
Abstract
Extrinsic and intrinsic factors may influence the activity budget of wild animals, resulting in a variation in the time spent in different activities among populations or individuals of the same species. In this study, we examined how extrinsic and intrinsic factors affect the behaviour of the alpine marmot (Marmota marmota), a hibernating social rodent inhabiting high-elevation prairies in the European Alps. We collected behavioural observations during scan sampling sessions on marked individuals at two study sites with different environmental characteristics. We used Bayesian hierarchical multinomial regression models to analyse the influence of both intrinsic (sex and age-dominance status) and extrinsic (environmental and climatic variables) factors on the above-ground activity budget. Marmots spent most of their time above ground foraging, and were more likely to forage when it was cloudy. Extrinsic factors such as the site, period of the season (June, July–August, and August–September), and time of the day were all related to the probability of engaging in vigilance behaviour, which reaches its peak in early morning and late afternoon and during July, the second period included in the study. Social behaviours, such as affiliative and agonistic behaviours, were associated mostly with sex and age-dominance status, and yearlings were the more affiliative individuals compared to other status. Overall, our results suggest that in alpine marmots, intrinsic factors mostly regulate agonistic and affiliative behaviours, while extrinsic factors, with the unexpected exception of temperature, affect the probabilities of engaging in all types of behavioural categories.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In animals, activity budget allocation is strongly influenced by multiple factors, such as energetic demands, food abundance, predation pressure, and climatic conditions (Belovsky & Slade 1986; Ebensperger & Hurtado 2005; Armitage & Salsbury 2016). In various mammal species, both extrinsic factors, such as climatic variability, environmental characteristics, season, and group size (Armitage et al. 1996; Arnold et al. 2006; Hannon et al. 2006; Bourgoin et al. 2008; Scantlebury et al. 2012), and intrinsic factors, such as age, sex, reproductive status, and dominance status (Oakes et al. 1992; Hamel & Coté 2008; Aniano et al. 2016; Armitage & Salsbury 2016), have been shown to influence activity budgets. The time each individual allocates to any activity has been shown to be the result of a trade-off between the necessity to fulfil its energetic requirement (i.e. foraging) and the need to devote enough time to other activities (e.g. vigilance or social behaviours; Ebensperger & Hurtado 2005).
We expect this trade-off to be particularly severe in animals living at high elevations where extrinsic factors may be more severe and demanding (Laszlo & Grabherr 2009), limiting an individual’s flexibility in adapting its activity budget. High temperatures during summers and adverse weather conditions have been shown to affect the activity patterns of alpine ungulates: for example, Alpine ibex (Capra ibex) migrate to areas at higher elevations when temperatures rise, despite these areas being characterized by lower quality vegetation (Mason et al. 2017), and chamois (Rupicapra rupicapra) reduce foraging activity during intense rain or very hot days (Pęska & Ciach 2018). On the other hand, intrinsic characteristics have been shown to affect several aspects of an individual’s activity budget: male and female ungulates, for example, may differ in their foraging rate. In bighorn sheep (Ovis canadensis), ewes spend more time foraging and walking compared to rams (Ruckstuhl 1998), while Appenine chamois (Rupicapra pyrenaica ornata) males show a higher intensity of foraging compared to females (Ferretti et al. 2014). In mountain goats (Oreamnos americanus), lactating females tend to spend more time feeding compared to non-lactating females, because of their higher energetic demands (Hamel & Côté 2008).
In the alpine marmot (Marmota marmota), a hibernating social rodent, time constraints are intense since individuals have to accumulate enough fat reserves during the short vegetative season typical of the alpine environment (Billings 1974; around 4 months at our site) to survive hibernation. At the same time, due to their complex social system, alpine marmots need to invest time and energy in social interactions (Panaccio et al. 2021). An exhaustive study on activity budget allocation is still lacking for alpine marmots, contrary to other related marmot species such as the yellow-bellied marmot (Marmota flaviventris; Armitage & Salsbury 2016).
Here, we explored the effects of extrinsic (temperature, time, period of the season, weather, site) and intrinsic (sex, age, and dominance status) factors on the daytime, above-ground, activity budget of alpine marmots. Furthermore, we tested the following predictions: (i) Considering that the alpine marmot is a fat-storing hibernating species, we expected foraging activity to be the most common behaviour during the active season (in agreement with results by Hannon et al. 2006). (ii) As a species adapted to low temperatures (Armitage 2017), we expected time of the day, weather conditions, and period of the season to affect animal activity in agreement with results on yellow-bellied marmots (Armitage & Salsbury 2016); specifically, we expect marmots to forage more in the second part of the season (July–August) compared to the first (June) and the third (August–September). Overall, we expected marmots to be scarcely active (foraging, moving) at high temperatures, at mid-day, and on sunny days. (iii) Based on a previous study highlighting different levels of vigilance in different areas (Ferrari et al. 2009), we expected vigilance behaviour to be affected by environmental characteristics of the site (e.g. visibility). (iv) As a social and territorial species, we expected agonistic and affiliative interactions to represent a substantial proportion of the activity budget, but to vary seasonally. We expected both inter- and intra-group agonistic interactions to be predominant early in the season, when animals delimit family territories and establish hierarchies, while intra-group affiliative interactions, which may have a joint function across all the family members, were expected to be stable across the whole season (Pasquaretta et al. 2015). (v) As alpine marmots are a cooperative breeding species in which mainly dominant individuals have access to reproduction and juveniles and subordinates remain in the family group as helpers (Pasquaretta et al. 2015; Berget et al. 2018), we expected that socially related activities (affiliative and agonistic interactions) should vary according to individual characteristics such as sex, age, and dominance status (Blumstein & Armitage 1999). We expected dominant male individuals to have more agonistic interactions compared to other sex-age classes in order to maintain their status, while we expected pups and yearlings to be more frequently engaged in affiliative behaviours.
To test our predictions, we collected alpine marmot activity budget data at two study areas located at different elevations, with different habitat characteristics. We used a Bayesian hierarchical multinomial regression model approach to analyse the data (Koster & McElreath 2017). This study provides the first detailed description of the activity budget of alpine marmots that can be used as a reference tool to further investigate variation in behavioural activities in this species.
Materials and methods
Study species
The alpine marmot (Marmota marmota) is a diurnal herbivorous species living in open habitats at elevations ranging from 1500 to 3000 m a.s.l., where animals must cope with a short vegetative season and extremely variable weather conditions (Macchi et al. 1992; Allainé et al. 1994; Garin et al. 2008; Zanet et al. 2017). Alpine marmots hibernate during the winter (in the Alps from October to April), and they mate after emerging from their winter burrows in spring. Gestation is approximately 33–35 days, and following birth, pups are nursed for approximately 40 days (Hacklaender & Arnold 1999). This species is highly social: individuals live in family territorial groups ranging from 2 to 20 individuals with the dominant pair, subordinate adult individuals (older than 2 years old), yearlings, and pups born in the summer (Arnold & Dittami 1997). Within the family, dominant individuals use direct agonistic interactions to establish and maintain the hierarchy and, consequently, access to reproduction. Affiliative interactions are used by all members to increase cohesion in the group (Arnold & Dittami 1997; Hackländer et al. 2003).
Study area
The data analysed in this study were collected during summer 2008, from April to September, in the Orvieilles alpine marmot study area in Valsavarenche (Gran Paradiso National Park, North-western Italian Alps, 45° 34′ N, 7° 11′ E). Captures and behavioural data collection described below were done at two sites: the first (hereafter referred to as the low site) was located at 2100 m a.s.l. This site was characterized by proximity to a larch (Larix decidua) forest, several rocks, and isolated big boulders, resulting in discontinuous visibility of approaching terrestrial predators for most of this site. Moreover, by having 3 different hiking trails pass through it, this area is an easy and common destination for hikers. Groups of hikers who stop to eat or rest may disturb and interrupt marmot activity (Mainini et al. 1993; CF, personal observation). The second site (hereafter referred to as the high site) was located at 2300 m a.s.l. and was far from forest edges. This area was characterized by the absence of rocks and big boulders, as well as minimal shrub cover resulting in optimal visibility for alpine marmots against terrestrial predators. In both areas, the main predators of alpine marmots are red foxes (Vulpes vulpes) and golden eagles (Aquila chrysaetos). This area has only one hiking trail crossing the area, and it is not considered a hiking destination. As a result, hikers usually continue walking along the trail to reach other destinations, representing therefore a minor disturbance for marmots.
Capture and individual marking
The beginning of the active season (late April to end of May) was mainly devoted to trapping and marking individuals within the framework of a long-term alpine marmot research project running since 2006 in this area (Ferrari et al. 2013). Captures were done using Tomahawk traps (150 × 30 × 30 cm, Tomahawk live traps, Hazelhurst, WI, USA) located near the entrances of marmot burrows and baited with either lettuce, Alpine dandelion (Taraxacum alpinum), or salt. Traps were activated at 6:00, checked constantly from a distance using spotting scopes, and closed at 20:00. After a marmot was captured, the trap was covered with a gunnysack and the marmot was transferred into another gunnysack that had a tear-off opening to facilitate ear-marking. Marmots were not anesthetized but they were kept within the gunnysack during the whole time to avoid excessive stress, while we followed a standard protocol of operations including marking, sexing, and weighing (for details, see Costantini et al. 2012; Ferrari et al. 2013). Each individual was marked with a unique combination of coloured plastic ear tags (Minirotag, 5-cm length, Ghislandi and Ghislandi, Bergamo, Italy) on its two ears for visual identification. Pups were marked using fur bleach (Moldus, Italy) since their ears were too small and frail to be tagged. Upon first capture, each individual was sexed and classified as a pup, yearling, or adult. All individuals were also permanently tagged with an under-skin pit tag (Bayer Animal Coder, Bayer S.p.A., Milan, Italy) for long-term identification. In this study, we analysed the behaviours of 48 marked individuals belonging to 11 families in summer 2008 (see Table 1 for details).
Data collection
We collected behavioural data using scan sampling (Martin & Bateson 1993) from 12 Jun. until 19 Sep. 2008. With this method, a whole group of individuals was scanned at regular intervals, and observers noted the activity of each marmot exactly at the instant it was observed (Martin & Bateson 1993). By being diurnal and territorial, and thus easily observable when above ground, scan sampling is a suitable method to collect behavioural data on this species. Scan sampling was conducted 5 days per week alternating among the different family groups by using long-view scopes (Swarovski 30x75 and Nikon ED82 25-56X82) following the method described in Pasquaretta et al. (2012, 2015). Scan sampling sessions, lasting 1 h each, were conducted for the whole day from 07:00 to 20:00 (Central European Summer time), with a break between 13:00 and 14:00 for the entire time of this study. We attempted to have an equal number of scan sampling sessions for each family we monitored. The sessions for each family were uniformly distributed during the day over the entire season. To avoid temporal autocorrelation among observations, each family was observed only once per day. Two observers recorded the behaviours performed by each visible individual every 5 min, obtaining a maximum of 12 behaviours per individual per hour. Behaviours were categorized according to the ethogram described in Table 2. Observations were carried out from fixed locations at a variable distance from each family (range 100–400 m), chosen for the best visibility of the entire territory of each focal family group. The marmots we did not see during a scan sampling session were not necessarily underground, as it is possible that the observer simply did not detect them while scanning the territory (e.g. individual hidden by rocks). We were therefore unable to distinguish between time in burrow and time above ground but out of sight. Young of the year were excluded from the analysis.
Similar to a previous study on yellow-bellied marmots (Armitage & Salsbury 2016), the active season was divided into three periods according to marmot biology (Table 3). Weather conditions were always noted by the same observer during each scan and were classified based on the presence of sun and clouds into three classes (Table 3). Observations during hours of intense rain were not conducted to ensure the safety of observers and because the visibility was very low. Furthermore, in cases of intense rain, marmots generally are not visible above ground (CF, pers. obs.). Observations were resumed when weather conditions allowed the observation of active animals. Temperature data (one value per hour) were obtained from the Aosta Valley regional weather station located at the low site. While we did not have a direct measure of solar radiation, the hour of the day, weather, and temperature have been shown to affect radiation on the surface in the alpine environment (Rangwala & Miller 2012; Rottler et al. 2019).
We assigned the dominance status of members of each family based on observations of social interactions (Panaccio et al., 2021). Dominant individuals had the highest frequency of agonistic interactions with a positive outcome. In most cases, individuals with the highest dominance rank based on social interactions were also the reproductive pair. In family groups where reproduction did not take place, we used only the social rank to determine the dominance status.
Statistical analyses
Behavioural data were analysed with a Bayesian hierarchical multinomial model (Koster & McElreath 2017). This approach is relatively new in the field of behavioural analysis that considers the dependent variable as a multinomial character (i.e. it considers the relative frequency of each behavioural category at the same time, instead of analysing each behaviour singularly as a dependent variable in separate models), with the possibility of accounting for repeated observations on individuals (Koster & McElreath 2017).
Following the procedure described in Koster and McElreath (2017), we ran the multinomial model with all the behaviours composing the ethogram (Table 2) as a response variable. We ran four different models to test for the structure and the relative importance of random effect terms:
-
Mod1: a model with ID (individual) as a random effect term and no fixed effects
-
Mod2: a model with ID nested in Family as random effect terms and no fixed effects
-
Mod3: a model with ID as a random effect term and all the variables described in Table 3 as fixed effects
-
Mod4: a model with ID nested in Family as random effect terms and all the variables described in Table 3 as fixed effects
We included a random effect for Family to reflect the fact that members of families are mostly related to each other and share the same environmental conditions. All model specifications included 3 MCMC chains of 2000 iterations each, of which 1000 were for the warm-up. Model convergence was visually assessed for each model. We then compared the four models using the widely applicable information criterion (WAIC), an extension of the Akaike information criterion (AIC) commonly used in Bayesian analysis (Watanabe 2010). This criterion estimates pointwise out-of-sample prediction accuracy; i.e. it uses the computed log pointwise posterior predictive density (LPPD) and it corrects for the number of parameters used in the models to adjust for overfitting (Vehtari et al. 2017).
To assess behavioural variation in relation to continuous variables (i.e. to establish if a continuous variable affected the probability of observing a certain behaviour), we determined if the 95% credible intervals (CI) around the coefficients estimates included zero. Time of the day was modelled with a quadratic function. To assess differences among categorical effects, we used the distribution of each of the contrasts considered. Specifically, we computed the 95% credibility intervals (95%CI) by using the 95th percentile of samples from the posterior density. We considered that the fixed factor had an effect when the percentile intervals of the contrasts did not include zero or had zero at an extreme. We estimated the conditional probability of engaging in the different activities based on all possible combinations of the different possible values of the categorical predictors. All estimated coefficients presented in the main text are for models of the logarithm of the odds of each response (logit scale). All predictions presented in the figures have been back-transformed to the probability scale.
We used R version 3.3.2 (R Core Team, 2016) with the packages RStan (Stan Development Team, 2016), rethinking (McElreath 2016), and chron (James & Hornik 2017) for all analyses. We wrote the Bayesian hierarchical multinomial models in STAN language (Carpenter et al. 2017), adapting the code provided by Koster and McElreath (2017). Data and code used in this manuscript are available at https://data.mendeley.com/datasets/66gg4mzn22/draft?a=d9a1053e-8b07-48f8-9d35-a65757a0e278.
Results
We analysed 3515 behavioural observations collected during 146-h-long scan sampling sessions on 48 marked individuals of different ages and sexes belonging to 12 different marmot families (Table 1). Foraging was the most represented activity, accounting for 54% of the behavioural observations (Table 2). Sitting in front of the burrow and vigilance were almost equal in their percentage (15% and 14.7% respectively). Moving/exploring accounted for 4.6% of behavioural observations, affiliative behaviours were 8.6%, and agonistic behaviours were 2%, while other behaviours accounted for only about 1% of observations.
The WAIC model selection indicated that the mixed effect model with ID nested in Family as random effect terms, in addition to all the variables described in Table 3 as fixed effects (Mod4), had a better fit than the other 3 models (Table 4), and we therefore selected this model for all further analyses. For all results below, we report the conditional probabilities for our reference condition (i.e. adult females during the first period (June), with cloudy weather, in the low site, at 08:00) and indicate in the text if different combinations of the predictors showed different trends. All estimates of the model are provided for the reference model (ESM 1).
Temperature
Our results did not show an effect temperature on the probability of any of the behaviours, for which all of the credible intervals included zero. All coefficients and the relative credible intervals are provided in the Electronic Supplemental Material (ESM_1).
Time of the day
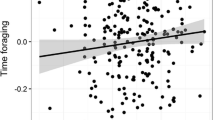
The probability of all behaviours varied according to the time of the day (Fig. 1). Affiliative behaviours had a peak between 07:00 and 09:00, after which they started to decrease slowly across the day, reaching their lowest probability of occurrence in the evening (β = 0.15, 95%CI = −0.75,0.39; β2 = 0.87, 95%CI = 0.27,1.46, where β is the time and β2 is the quadratic term of time). Foraging increased constantly from the morning to the late afternoon (17:00) when it started to decrease (β = 0.27, 95%CI = −0.29,0.81; β2 = 0.86, 95%CI = 0.25,1.44). Sitting in front of the burrow had its highest probability between 07:00 and 10:00, and then it was nearly zero until 16:00 when it started to increase again until the evening (β = −0.26, 95%CI = −0.83,0.29; β2 = 1.43, 95%CI = 0.83,2.03). Vigilance was highest from 07:00 to 11:00, while moving/exploring peaked in the mid-hours of the day (β = −0.09, 95%CI = −0.65,0.48; β2 = 1.13, 95%CI = 0.52,1.72).

Model prediction of behaviours as a function of time of the day. The shaded regions are the 90% percentiles as calculated from the posteriors of model Mod4 (Table 4)
Period of the season
The period of the season was related to the probability of foraging, sitting in front of the burrow, vigilance, and moving/exploring, as well as affiliative behaviours, and agonistic interactions (Figure 2; Table 5a). Foraging and sitting in front of the burrow had a higher probability in the first and third periods compared to the second, while vigilance had a higher probability in July, the second period, compared to the other two periods. Moving/exploring had a higher probability during the first period compared to the second and third periods, as well as a higher probability during the second compared to the third. The probability of affiliative interactions was just barely higher in the first period compared to the second and was lower in the third period compared to both previous ones. The probability of agonistic interactions did not differ between the first and second periods; however, agonistic interactions were more common in the second period compared to the third.

Model prediction of behaviours as a function of the period of the season. As period is not a continuous variable, we could not represent a mean value of the variable. Bars represent the credible intervals calculated at 95% of the posterior samples of the model
Weather conditions
Weather conditions were related to the probability of foraging, sitting in front of the burrow, and affiliative behaviours (Table 5b). Alpine marmots had a higher probability of being observed foraging on cloudy days compared to variable and sunny days. Sitting in front of the burrow had a lower probability on cloudy days compared to both sunny and variable days. Affiliative behaviours had a lower probability on cloudy and variable days compared to sunny days, but these behaviours were similar on cloudy and variable days. Vigilance, agonistic, and moving/exploring behaviours were related to weather conditions.
Site
The probability of foraging, sitting in front of the burrow, moving/exploring, and affiliative behaviour did not differ between the two sites. There was also no difference in agonistic behaviour between the two sites. Vigilance behaviour was slightly higher in individuals inhabiting the low site compared to those living in the high site (Table 5c).
Sex
The sex of an individual was related to the probability of moving/exploring, agonistic, and affiliative behaviours (Table 5d). The probability of moving/exploring was barely higher in females compared to males, as well as the probability of engaging in affiliative interactions, which was higher in females than males. On the contrary, agonistic behaviour had a slightly higher probability to be seen in males compared to females.
Age and dominance status of individuals
Age and dominance status were related to the probability of moving/exploring as well as affiliative and agonistic behaviours (Table 5e). Affiliative behaviours varied among all age classes and dominance statuses. Dominant adult males had a slightly higher probability of affiliative interactions compared to subordinate males; this difference was not found among females. Among males, a lower probability of engaging in affiliative interactions was found for dominant compared to yearlings; this result was not found among females. Yearlings also showed a higher probability to engage in affiliative interactions compared to subordinate adults, which therefore represent the category with a lower probability of being engaged in affiliative interactions. Compared to yearlings, dominant individuals showed a higher probability of moving/exploring, while their agonistic behaviour was just slightly higher. All other combinations between predictors had the same differences among age and status categories.
Discussion
Results of this study indicate that extrinsic factors were related to the probabilities of engaging in all behavioural activities of alpine marmots, while intrinsic factors mostly regulated social behavioural activities. As predicted, and in agreement with other studies investigating activity budgets in the genus Marmota (Holmes 1984; Armitage et al. 1996; Perrin & Coulon 1993; Hannon et al. 2006; but see Armitage & Salsbury 2016), our results indicated that foraging was the most common activity in alpine marmots and that cloudiness is a key predictor of marmot activity. In the yellow-bellied marmot, the activity of individuals outside of burrows was positively affected by cloud cover. However, contrary to our results on alpine marmots, the activity of yellow-bellied marmots was negatively related to solar radiation and temperature (Armitage & Salsbury 2016). Resting in a static position in front of the burrow entrance (an activity which may include basking; Armitage & Chiesura-Corona 1994) and vigilance were the two behaviours with the highest probabilities after foraging.
In our study, temperature was the only covariate that did not affect the activity budget of alpine marmots. This was unexpected given previous literature on other marmot species and considering that other factors indirectly linked to temperature and solar radiation (Rangwala & Miller 2012; Rottler et al. 2019), such as the time of the day and the period of the season, affect the activity budget of alpine marmots. However, the limit given by temperature to a marmot activity is better described by the standard operative temperature of an individual, an index which integrates temperature of the environment, wind, and solar radiation (Armitage 2017) and has some flexibility in its extremes. We thus suggest that all these values, if possible, and not temperature alone should be considered in future studies of activity budget in marmot.
Foraging activity was high in the first period (June) and decreased in the second (July–August), the period characterized by higher vegetation productivity (Ranghetti et al. 2016). This suggests that the availability of high-quality food resources during July reduces the time necessary to fulfil the energetic demands of the alpine marmots, similar to findings on mouflon (Ovis gmelini musimon × Ovis sp.; Bourgoin et al. 2008). Previously, we showed that vegetation starts to grow around the end of May–early June at our study site (Ranghetti et al. 2016). We therefore suggest that, in the first period, marmots maximize their foraging effort to regain energy after hibernation, and during the second period, marmots can devote more time to other activities, such as social interactions. Similarly, in the third period just prior to winter hibernation, the time devoted to foraging increases, probably due to a lower quality of vegetation and the need to increase fat reserves, with a concomitant decrease in social interactions.
Foraging increased during the day, suggesting that this activity may be more favourable when visibility is good and the temperature is not too low as it is at high altitudes in the early morning and late afternoon, when the sun is below the horizon. Foraging activity is energetically demanding due to the costs associated with food searching and processing, and it can be less favourable on cloudless days, when animals are exposed to direct solar radiation. Accordingly, our results suggest that, on sunny days, alpine marmots spend more time sitting in front of their burrow, while they are more likely to forage when clouds cover the sun, resulting in lower solar radiation (Pyrgou et al. 2019; Rottler et al. 2019). Indeed, solar radiation is the variable influencing the most heat transfer through a marmot’s fur: at mid-day, with direct solar radiation, the basal heat production of a marmot can increase by 10 times, generating a major heat stressor in marmots (Armitage 2009; Melcher et al. 1990). But alpine marmots do not appear to have any mechanisms to disperse excess heat besides burrowing (Webb 1979; Melcher et al. 1990), laying under big stones, orienting their bodies such that they limit direct exposure to the sun during the hours of intense radiation (Armitage 2009), and adjusting activity above ground (Armitage 2009), as for example foraging more on cloudy than cloudless days.
Furthermore, we found that sitting in front of their burrows is more frequent during the early morning and late afternoon in alpine marmots. On the contrary, in yellow-bellied marmots, Armitage and Salsbury (2016) collected most observations of sitting and resting behaviour during mid-day. In the alpine habitat inhabited by alpine marmots, characterized by few, if any, shaded areas in the open alpine prairies, it is possible that marmots prefer to burrow under the ground instead of remaining sitting in front of their burrows for long periods during the hours of highest solar radiation. This behaviour has been already documented in Cape ground squirrels Xerus inauris, where individuals tend to go underground during hours of intense heat to lower their body temperature (Scantlebury et al. 2012).
Vigilance was more frequently observed at the low-elevation site, characterized by discontinuous visibility, compared to the high-elevation site, confirming results documented in the same species (Ferrari et al. 2009). Similarly, yellow-bellied marmots were less vigilant in areas of low vegetation compared to those with high vegetation (Chmura et al. 2016): for a prey species like marmot, visual obstacles represented by big rock or high vegetation may decrease the possibility of spotting a predator during foraging activity, resulting in the necessity of increasing vigilance rates. Considering the different levels of human disturbance at the two sites (i.e. higher at the low site; CF, pers. obs.), we cannot exclude human disturbance, and not only natural predators favour the observed higher vigilance rate at the low site, as has been shown in other wild mammals (Pęska & Ciach 2018).
We predicted differences in vigilance at the two sites, but we also found that vigilance behaviour varied in response to most of the extrinsic covariates included in our model. Specifically, we found that marmots were more likely to be vigilant in the early morning and late afternoon. When the sun is rising, or is going down, visibility may be reduced, which may increase the risk of predation for alpine marmots (Semenov et al. 2000), especially from red foxes, an important predator of marmots at our study area (Cagnacci et al. 2003). Vigilance activity changed also over the season, reaching its peak during the second period, when pups emerge from their burrows, to forage, and play with other members of their group on their families’ territory. We have observed that red foxes visit areas inhabited by alpine marmots more frequently as pups emerge from their burrows (CF, unpublished data), and therefore, this period likely requires more anti-predatory vigilance as demonstrated in meerkats (Suricata suricatta; Santema & Clutton-Brock 2013). We thus suggest that the higher vigilance observed in the second period may be related to a higher predation risk for the pups.
Moving/exploring was more frequently observed in the second period compared to the rest of the season. During the first period, marmots may limit exploratory activity due to the time they need to forage to recover after the winter period. The observed decrease of moving/exploring in the third and last period is related to a phase of pre-hibernation for the species when the overall activity of marmots decreases (Webb 1979; Perrin & Coulon 1993; Kŏrtner & Heldmaier 1995). Accordingly, affiliative behaviour was at its highest level in period A, supporting the results of Perrin et al. (1993), who suggested that early in the season, post-emergence from hibernation is important for establishing a stable hierarchy within family groups.
As expected, dominant individuals had a higher probability of agonistic and moving/exploring behaviour to defend the family territory, regularly moving all around to mark the boundaries and to eventually interact with intruders exhibiting agonistic display or direct attack (Lenti Boero 1995). In a previous study on the same population, Pasquaretta et al. (2015) found that dominant males are also more often involved in agonistic interactions with both members of the family and intruders, compared with agonistic interactions carried out by other members of the family group. These interactions, on which we based our dominance status ranking (Panaccio et al. 2021), are observed with a higher frequency in the second period, coinciding with the emergence of the pups. Wandering individuals attempting to take over the territory represent a risk for dominant individuals and their offspring, which could be killed by the intruder (Hackländer & Arnold 1999). Our experience in the field indicates that early May is the period with the highest frequency of territorial quarrels, but we had no appropriate data to include this period in our analysis. We are also aware that food resources are still scarce in May at the elevations we studied marmots, while in June, vegetation starts to grow. We suggest that in June, marmots spend most of their time foraging to recover body mass and energy, while, during July, marmots devote more time to agonistic behaviours because there is greater food availability (Ranghetti et al. 2016) and the young of the year need to be protected.
Yearlings were more likely to have affiliative interactions compared to dominant and subordinate adults, a result which has been found in the yellow-bellied marmot (Armitage 2012) such as in other social mammals (Clutton-Brock & Sheldon 2010). At this age, individuals are not reproductively mature yet, which likely favours a higher frequency of affiliative interactions with individuals of all age classes within the family group. Among adults, dominant ones have a higher probability of engaging in affiliative interactions compared to subordinates, and females more than males. Dominant individuals often perform and receive this type of interaction, which serves to maintain the hierarchy and reinforce the cohesion of the group, with a higher number of individuals compared to subordinates (Clutton-Brock & Huchard 2013).
Social interactions were little represented in our data; in particular, agonistic interactions represented only 2% of the observations. This could be partially due to the fact that agonistic interactions are generally rare in marmots and therefore may be underrepresented in the activity budget estimated with scan sampling. We suggest that, in future studies focused on social interactions, observers use an “all occurrence” sampling method for categories of behaviours scarcely observable but of high relevance for the species such as agonistic interactions (Martin & Bateson, 1993; Blumstein et al. 2018).
Conclusion
In the present study, we detailed the above-ground, daytime activity budget of alpine marmots in relation to extrinsic and intrinsic factors using a Bayesian hierarchical multinomial regression approach (Koster & McElreath 2017). Despite being a well-studied species, a complete description of the activity budget of alpine marmots was lacking, and this study fulfils this gap in the literature and provides a useful tool for future studies on this species.
Our results indicate that all the extrinsic variables related to climatic and environmental conditions, such as weather, site, seasonal period, and time of day but with the exception of temperature, are associated with all the behaviours included in the activity budget of this species. Thus, external conditions, which are generally the same for all marmots, influence all individuals no matter their age, sex, and dominance status. We suggest that temperature of the environment alone may fail in detecting variation in activity during the day, while solar radiation may be a clearer driver of marmot behaviour to be considered in future studies. Intrinsic factors, such as age, sex, and dominance status are associated with activities related to the social dynamics of alpine marmots, such as affiliative, agonistic, and moving/exploring behaviours.
In the future, we suggest collecting data on individual activity budget allocation on recognizable individuals during multiple consecutive years and considering individual’s phenotypic plasticity (sensu Nussey et al. 2007) in response to inter-annual environmental variation. In species influenced by climatic and environmental habitat changes, the plasticity of an organism may favour adaptation to the new environmental conditions (Charmantier et al. 2008; Maldonado et al. 2017): the alpine marmot, a species scarcely adapted to high temperature and dry environment (Armitage 2013), will be a pivotal subject to study the possibility, if any, for alpine species to cope with a changing environment.
Availability of data, material, and code
The data and code used in this manuscript are available at https://data.mendeley.com/datasets/66gg4mzn22/draft?a=d9a1053e-8b07-48f8-9d35-a65757a0e278.
Change history
18 July 2022
Missing Open Access funding information has been added in the Funding Note.
References
Allainé D, Rodrigue I, Le Berre M, Ramousse R (1994) Habitat preferences of alpine marmots, Marmota marmota. Can J Zool 72:2193–2198
Allainé D (2000) Sociality, mating system and reproductive skew in marmots: evidence and hypotheses. Behav processes 51(1–3):21–34
Allainé D, Theuriau F (2004) Is there an optimal number of helpers in Alpine marmot family groups? Behav Ecol 15:916–924
Aniano L, Ungerfeld R (2016) Time budget of socially high and low ranked pampas deer (Ozotoceros bezoarticus) females. North-Western J of Zool 12:58–64
Armitage KB, Chiesura-Corona M (1994) Time and wariness in yellow-bellied marmots. IBEX J Mountain Ecol 2:1–8
Armitage KB, Salsbury CM, Barthelmess EL, Gray RC, Kovach A (1996) Population time budget for the yellow-bellied marmot. Ethol Ecol Evol 8:67–95
Armitage KB (2009) Fur color diversity in marmots. Ethol Ecol Evol 21:183–194
Armitage KB (2012) Sociality, individual fitness and population dynamics of yellow-bellied marmots. Mol Ecol 21:532–540
Armitage KB (2013) Climate change and the conservation of marmots. Natural Science 5:36–43
Armitage KB, Salsbury CM (2016) Pattern and variation of the time budget of yellow-bellied marmots. Ethol Ecol Evol 28(3):329–365
Armitage KB (2017) Hibernation as a major determinant of life-history traits in marmots. J Mammalogy 98(2)
Arnold W (1989) The evolution of marmot sociality: II. Costs and benefit of joint hibernation. Behav Ecol Sociobiol 27:239–246
Arnold W, Ruf T, Kuntz R (2006) Seasonal adjustment of energy budget in a large wild mammal, the Przewalski horse (Equus ferus przewalskii) II. Energy expenditure. J Ex Biol 209:4566–4573
Beauchamp G (2014) Social predation: how group living benefits predators and prey. Academic Press
Belovsky GE, Slade JB (1986) Time budgets of grassland herbivores: body size similarities. Oecol 70:53–62
Berger V, Lemaître JF, Allainé D, Gaillard JM, Cohas A (2018) Early and adult social environments shape sex-specific actuarial senescence patterns in a cooperative breeder. Am Natur 192:525–536
Billings WD (1974) Adaptations and origins of alpine plants. Arctic and Alpine Research 6(2):129–142
Blumstein D, Armitage K (1999) Cooperative breeding in marmots. Oikos 84:369–382
Blumstein DT, Williams DM, Lim AN, Kroeger S (1871) Martin JGA (2018) Strong social relationships are associated with decreased longevity in a facultatively social mammal. Proc R Soc B 285
Bourgoin G, Garel M, Blanchard P, Dubray D, Maillard D, Gaillard JM (2008) Determinants of seasonal variation in activity patterns of mouflon. Can J Zool 86:1410–1418
Cagnacci F, Lovari S, Meriggi A (2003) Carrion dependence and food habits of the red fox in an Alpine area. Ita J Zool 70:31–38
Carey HV (1985) The use of foraging areas by yellow-bellied marmots. Oikos 44:273–279
Carpenter B, Gelman A, Hoffman MD, Lee D, Goodrich B, Betancourt M, Brubaker M, Guo J, Li P, Riddell A (2017) Stan: a probabilistic programming language. J Statist Soft 76(1)
Charmantier A, McCleery RH, Cole RL, Perrins C, Kruuk LEB, Sheldon BC (2008) Adaptive phenotypic plasticity in response to climate change in a wild bird population. Science 9:800–803
Chmura HE, Wey TW, Blumstein DT (2016) Assessing the sensitivity of foraging and vigilance to internal state and environmental variables in yellow-bellied marmots (Marmota flaviventris). Behav Ecol Sociobiol 70(11):1901–1910
Cohas A, Yoccoz NG, Allaine D (2007) Extra-pair paternity in alpine marmots, Marmota marmota: genetic quality and genetic diversity effects. Behav Ecol Sociobiol 61:1081–1092
Costantini D, Ferrari C, Pasquaretta C, Cavallone E, Carere C, von Hardenberg A, Réale D (2012) Interplay between plasma oxidative status, cortisol and coping styles in wild alpine marmots (Marmota marmota). J Ex Biol 215:374
Clutton-Brock T, Sheldon BC (2010) Individuals and populations: the role of long-term, individual-based studies of animals in ecology and evolutionary biology. Trends Ecol Evol 25:562–573
Clutton-Brock T, Huchard E (2013) Social competition and selection in males and females. Philos T R Soc B 368:1631
Ebensperger LA, Hurtado MJ (2005) Seasonal changes in the time budget of degus. Behav 142:91–112
Ferrari C, Bogliani G, von Hardenberg A (2009) Alpine marmots (Marmota marmota) adjust vigilance behaviour according to environmental characteristics of their surroundings. Ethol Ecol Evol 21:355–364
Ferrari C, Pasquaretta C, Carere C, Cavallone E, von Hardenberg A, Réale D (2013) Testing for the presence of coping styles in a wild mammal. Anim Behav 85:1385–1396
Ferretti F, Costa A, Corazza M, Pietrocini V, Cesaretti G, Lovari S (2014) Males are faster foragers than females: intersexual differences of foraging behaviour in the Apennine chamois. Behav Ecol Sociobiol 68:1335–1344
Garin I, Aldezabal A, Herrero J, García-Serrano A, Remón LJ (2008) Diet selection of the alpine marmot (Marmota M. marmota L.) in the Pyrenees. Revue d’ Ecologie 63:383–390
Grant EC (1963) An analysis of the social behaviour of the male laboratory rat. Behav 21:260–281
Hackländer K, Arnold W (1999) Male-caused failure of female reproduction and its adaptive value in alpine marmots (Marmota marmota). Behav Ecol 10:592–597
Hackländer K, Möstl E, Arnold W (2003) Reproductive suppression in female Alpine marmots. Marmota marmota. Animal Behav 65(6):1133–1140
Hamel S, Côté SD (2008) Trade-offs in activity budget in an alpine ungulate: contrasting lactating and nonlactating females. Anim Behav 75:217–227
Hannon MJ, Jenkins SH, Crabtree RL, Swanson AK (2006) Visibility and vigilance: behavior and population ecology of Uinta ground squirrels (Spermophilus armatus) in different habitats. J Mammal 87:287–295
Holmes WG (1984) Predation risk and foraging behaviour of the hoary marmots in Alaska. Behav Ecol Sociobiol 15:293–301
James D, Hornik K (2017) chron: Chronological Objects which Can Handle Dates and Times. R package version 2:3–51
Kŏrtner G, Heldmaier G (1995) Body weight cycles and energy balance in the Alpine marmot (Marmota marmota). Physiol Zool 68:149–163
Koster J, McElreath R (2017) Multinomial analysis of behavior: statistical methods. Behav Ecol Sociobiol 71:138
Laszlo N, Grabherr G (2009) The biology of alpine habitats. Oxford University Press
Lenti Boero D (1995) Scent deposition behavior in Alpine marmots (Marmota marmota L.): its role in territorial defence and social communication. Ethol 100:26–38
Macchi E, Bassano B, Durio P, Tarantola M, Vita A (1992) Ecological parameters affecting the settlement’s choice in Alpine marmot (Marmota marmota). Proceedings of the First International Symposium on Alpine Marmot and the genus Marmota. Bassano B, Durio P, Gallo Orsi U, Macchi E, editors. 123-127
Mainini B, Neuhaus P, Ingold P (1993) Behavior of marmots Marmota marmota under the influence of different hiking activities. Biological Conserv 64(2):161–164
Maldonado-Chaparro AA, Read DW, Blumstein DT (2017) Can individual variation in phenotypic plasticity enhance population viability? Ecological Modelling 352:19–30
Martin P, Bateson PPG (1993) Measuring behaviour: an introductory guide (2nd ed.). Cambridge University Press
Mason THE, Brivio F, Stephens PA, Apollonio M, Grignolio S (2017) The behavioral trade-off between thermoregulation and foraging in a heat-sensitive species. Behav Ecol 28:908–918
McElreath R (2016) rethinking: Statistical Rethinking book package. R package version 1:59
Melcher J, Armitage K, Porter W (1990) Thermal influences on the activity and energetics of yellow-bellied marmots (Marmota flaviventris). Physiological Zool 63:803–820
Nussey DH, Wilson AJ, Brommer JE (2007) The evolutionary ecology of individual phenotypic plasticity in wild populations. J Evol Biol 20:831–844
Oakes EJ, Harmsen R, Eberl C (1992) Sex, age and seasonal differences in the diets and activity budgets of muskoxen (Ovibos moschatus). Can J Zool 70:605–616
Pasquaretta C, Bogliani G, Ranghetti L, Ferrari C, von Hardenberg A (2012) The Animal Locator: a new method for accurate and fast collection of animal locations for visible species. Wildlife Biol 18:202–214
Pasquaretta C, Busia L, Ferrari C, Bogliani G, Réale D, von Hardenberg A (2015) Helpers influence on territory use and maintenance in Alpine marmot groups. Behav 152:1391–1412
Perrin C, Coulon MLB, Le Berre M (1993) Social behavior of alpine marmots (Marmota marmota): seasonal, group, and individual variability. Can J Zool 71(10):1945–1953
Pęska L, Ciach M (2018) Daytime activity budget of an alpine ungulate (Tatra chamois Rupicapra rupicapra tatrica): influence of herd size, sex, weather and human disturbance. Mammal Research 63:443–453
Pyrgou A, Santamouris M, Livada I (2019) Spatiotemporal analysis of diurnal temperature range: effect of urbanization, cloud cover, solar radiation and precipitation. Climate 7(7):89
R Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria https://www.R-project.org/
Ranghetti L, Bassano B, Bogliani G, Palmonari A, Formigoni A, Stendardi L, von Hardenberg A (2016) MODIS time series contribution for the estimation of nutritional properties of alpine grassland. European J Remote Sensing 49:691–718
Rangwala I, Miller JR (2012) Climate change in mountains: a review of elevation-dependent warming and its possible causes. Climatic Change 114(3–4):527–547
Rogora M, Frate L, Carranza M, Freppaz M, Stanisci A, Bertani I, Bottarin R, Brambilla A, Canullo R, Carbognani M, Cerrato C, Chelli S, Cremonese E, Cutini M, Di Musciano M, Erschbamer B, Godone D, Iocchi M, Isabellon M et al (2018) Assessment of climate change effects on mountain ecosystems through a cross-site analysis in the Alps and Apennines. Science of The Total Environment 624:1429–1442
Rottler E, Kormann C, Francke T, Bronstert A (2019) Elevation-dependent warming in the Swiss Alps 1981–2017: features, forcings and feedbacks. International J Climatology 39(5):2556–2525
Ruckstuhl KE (1998) Foraging behaviour and sexual segregation in bighorn sheep. Anim Behav 56(1):99–106
Ruckstuhl KE (1999) To synchronise or not to synchronise: a dilemma for young bighorn males? Behav 136(6):805–818
Ruckstuhl KE, Neuhaus P (2002) Sexual segregation in ungulates: a comparative test of three hypotheses. Biological Reviews 77:77–96
Santema P, Clutton-Brock T (2013) Meerkat helpers increase sentinel behavior and bipedal vigilance in the presence of pups. Anim Behav 85(3):655–661
Scantlebury M, Danek-Gontard M, Bateman PW, Bennett NC, Manjerovic MB, Joubert KE, Waterman JM (2012) Seasonal patterns of body temperature daily rhythms in group-living Cape ground squirrels Xerus inauris. PLOS ONE 7(4)
Semenov Y, Ramousse R, Le Berre M (2000) Effet de la lumière et de la température sur le rythme d’activité de la marmotte alpine (Marmota marmota Linné, 1758) en milieu naturel. Can J Zool 78:1980–1986
Stan Development Team. 2016. RStan: the R interface to Stan, version 2.10.1, http://mc-stan.org/
Vehatari A, Gelman A, Gabry J (2017) Practical Bayesian model evaluation using leave-one-out cross-validation and WAIC. Stat Comput 27:1413–1432
Watanabe S (2010) Asymptotic equivalence of Bayes cross validation and widely applicable information criterion in singular learning theory. J Machine Learning Research 11:3571–3594
Webb DR (1979) Environmental harshness, heat stress and Marmota flaviventris. Oecol 44:390–395
Zanet S, Miglio G, Ferrari C, Bassano B, Ferroglio E, von Hardenberg A (2017) Higher risk of gastrointestinal parasite infection at lower elevation suggests possible constraints in the distributional niche of Alpine marmots. PLOS ONE 12(8)
Acknowledgements
The authors thank all the students who contributed to the marmot project and the Surveillance Service of the Park for the logistic support. We thank K. Horvath for revising the English language.
Funding
Open access funding provided by Università degli Studi di Torino within the CRUI-CARE Agreement. At the time of this study, CF was funded with Bourse excellence by UQAM, Government of Québec (CA), and CP was funded with a doctoral fellowship by the University of Pavia (IT).
Author information
Authors and Affiliations
Contributions
CF: conceptualization, data analysis, writing. CP: data collection, reviewing. EC: support with data analysis. LR: data collection, reviewing. GB: reviewing. SB: reviewing. AR: reviewing. BB: reviewing. AvH: conceptualization, data analysis, reviewing.
Corresponding author
Ethics declarations
Statement on the welfare of the animals
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. All procedures performed in this study involving animals were in accordance with the ethical standards of ISPRA (Higher Institute for Environmental Protection and Research, Protocol No. 016970, April 2009) for captures of wild animals. Marmots were trapped using 19 Tomahawk traps located in close proximity to burrow entrances. Traps, opened in the early morning and closed in the evening, were monitored during the day by observers from a distance. Handling of animals lasted for a mean of 30 ± 8 min, and marmots were released in the place of capture as soon as the operations ended. All tagged individuals were observed behaving in the usual way within few hours from capture.
Conflict of interest
The authors declare no competing interests.
Additional information
Communicated by: Quinn Fletcher
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ferrari, C., Pasquaretta, C., Caprio, E. et al. Extrinsic and intrinsic factors affecting the activity budget of alpine marmots (Marmota marmota). Mamm Res 67, 329–341 (2022). https://doi.org/10.1007/s13364-022-00623-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-022-00623-9