-like dopamine receptors in the dentate gyrus in the acquisition, expression, and extinction of the morphine-induced conditioned place preference in rats. Behav Brain Res. 2018;353:185–93. https://doi.org/10.1016/j.bbr.2018.07.018.
Corlett PR, Krystal JH, Taylor JR, Fletcher PC. Why do delusions persist? Front Hum Neurosci. 2009;3:12. https://doi.org/10.3389/neuro.09.012.2009.
Acknowledgements
This work was supported by National Natural Science Foundation of China (32071028), Natural Science Foundation of Beijing Municipality (5202023), CAS-VPST Silk Road Science Fund 2021 (GJHZ202129), and CAS Key Laboratory of Mental Health, Institute of Psychology.
Human and animal rights and informed consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Author information
Authors and Affiliations
Contributions
The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing of interest
The authors declare no conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Neuroscience & Addiction
Rights and permissions
About this article

Cite this article
Dai, Zh., Xu, X., Chen, Wq. et al. The role of hippocampus in memory reactivation: an implication for a therapeutic target against opioid use disorder. Curr Addict Rep 9, 67–79 (2022). https://doi.org/10.1007/s40429-022-00407-w
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40429-022-00407-w