
Abstract
Our study investigated the possibility of using the red mason bee Osmia bicornis as a pollinator of forest seed orchards of frequently grown European trees: insect - pollinated Tilia cordata and Prunus avium. Moreover, we checked whether the presence of O. bicornis has an impact on the pollination results of Quercus petraea, which is wind - pollinated. Data were collected from 8 forest seed orchards in western Poland between 2018 and 2020. We assessed whether the presence of O. bicornis populations affects the number and mass of seeds and seed quality. Supplementing forest orchards with O. bicornis affected seed yield. The generalized linear mixed models indicated that the number of seeds and the proportion of viable seeds, but not seed mass, were higher in the Tilia cordata trees, when additionally pollinated with O. bicornis added as a pollinator.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
In forestry, seed orchards are plantations of high - quality seeds of known origin (Kowalczyk et al. 2011), capable of germinating and furthering forest production (Chałupka et al. 2011). Seed orchards are created to obtain an abundant harvest of seeds with increased genetic value and to facilitate seed harvesting (Kocięcki 1965; Kowalczyk et al. 2011). Moreover, seed orchards constitute a valuable base for protecting the genetic variability of forest trees and the production of seed that is well adapted to specific environments (Nanson 1972; Kowalczyk et al. 2011). Due to the growing demand for seeds of various species of trees over the last several decades, the creation of new seed plantations has been observed (Ruotsalainen 2014; Gömöry et al. 2020; Liesebach et al. 2021).
Wild bee species, the main group that provides the ecosystem service of pollination (Garibaldi et al. 2011, 2013), have been declining in many parts of the world (e.g., Potts et al. 2010; Goulson et al. 2015). Insect pollination services are worth 14.6 billion EUR annually in the EU and more than 24 billion USD in the USA (Leonhardt et al. 2013). Global pollinator decline and lack of their services have a significant negative economic effect on crop production worldwide (Kevan and Viana 2003). The observed decline in the populations of pollinators is noticeable in horticulture and agriculture (Kearns et al. 1998; Klein et al. 2007; Radmacher and Strohm 2010; Bąk and Wilde 2003). Moreover, the yield from some seed orchards has also been low, and insufficient pollination has been suggested as a possible cause (Moncur et al. 1995). Commercially managed species such as honey bees, Apis mellifera (Apidae), are used to supplement wild bees to prevent low yields of various crops (Allsopp et al. 2008; Breeze et al. 2011; Garibaldi et al. 2013). Honey bees were also introduced in forest seed orchards of insect - pollinated trees such as Eucalyptus and Acacia to increase seed quantity and quality (Hardi and Kanso 1995; Moncur et al. 1995). Managed solitary bees of the genus Osmia also proved to be effective pollinators in several crops (Batra 1978; Torchio and Asensio 1985; Bosch and Kemp 2002). Osmia bicornis (Linnaeus 1758) (previously Osmia rufa) has been used as an additional pollinator in European fruit tree orchards and on other crops such as strawberries and oilseed rape from the 1970s (Hansted et al. 2014; Sedivy and Dorn 2014; Wilkaniec and Radajewska 1997; Gruber et al. 2011). However, the use of managed solitary bees as supplemental crop pollinators is still limited.
O. bicornis is a widely distributed univoltine, solitary, polylectic species native to Europe (Krunic and Stanisavljevic 2006). O. bicornis nest gregariously in artificial nesting materials. It is active from mid - April until the end of June in Central Europe (Steffan - Dewenter and Schiele 2008), but commercial rearing techniques ensure the availability of adults for release at a desired time of the year to synchronize bee emergence and activity with the bloom of a target crop (Bosch and Blas 1994; Krunic et al. 1995). The use of managed bees to supplement pollination positively affects the yield and quality of fruits (e.g., strawberries, fruit trees) compared to pollination by wild pollinators alone (Lye et al. 2011; Ryder et al. 2020). However, there is a lack of information on managed O. bicornis in forest orchard production systems. The role of O. bicornis in forest orchards has yet to be quantified, and managed wild bees are not commonly incorporated into forest orchard management. Therefore, research is required to support the optimization of pollination services in the forest orchard. This study investigates the hypothesis that supplementing pollination by managed O. bicornis will increase the yield and the seed quality of forest orchards.
This study aimed to investigate the possibility of using O. bicornis as a pollinator of forest seed orchards of frequently grown European insect - pollinated trees: Tilia cordata and Prunus avium. Moreover, we checked whether the presence of O. bicornis has an impact on the pollination results of Quercus petraea, which is wind - pollinated. The following questions were addressed: (1) does the presence of O. bicornis populations in a seed orchard increase the number and mass of seeds? and (2) does O. bicornis pollination also affect seed quality?
2 Materials and methods
2.1 Study area
Small - leaved lime (a.k.a. linden) (Tilia cordata Mill.), sessile oak (Quercus petraea (Matt.) Liebl., and wild cherry (Prunus avium L.) seed orchards located in western Poland were selected for the research. T. cordata was cultivated in 4 forest districts: Jastrowie (53°39″N; 116°74ʺE), Pniewy (52°48″N; 16°26″E), Łopuchówko (52°57″N; 17°09″E), and Świerczyna (53°42″N; 16°21″E). The sizes of these orchards were as follows: 4.00, 4.32, 2.20, and 4.43 ha. Q. petraea cultivated areas were located in Tuczno (53°22ʺN; 16°09″E) and Złocieniec (53°45″N; 16°07″E) and covered the area of 7.89 and 4.76 ha. P. avium orchards were located in Łopuchówko (52°57″N; 17°10″E) and Świerczyna (53°46″N; 16°30″E) and had areas of 2.18 and 3.98 ha respectively. These seed orchards were established at different times: Jastrowie in 1987, Łopuchówko (Prunus in 2007, Tilia in 1996), Pniewy in 1996, Świerczyna (Prunus in 2010, Tilia in 2000), Tuczno in 2005, Złocieniec in 2005.
2.2 Study species
2.2.1 Tilia cordata
The range of Tilia cordata Mill. covers almost the whole continental part of Europe. Its distribution is suboceanic to subcontinental. It is most abundant from north - eastern France through central Germany and Poland to central Russia and northern Ukraine (Radoglou et al. 2009). Their importance as a significant part of forest biodiversity is expected to increase, especially in the changing environment, due to climate change (Radoglou et al. 2009; De Jaegere et al. 2016; Zhelev et al. 2020). Because of their esthetic and cultural value, lime trees are also often planted in urban areas of Central Europe as park trees and along avenues and roads (Weryszko - Chmielewska and Sadowka 2010). Tilia cordata flowers in July. Limes are principally insect - pollinated trees, though wind pollination also occurs. Bees are the main visitors and pollinators, even if other generalist insects are also observed on Tilia trees (Anderson 1976; Pigott 1991). Flowers of T. cordata are regarded as nutritionally very attractive for bees (Jabłoński et al. 2000).
2.2.2 Prunus avium
Modern silvicultural and management regimes should ensure the forest ecosystem biodiversity and obtain valuable wood that could increase the income of forest owners. Wild cherry (Prunus avium L.) is a very valuable forest tree species and one of noble hardwoods. It is a mainly European species and served as the origin of all cultivated forms. It blooms from April to May. Flowers are pollinated mainly by honey bees and wild bees, and the trees are generally not self - fertile (Welk et al. 2016).
2.2.3 Quercus petraea
Quercus petraea (Matt.) Liebl. is a widespread species native to most of Europe, found from Scandinavia to the Iberian Peninsula. The wood from oaks is hard and durable and has been valued for centuries. Dieback and decline in oak have been reported in Europe since the early 1900 s and have been attributed to single or combined effects of various abiotic factors. Q. petraea co - occur at many sites as the main component of temperate deciduous mixed forests. It is also appreciated as a park and roadside tree. The sessile oak is monoecious and wind - pollinated. Although temperate wind - pollinated species are usually discounted as bee resources, there is evidence that bees collect such pollen (Saunders 2018), and some wind - pollinated trees may be good protein sources (Roulston and Cane 2000). O. bicornis representatives eagerly visit oak flowers and collect large quantities of pollen (Hallmen and van Leeuwen 1990). Therefore, we included plantations of this species in our research to check whether the presence of additionally introduced pollinators may impact the yield of these trees.
2.3 Data collection
Data were collected from the 8 forest seed orchards for 3 years each (2018–2020). Twenty - four (12 experimental and 12 control) trees of Tilia cordata and 12 (6 experimental and 6 control) Prunus avium trees and 12 (6 experimental and 6 control) Quercus petraea trees were selected. The experimental and paired control trees without O. bicornis were from one clone for genetic compatibility. Selected trees (experimental and control) were at least 500 m away from each other. They were the same age and size and had similar diameters at breast height (DBH). Sixteen artificial bee nests and cocoons with Osmia bicornis were placed next to every experimental tree. Artificial nests consisted of 60–70 reeds (Phragmites australis) of different diameters (5–8 mm) and 20 cm length placed in plastic tubes. The artificial nests were located in dry and sunny places, 100 cm above the ground (Appendix, Figure A1). Cocoons were removed from nest cells in the winter months and maintained in a cold room under conditions of continuous darkness with the temperature of 4 °C. Four to five days before being moved to Prunus and Quercus experimental plots, cocoons were transferred to 20 °C. Placing the release boxes in the orchards was synchronized with the flowering of the trees. The 1000 O. bicornis cocoons were placed in each release box.

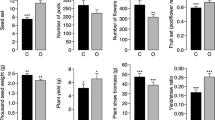
Mean (whiskers, SE) of the proportion of viable seeds, number of seeds, and mass of thousand seeds (g) for control (without supplementing pollination by O. bicornis) and experiment (with supplementing pollination by O. bicornis) trees of Tilia cordata, Quercus petraea, and Prunus avium. Asterisks indicate significant differences (*p < 0.05, **p < 0.01), n = number of trees sampled.
Seeds were collected from the selected experimental and control trees to determine the quantity and quality of the seeds. The lime seeds came from branches of similar length and number of lateral branches located at 3 height levels (1.5, 3, and 4.5 m). Samples were additionally collected from 4 different geographic directions for each of the levels, i.e., 12 times from each tree. For oak and cherry trees, all seeds from the trees were collected. After harvesting, seed samples were assessed using the cutting method commonly used to evaluate seed material in forestry (Załęski et al. 2000). The following seed parameters were determined: the weight of a thousand seeds, the number of seeds: empty, healthy, rotten, and the ability to germinate. The results concerning the number of obtained seeds and their weight and germination capacity were compared between the experimental and control trees. The experiment was carried out so that the trees of the corresponding clones, which were control trees in the first year of the study, were experimental in the next year, and vice versa, only changing the location of aggregations of bees.
The number of samples obtained for Prunus and Quercus was lower than planned due to the lack of fruit production due to spring frost damage and water deficits.
2.4 Statistical analysis
We assessed differences in the proportion of viable seeds, the number of seeds, and the mass of the thousand seeds between the experiment (supplemental pollination by O. bicornis) and control (without supplemental pollination by O. bicornis) groups separately for tree species using generalized linear mixed - effects models (GLMMs) with year and site as random factors and restricted maximum - likelihood (REML) estimator. We assumed the Gaussian distribution of mass of a thousand seeds, negative binomial of the number of seeds, and beta distribution of the proportion of viable seeds. Additionally, we assessed differences in the proportion of empty seeds of Tilia cordata and the proportion of non - viable seeds for both tree species between the control and experimental group. Prior to analyses, the mass of the thousand seeds was logarithm - transformed (log10). The proportion of viable seeds was treated as the response variable after applying a small, linear transformation to eliminate zeroes and ones (Smithson and Verkuilen 2006). Because of the small sample size for Prunus avium, we used a non - parametric U- Mann–Whitney test to assess differences in the proportion of viable seeds, the mass of the thousand seeds, and the number of seeds between the experimental (n = 4) and control groups (n = 4).
Additionally, we calculated the standardized effect size (Cohen d) for our response variable for experimental and control groups, separately for tree species, calculated using the eff_size() function in the emmeans package (Lenth 2022). Standardized effect sizes were calculated by dividing pairwise differences in estimates by the SD of the experiments (with Osmia bee) and controls (without Osmia bee) providing the context for those differences. Zero Cohen d values signify no difference in response variables between experimental and control groups, while positive and negative g values indicate trends toward increased and decreased effect of bees on response variables respectively. Next, we visualized the predicted effect size with corresponding 95 % confidence intervals for tree species separately for response variables.
The analyses were carried out in R 4.0.2 (R Core Developmental Team 2020). GLMMs were carried out using the lme4 (Gaussian and negative binomial distribution) and glmmTMB (bet a distribution) packages (Bates et al. 2015; Brooks et al. 2017), and the data visualization using the ggplot2 package (Wickham 2009).
3 Results
All O. bicornis emerged from the cocoons exposed in Prunus and Quercus forest seed orchards in the spring, while in the summer, single cocoons remained unemerged in Tilia orchards. O. bicornis individuals nested in prepared artificial nests. They were also observed on experimental trees.
The generalized linear mixed models indicated that the proportion of viable seeds (p = 0.020) and the number of seeds (p = 0.009) in Tilia cordata significantly differed between the experimental and control groups (Figure 1). The number of seeds and the proportion of viable seeds were higher for trees additionally pollinated by O. bicornis. For the rest tree species, we did not find any significant differences in tested response variables (p > 0.05; Table I).
In Figure 2, we present the effect size (Cohen d) with corresponding 95 % confidence intervals for each tree species separately for tested response variables. The introduction of O. bicornis to forest seed orchards has a positive effect on the number of seeds and the proportion of viable seeds but the effects were not significant.

Effect size of supplementing pollination by O. bicornis on number of seeds, mass of thousand seeds (g), and proportion of viable seeds. The points represent mean effect sizes and 95 % confidence intervals.
4 Discussion
In this study, significant differences between treatments in two of three key characteristics of tree seed production were found. The number of seeds and the proportion of viable seeds were higher in the Tilia cordata trees where pollinating insects were supplemented with commercially reared Osmia bicornis. No differences emerged across pollination treatments on Quercus petraea and Prunus avium. Additional analyses showed that the introduction of bees to forest seed orchards has a positive effect on the number of seeds and the proportion of viable seeds.
Insect pollinators are a crucial component in providing vital ecosystem services to crops and wild plants, and the lack of pollinators frequently limits seed production. The decrease in the number of pollinating insects causes crop production to be increasingly dependent on managed pollinators. Only a few works is there information on the use of honey bees in the seed production of forest tree species (Loneragan 1979; Moncur et al. 1995; Hardi and Kanso 1995). Implementation of a native bee commercially developed and implemented for pollination can increase yield to meet the rising demand for forest products. To the best of our knowledge, this is the first field study in which the impact of solitary bee implementation in forest tree seed orchards has been studied for their effects on the seed yield. O. bicornis is widely distributed across Europe and is commonly used for commercial agriculture. Our research demonstrated that the red mason bee also has the potential as a forest orchard pollinator. It is thus possible to increase pollination and seed production with O. bicornis populations.
Moreover, because of its relatively short foraging range, O. bicornis would probably limit the pollination with pollen from outside the seed orchard compared to other commercial pollinators. Seed plantations are a particular type of forest cultivation established to produce seeds of high utility value used for afforestation (Kess and El - kassaby 2015). Pollen from individuals growing outside this area is considered contamination. Breeding procedures are limited to isolating seed plantations from potential sources of foreign pollen as effectively as possible (Adams and Burczyk 2000). With abundant food, O. bicornis bee flies 100–200 m from the nest. It is a generalist pollen forager collecting pollen from many plant families (Haider et al. 2013). However, their foraging behavior often shows a relatively high pollen constancy as these bees tend to prefer specific pollen sources when they are abundant near bee nesting sites (Radmacher and Strohm 2010; Sedivy and Dorn 2014). O. bicornis females specialize temporarily on plant species with high pollen availability to maximize pollen collection mass over time (Radmacher and Strohm 2010). This trait is advantageous for pollination in forest seed orchards. Moreover, Osmia bees forage at lower temperatures and in less favorable weather conditions and, therefore, for a more extended period per day than honey bees (Vicens and Bosch 2000). This feature is valuable for pollinating early blooming tree species such as cherry. Ryder et al. (2020) indicated that Prunus avium fruits from orchards with supplemented pollination by Osmia bicornis were larger and heavier at harvest than those produced with wild pollinators alone. An increase in fruit quality has been reported in cherry orchards when O. bicornis contributes to pollination (Kuhn and Ambrose 1984; Wilkaniec and Radajewska 1997). Our research also indicated a positive effect of supplementing O. bicornis on Prunus cultivation. However, a too - small sample size influenced the obtained results.
A strong preference for oak pollen is consistent with many studies on O. bicornis (e.g., Ruddle et al. 2018; Šlachta et al. 2020; Bednarska et al. 2022). Oak is wind - pollinated and offers large quantities of pollen. Other researchers have also confirmed the increased preference of red mason bees for anemophilous plants (Wilkaniec and Radajewska 1997; Teper 2007). As a wind - pollinated plant, oak is not dependent on pollination by insects. However, in the case of many wind - pollinating plants, pollinating insects may impact their yield. In our research, the presence of O. bicornis did not adversely affect the yielding of oak trees. We observed a slight trend of increasing the number and mass of seeds, but the results were not significant.
4.1 Implications on seed orchard management
The results of this study can be used for improved seed production of forest orchards. Our study indicates the potential use of the managed species of the wild bee as a complementary pollinator in seed orchards. Seed production in a seed orchard can be increased by supplemental mass pollination by red mason bees, thanks to an increase in the number of seeds and the proportion of viable seeds.
The food resources of forest environments are relatively low. Therefore, the bee density in forest environments is lower than in open habitats (Cierzniak 2003). This can pose a problem in the availability of an adequate number of pollinators in forest seed orchards that require pollinating insects. Low numbers of Apis mellifera and naturally occurring wild bees were recorded during the flowering period of examined forest seed orchards (unpublished data). That is why using managed species of solitary bees may be beneficial. High pollination efficiency, easy handling of the nests, and the lack of aggressiveness make it worth using Osmia bicornis as an alternative to honey bees. Nesting in large aggregations of these solitary bees and the possibility of using various artificial nesting materials are conducive to successful mass breeding. Moreover, the emergence of the adult can be manipulated to the flowering period of trees in forest orchards. Furthermore, managed honey bee colonies often suffer from increased prevalence of Varroa destructor parasites and viral, bacterial, and fungal diseases (Genersch 2010; Geslin et al. 2017), which, in turn, increases the costs of managing the hives, which takes both time and money. In the case of Osmia bicornis breeding, these problems are limited.
However, comprehensive and sustainable management should aim not only to improve pollination efficiency but also to maintain stable bee populations in the environment of forest seed orchards. That is why further research should also investigate the reproduction of O. bicornis in forest orchards to assess the costs and benefits of its commercial use and allow comparisons with alternatively managed pollinators. Also, more study is needed to make recommendations for ensuring the maintenance of supplementing pollination services by Osmia bicornis in forest ecosystems. Another issue is the impact of managed O. bicornis on local populations. Concern has arisen that managed bees may negatively affect wild bees (e.g., through competition with wild bees and pathogen spillover). Research on this subject is focused on the most common and widely used managed bees, honey bees and bumble bees. In contrast, the information concerning other managed bees, including O. bicornis, is limited (Mallinger et al. 2017). Moreover, other possible solutions for increasing pollinator populations in forest orchards require consideration. Trees in plantations provide a rich source of food, but for a limited time. For this reason, additional wildflower plantings can furnish wild bees with nectar and pollen throughout the season and ensure greater abundance and stability of pollinator populations, thus supporting forest seed orchard pollination. Moreover, decreasing pesticide use or increasing nesting sites can also be beneficial.
Data availability
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Code availability
Not applicable.
References
Adams WT, Burczyk J (2000) Magnitude and implications of gene flow in gene conservation reserves. Forest conservation genetics: principles and practice, 215–244
Allsopp MH, De Lange WJ, Veldtman R (2008) Valuing insect pollination services with cost of replacement. PLoS ONE 3(9):e3128
Anderson GJ (1976) The pollination biology of Tillia. Am J Bot 63(9):1203–1212
Bates D, Kliegl R, Vasishth S, Baayen H (2015) Parsimonious mixed models. arXiv preprint https://doi.org/arXiv:1506.04967
Batra SW (1978) Aggression, territoriality, mating and nest aggregation of some solitary bees (Hymenoptera: Halictidae, Megachilidae, Colletidae, Anthophoridae). J Kansas Entomol Soc 547–559
Bąk B, Wilde J (2003) Tempo rozwoju populacji murarki ogrodowej – Osmia rufa (L.) (Hymenoptera:Apidae) w sztucznych gniazdach trzcinowych. Wiad Entomol 22(3):161–167
Bednarska AJ, Mikołajczyk Ł, Ziółkowska E, Kocjan K, Wnęk A, Mokkapati JS, ... Laskowski R (2022) Effects of agricultural landscape structure, insecticide residues, and pollen diversity on the life - history traits of the red mason bee Osmia bicornis. Sci Total Environ 809:151142
Bosch J, Blas M (1994) Foraging behaviour and pollinating efficiency of Osmia cornuta and Apis mellifera on almond (Hymenoptera, Megachilidae and Apidae). Appl Entomol Zool 29(1):1–9
Bosch J, Kemp WP (2002) Developing and establishing bee species as crop pollinators: the example of Osmia spp. (Hymenoptera: Megachilidae) and fruit trees. Bull Entomol Res 92(1):3–16
Breeze TD, Bailey AP, Balcombe KG, Potts SG (2011) Pollination services in the UK: how important are honeybees? Agr Ecosyst Environ 142(3–4):137–143
Brooks ME, Kristensen K, Van Benthem KJ, Magnusson A, Berg CW, Nielsen A, ... Bolker BM (2017) glmmTMB balances speed and flexibility among packages for zero - inflated generalized linear mixed modeling. R Journal 9(2):378–400
Cierzniak T (2003) Ekologia pszczół w dynamicznym kręgu zbiorowisk grądowych. Wydaw. Akademii Bydgoskiej im. Kazimierza Wielkiego
Chałupka W, Matras J, Barżdajn W, Blonkowski S, Burczyk J, Fonder W, Grądzki T, Gryzło Z, Kacprzak P, Kowalczyk J, Kozioł C, Pytko T, Rzońca Z, Sabor J, Szeląg Z, Tarasiuk S (2011) Program zachowania leśnych zasobów genowych i hodowli selekcyjnej drzew w Polsce na lata 2011–2035. Centrum Informacyjne Lasów Państwowych. Warszawa 135–136. ISBN 978–83–61633–60–0
De Jaegere T, Hein S, Claessens H (2016) A review of the characteristics of small - leaved lime (Tilia cordata Mill.) and their implications for silviculture in a changing climate. Forests 7(3):56. https://doi.org/10.3390/f7030056.
Garibaldi LA, Aizen MA, Klein AM, Cunningham SA, Harder LD (2011) Global growth and stability of agricultural yield decrease with pollinator dependence. Proc Natl Acad Sci 108(14):5909–5914
Garibaldi LA, Steffan - Dewenter I, Winfree R, Aizen MA, Bommarco R, Cunningham SA, ... Klein AM (2013) Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 339(6127):1608–1611
Genersch E (2010) Honey bee pathology: current threats to honey bees and beekeeping. Appl Microbiol Biotechnol 87(1):87–97
Geslin B, Gauzens B, Baude M, Dajoz I, Fontaine C, Henry M, ... Vereecken NJ (2017) Massively introduced managed species and their consequences for plant–pollinator interactions. Adv Ecol Res 57:147–199 Academic Press
Goulson D, Nicholls E, Botías C, Rotheray EL (2015) Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347(6229):1255957
Gömöry D, Himanen K, Tollefsrud M, Uggla C, Kraigher H, Bordács S, ... Bozzano M (2020) Genetic aspects linked to production and use of forest reproductive material (FRM) collecting scientific evidence for developing guidelines and decision support tools for effective FRM management
Gruber B, Eckel K, Everaars J, Dormann CF (2011) On managing the red mason bee (Osmia bicornis) in apple orchards. Apidologie 42(5):564–576
Haider M, Dorn S, Müller A (2013) Intra - and interpopulational variation in the ability of a solitary bee species to develop on non - host pollen: implications for host range expansion. Funct Ecol 27(1):255–263
Hallmen M, van Leeuwen JFN (1990) The importance of oak (Quercus sp.) for a population of solitary bees Osmia rufa in the Hanau area [Germany]. Mitteilungen des Int Entomol Vereins e.V. 15(3 / 4):79–89
Hansted L, Grout BW, Toldam - Andersen TB, Eilenberg J (2014) An assessment of Osmia rufa (syn. bicornis) as a pollinator of the sour cherry (Prunus cerasus) cv. Stevnsbaer in eastern Denmark. J Apicult Res 53(1):177–182
Hardi T, Kanso A (1995) The role of European honey bees (Apis mellifera L.) in the pollination of a eucalypt plantation in Pati. Honeybee Science 16:24–26
Jabłoński B, Kołtowski Z, Szklanowska K, Teper D (2000) The investigations on flowering of lime trees. Pszczelarstwo 7:4–5
Kearns CA, Inouye DW, Waser NM (1998) Endangered mutualism: the conservation of plant - pollinator interactions. Annu Rev Ecol Syst 29:83–112
Kess T, El - Kassaby YA (2015) Estimates of pollen contamination and selfing in a coastal Douglas - fir seed orchard. Scand J for Res 30(4):266–275
Kevan PG, Viana BF (2003) The global decline of pollination services. Biodiversity 4(4):3–8
Klein AM, Vaissiere BE, Cane JH, Steffan - Deventer I, Cunningham SA, Kremen C, Tscharntke T (2007) Importance of pollinators in changing landscapes for worldcrops. Proc. R.Soc. London 274:303–313
Kocięcki S (1965) Plantacje nasienne ze szczepów - zakładanie i prowadzenie. Sylwan 109(1):51–63
Kowalczyk J, Markiewicz P, Chałupka W, Matras J (2011) Plantacje nasienne – rola i znaczenie w gospodarce leśnej. Las Polski 22:18–20
Krunic M, Pinzauti M, Felicioli A, Stanisavljevic LJ (1995) Further observations on Osmia cornuta Latr. and O.[Osmia] rufa L. as alternative fruit pollinators, domestication and utilization. Arch Biol Sci (Yugoslavia)
Krunić MD, Stanisavljević LŽ (2006) Supercooling points and diapause termination in overwintering adults of orchard bees Osmia cornuta and O. rufa (Hymenoptera: Megachilidae). Bull Entomol Res 96(3):323–326
Kuhn ED, Ambrose JT (1984) Pollination of ‘Delicious’ apple by megachilid bees of the genus Osmia (Hymenoptera: Megachilidae). J Kansas Entomol Soc 169–180
Lenth R (2022) Emmeans: estimated marginal means, aka least - squares means. R package version 1.8.2. https://doi.org/https:// CRAN.R - project.org / package = emmeans
Leonhardt SD, Gallai N, Garibaldi LA, Kuhlmann M, Klein AM (2013) Economic gain, stability of pollination and bee diversity decrease from southern to northern Europe. Basic Appl Ecol 14(6):461–471
Liesebach H, Liepe K, Bäucker C (2021) Towards new seed orchard designs in Germany–a review. Silvae Genetica 70(1):84–98
Loneragan OW (1979) Karri (Eucalyptus diversicolor F. Muell) phenological studies in relation to reforestation. For Dept West Aust Bull 90
Lye GC, Jennings SN, Osborne JL, Goulson D (2011) Impacts of the use of nonnative commercial bumble bees for pollinator supplementation in raspberry. J Econ Entomol 104(1):107–114
Mallinger RE, Gaines - Day HR, Gratton C (2017) Do managed bees have negative effects on wild bees?: a systematic review of the literature. PLoS ONE 12(12):e0189268
Moncur MW, Mitchell A, Fripp Y, Kleinschmidt GJ (1995) The role of honey bees (Apis mellifera) in eucalypt and acacia seed production areas. Commonw For Rev 350–354
Nanson A (1972) The provenance seedling seed orchard. Silvae Genetica 21:243–248
Pigott CD (1991) Biological flora of the British Isles. No. 174. Tilia cordata Miller. J Ecol 79(4):1147–1207
Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, Kunin WE (2010) Global pollinator declines: trends, impacts and drivers. Trends Ecol Evol 25(6):345–353
Radmacher S, Strohm E (2010) Factors affecting offspring body size in the solitary bee Osmia bicornis (Hymenoptera, Megachilidae). Apidologie 41:169–177
Radoglou K, Dobrowolska D, Spyroglou G, Nicolescu VN (2009) A review on the ecology and silviculture of limes (Tilia cordata Mill., Tilia platyphyllos Scop. and Tilia tomentosa Moench.) in Europe. Die Bodenkultur 60(3):7–17
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Roulston TM, Cane JH (2000) Pollen nutritional content and digestibility for animals. Plant Syst Evol 222:187–209
Ruddle N, Elston C, Klein O, Hamberger A, Thompson H (2018) Effects of exposure to winter oilseed rape grown from thiamethoxam - treated seed on the red mason bee Osmia bicornis. Environ Toxicol Chem 37(4):1071–1083
Ruotsalainen S (2014) Increased forest production through forest tree breeding. Scand J for Res 29(4):333–344
Ryder JT, Cherrill A, Prew R, Shaw J, Thorbek P, Walters KF (2020) Impact of enhanced Osmia bicornis (Hymenoptera: Megachilidae) populations on pollination and fruit quality in commercial sweet cherry (Prunus avium L.) orchards. J Apicult Res 59(1):77–87
Saunders ME (2018) Insect pollinators collect pollen from wind - pollinated plants: implications for pollination ecology and sustainable agriculture. Insect Conservation and Diversity 11(1):13–31
Sedivy C, Dorn S (2014) Towards a sustainable management of bees of the subgenus Osmia (Megachilidae; Osmia) as fruit tree pollinators. Apidologie 45(1):88–105
Šlachta M, Erban T, Votavová A, Bešta T, Skalský M, Václavíková M, ... Cudlín P (2020) Domestic gardens mitigate risk of exposure of pollinators to pesticides—an urban - rural case study using a red mason bee species for biomonitoring. Sustainability 12(22):9427
Smithson M, Verkuilen J (2006) A better lemon squeezer? Maximum - likelihood regression with beta - distributed dependent variables. Psychol Methods 11(1):54
Steffan - Dewenter I, Schiele S (2008) Do resources or natural enemies drive bee population dynamics in fragmented habitats. Ecology 89(5):1375–1387
Teper D (2007) Food plants of the red mason bee (Osmia rufa L.) determined based on a palynological analysis of faeces. Journal of Apicultural Science 51:55–62
Torchio PF, Asensio E (1985) The introduction of the European bee, Osmia cornuta Latr., into the US as a potential pollinator of orchard crops, and a comparison of its manageability with Osmia lignaria propinqua Cresson (Hymenoptera: Megachilidae). J Kansas Entomol Soc 42–52
Vicens N, Bosch J (2000) Weather - dependent pollinator activity in an apple orchard, with special reference to Osmia cornuta and Apis mellifera (Hymenoptera: Megachilidae and Apidae). Environ Entomol 29(3):413–420
Welk E, De Rigo D, Caudullo G (2016) Prunus avium in Europe: distribution, habitat, usage and threats. European Atlas of Forest Tree Species. Publ. Off, EU, Luxembourg, p 3
Weryszko - Chmielewska E, Sadowska DA (2010) The phenology of flowering and pollen release in four species of linden (Tilia L.). J Apicult Sci 54(2):99–108
Wickham H (2009) Getting started with qplot. In ggplot2 (pp. 9–26). Springer, New York, NY
Wilkaniec Z, Radajewska B (1997) Solitary bee Osmia rufa L. (Apoidea, Megachilidae) as pollinator of strawberry cultivated in an unheated plastic tunnel. Acta Hortic 439(1):227–234
Załęski A, Aniœko E, Kantorowicz W, Sobczak H (2000) Zasady i metodyka oceny nasion w Lasach Państwowych. CILP, Warszawa 180
Zhelev P, Tsvetanov N, Aneva I, Ivanov D (2020) Diversity, genetic resources and conservation of tilia species in Bulgaria. Forestry 26(2):503–513
Funding
Research conducted by the statutory funding No. 506-511-04-00 of the Faculty of Veterinary Medicine and Animal Science Poznań University of Life Sciences, Poland; Department of Zoology.
Author information
Authors and Affiliations
Contributions
MK: methodology, investigation, writing – original draft. WBC: conceptualization, writing – original draft. ŁD: statistical analysis, writing – review and editing. MF: conceptualization, methodology, supervision.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Manuscript editor: Peter Rosenkranz
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
Cite this article
Kęsy, M., Banaszak - Cibicka, W., Dylewski, Ł. et al. Effect of Osmia bicornis supplemental pollination on seed yield of forest seed orchards. Apidologie 54, 32 (2023). https://doi.org/10.1007/s13592-023-01013-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13592-023-01013-z