
Abstract
Ligand-binding properties of β-lactoglobulin (β-lg) are well documented, but the subsequent biological functions are still unclear. Focusing on fatty acids/β-lg complexes, the structure-function relationships are reviewed in the light of the structural state of the protein (native versus non-native aggregated proteins). After a brief description of β-lg native structure, the review takes an interest in the binding properties of native β-lg (localization of binding sites, stoichiometry, and affinity) and the way the interaction affects the biological properties of the protein and the ligand. The binding properties of non-native aggregated forms of β-lg that are classically generated during industrial processing are also related. Structural changes modify the stoichiometry and the affinity of β-lg for fatty acids and consequently the biological functions of the complex. Finally, the fatty acid-binding properties of other whey proteins (α-lactalbumin, bovine serum albumin) and some biological properties of the complexes are also addressed. These proteins affect β-lg/fatty acids complex in whey given their competition with β-lg for fatty acids.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Milk is a complete food for the mammalian neonate, supplying carbohydrates, fatty acids (FA), vitamins, inorganic elements, and proteins (Jensen 1995). Bovine milk proteins (average concentration of 34 g.L) are divided into casein and whey protein fractions, which represent about 80 and 20% of the whole milk proteins, respectively. The major whey proteins are β-lactoglobulin (β-lg), α-lactalbumin (α-la), and bovine serum albumin (BSA). The amino acid composition of whey proteins (high proportion of cysteine, tryptophan, and branched amino acids), their ability to bind various ligands, and the presence of bioactive peptides within their sequence, give them excellent nutritional properties. Additionally, whey proteins have important techno-functional properties in food systems such as their ability to act as emulsifiers, gelling, and texturing agents (Smithers 2008).
β-lg, the major whey protein in bovine milk, is an extensively studied protein and is known to bind hydrophobic ligands such as FA or vitamins. Nevertheless, beyond the nutritional contribution of the individual components (β-lg and ligands), the biological functions of the complex protein/ligand are still speculative. Putative roles would be (1) increase of FA absorption (Perez et al. 1992), (2) modification of the kinetics of the enzymatic hydrolysis of the protein (Puyol et al. 1993; Mandalari et al. 2009), (3) protection of sensitive ligands against oxidation (Futterman and Heller 1972) or other stresses, and (4) modification of the bioaccessibility of the ligands (Puyol et al. 1995; Riihimäki-Lampén 2009). Additionally, in food products, the binding properties and consequently the biological properties of β-lg/ligand complexes may be affected by the structure of β-lg, and/or the presence of other proteins capable of competing with β-lg for ligand binding. Information regarding the interactions of non-native species of β-lg with hydrophobic ligands and the competition between β-lg and other proteins for hydrophobic ligands are poorly described in the literature. Improving our knowledge of the impact of interactions between nutrients in food products and their biological functions will help to design food products with optimized nutritional properties. It will also support a more detailed nutritional picture of milk and milk products.
The aim of this review is to map the current knowledge on the binding of native and non-native forms of β-lg with hydrophobic ligands, focusing on FA, as well as the subsequent biological properties of the complexes when available. Since β-lg is often in mixture with other whey proteins in food products, current knowledge on the binding of FA to other whey proteins (α-la, BSA) is also considered.
2 β-lactoglobulin structure
β-Lg is present in the milk of ruminants and non-ruminants such as pig, horse, dog, cat, dolphin, and marsupials (Pérez et al. 1989; Sawyer and Kontopidis 2000). Interestingly, it is absent in human, lagomorph, and rodent milks (Sawyer and Kontopidis 2000). With a concentration of 3.2 g.L in mature bovine milk, β-lg accounts for about 10% of the total milk proteins and about 50–60% of the total whey proteins. There are several genetic variants of β-lg, with the A and B variants being the most common (Godovac-Zimmermann et al. 1996). These two variants both contain 162 amino acids, but they differ by two amino acids at positions 64 and 118. Variant A has an aspartic acid residue at position 64 and a valine residue at position 118, while variant B has glycine and alanine in these positions, respectively. Both variants contain five cysteine (Cys) residues, located at positions 66, 106, 119, 121, and 160. These cysteines form two disulfide bonds, between Cys66 and Cys160, and between Cys106 and Cys119 (Papiz et al. 1986; Brownlow et al. 1997). Cys121 is a free thiol that lies buried in the center of β-lg structure (Qin et al. 1998; Burova et al. 1998) and participates to the stability of the native protein (Barbiroli et al. 2011; Croguennec et al. 2004; Jayat et al. 2004).
The secondary structure of the β-lg is composed of 15% α-helix, 50% β-sheet, and 15–20% reverse turn (Creamer et al. 1983; Sawyer and Kontopidis 2000). The nine β-strands labelled from A to I, which form two β-sheets, and the three turns α-helix are arranged to form the β-lg globular structure. Tertiary structure of the β-lg is shown in Fig. 1. In aqueous solution and at neutral pH, the β-sheets form a flattened and conical barrel, called a calyx (Papiz et al. 1986; Brownlow et al. 1997). This barrel has its two β-sheets connected by strand A on one side, while a secondary connection is formed between strands D and E. The α-helix is entrapped between the strands A and H and is followed by the ninth β-strand called I (Sawyer and Kontopidis 2000). In the native protein, disulfide bonds link strands G to H (Cys106–Cys119) and strand D to the C-terminal (Cys66–Cys160). The free thiol group is inaccessible to solvents in the native protein structure and consequently is unavailable for reactions under physiological conditions (Qin et al. 1998; Burova et al. 1998). The calyx is closed at one end by the N-terminal loop and can be closed at the other by the EF loop in low pH conditions. β-lg belongs to the lipocalin protein family, which typically contains a β-barrel, inside which small hydrophobic molecules can be found (Flower 1996). Variants A and B of β-lg have similar tertiary structure at neutral pH as the amino acids that differ between the two variants are located on a mobile surface loop and in the hydrophobic core (Sawyer and Kontopidis 2000; Qin et al. 1999). Under physiological conditions, β-lg is in equilibrium between monomers and non-covalent dimers. Protein concentration, pH, ionic strength, and temperature affect this equilibrium and consequently the proportion of monomers and non-covalent dimers in solution (Aymard et al. 1996; Renard et al. 1998; Verheul et al. 1999; Mercadante et al. 2012).

A schematic view of the main-chain fold of bovine β-lg in interaction with linoleic acid in its central cavity. The graphic was performed with the UCSF Chimera package. Chimera is developed by the Resource for Biocomputing, Visualization, and Informatics at the University of California, San Francisco (supported by NIGMS P41-GM103311, http://www.cgl.ucsf.edu/chimera). RCSB PDB code 4DQ4 (Loch et al. 2012b)
Despite structural similarities between variants A and B, several differences in physical and chemical properties were reported; it concerns difference in pI (Yan et al 2013), stability of native dimers (Mercadante et al 2012), thermal denaturation temperature (Manderson et al 1999) and denaturation reaction rate (O’Kennedy et al 2006), susceptibility to chemicals (Bouhallab et al. 2004; Boye et al. 2004), and affinity for FA (Loch et al. 2013b).
3 β-lactoglobulin/ligand-binding properties
In 1949, McMeekin et al. (1949) described the binding of sodium dodecyl sulfate (SDS) to native β-lg. Since then, native β-lg has been demonstrated to bind numerous hydrophobic ligands such as retinol, vitamin D, cholesterol, curcumin, FA and their derivatives, and polycycles such as protoporphyrine IX, aromatic compounds, catechin, and cations (Lišková et al. 2011; Le Maux et al. 2012; Puyol et al. 1994; Smith et al. 1983; O'Neill and Kinsella 1988; Futterman and Heller 1972; Patocka and Jelen 1991; Wang et al. 1997; Sneharani et al. 2010; Kanakis et al. 2011; Dufour et al. 1990, 1992; Liu et al. 2011). FA are carboxylic acids with hydrocarbon chains varying from 4 to 30 carbon units. In general, FA are poorly soluble in water. FA solubility decreases with an increase in chain length. This is illustrated by the solubility of caprylic acid (C12:0) and stearic acid (C18:0) at 20 °C, which are 0.08 g/100 g water and 0.0003 g/100 g water, respectively (Lide 2003). The chain length increases the van der Waals interactions between two adjacent FA bringing them closer to each other (Tanford 1980). Moreover, the solubility of FA of similar length chain increases with the number of C = C double bonds in the aliphatic chain because the curvature of the FA increases the distance between FA (Beneventi et al. 2001). At pH value above the pKa of the carboxylic group, FA are more soluble. In aqueous environments, their hydrophilic head allows the FA to be stabilized by organization into micelles and bilayers. Above the critical micelle concentration (CMC) and below the critical micelle temperature, FA are in equilibrium between micelles and free FA. Such equilibrium and FA solubility affect their interaction and binding to receptors such as proteins.
3.1 Localization of hydrophobic ligand-binding sites
Native β-lg has one main well-defined binding site per monomer, and depending on the ligand and medium conditions, one or more secondary binding sites were reported. The main binding site of native β-lg for hydrophobic ligands is formed by the calyx of the protein as shown in Fig. 1 (Wu et al. 1999; Kontopidis et al. 2004; Ragona et al. 2000). Crystallographic study clearly showed the binding of palmitate in the calyx of β-lg (Wu et al. 1999). The complex is stabilized by hydrophobic interactions in the calyx and hydrogen bonding involving the FA carboxylic group and β-lg Glu62 and Lys69 at the entrance of the calyx (Loch et al. 2013a). Several studies have also indicated a secondary binding site on the protein monomer. β-lg can simultaneously bind two different ligands at two different sites as shown for retinol and protoporphyrine IX (Dufour et al. 1990) as well as for a mixture of palmitic acid and retinoid (Narayan and Berliner 1998). However, Puyol et al. (1991) found that palmitic acid and retinol have the same binding site. In order to identify the competitive binding of two ligands, these studies determined the difference of binding constants in the presence of one or two ligands. However, these two research groups did not use the same methods (ultrafiltration versus fluorescence), which could explain the discrepancy of observed results about concomitant binding of palmitic acid and retinol. Subsequent studies confirmed the presence of two distinct binding sites. The second binding site on β-lg was hypothesized to be located in the hydrophobic pocket formed by the α-helix and the β-barrel, next to the dimer interface (Kontopidis et al. 2004; Yang et al. 2008; Wu et al. 1999). This was confirmed experimentally on the crystal structure of β-lg/vitamin D3 complex (Yang et al. 2008). These authors located the second binding site between the α-helix and the I β-strand. Another possible binding site was suggested to be located at the dimer interface (Wang et al. 1998). Using an ultrafiltration-based methodology, the binding of C14-palmitate to β-lg was found to be protein-concentration dependent (Wang et al. 1998). An increase in protein concentration from 1 to 200 μM, which increased the fraction of dimers in solution, resulted in the formation of a hydrophobic pocket for the binding of palmitate. This binding site was described by the authors on the β-lg dimer interface, binding 2 mol of palmitate per dimer of β-lg with an affinity constant in the order of 105 M−1. Another set of binding site with a weaker affinity was described on the surface of the monomer; the fitting equation highlighted that 24 mol of palmitate would bind to 1 mol of β-lg. This set of binding site disappears with the formation of the dimer. Note that the same study found only one binding site of palmitate at protein concentration of 20 μM when fluorescence technique was used instead of ultrafiltration (Wang et al. 1998). Other authors described also one binding site of palmitic acid per β-lg using equilibrium partition or NMR methods (Spector and Fletcher 1970; Ragona et al. 2000). The effect of pH and ionic strength on the number of binding sites was also illustrated in the case of binding of vitamin D3 to β-lg (Forrest et al. 2005). At low pH values, when the EF loop is closed, the binding can occur only at an external binding site. By varying pH and ionic strength, these authors showed that vitamin D3 binds to the protein in the calyx and at the surface of the protein. The latter binding site exhibited a lower affinity when the protein is monomeric, whereas affinity increased significantly when β-lg is dimeric.
3.2 Stoichiometry and association constants
The binding properties between β-lg and different FA are generally characterized by stoichiometry (molar ratio of ligand bound to the protein, n) and association constant (K a). Different methods have been used to access the values of these parameters including partition equilibrium, ultrafiltration, isothermal titration calorimetry (ITC), mass spectrometry, affinity chromatography, fluorescence, or nuclear magnetic resonance (Table 1).
Table 1 gives an overview of the parameters of interactions between numerous FA and native β-lg. Studies performed on binding FA with genetically or chemically modified β-lg are not considered here. Pérez et al. (1989) showed that bovine β-lg binds FA in milk, at a molar ratio of 1 mol of FA per mole of β-lg dimer. Palmitic acid and oleic acid, which are the major FA in milk, were found to be the main lipids bound to β-lg. The major part of reported binding was performed at neutral pH as β-lg does not bind easily hydrophobic ligands at low pH. Indeed, the binding of myristic, palmitic, and oleic acids to β-lg observed at pH 7 was not detectable at pH 3 (Frapin et al. 1993). The interaction between β-lg and palmitic acid was described to be reversible from pH 2.4 to 7.3 (Ragona et al. 2000). Similarly, Dufour et al. (1994a) highlighted that the interaction of β-lg/retinol was pH-dependent; its strength increased in the range 3 to 8. Using fluorescence, Dufour et al. (1992) found that cis-parinaric acid (C18:4), a very long chain fatty acid (VLCFA), binds β-lg at pH 3 with a high K a (K a = 4.7 × 107 M−1) but low n (n = 0.2). By increasing the pH to 7, these authors observed an increase in n values with no change in K a. Interestingly, an increase of binding affinity (Ka) by increasing pH, from 6 to 8, was found for short, medium, and long chain FA (Collini et al. 2003).
The binding affinity of short chain fatty acids (SCFA) to β-lg is relatively low (Collini et al. 2003; Loch et al. 2012a). The association constant of FA to β-lg increases with increasing chain length: between 102 M−1 and 104 M−1 for SCFA, between 103 M−1 and 106 M−1 for medium chain fatty acids (MCFA), and between 104 M−1 and 107 M−1 for LCFA and VLCFA. The increase in affinity constant with the length of the hydrocarbon chain is related to the increased hydrophobic interactions between the ligand and the hydrophobic calyx and maybe also to the lower solubility of FA in aqueous solution (Lide 2003; Beneventi et al. 2001). Palmitic acid was shown to have the highest K a among tested LCFA (Frapin et al. 1993; Spector and Fletcher 1970), supporting the idea that the β-lg calyx is most suited to bind FA with a 16-length carbon chain. For FA longer than 16 carbons, the hydrocarbon chain distorts to accommodate the calyx or the carboxylic group juts out of the calyx entrance (Loch et al. 2013a). However, the structural constraints imposed by the number and position of double bonds within FA only weakly affect its interaction parameters with β-lg (Frapin et al. 1993).
Most authors agree that the number of ligands bound per mole of protein monomer (n) is one, located at the main binding site (calyx). Although experimental values of the stoichiometry often diverge slightly from 1, a value of 1 agrees with crystallography measurements. In addition, for some ligands, a second set of binding sites located at β-lg surface was suggested by several authors. This second class of binding sites has a lower affinity (103 to 104 M−1) than the main binding site but n can vary substantially, from 2 to 24. These discrepancies in the values obtained for the binding parameters between β-lg and FA may simply be attributed to the diversity of used methodologies (fluorescence, partition equilibrium, and ITC). Otherwise, small association constants values, different sets of binding sites with close association constants, or other phenomena such as protein structural changes induced by ligand binding and modification of ligand solubility in the presence of protein may distort data and cause artifacts (Muresan et al. 2001). In addition, traces of impurities in the β-lg sample (other proteins, non-native forms of β-lg) may also affect data interpretation. In addition, calculated values for both K a and n can be highly dependent on the fitting procedure, as demonstrated by Frapin et al. (1993).
4 Biological properties of the β-lactoglobulin/ligand complexes
β-lg is the only major whey protein whose biological role, other than nutritional value, is still unknown. β-lg could participate to the digestion of milk lipids during the neonatal period by sequestering FA that inhibit pregastric lipase, enhancing consequently enzyme activity (Perez et al. 1992). It was thought for some time that β-lg may play a role in the transport of retinol from the mother to the neonate because of its homology with serum retinol-binding protein and its ability to bind retinol in vitro (Puyol et al. 1991). However, β-lg was shown to bind mainly FA but not retinol in milk (Kontopidis et al. 2002; Perez et al. 1992). As β-lg is not expressed in the milk of all species, it is unlikely that its ability to bind FA is its primary function. Indeed, pig and horse β-lg seem to have no affinity for FA (Perez et al. 1993). This failure to bind FA was attributed to the β-lg amino acids sequence variation, particularly at the C-terminus side, and the inability of porcine β-lg to form native dimers of the protein (Frapin et al. 1993).
4.1 Impact of ligand binding on the properties of the protein
The accessibility of the calyx of β-lg to ligand is dependent on pH. At pH below 6.0–6.5, the EF loop moves to a closed position preventing the binding of ligands (Qin et al. 1998). Therefore, interaction of FA to β-lg can only impact on the protein structure at non-acidic pH. Interestingly, Puyol et al. (1993) demonstrated that the binding of palmitic acid to β-lg has a protective effect on the protein against hydrolysis. In contrast, the binding of retinol to β-lg did not have a similar protective effect toward hydrolysis (Puyol et al. 1993; Mandalari et al. 2009). Besides, the protective effect of phosphatidylcholine against proteolysis of native β-lg could be due to the binding of the lipid that blocks the action of proteases for steric reasons (Mandalari et al. 2009).
Binding of FA, e.g., palmitate, to β-lg also protects the protein against heat-induced and chaotrope-induced denaturation (Barbiroli et al. 2011). FA acts by stabilizing the calyx at the hydrophobic interface between the barrel itself and the long helix, where the thiol group of Cys121 is buried. Other studies showed that binding of ligands such as myristic acid, conjugated linoleic acid (CLA), SDS, anilino naphthalene sulfonic acid (ANS), or retinol to β-lg stabilized the protein against both heat-induced and pressure-induced denaturation treatment (Considine et al. 2005, 2007). In many cases, changes in the structure of β-lg consecutive to ligand binding were reported. Thus, changes in the β-lg amide I band in the presence of tea polyphenols and aromatic compounds were observed (Kanakis et al. 2011; Tavel et al. 2010). Kanakis et al. (2011) attributed the structural changes to the large size of the ligands. In contrast, binding of linoleate or phosphatidylcholine to β-lg does not modify the protein secondary structure (Lefevre and Subirade 2000; Le Maux et al. 2012). However, β-lg aggregation in the presence of lipids has been reported (De Franceschi et al. 2011). Below the temperature of denaturation of the protein, CLA binding to β-lg induces the formation of complexes, approximately 170 nm in size, at pH 7 (Jiang and Liu 2010). Interestingly, the formation of covalent protein dimers and trimers consecutively to the interaction between β-lg and oleate or linoleate after heating at 60 °C was reported (Lišková et al. 2011; Le Maux et al. 2012).
4.2 Impact of the protein/fatty acid interaction on the properties of the ligand
Binding FA to proteins such as β-lg can modify the bioaccessibility of FA for biological cells. This can be measured by investigating FA transport into cells or FA cytotoxicity. LCFA are known to be cytotoxic to cells when taken up by cells (Lu et al. 2010). The cytotoxic effect of the FA when bound to β-lg either increases or decreases depending on the FA solubility. For instance, when CLA (100 μM) was part of a CLA/β-lg complex, its cytotoxicity on Caco-2 cells after 48 h exposure was 30% greater than that of free CLA (Jiang and Liu 2010). Similarly, the toxic effect of oleate/oleic acid was increased when bound to β-lg or α-la (Knyazeva et al. 2008; Lišková et al. 2011). Oleic acid/α-la complex exhibited a ∼40% increase in cytotoxic effect on human larynx carcinoma cells compared to free oleic acid (Knyazeva et al. 2008). In contrast, 58 μM of linoleate (in the absence of protein) decreased Caco-2 cell viability by 50%, whereas linoleate bound to β-lg had no effect on cell viability up to a concentration of 150 μM (Le Maux et al. 2012).
Jiang and Liu (2010) showed a twofold increase in CLA uptake by Caco-2 cells exposed to 100 μM CLA for 4 h from 45.8 μM to 85.9 μM when bound to β-lg as compared to free CLA. However, using a Caco-2 monolayer that mimics the intestinal barrier in combination with confocal imaging, the transport of linoleate into Caco-2 cells was decreased in the presence of β-lg in agreement with the cytotoxicity study (Le Maux et al. 2013b). Retinol and palmitic acid were also transported more efficiently across the monolayer in its free form than in complex with β-lg (Puyol et al. 1995; Riihimäki-Lampén 2009). Nevertheless, no significant change was detected for cholesterol transport under similar conditions (Puyol et al. 1995). Interestingly, oleic acid in lipidic micelles with taurocholate was absorbed more efficiently than oleic acid bound to BSA in Caco-2 cells (Levin et al. 1992). This can be explained by the emulsifying properties of taurocholate which may act as a free FA solubility enhancer (Levin et al. 1992).
Recent studies based on direct measurement of oleic acid in solution put back into question the increased cytotoxic effect of oleic acid/protein complex compared to free oleic acid. They argue that free oleic acid and oleic acid in complex with proteins at the same FA concentration have comparable cytotoxic effects toward various cells, with the protein alone having no effect (Brinkmann et al. 2011; Lišková et al. 2011). The above mentioned increased cytotoxic effect of oleic acid/β-lg or oleic acid/α-la complexes may relate to the difference of solubility of the FA in the absence and presence of protein with regard to the CMC of oleic acid. Oleic acid has a low solubility in aqueous solution, its CMC at pH 8.3, in the presence of 1 mM EDTA and no added salts, is 20 μM and 69 μM at 17 °C and 45 °C, respectively (Knyazeva et al. 2008). Therefore, the amount of available oleic acid (soluble) for the cells is low in the absence of proteins. The binding of oleic acid to β-lg or other proteins increases the amount of oleic acid in solution and consequently may increase the concentration of oleic acid accessible to the cells. Since solubility of FA increases with the number of C = C double bond along the FA aliphatic chain (Beneventi et al. 2001), the solubility of LA (C18:2) is higher than that of oleic acid (C18:1). Under the experimental conditions used by Collin et al. (2010), linoleate has a CMC of 2 mM. The impact of the FA solubility was highlighted by Norman et al. (1988) who found a higher cytotoxic effect of linoleate, the soluble salt form of linoleic acid, on the epithelial mouse cells, Ehrlich ascites tumor, compared to linoleic acid.
Docosahexaenoic acid (DHA) with β-lg protected DHA from oxidation at pH 7 and 40 °C, compared to DHA alone (Zimet and Livney 2009). Similarly, retinol was protected against enzymatic oxidation by BSA and β-lg (Futterman and Heller 1972). It has also been shown that retinol and β-carotene were protected against degradation by heating, oxidation, and irradiation by complexation with β-lg (Hattori et al. 1995).
5 Impact of the structure of β-lg on its fatty acid-binding properties
β-lg structure is highly sensitive to the processing conditions used in food manufacture, especially heat treatments, which are routinely used to change food textures or reduce microbial load. Such treatments denature the protein, leading to the formation of non-native monomers, oligomers and higher aggregates of β-lg in food products. Hence, with the exception of raw milk, β-lg is often found in a non-native conformation in food products. These changes in protein structure may have a strong impact on the binding properties of β-lg. However, few data are available on how FA interact with oligomers and aggregates of β-lg.
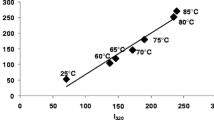
During the denaturation of β-lg, the low energy bonds that stabilize the protein weaken and buried amino acids become accessible to the solvent. The increased mobility of amino acid side chains allow intramolecular sulfhydryl-disulfide bond exchange reactions, resulting in the exposure of one of β-lg cysteine residues on the surface of the protein (Croguennec et al. 2003). The exposed sulfhydryl groups and hydrophobic amino acids are involved in the formation of aggregates (Havea et al. 1998; Sawyer 1968; Hoffmann and van Mil 1999). The modification of Cys121 in the β-lg protein weakens or totally eliminates the FA-binding capacity of β-lg by steric hindrance, which demonstrates the importance of protein structural integrity on ligand binding (Narayan and Berliner 1998). (Dufour et al. 1994b) reported an irreversible dissociation of β-lg/cis-parinaric acid complexes at high pressure, 350 MPa. Denatured β-lg, obtained after β-lg exposure to denaturing agents or high temperature, has a weaker affinity for palmitate compared to native β-lg (Spector and Fletcher 1970). Chemically modified β-lg (reductive ethylation of lysine residues in β-lg and methylated β-lg derivative) has a higher K a for benzo(α)pyrene and ellipticine compared to native β-lg (Dufour et al. 1992). However, a higher stoichiometry (n) was observed for the binding of cis-parinaric acid to denatured β-lg compared to native β-lg. Also, heating of β-lg at 75 °C for 10 to 20 min modified the binding parameters of 2-nonanone, with a decrease in binding affinity and an increase in stoichiometry (O'Neill and Kinsella 1988). By contrast, an increasing amount of linoleate bound per β-lg monomer was observed when the protein is heat denatured compared to native β-lg, without significant change of binding constant (Le Maux et al. 2013a). This was attributed to the exposure of internal hydrophobic patches on the surface of denatured β-lg molecules or the formation of hydrophobic cavities in β-lg aggregates. This binding difference increased linoleate uptake by Caco-2 cells (Le Maux et al. 2013a). Similarly, heated β-lg decreased the binding constant with vitamin D3, and therefore, uptake of vitamin D3 was decreased in mice (Yang et al. 2009). By heating β-lg at 100 °C for 16 min, binding of vitamin D3 to the calyx was ineffective although the external binding site remained thermally stable (Yang et al. 2008). Shpigelman et al. (2010) showed that binding of epigallocatechin gallate (EGCG) to β-lg resulted in the formation of nanoparticles of ∼50 nm when the protein was heat denatured (75–85 °C, 20 min). Binding of EGCG to denatured β-lg was stronger than to native β-lg. The large size of EGCG probably restricts its binding to native β-lg.
6 Interaction of FA with other whey proteins
In whey, β-lg coexists with other proteins which can modify the properties of the β-lg/FA complexes. The other major whey proteins, α-la and BSA, are also known to bind FA (Fletcher et al. 1971; Barbana et al. 2011). Consequently, in a complex mixture such as whey, the FA will exist in complex equilibrium between the total FA/protein complexes and its free form. The proportion of free FA will depend on the composition of the whey, the solubility of the FA, the FA/protein molar ratio, the proportion of each protein in the mixture, the structural state of the proteins, and the physicochemical conditions of the medium. Whey derived from cheese manufacture has very wide compositional variability; therefore, each whey has its own specific amounts of free and complexed FA. Together, these parameters could combine to affect positively or negatively the biological properties of the FA. However, to date, data on the biological properties of FA in the presence of several proteins are scarce.
α-Lactalbumin is the second major bovine whey protein, accounting for about 20% of the total whey proteins and is the principle protein in human milk due to the absence of β-lg. α-La has 123 amino acids and a molecular weight of 14.2 kDa. It contains eight cysteine residues, all engaged in disulfide bonds (Cys6-Cys120; Cys28-Cys111; Cys61-Cys77; Cys73-Cys91) that stabilize the tertiary structure of the protein. This protein has an important role in lactose synthesis, regulating the lactose synthase enzyme (Grobler et al. 1994; Fox and McSweeney 1998). Moreover, α-la can bind one calcium ion per protein (Hiraoka et al. 1980). In the absence of calcium, α-la (apo form) can bind FA such as oleic acid. Oleic acid/α-la complexes—known as HAMLET or BAMLET—constitute one of the most famous FA/α-la complexes because of its apoptotic activity on tumor cells (Knyazeva et al. 2008; Lišková et al. 2010; Fontana et al. 2013). Barbana et al. (2011) reviewed the interaction of FA with α-la and reported several binding sites (3 to 5) which have a similar association constant to FA/β-lg. The association constant between oleic acid and bovine α-la decreased from 2 × 105 M−1 to 2 × 104 M−1 when the temperature is decreased from 45 to 17 °C (Knyazeva et al. 2008; Barbana et al. 2006, 2008). Association constant values of 4.6 × 106 M−1 and 5.4 × 105 M−1 were reported for oleic acid/α-la and palmitic acid/α-la complexes, respectively (Barbana et al. 2006, 2008).
BSA is the third major protein of the bovine whey proteins, constituting about 10% of the total whey proteins. It consists of 582 amino acids with a molecular weight of 66.4 kDa (Cayot and Lorient 1998). It is a monomeric protein containing one sulfhydryl group and 17 disulfide bonds. BSA binds large amounts of hydrophobic molecules. BSA plays an important role in the digestion of milk fat due to its ability to bind free FA. The interactions of serum albumin with FA have been reviewed by van der Vusse (2009). Selective binding was found for radio-labelled palmitate which binds to BSA and β-lg but not to α-la (Perez et al. 1993). Palmitate was shown to bind to BSA with a higher affinity constant than to β-lg and with a higher stoichiometry (0.5 mol of palmitate bound per monomer of sheep β-lg versus 4.5 mol for sheep albumin). Palmitate and oleate bind to BSA with an association constant of approximately 107 M−1 and with a stoichiometry between 1 and 8. Fletcher et al. (1971) reported eight association constants between 107 and 104 M−1 for FA with 14, 16, and 18 carbons. Choi et al. (2002) described a decrease in the stoichiometry with increasing chain length of the FA. Spector and Fletcher (1970) demonstrated that for a palmitate/protein molar ratio of 0.8, palmitate bound to BSA was taken up by Ehrlich ascites tumor cells 20 times slower than palmitate bound to β-lg. This reduced bioaccessibility is attributed to the tighter binding of palmitate to BSA compared to β-lg; therefore, the FA was less bioaccessible to the cells when bound to BSA.
7 Conclusion
The molecular structure of bovine β-lg has been extensively studied. Its globular structure allows it to naturally bind hydrophobic ligands such as FA. Its biological function, other than nutrition, is unknown but it may serve to deliver FA to the enterocyte. However, the ability to bind hydrophobic components is species dependent, as some variants of β-lg are unable to bind FA.
The main binding site for hydrophobic ligands is the β-lg calyx. A secondary binding site is located at the surface of the protein next to the dimer interface. This explains why β-lg in its monomeric or dimeric native form does not have the same stoichiometry. The stoichiometry of the β-lg/ligand is difficult to measure unambiguously as methodologies used are mainly indirect. Even if affinity constants are of the same order of magnitude between studies, ratios of FA bound to β-lg may diverge largely. The β-lg affinity for FA is dependent on the properties of the FA. Palmitic acid is the most tightly bound FA to the β-lg structure. In manufactured and processed food products, β-lg exists under very different structures which alter its binding properties for ligands.
The interaction of β-lg with ligands such as FA modifies the bioaccessibility of the ligand. It would appear that when the FA solubility is low, β-lg increases its bioaccessibility, whereas this interaction decreases the FA bioaccessibility when the ligand has a high solubility. This constitutes a means to control FA uptake during digestion. As such, β-lg may be used in food matrices to influence the bioaccessibility of FA.
References
Aymard P, Durand D, Nicolaï T (1996) The effect of temperature and ionic strength on the dimerisation of β-lactoglobulin. Int J Biol Macromol 19:213–221
Barbana C, Perez MD, Sanchez L, Dalgalarrondo M, Chobert JM, Haertle T (2006) Interaction of bovine alpha-lactalbumin with fatty acids as determined by partition equilibrium and fluorescence spectroscopy. Int Dairy J 16(1):18–25
Barbana C, Perez MD, Pocovi C, Sanchez L, Wehbi Z (2008) Interaction of human alpha-lactalbumin with fatty acids: determination of binding parameters. Biochemistry-Moscow 73(6):711–716
Barbana C, Sanchez L, Perez MD (2011) Bioactivity of alpha-lactalbumin related to its interaction with fatty acids: a review. Crit Rev Food Sci Nutr 51(8):783–794
Barbiroli A, Bonomi F, Ferranti P, Fessas D, Nasi A, Rasmussen P, Iametti S (2011) Bound fatty acids modulate the sensitivity of bovine beta-lactoglobulin to chemical and physical denaturation. J Agric Food Chem 59(10):5729–5737
Beneventi D, Carré B, Gandini A (2001) Precipitation and solubility of calcium soaps in basic aqueous media. J Colloid Interface Sci 237(1):142–144
Bouhallab S, Henry G, Caussin F, Croguennec T, Fauquant J, Mollé D (2004) Copper-catalyzed formation of disulfide-linked dimer of bovine β-lactoglobulin. Lait 84:517–525
Boye JI, Ma CY, Ismail A (2004) Thermal stability of β-lactoglobulins A and B: effect of SDS, urea, cysteine and N-ethylmaleimide. J Dairy Res 71:207–215
Brinkmann CR, Heegaard CW, Petersen TE, Jensenius JC, Thiel S (2011) The toxicity of bovine alpha-lactalbumin made lethal to tumor cells is highly dependent on oleic acid and induces killing in cancer cell lines and noncancer-derived primary cells. FEBS J 278(11):1955–1967
Brownlow S, Cabral JHM, Cooper R, Flower DR, Yewdall SJ, Polikarpov I, North ACT, Sawyer L (1997) Bovine beta-lactoglobulin at 1.8 angstrom resolution—still an enigmatic lipocalin. Structure 5(4):481–495
Burova TV, Choiset Y, Tran V, Haertle T (1998) Role of free Cys121 in stabilization of bovine beta-lactoglobulin B. Protein Eng 11(11):1065–1073
Cayot P, Lorient D (1998) Structures et technofonctions des protéines du lait. Technique et Documentation Lavoisier, Paris
Choi JK, Ho J, Curry S, Qin DH, Bittman R, Hamilton JA (2002) Interactions of very long-chain saturated fatty acids with serum albumin. J Lipid Res 43(7):1000–1010
Collin F, Hindo J, Thérond P, Couturier M, Cosson C, Jore D, Gardès-Albert M (2010) Experimental evidence of the reciprocal oxidation of bovine serum albumin and linoleate in aqueous solution, initiated by HO-free radicals. Biochimie 92(9):1130–1137
Collini M, D'Alfonso L, Molinari H, Ragona L, Catalano M, Baldini G (2003) Competitive binding of fatty acids and the fluorescent probe 1-8-anilinonaphthalene sulfonate to bovine beta-lactoglobulin. Protein Sci 12(8):1596–1603
Considine T, Patel HA, Singh H, Creamer LK (2005) Influence of binding of sodium dodecyl sulfate, all-trans-retinol, palmitate, and 8-anilino-1-naphthalenesulfonate on the heat-induced unfolding and aggregation of β-lactoglobulin B. J Agric Food Chem 53(8):3197–3205
Considine T, Patel HA, Singh H, Creamer LK (2007) Influence of binding conjugated linoleic acid and myristic acid on the heat-and high-pressure-induced unfolding and aggregation of beta-lactoglobulin B. Food Chem 102(4):1270–1280
Creamer LK, Parry DAD, Malcolm GN (1983) Secondary structure of bovine beta-lactoglobulin B. Arch Biochem Biophys 227(1):98–105
Croguennec T, Bouhallab S, Mollé D, O’Kennedy BT, Mehra R (2003) Stable monomeric intermediate with exposed Cys-119 is formed during heat denaturation of beta-lactoglobulin. Biochem Biophys Res Commun 301(2):465–471
Croguennec T, O’Kennedy BT, Mehra R (2004) Heat-induced denaturation/aggregation of beta-lactoglobulin A and B: kinetics of the first intermediates formed. Int Dairy J 14(5):399–409
De Franceschi G, Frare E, Pivato M, Relini A, Penco A, Greggio E, Bubacco L, Fontana A, de Laureto PP (2011) Structural and morphological characterization of aggregated species of alpha-synuclein induced by docosahexaenoic acid. J Biol Chem 286(25):22262–22274
Dufour E, Marden MC, Haertlé T (1990) Beta-lactoglobulin binds retinol and protoporphyrin IX at two different binding sites. FEBS Lett 277(1):223–226
Dufour E, Roger P, Haertlé T (1992) Binding of benzo(α)pyrene, ellipticine, and cis-parinaric acid to beta-lactoglobulin: influence of protein modifications. J Protein Chem 11(6):645–652
Dufour E, Genot C, Haertlé T (1994a) Beta-lactoglobulin binding properties during its folding changes studied by fluorescence spectroscopy. BBA-Protein Struct M 1205(1):105–112
Dufour E, Hoa GHB, Haertlé T (1994b) High-pressure effects on β-lactoglobulin interactions with ligands studied by fluorescence. BBA-Protein Struct M 1206(2):166–172
Fletcher JE, Spector AA, Ashbrook JD (1971) Analysis of long-chain free fatty acid binding to bovine serum albumin by determination of stepwise equilibrium constants. Biochemistry 10(17):3229–3232
Flower DR (1996) The lipocalin protein family: structure and function. Biochem J 318(1):1–14
Fontana A, Spolaore B, Polverino de Laureto P (2013) The biological activities of protein/oleic acid complexes reside in the fatty acid. Biochim Biophys Acta (BBA) - Proteins Proteomics 1834(6):1125–1143
Forrest SA, Yada RY, Rousseau D (2005) Interactions of vitamin D3 with bovine beta-lactoglobulin A and beta-casein. J Agric Food Chem 53(20):8003–8009
Fox PF, McSweeney PLH (1998) Dairy chemistry and biochemistry. Chapman & Hall, London
Frapin D, Dufour E, Haertle T (1993) Probing the fatty acid binding site of beta-lactoglobulins. J Protein Chem 12(4):443–449
Futterman S, Heller J (1972) The enhancement of fluorescence and the decreased susceptibility to enzymatic oxidation of retinol complexed with bovine serum albumin, beta-lactoglobulin, and the retinol-binding protein of human plasma. J Biol Chem 247(16):5168–5172
Godovac-Zimmermann J, Krause I, Baranyi M, Fischer-Frühholz S, Juszczak J, Erhardt G, Buchberger J, Klostermeyer H (1996) Isolation and rapid sequence characterization of two novel bovine beta-lactoglobulins I and J. J Protein Chem 15(8):743–750
Grobler JA, Wang M, Pike A, Brew K (1994) Study by mutagenesis of the roles of two aromatic clusters of alpha-lactalbumin in aspects of its action in the lactose synthase system. J Biol Chem 269(7):5106–5114
Hattori M, Watabe A, Takahashi K (1995) Beta-lactoglobulin protects beta-ionone related compounds from degradation by heating, oxidation, and irradiation. Biosci Biotechnol Biochem 59(12):2295–2297
Havea P, Singh H, Creamer LK, Campanella OH (1998) Electrophoretic characterization of the protein products formed during heat treatment of whey protein concentrate solutions. J Dairy Res 65(01):79–91
Hiraoka Y, Segawa T, Kuwajima K, Sugai S, Murai N (1980) Alpha-lactalbumin: a calcium metalloprotein. Biochem Biophys Res Commun 95(3):1098–1104
Hoffmann MA, van Mil PJ (1999) Heat-induced aggregation of β-lactoglobulin as a function of pH. J Agric Food Chem 47(5):1898–1905
Jayat D, Gaudin J-C, Chobert J-M, Burova TV, Holt C, McNae I, Sawyer L, Haertlé T (2004) A recombinant C121S mutant of bovine beta-lactoglobulin is more susceptible to peptic digestion and to denaturation by reducing agents and heating. Biochemistry 43(20):6312–6321
Jensen RG (1995) Handbook of milk composition. Academic, San Diego
Jiang HR, Liu N (2010) Self-assembled beta-lactoglobulin-conjugated linoleic acid complex for colon cancer-targeted substance. J Dairy Sci 93(9):3931–3939
Kanakis CD, Hasni I, Bourassa P, Tarantilis PA, Polissiou MG, Tajmir-Riahi H-A (2011) Milk beta-lactoglobulin complexes with tea polyphenols. Food Chem 127(3):1046–1055
Knyazeva EL, Grishchenko VM, Fadeev RS, Akatov VS, Permyakov SE, Permyakov EA (2008) Who is Mr. HAMLET? Interaction of human alpha-lactalbumin with monomeric oleic acid. Biochemistry 47(49):13127–13137
Kontopidis G, Holt C, Sawyer L (2002) The ligand-binding site of bovine beta-lactoglobulin: evidence for a function? J Mol Biol 318(4):1043–1055
Kontopidis G, Holt C, Sawyer L (2004) Invited review: beta-lactoglobulin: binding properties, structure, and function. J Dairy Sci 87(4):785–796
Le Maux S, Giblin L, Croguennec T, Bouhallab S, Brodkorb A (2012) Beta-lactoglobulin as a molecular carrier of linoleate: characterization and effects on intestinal epithelial cells in vitro. J Agric Food Chem 60(37):9476–9483
Le Maux S, Bouhallab S, Giblin L, Brodkorb A, Croguennec T (2013a) Complexes between linoleate and native or aggregated beta-lactoglobulin: interaction parameters and in vitro cytotoxic effect. Food Chem 141(3):2305–2313
Le Maux S, Brodkorb A, Croguennec T, Hennessy AA, Bouhallab S, Giblin L (2013b) Beta-lactoglobulin-linoleate complexes: in vitro digestion and the role of protein in fatty acid uptake. J Dairy Sci 96(7):4258–4268
Lefevre T, Subirade M (2000) Interaction of beta-lactoglobulin with phospholipid bilayers: a molecular level elucidation as revealed by infrared spectroscopy. Int J Biol Macromol 28(1):59–67
Levin M, Talkad V, Gordon J, Stenson W (1992) Trafficking of exogenous fatty acids within Caco-2 cells. J Lipid Res 33(1):9–19
Lide D (2003) Handbook of chemistry and physics, 84th Edition. CRC, London
Lišková K, Kelly AL, O'Brien N, Brodkorb A (2010) Effect of denaturation of α-lactalbumin on the formation of BAMLET (bovine alpha-lactalbumin made lethal to tumor cells). J Agric Food Chem 58(7):4421–4427
Lišková K, Auty MAE, Chaurin V, Min S, Mok KH, O'Brien N, Kelly AL, Brodkorb A (2011) Cytotoxic complexes of sodium oleate with beta-lactoglobulin. Eur J Lipid Sci Tech:1207–1218
Liu L, Kitova EN, Klassen JS (2011) Quantifying protein-fatty acid interactions using electrospray ionization mass spectrometry. J Am Soc Mass Spectrom 22(2):310–318
Loch J, Polit A, Gerocki A, Bonarek P, Kurpiewska K, Dziedzicka-Wasylewska M, Lewinski K (2011) Two modes of fatty acid binding to bovine beta-lactoglobulin-crystallographic and spectroscopic studies. J Mol Recognit 24(2):341–349
Loch J, Polit A, Bonarek P, Olszewska D, Kurpiewska K, Dziedzicka-Wasylewska M, Lewiński K (2012a) Structural and thermodynamic studies of binding saturated fatty acids to bovine beta-lactoglobulin. Int J Biol Macromol 50(4):1095–1102
Loch J, Polit A, Bonarek P, Ries D, Kurpiewska K, Dziedzicka Wasylewska M, Lewiński K (2012b) Bovine beta-lactoglobulin complex with linoleic acid. http://www.cgl.ucsf.edu/chimera (accessed 5 June 2012)
Loch JI, Bonarek P, Polit A, Riès D, Dziedzicka-Wasylewska M, Lewiński K (2013a) Binding of 18-carbon unsaturated fatty acids to bovine β-lactoglobulin—structural and thermodynamic studies. Int J Biol Macromol 57:226–231
Loch JI, Bonarek P, Polit A, Świątek S, Dziedzicka-Wasylewska M, Lewiński K (2013b) The differences in binding 12-carbon aliphatic ligands by bovine β-lactoglobulin isoform A and B studied by isothermal titration calorimetry and X-ray crystallography. J Mol Recognit 26(8):357–367
Lu XF, He GQ, Yu HN, Ma Q, Shen SR, Das UN (2010) Colorectal cancer cell growth inhibition by linoleic acid is related to fatty acid composition changes. J Zhejiang Univ-SCI B 11(12):923–930
Mandalari G, Mackie AM, Rigby NM, Wickham MSJ, Mills EN (2009) Physiological phosphatidylcholine protects bovine beta-lactoglobulin from simulated gastrointestinal proteolysis. Mol Nut Food Res 53(S1):S131–S139
Manderson GA, Hardman MJ, Creamer LK (1999) Effect of heat treatment on bovine β-lactoglobulin A, B, and C explored using thiol availability and fluorescence. J Agric Food Chem 47:3617–3627
McMeekin T, Polis B, DellaMonica E, Custer J (1949) A crystalline compound of beta-lactoglobulin with dodecyl sulfate. J Am Chem Soc 71(11):3606–3609
Mercadante D, Melton LD, Norris GE, Loo TS, Williams MAK, Dobson RCJ, Jameson GB (2012) Bovine β-lactoglobulin is dimeric under imitative physiological conditions: dissociation equilibrium and rate constants over the pH range of 2.5–7.5. Biophys J 103:303–312
Muresan S, van der Bent A, de Wolf FA (2001) Interaction of beta-lactoglobulin with small hydrophobic ligands as monitored by fluorometry and equilibrium dialysis: nonlinear quenching effects related to protein–protein association. J Agric Food Chem 49(5):2609–2618
Narayan M, Berliner LJ (1998) Mapping fatty acid binding to beta-lactoglobulin: ligand binding is restricted by modification of Cys 121. Protein Sci 7(1):150–157
Norman A, Bennett LR, Mead JF, Iwamoto KS (1988) Antitumor activity of sodium linoleate. Nutr Cancer 11(2):107–115
O'Kennedy BT, Mounsey JS, Murphy F, Pesquera L, Mehra R (2006) Preferential heat-induced denaturation of bovine beta-lactoglobulin variants as influenced by pH. Milchwissenschaft-Milk Sci Int 61(4):366–369
O'Neill T, Kinsella J (1988) Effect of heat treatment and modification on conformation and flavor binding by beta-lactoglobulin. J Food Sci 53(3):906–909
Papiz MZ, Sawyer L, Eliopoulos EE, North ACT, Findlay JBC, Sivaprasadarao R, Jones TA, Newcomer ME, Kraulis PJ (1986) The structure of beta-lactoglobulin and its similarity to plasma retinol-binding protein. Nature 324(6095):383–385
Patocka G, Jelen P (1991) Calcium association with isolated whey proteins. Can Inst Food Sci Technol J 24(5):218–223
Pérez MD, de Villegas CD, Sánchez L, Aranda P, Ena JM, Calvo M (1989) Interaction of fatty acids with beta-lactoglobulin and albumin from ruminant milk. J Biochem (Tokyo) 106(6):1094–1097
Perez MD, Sanchez L, Aranda P, Ena J, Oria R, Calvo M (1992) Effect of beta-lactoglobulin on the activity of pregastric lipase. A possible role for this protein in ruminant milk. BBA-Lipid Lipid Met 1123(2):151–155
Perez MD, Puyol P, Ena JM, Calvo M (1993) Comparison of the ability to bind lipids of beta-lactoglobulin and serum albumin of milk from ruminant and non-ruminant species. J Dairy Res 60(1):55–63
Puyol P, Perez MD, Ena JM, Calvo M (1991) Interaction of bovine beta-lactoglobulin and other bovine and human whey proteins with retinol and fatty acids. Agric Biol Chem 55(10):2515–2520
Puyol P, Perez MD, Mata L, Ena JM, Calvo M (1993) Effect of retinol and fatty acid binding by bovine beta-lactoglobulin on its resistance to trypsin digestion. Int Dairy J 3(7):589–597
Puyol P, Perez MD, Peiro JM, Calvo M (1994) Effect of binding of retinol and palmitic acid to bovine beta-lactoglobulin on its resistance to thermal denaturation. J Dairy Sci 77(6):1494–1502
Puyol P, Dolores Perez M, Sanchez L, Ena JM, Calvo M (1995) Uptake and passage of beta-lactoglobulin, palmitic acid and retinol across the Caco-2 monolayer. BBA Biomembranes 1236(1):149–154
Qin BY, Bewley MC, Creamer LK, Baker HM, Baker EN, Jameson GB (1998) Structural basis of the Tanford transition of bovine beta-lactoglobulin. Biochemistry 37(40):14014–14023
Qin BY, Jameson GB, Bewley MC, Baker EN, Creamer LK (1999) Functional implications of structural differences between variants A and B of bovine beta-lactoglobulin. Protein Sci 8(1):75–83
Ragona L, Zetta L, Fogolari F, Molinari H, Pérez DM, Puyol P, Kruif KD, Löhr F, Rüterjans H (2000) Bovine beta-lactoglobulin: interaction studies with palmitic acid. Protein Sci 9(7):1347–1356
Renard D, Lefebvre J, Griffin MCA, Griffin WG (1998) Effects of pH and salt environment on the association of β-lactoglobulin revealed by intrinsic fluorescence studies. Int J Biol Macromol 22:41–49
Riihimäki-Lampén L (2009) Interactions of natural products with beta-lactoglobulins, members of the lipocalin family. University of Helsinki
Sawyer W (1968) Heat denaturation of bovine β-lactoglobulins and relevance of disulfide aggregation. J Dairy Sci 51(3):323–329
Sawyer L, Kontopidis G (2000) The core lipocalin, bovine beta-lactoglobulin. BBA-Protein Struct M 1482(1–2):136–148
Shpigelman A, Israeli G, Livney YD (2010) Thermally-induced protein-polyphenol co-assemblies: beta lactoglobulin-based nanocomplexes as protective nanovehicles for EGCG. Food Hydrocoll 24(8):735–743
Smith LM, Fantozzi P, Creveling RK (1983) Study of triglyceride-protein ineraction using a microemulsion-filtration method. J Am Oil Chem Soc 60(5):960–967
Smithers GW (2008) Whey and whey proteins—from ‘gutter-to-gold’. Int Dairy J 18(7):695–704
Sneharani AH, Karakkat JV, Singh SA, Rao AGA (2010) Interaction of curcumin with beta-lactoglobulin—Stability, spectroscopic analysis, and molecular modeling of the complex. J Agric Food Chem 58(20):11130–11139
Spector AA, Fletcher JE (1970) Binding of long chain fatty acids to beta-lactoglobulin. Lipids 5(4):403–411
Tanford C (1980) The hydrophobic effect: formation of micelles and biological membranes, 2nd edn. Wiley, New York
Tavel L, Moreau C, Bouhallab S, Li-Chan ECY, Guichard E (2010) Interactions between aroma compounds and beta-lactoglobulin in the heat-induced molten globule state. Food Chem 119(4):1550–1556
van der Vusse GJ (2009) Albumin as fatty acid transporter. Drug Metab Pharmacokinet 24(4):300–307
Verheul M, Pedersen JS, Roefs SPFM, de Kruif KG (1999) Association behavior of native β-lactoglobulin. Biopolymers 49:11–20
Wang Q, Allen JC, Swaisgood HE (1997) Binding of vitamin D and cholesterol to beta-lactoglobulin. J Dairy Sci 80(6):1054–1059
Wang QWQ, Allen JC, Swaisgood HE (1998) Protein concentration dependence of palmitate binding to beta-lactoglobulin. J Dairy Sci 81(1):76–81
Wu SY, Pérez MD, Puyol P, Sawyer L (1999) Beta-lactoglobulin binds palmitate within its central cavity. J Biol Chem 274(1):170–174
Yan Y, Seeman D, Zheng B, Kizilay E, Xu Y, Dubin PL (2013) pH-Dependent aggregation and disaggregation of native β-lactoglobulin in low salt. Langmuir 29:4584–4593
Yang MC, Guan HH, Liu MY, Lin YH, Yang JM, Chen WL, Chen CJ, Mao SJT (2008) Crystal structure of a secondary vitamin D3 binding site of milk beta-lactoglobulin. Proteins 71(3):1197–1210
Yang MC, Chen NC, Chen CJ, Wu CY, Mao SJT (2009) Evidence for beta-lactoglobulin involvement in vitamin D transport in vivo—role of the γ-turn (Leu-Pro-Met) of beta-lactoglobulin in vitamin D binding. FEBS J 276(8):2251–2265
Zimet P, Livney YD (2009) Beta-lactoglobulin and its nanocomplexes with pectin as vehicles for ω-3 polyunsaturated fatty acids. Food Hydrocoll 23(4):1120–1126
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Le Maux, S., Bouhallab, S., Giblin, L. et al. Bovine β-lactoglobulin/fatty acid complexes: binding, structural, and biological properties. Dairy Sci. & Technol. 94, 409–426 (2014). https://doi.org/10.1007/s13594-014-0160-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13594-014-0160-y