
Abstract
Fruit cracking caused by rainfall prior to harvest, a major problem in sweet cherry production, is being exacerbated by climate change. Currently, pre-harvest spraying with calcium salt solutions is the prevalent technique to reduce fruit cracking in cherry orchards not covered by plastic roofs. This study evaluated the effectiveness of canopy-applied silicon in the reduction of sweet cherry cracking under different field conditions. Four field trials were conducted on mature trees of the cultivars Van, New Star, and Emperor Francis. Treatments included water (control), calcium chloride, and sodium silicate. Multiple sprays (three) were applied weekly from fruit onset of color to approximately 1 week before harvest. The results showed that under conditions conducive to cracking, sodium silicate reduced the percentage of cracked fruits to a similar or larger extent than calcium chloride. This study highlights how canopy-applied silicon sources may effectively contribute to reducing cherry cracking, acting as an alternative technique to other preventive methods.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Fruit cracking is a physiological disorder that can dramatically depreciate cherry fruits, causing heavy economic losses starting from the fruit’s onset of color (Sekse 1995; Sekse et al. 2005; Simon 2006; Knoche and Winkler 2017; Correia et al. 2018). Recently, climate change has intensified the occurrence and damages caused by rain-induced cracking in sweet cherry and other important horticultural crops. Fruits are particularly vulnerable to cracking from onset of color to maturity, mostly in conjunction with high atmospheric humidity or rainfall events (Christensen 1996).
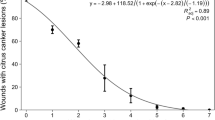
The harvest value is extremely compromised when the incidence of cracking is higher than 20–30% (Hansen and Proebsting 1996). This is recurrent for susceptible genotypes, such as cv. Van, where cracking can reach almost 100% of the marketable fruits. Moreover, lesions localized on the drupe represent a preferential access route to the spread of pathogens such as Monilinia laxa and Botrytis cinerea (Børve et al. 2000), causing additional losses.
Many hypotheses have been proposed to explain the phenomenon of fruit cracking, even though the responsible mechanisms are not completely elucidated (Correia et al. 2018).
The incidence and severity of cherry cracking are influenced by genotypic, environmental, and agronomic factors (Correia et al. 2018). The susceptibility to cracking differs largely among cherry cultivars (Sekse 1995; Christensen 1996; Demirsoy and Demirsoy 2004; Measham et al. 2009; Correia et al. 2018) and can be modulated by the rootstock (Cline et al. 1995; Sekse 1995; Simon et al. 2004). In addition, the fruit cuticle plays a crucial role in cracking susceptibility (Martin and Rose 2014; Tafolla-Arellano et al. 2018; Wang et al. 2021).
The reduction of cherry cracking often involves the use of physical-mechanical systems (plastic roofs, helicopters, or air blast blowers), chemicals (calcium-based compounds, mineral sprays, antitranspirants, and growth regulators), or biostimulants (e.g., seaweed extracts) (Sekse 1995; Sekse et al. 2005; Simon 2006; Knoche and Winkler 2017; Correia et al. 2018). Plastic roofs reduce cracking damage but are costly (Simon 2006; Correia et al. 2018).
Pre-harvest calcium sprays generally reduce sweet cherry cracking (Demirsoy and Bilgener 1998; Sekse 1998), as calcium plays a critical role in maintaining the structural integrity and firmness of the cell wall of the fruits (Christensen 1972; Sekse 1995). However, calcium sprays can reduce fruit size (Looney 1986; Facteau et al. 1987).
The techniques used to reduce fruit cracking are hardly effective when its incidence is high (e.g. from frequent rainfall) and present some drawbacks. Consequently, it is necessary to develop agronomic strategies that prevent the final appearance of macrocracks, without generating adverse impacts on the fruits and the tree. Modifications of cell wall properties (extensibility and stability) of the epidermis and the preservation of fruit cuticles by agronomic means are likely to contribute to reduced cracking.
Silicon (Si) is a beneficial element whose positive effects are mainly associated with its high deposition in plant tissues, enhancing their strength and rigidity (Ma and Takahashi 2002; Ma and Yamaji 2006). Si is capable of increasing cell wall extensibility and thereby the elongation process in the meristematic zone (Hossain et al. 2002; Hattori et al. 2003; Bat-Erdene et al. 2021). In addition, Si forms a protective layer that prevents the penetration of water and fungal pathogens and stimulates defense reactions against pathogens (Ma and Takahashi 2002; Ma and Yamaji 2006; Bat-Erdene et al. 2021).
The objective of this study is to evaluate the effectiveness of canopy-applied Si sources in the reduction of sweet cherry cracking through a range of field experiments under different environmental conditions.
2 Materials and methods
2.1 Plant material and growing conditions
Sweet cherry (Prunus avium) mature trees of cultivars Van, Emperor Francis, and New Star were subjected to canopy-applied treatment in four experimental sites, of which three were located in Northern Italy and one in Upper Midwestern, United States. Meteorological data for the period between onset of color and harvest are presented in Table 1.
Experiment 1 was conducted in 2002 in a mature cherry orchard located in Savignano sul Panaro (Modena, Italy) (44°29’N 11°02’E; 217 m a.s.l.) on plants of the cv. Van grafted on Colt.
Experiment 2 was conducted in 2003 in a mature cherry orchard located at the Clarksville Research Center, Michigan State University (Clarksville, Michigan, USA) (42°52’N 85°15’W; 251 m a.s.l.) on plants of the cv. Emperor Francis grafted on a cherry seedling.
Experiment 3 was carried out in 2005 in Vignola (Modena, Italy), (44°28’N 11°00’E; 125 m a.s.l.), in a mature orchard, on plants of cv. Van grafted on a cherry seedlings.
Experiment 4 was carried out in 2007 in Castelfranco Emilia (Modena, Italy), (44°34’N 11°05’E; 35 m a.s.l.) in a mature orchard, on plants of cv. New Star grafted on a cherry seedling.
2.2 Canopy-applied calcium and silicon treatments
Field experiments were conducted, during one growing season, in a completely randomized block design with three replicates. The following treatments were compared: (1) control (distilled water), (2) calcium chloride (CaCl2) (5 g L−1), and (3) sodium silicate (Na2OSiO2)3 (36–40°Bé) (2.3 g L−1). Each treatment was applied to a single branch within a tree. The pH of all solutions was adjusted to 5.5 using HCl. Solutions were applied in the late afternoon to the canopy through pressurized nebulizers, until the complete dripping of fruits and leaves.
Three (Experiments 1–3) and four (Experiment 4) randomly selected trees were sprayed for each treatment and separated by at least two untreated trees along consecutive plots. Treatments were performed weekly from the onset of color until commercial harvest, for a total of three applications.
2.3 Sampling and measurement methods
The effects of treatments were evaluated at harvest. The percentage of cracked fruits was calculated based on the total number of treated fruits. In Experiments 1 and 2, the types of cracks (peduncular, dorsal, and apical end) were also assessed.
2.4 Fruit quality parameters
Fruit quality was evaluated, for all experiments, by fruit weight and soluble solids content (TSS) (Atago Digital Refractometer, Optolab, Modena, Italy). Fruit firmness (Effegi FT 011, tip ø 5 mm, Ravenna, Italy) (Experiments 1 and 4), pH, and juice titratable acidity (Experiment 4) (Crison Titromatic 1 S, Barcelona, Spain) were also determined.
Cherry skin color parameters were determined (Experiment 1) via a chromameter (CR-200 Chromameter, Minolta Co. Ltd., Osaka, Japan) using the digital representation model HLS (hue, lightness, and saturation). The hue axis (H) includes red at 0° and 360°. The brightness axis (L) ranges from 0 (black) to 100 (white). The saturation axis (S) ranges from 0 (darker color) to 100 (more intense color).
2.5 Silicon localization by cryo-scanning electron microscopy and energy-dispersive X-ray microanalysis
Analysis of the deposition of Si on the surface of the fruits and its possible penetration into the internal tissues was performed on the frozen-hydrated, whole, and freeze-fractured samples of cvs. Van (Experiment 3) and New Star (Experiment 4) by cryo-scanning electron microscopy (Cryo-SEM) and energy-dispersive X-ray microanalysis (McCully et al. 2009, 2010; Minnocci et al. 2018). At harvest, a portion of the surface and a parallelepiped of the regular Sect. (5 × 5 mm) from the epidermis until the seed of unwashed fruits (without stalk) were sampled. The fruit portions were immediately cryo-fixed by immersion in liquid nitrogen (−196 °C) and stored until Cryo-SEM analysis. For the internal tissue analysis, the frozen-hydrated (FH) samples were first mounted under liquid nitrogen gas in an aluminum stub with Tissue-Tek, freeze-fractured inside the liquid nitrogen to expose the internal texture, transferred to a dedicated cryo-preparation chamber (SEM Cryo Unit, SCU 020, Bal-Tech, Balzers, Liechtenstein), surface etched for 3 min at −80 °C under high vacuum (P < 2 × 10− 4 Pa), and sputter-coated with 8-nm gold in an argon atmosphere (P < 2.2 × 10− 2 Pa) to produce an electrically conductive surface. FH specimens were finally transferred to the cryo-stage (−180 °C) inside a scanning electron microscope (Philips SEM 515, Eindhoven, the Netherlands). Energy-dispersive X-ray microanalysis of FH specimens was performed with SEM using an acceleration voltage of 17 kV, a take-off angle of 16.5°, and a working distance (sample to the final lens of the SEM instrument) of 12.0 mm. Spectra from 0 to 20 keV were acquired for 120 s (live time) with a dead time of less than 20%, and collected at increments of 10 eV per channel with the electron beam focused on a spot area (diameter 40 nm) in the center of the selected cells (Fig. 1D). The background and element-specific peak spectra were analyzed using the program EDAX DX-4 2.0 (EDAX, San Francisco, CA), which fully deconvolutes the spectra and allows corrections for interference between elements (Servili et al. 2008).

(A): cv. Van, sodium silicate residue on the outer surface of the treated fruit; (B): cv. New Star, transverse freeze-fracture of exocarp in correspondence of a residual drop of sodium silicate; (C) the same plane of fracture of (B) at a lower magnification to display the different cell types examined with X-ray microanalysis; and (D) an X-ray spectrum representative of those detected both in cells of the epidermis and in the mesocarp, with the silicon content (SiK) highlighted in green
2.6 Statistical analysis
Data were subjected to analysis of variance, and the comparison between treatments was performed by the Student Newman Keuls (SNK) test (P ≤ 0.05).
3 Results
3.1 Effects of canopy-applied sodium silicate and calcium chloride on sweet cherry cracking
3.1.1 Experiment 1
The application of Si and CaCl2 reduced to a similar extent the incidence of cracked fruits compared to water-sprayed fruits (Table 2). Most fruits showed cracks in the apical portion, and the type of cracking was not affected by treatments (Table 2). The application of sodium silicate to the canopy decreased the average weight of the fruit at harvest in comparison to control plants (Table 3), whereas fruit firmness was not affected by treatments (Table 3). Si treatment reduced hue, brightness, and saturation (Table 4).
3.1.2 Experiment 2
The highest percentage of cracked fruits was recorded in control plants. This value was reduced, with similar effectiveness, in the branches treated with sodium silicate and calcium salt (Table 2). In addition, differences were observed also in the type of cracking (Table 2). The supply of calcium chloride to the canopy reduced the average weight of the fruit and the soluble solids, while the sodium silicate increased the latter parameter (Table 3).
3.1.3 Experiment 3
The low rainfall that occurred during the experiment (Table 1) prevented the occurrence of cracking. Negligible cracking incidence was observed (less than 0.5%; data not shown), and it was not influenced by treatment. Similarly, treatments did not affect fruit weight at harvest (Table 3). Nevertheless, total soluble solids were enhanced by canopy-applied sodium silicate (Table 3).
3.1.4 Experiment 4
Cracked fruits were higher than 50%, except in plants treated with the Si source (Table 2). Only the latter treatment reduced the percentage of damaged fruit compared to the control, whereas calcium chloride was not effective. On cracked fruits, most of the fractures were localized in the peduncular position, with no differences induced by treatments (Table 2). The analysis of the main quality parameters did not show differences due to treatments (Table 3).
3.2 Cryo-SEM and energy-dispersive X-ray microanalysis of fruits subjected to sodium silicate treatments
The analysis of frozen-hydrated samples of cherry fruits subjected to sodium silicate treatments by Cryo-SEM and energy-dispersive X-ray microanalysis demonstrated the presence of Si deposits on the surface of the fruits (Fig. 1A). The assay determined that even with the presence of sodium silicate on the outer surface of the fruit in the form of droplets remained following treatment (Fig. 1A and B), the cells below the cuticle do not show a detectable amount of Si (Fig. 1C), whose silicon X-ray signal is always maintained below the level of the background (Fig. 1D, SiK, in green). This was observed by analyzing the cells of the mesocarp (Fig. 1C) and those of the epidermis, even at a distance of 50–60 μm from the drops of sodium silicate remaining on the surface (Fig. 1B). The same results were obtained from the analysis of frozen-hydrated samples of cherry fruits subjected to calcium chloride treatments (data not shown).
4 Discussion
Our results, obtained in different experimental conditions conducive to cracking, showed that canopy-applied sodium silicate during fruit ripening was effective in reducing sweet cherry cracking at harvest (Table 2). The effectiveness of sodium silicate was similar or higher than calcium chloride (Table 2), the most adopted technique for mitigating cracking.
In sweet cherry, three types of cracking have been described (Christensen 1996): in the cheek, in the apical end, and in the stem cavity region of the fruit. Multiple cracking, resulting from the combination of these three types, often occurs. Our data suggest that meteorological conditions influenced the incidence and type of cracking (Tables 1 and 2). Data indicate that high rainfall (Table 1) increased the occurrence of cracking in the stem cavity (Table 2). When low rainfall and low humidity occurred, no fruit cracking was observed. However, when rainfall was low but high humidity was recorded (Table 1), cracking was observed with the dominant type of cracking in the apical region (Table 2).
The results obtained via cryo-SEM and energy-dispersive X-ray microanalysis indicate that Si was localized only on the surface of the fruit, without any significant detection of its penetration into internal tissues (Fig. 1). However, some Si in traces, at concentrations below the resolution capability of the instrument, could be present in the observed tissues.
Although Si is not considered an essential element for plant nutrition and its questionable role in plant biology have been reported (Coskun et al. 2019), many authors have shown its beneficial effects in a variety of species and environmental conditions (Ma and Yamaji 2006; Coskun et al. 2019). The favorable effects of Si on the crop are often associated with the reinforcement of the cell walls due to the deposition of Si in the form of amorphous silica and opal phytoliths (Epstein 1999). Si displays a notable ability to increase the extensibility of the cell wall, a prerequisite for growth (Ma and Takahashi 1993). The application of Si to rice seedlings promoted plant growth mainly by enhancing cell elongation of the epidermal cells in the young shoot tissues and increased cell wall extensibility (Hossain et al. 2002).
When treated with Si, cell walls of sorghum roots were strengthened in the mature basal region, and cell walls in the apical and subapical zones showed an increase in extensibility, which resulted in a promotion of root elongation (Hattori et al. 2003). Si has also been reported to improve tissue extensibility and enhance cell enlargement in cucumber plants under salt and drought stress (Ouzounidou et al. 2016).
Si can modify the mechanical properties of tissues, contributing to cell wall rigidity and reinforcement, as it interacts with primary cell wall constituents (Currie and Perry 2007); at the same time, it increases the elasticity of the cell wall during distension and extension growth (Pilon-Smits et al. 2009).
Our data (Table 3; Fig. 1) indicate that canopy-applied Si did not build up a protective film covering the whole fruit surface, which would have prevented water exchanges between the fruit and the outside slowing down phloem translocation and consequent accumulation of sugars in the fruit. Such effects were not recorded in any of the experiments (Table 3).
The reduction of cracks also plays a role in the reduction of pathogen incidence (Børve et al. 2000). Therefore, Si applied to the canopy could also reduce the incidence of monilia (Monilinia spp.) and Botrytis cinerea on fruits. Si forms a physical barrier and stimulates defense reactions against pathogens (Ma and Takahashi 2002; Ma and Yamaji 2006; Wang et al. 2017; Bat-Erdene et al. 2021).
Additionally, Si treatment influenced color (Experiment 1; Table 4), reducing hue and brightness. Chromatic functions of chroma and hue are negatively correlated with anthocyanin levels in the fruit skin (Gonçalves et al. 2007), indicating higher concentrations of anthocyanins in the berry skin. In apples, canopy-applied Si has been shown to stimulate anthocyanin accumulation and induce skin color traits (Karagiannis et al. 2021).
Canopy-applied Si resulted in a significant increase in total soluble solids of cherry fruits (Experiments 2 and 3; Table 3), an effect attributable to the stimulating action of Si on the leaf photosynthetic activity (Ma and Takahashi 2002; Ma and Yamaji 2006; Bat-Erdene et al. 2021).
Research outcomes on fruit cracking highlight the relevance of mechanical properties of fruit epidermal cell wall strength, overall extensibility, elasticity, and cuticular membrane composition (Alkio et al. 2012; Balbontín et al. 2014; Quero-García et al. 2021). An elastic epidermis will increase the tolerance to cracking since it is more capable of accommodating the increase in flesh volume during fruit growth and reducing the formation of cuticular fractures. Si appears to be a good candidate to achieve both of these conditions in the fruit. Therefore, these parameters should be included in further experiments.
5 Conclusions
Fruit cracking is a serious concern for cherry growers worldwide. Si is a beneficial element, known for its effectiveness against various biotic and abiotic stresses and therefore of interest in sustainable cherry cultivation. The current study highlights how canopy-applied Si sources may effectively contribute to reducing cherry cracking, acting as an alternative technique to other preventive methods. Future studies should gain further knowledge on Si mode of actions and Si nutrition in different cherry rootstocks and cultivars.
References
Alkio M, Jonas U, Sprink T, van Nocker S, Knoche M (2012) Identification of putative candidate genes involved in cuticle formation in Prunus avium (sweet cherry) fruit. Ann Bot 110:101–112. https://doi.org/10.1093/aob/mcs087
Balbontín C, Ayala H, Rubilar J, Cote J, Figueroa CR (2014) Transcriptional analysis of cell wall and cuticle related genes during fruit development of two sweet cherry cultivars with contrasting levels of cracking tolerance. Chil J Agric Res 74:162–169. https://doi.org/10.4067/S0718-58392014000200006
Bat-Erdene O, Szegő A, Gyöngyik M, Mirmazloum I, Papp I (2021) Effects of silicon in plants with particular reference to horticultural crops - review article. Int J Hortic Sci 27. https://doi.org/10.31421/ijhs/27/2021/9096
Børve J, Sekse L, Stensvand A (2000) Cuticular fractures promote postharvest fruit rot in sweet cherries. Plant Dis 84:1180–1184. https://doi.org/10.1094/PDIS.2000.84.11.1180
Christensen JV (1972) Cracking in cherries: III. Determination of cracking susceptibility. Acta Agric Scand 22:128–136. https://doi.org/10.1080/00015127209433471
Christensen JV (1996) Rain-induced cracking of sweet cherries: its causes and prevention. In: Webster AD, Looney NE (eds) Cherries: Crop Physiology, production and uses. CAB International, UK, pp 297–327
Cline JA, Meland M, Sekse L, Webster AD (1995) Rain cracking of sweet cherries: II. Influence of rain covers and rootstocks on cracking and fruit quality. Acta Agric Scand Sect B — Soil Plant Sci 45:224–230. https://doi.org/10.1080/09064719509413108
Correia S, Schouten R, Silva AP, Gonçalves B (2018) Sweet cherry fruit cracking mechanisms and prevention strategies: a review. Sci Hortic 240:369–377. https://doi.org/10.1016/j.scienta.2018.06.042
Coskun D, Deshmukh R, Sonah H, Menzies JG, Reynolds O, Ma JF, Kronzucker HJ, Bélanger RR (2019) The controversies of silicon’s role in plant biology. New Phytol 221:67–85. https://doi.org/10.1111/nph.15343
Currie HA, Perry CC (2007) Silica in plants: biological, biochemical and chemical studies. Ann Bot 100:1383–1389. https://doi.org/10.1093/aob/mcm247
Demirsoy LK, Bilgener S (1998) The effects of preharvest calcium hydroxide applications on cracking in 0900 ‘Ziraat’, ‘Lambert’ and ‘Van’ sweet cherries. Acta Hortic 657–662. https://doi.org/10.17660/ActaHortic.1998.468.82
Epstein E (1999) Silicon. Annu Rev Plant Physiol. Plant Mol Biol 50:641–664. https://doi.org/10.1146/annurev.arplant.50.1.641
Facteau TJ, Rowe KE, Chestnut NE (1987) Response of Bing and Lambert sweet cherry fruit to preharvest calcium chloride applications. HortScience 22:271–273
Gonçalves B, Silva AP, Moutinho-Pereira J, Bacelar E, Rosa E, Meyer AS (2007) Effect of ripeness and postharvest storage on the evolution of colour and anthocyanins in cherries (Prunus avium L). Food Chem 103:976–984. https://doi.org/10.1016/j.foodchem.2006.08.039
Hansen EJ, Proebsting EL (1996) Cherry nutrient requirements and water relations. In: Webster AD, Looney NE (eds) Cherries: Crop Physiology, production and uses. CAB International, UK, pp 243–257
Hattori T, Inanaga S, Tanimoto E, Lux A, Luxová M, Sugimoto Y (2003) Silicon-induced changes in viscoelastic properties of sorghum root cell walls. Plant Cell Physiol 44:743–749. https://doi.org/10.1093/pcp/pcg090
Hossain MT, Mori R, Soga K, Wakabayashi K, Kamisaka S, Fujii S, Yamamoto R, Hoson T (2002) Growth promotion and an increase in cell wall extensibility by silicon in rice and some other Poaceae seedlings. J Plant Res 115:0023–0027. https://doi.org/10.1007/s102650200004
Karagiannis E, Michailidis M, Skodra C, Molassiotis A, Tanou G (2021) Silicon influenced ripening metabolism and improved fruit quality traits in apples. Plant Physiol Biochem 166:270–277. https://doi.org/10.1016/j.plaphy.2021.05.037
Knoche M, Winkler A (2017) Rain-induced cracking of sweet cherries. In: Quero-García J, Lezzoni A, Puławska J, Lang G (eds) Cherries: botany, production and uses. CABI, Wallingford, pp 140–165
Looney NE (1986) Benefits of calcium sprays below expectations in BC tests. Goodfruit Grower 36:7–8
Ma JF, Takahashi E (2002) Soil, fertilizer, and plant silicon research in Japan. Elsevier, Amsterdam; Boston
Ma JF, Takahashi E (1993) Interaction between calcium and silicon in water-cultured rice plants. Plant Soil 148:107–113. https://doi.org/10.1007/BF02185390
Ma JF, Yamaji N (2006) Silicon uptake and accumulation in higher plants. Trends Plant Sci 11:392–397. https://doi.org/10.1016/j.tplants.2006.06.007
Martin LBB, Rose JKC (2014) There’s more than one way to skin a fruit: formation and functions of fruit cuticles. J Exp Bot 65:4639–4651. https://doi.org/10.1093/jxb/eru301
McCully ME, Canny MJ, Huang CX, McCully ME, Canny MJ, Huang CX (2009) Cryo-scanning electron microscopy (CSEM) in the advancement of functional plant biology. Morphological and anatomical applications. Funct Plant Biol 36:97–124. https://doi.org/10.1071/FP08304
McCully ME, Canny MJ, Huang CX, Miller C, Brink F, McCully ME, Canny MJ, Huang CX, Miller C, Brink F (2010) Cryo-scanning electron microscopy (CSEM) in the advancement of functional plant biology: energy dispersive X-ray microanalysis (CEDX) applications 1. Funct Plant Biol 37:1011–1040. https://doi.org/10.1071/FP10095
Measham PF, Bound SA, Gracie AJ, Wilson SJ, Measham PF, Bound SA, Gracie AJ, Wilson SJ (2009) Incidence and type of cracking in sweet cherry (Prunus avium L.) are affected by genotype and season. Crop Pasture Sci 60:1002–1008. https://doi.org/10.1071/CP08410
Minnocci A, Francini A, Romeo S, Sgrignuoli AD, Povero G, Sebastiani L (2018) Zn-localization and anatomical changes in leaf tissues of green beans (Phaseolus vulgaris L.) following foliar application of Zn-lignosulfonate and ZnEDTA. Sci Hortic 231:15–21. https://doi.org/10.1016/j.scienta.2017.12.002
Ouzounidou G, Giannakoula A, Ilias I, Zamanidis P (2016) Alleviation of drought and salinity stresses on growth, physiology, biochemistry and quality of two Cucumis sativus L. cultivars by Si application. Braz J Bot 39:531–539. https://doi.org/10.1007/s40415-016-0274-y
Pilon-Smits EA, Quinn CF, Tapken W, Malagoli M, Schiavon M (2009) Physiological functions of beneficial elements. Curr Opin Plant Biol 12:267–274. https://doi.org/10.1016/j.pbi.2009.04.009
Sekse L (1995) Fruit cracking in sweet cherries (Prunus avium L.). Some physiological aspects—a mini review. Sci Hortic 63:135–141. https://doi.org/10.1016/0304-4238(95)00806-5
Sekse L (1998) Fruit cracking mechanisms in sweet cherries (Prunus avium L.) - a review. Acta Hortic 637–648. https://doi.org/10.17660/ActaHortic.1998.468.80
Sekse L, Bjerke KL, Vangdal E (2005) Fruit cracking in sweet cherries - an integrated approach. Acta Hortic 471–474. https://doi.org/10.17660/ActaHortic.2005.667.68
Servili M, Minnocci A, Veneziani G, Taticchi A, Urbani S, Esposto S, Sebastiano L, Valmorri S, Corsetti A (2008) Compositional and tissue modifications induced by the natural fermentation process in table olives. J Agric Food Chem 56(15):6389–6396. https://doi.org/10.1021/jf8007019
Simon G (2006) Review on rain induced fruit cracking of sweet cherries (Prunus avium L.), its causes and the possibilities of prevention. Int J Hortic Sci 12:27–35. https://doi.org/10.31421/IJHS/12/3/654
Simon G, Hrotkó K, Magyar L (2004) Fruit quality of sweet cherry cultivars grafted on four different rootstocks. Acta Hortic 658:365–370. https://doi.org/10.17660/ActaHortic.2004.658.53
Tafolla-Arellano JC, Báez-Sañudo R, Tiznado-Hernández ME (2018) The cuticle as a key factor in the quality of horticultural crops. Sci Hortic 232:145–152. https://doi.org/10.1016/j.scienta.2018.01.005
Wang M, Gao L, Dong S, Sun Y, Shen Q, Guo S (2017) Role of silicon on plant–pathogen interactions. Front Plant Sci 8:701. https://doi.org/10.3389/fpls.2017.00701
Wang Y, Guo L, Zhao X, Zhao Y, Hao Z, Luo H, Yuan Z (2021) Advances in mechanisms and omics pertaining to fruit cracking in horticultural plants. Agronomy 11:1045. https://doi.org/10.3390/agronomy11061045
Acknowledgements
Authors gratefully acknowledge Nino Quartieri farm (Savignano sul Panaro, MO), Neri-Vezzali farm (Vignola, MO), Testoni farm (Castelfranco Emilia, MO), and the Research Center of the Michigan State University (Michigan, USA) for hosting the field trials. The authors kindly acknowledge Prof. Dr. Jian Feng Ma and Dr. Alessandra Lombini for the critical reading of the manuscript.
Funding
Open access funding provided by Alma Mater Studiorum - Università di Bologna within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Contributions
ADR conceived the study, designed and performed the experiments, and wrote the manuscript. MQ contributed to performing the experiment and to the revision of the manuscript. ARD contributed to the writing of the manuscript. AM performed the Cryo-SEM and energy-dispersive X-ray analysis and interpretation and contributed to the writing. LS conceptualized the Cryo-SEM and energy-dispersive X-ray analysis and contributed to the revision of the manuscript. GS performed the experiments and contributed to the revision of the manuscript. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Communicated by Heakeun Yun.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rombolà, A., Quartieri, M., Rodríguez-Declet, A. et al. Canopy-applied silicon is an effective strategy for reducing sweet cherry cracking. Hortic. Environ. Biotechnol. 64, 371–378 (2023). https://doi.org/10.1007/s13580-022-00486-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13580-022-00486-8