
Abstract
Environmental factors have strong influence on activity of alpine ungulates. However, the presence and activities of people in high mountains have been growing rapidly and have led to the advent of human-induced factors, which may modify the time budget. In this study, we examined the influence of natural and human-induced factors on the daytime budget of Tatra chamois Rupicapra rupicapra tatrica. On average, chamois spent 46% of their time foraging, 40% resting, 13% moving around and 1% on social behaviour. The amount of time devoted to particular categories of behaviour was influenced by the time of day, herd size, weather conditions and human disturbance. Human disturbance and the time of day had the highest effect on the proportion of foraging in the daytime budget, which increased as the day progressed and at greater distances from the nearest hiking trail. These two factors also increased the amount of time spent resting, which peaked during the afternoon hours and at greater distances from the nearest trail. The time spent moving around decreased with increasing herd size, distance from the nearest trail and as the day progressed. Males devoted less time to foraging and more to resting and moving around than females. The intensity of human-induced factors is particularly important for a population inhabiting a small, isolated area, as is the case with strictly high-mountain species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The amount of time individual animals allocate to mutually exclusive activities is known as their time budget. It depends on inter- and intraspecific interactions, on the prevailing abiotic conditions (Willmer et al. 2005) and also on the features of an individual animal, such as condition, sex and age. Deciding which activity to undertake depends on the needs and capabilities of a given individual and the availability of time and energy; in consequence, this affects its behaviour and governs its adaptability to the surrounding environment (Illius et al. 2002). The duration of particular activities is a trade-off between necessary activities and those feasible at a given moment (Lind & Cresswell 2005, Sansom et al. 2009).
In most species of social ungulates, males and females live separately except during the reproductive season (Côté et al. 1997, Ruckstuhl & Neuhaus 2002, Singh et al. 2010). This sex-dependent spatial segregation emerges from their different purposes in life: females are responsible for breeding success and raising young and males for reproductive success achieved by the transfer of genes best adapted to a given set of environmental conditions (Clutton-Brock et al. 1987, Bon et al. 1992, Pelletier 2005). Sexual segregation and the ability to form herds are also affected by predation and the availability of food (Jennings & Evans 1980, Dehn 1990). Females escorting young are exposed to a greater risk of predation than adult males, so behaviour is first and foremost governed by the need for safety; once this is assured, then the quality and availability of food become the prime concern. Consequently, family herds are more active in the search for food (Young & Isbell 1991, Bleich et al. 1997). In contrast, the larger and stronger males may remain in habitats with superior food resources even when the threat of predation exists (Main & Coblentz 1990, Miquelle et al. 1992). Other hypotheses explaining sexual segregation in ungulates predict that females of sexually size dimorphic ungulates are expected to select high quality food resources to meet their higher relative metabolic rates, in comparison to males, which may be more efficient in extracting sufficient energy from food resources with a lower quality (Main et al. 1996, Ruckstuhl & Neuhaus 2002). However, due to the trade-off between food quality and quantity on one side and the predation risk on the other side, ungulates show different use of habitats according to sex and age classes (Grignolio et al. 2007).
Predation interferes with the realisation of the time budget (Elgar 1989, Abramsky et al. 2002). Repeated disturbance, for example, being scared away, can adversely affect the balance of energy gains and losses which an individual invests in a particular behaviour or reduce reproductive success as expressed by the number of offspring. The energy wasted as a result of disturbance is kept to a minimum by herd formation: this dilutes the consequences of disturbance among the members of the herd and increases communal vigilance (Messier & Barrette 1985, Hebblewhite & Pletscher 2002). The strategy of herd formation enables each individual to participate in different activities, enhancing defensive effectiveness or achieving a dilution effect and/or confusion effect (e.g. Baruzzi et al. 2017). However, living in groups is expected to lead to increased interactions between individuals competing for resources such as food or sex partners and may also attract more predators (Beauchamp 2014).
Climatic conditions can directly affect animals by altering their physiological, behavioural or ecological reactions. On warm, windless days in arctic or alpine environments, the morning activity of large herbivores can peak earlier and their evening activity later, because their level of heat tolerance is lower than that of species adapted to warmer climates (Belovski & Slade 1986, Aublet et al. 2009, Bourgoin et al. 2011, Brivio et al. 2016). When temperatures are low, or rain or snow is falling, the animals may remain relatively inactive in order to save energy (Loe et al. 2007). Availability of food resources can also change the proportion of foraging in the time budget (Moncorps et al. 1997).
Besides natural factors like climatic conditions, spatial differences in availability/dispersion of food resources or predation pressure, the time budget may be modified by human factors (Podgórski et al. 2013). Pressure due to the activities of hikers, climbers, skiers and the like is present in protected areas, too: the Tatra National Park (southern Poland) is visited by 2.7 million people annually (Czochański 2002). The almost incessant presence of such large numbers of people on the 270 km of marked hiking trails must affect behaviour and ecology of wild animals, including the Tatra chamois Rupicapra rupicapra tatrica (Zwijacz-Kozica et al. 2013, Pęksa & Ciach 2015).
As a consequence, the Tatra chamois population is exposed to a set of natural and human-induced factors that may influence its time budget. The opportunity of simultaneously analysing the influence of a broad spectrum of co-existing factors on the time budget of a species inhabiting a protected area creates a model research system. The aim of the present work is to characterise the daytime budget of the Tatra chamois and to evaluate the influence of the natural factors shaping it, i.e. herd size, time of day, weather and age-sex composition. At the same time, we set up the hypothesis that the anthropogenic factor, i.e. the presence of people on hiking trails within areas used by chamois, is an important factor determining the daytime budget of these ungulates. Previous studies have suggested that human-induced disturbance has significant impact on the population of Tatra chamois (Zwijacz-Kozica et al. 2013, Pęksa & Ciach 2015). Our study evaluates whether the touristic disturbance may increase the proportion of time chamois devoted to movements, while reducing the proportion of time spent foraging and resting in the vicinity of hiking trails. If so, this study may be turned to practical use for controlling the movement of people in areas of great natural value.
Materials and methods
Study area
The Tatras (Carpathians) are the highest mountains in central Europe; they are the only alpine-type range between the Alps and the Caucasus. The Tatra foothills lie at an altitude of around 900 m amsl, while the highest peak, the Gerlach, is at 2655 m amsl. The Tatras cover an area of ca 800 km2: 20% of this is in Poland, the remainder in Slovakia. The whole Tatra massif is protected in the form of national parks: the Slovakian Tatranský Národný Park was formed in 1949 and the Polish Tatrzański Park Narodowy (Tatra National Park—TPN) came into being in 1954. In view of the high level of biodiversity in the Tatras and their unique geomorphology, these mountains have been designated an International Biosphere Reserve and are one of the areas protected by the Natura 2000 network.
The climate of the Tatras is shaped by air masses of different provenance; consequently, the weather can be extremely variable. Sixty-five percent of the air masses flowing towards the Tatras during the year are of polar-maritime origin, while 20% are polar-continental, 6% arctic and 3% tropical. These conditions give rise to abrupt changes in wind strength and direction, cloudiness and the appearance and intensity of precipitation (Limanówka et al. 2008). In the alpine zone of the massif, the mean annual temperature is − 0.7 °C, there is snow cover for an average of 221 days in the year, and the annual total precipitation is nearly 1800 mm.
The climate-vegetation zones in the Tatras change with altitude. At lower elevations (below 1550 m amsl), there are forests dominated by Norway spruce Picea abies, while higher up there is dwarf pine Pinus mugo scrub (to 1800 m amsl). Higher still there are alpine meadows (1800–2200 m amsl) and above them, rocks. Roe deer Capreolus capreolus, red deer Cervus elaphus and wild boar Sus scrofa are present in the Tatras, but they make only sporadic visits to the grasslands above the tree line. Large carnivores (wolf Canis lupus, brown bear Ursus arctos and lynx Lynx lynx) and raptors (golden eagle Aquila chrysaetos) are also present in the study area. As many as 2.7 million people visit the Polish Tatras every year (Czochański 2002). Hiking activity peaks in July and August, which are the summer holiday months (Pociask-Karteczka et al. 2007). Visitors are present on the trails from dawn to dusk, but the highest intensity of tourist activity takes place between 10 am and 2 pm (Buchwał & Fidelus 2010).
Target species
The Tatra chamois is a subspecies threatened with extinction (IUCN category EN—endangered) (Shackleton 1997). It has a small island-like distribution range of some 300 km2 that is limited to the highest parts of the Tatras (Jamrozy et al. 2007). In the 1990s, its numbers fell rapidly from over 1000 animals to barely 200, which caused its threat category in the Polish Red Data Book to be raised to CR—critically endangered. At present (2017), the entire population of the Tatra chamois has recovered to the number of ca 1300 animals (TPN—unpublished data). Results of annual inventories of Tatra chamois conducted in Polish and Slovakian national parks indicate that during the study period (2008–2010), a population size of approximately 750 individuals was estimated, from which 150 were recorded in the Polish part of Tatras (TPN—unpublished data).
The whole population of Tatra chamois inhabits the national park area, which are subject to passive conservation. Unlike other subspecies of the chamois (Knaus & Schröder 1975, Clarke 1986), the Tatra subspecies spends the whole year almost exclusively in the open areas above the tree line and the dwarf pine zone; it does not undertake seasonal vertical migrations (Jamrozy et al. 2007). The island-like nature of the Tatras and their separation from other alpine-like mountain ranges means that the Tatra chamois population is isolated and has a distribution restricted entirely to the highest parts of these mountains. The legal protection of the subspecies and the additional protection afforded by the area’s national park status guarantee the population and its habitat complete protection. The chamois as such are not hunted, nor is their habitat open to hunters, and instances of poaching are extremely few and far between (Jamrozy et al. 2007).
Field methods
Direct observations of chamois were made from early June to late September in 2008–2010. The fieldwork was carried out in the two main regions of the subspecies’ occurrence, which coincide with the areas subject to the strongest tourist pressure in the Polish Tatras. These are the Czerwone Wierchy range (2096 m amsl), along which runs a well-frequented hiking trail, and the vicinity of the Kasprowy Wierch peak (1987 m amsl), to which tourists/hikers can reach along several trails or directly by cable car. In both regions, the observer walked along fixed transects (routed along hiking trails), cutting across the areas where chamois occur (Fig. 1). The transect in the Czerwone Wierchy was 15 km long and covered a relative height of 1200 m, while that in the Kasprowy Wierch region was 18 km long, covering a relative height of 1050 m. Each transect walk began at 06:00 CET and lasted 6–7 h, although long observations of the chamois could prolong it to as much as 14 h. Surveys were done twice a week. A total of 104 surveys were carried out on each transect. To reduce the problem of pseudoreplication, consecutive walks started at opposite ends of the transects to rise the chance of finding different herds (as a herd we considered either group of two or more animals or single individual). Due to the legal restrictions (high regime of protection) and the conservation needs (low population size), the animals remained unmarked.

Map of the study area (Tatra National Park, Carpathians), transects overlap with hiking trails
Behavioural observations were carried out by scan sampling with instantaneous recording (Martin & Bateson 2007). Walking along the transect, the observer started watching a herd through binoculars (10 × 42) as soon as it came into view. The observation of a particular herd ceased after 4 h or earlier when visual contact with it was lost. After the end of the sampling bout (observation session), the observer resumed his walk along the transect in search of further animals. If the herd was recorded again during the same walk, it was excluded from further observation. Since the animals remained unmarked, we used clues as herd size, age-sex composition and natural marks of individuals (horns size and shape, the coat colouration) to recognise particular herd. The behaviour of each individual chamois was recorded at 10-min intervals, 10 s being allotted to categorising the behaviour of each animal. Four categories of behaviour were distinguished: (1) foraging (animals remained at one spot or walked slowly around while consuming portions of food), (2) resting (animals were lying on the ground, ruminating or sleeping), (3) moving around (a directed change in position over a distance without consuming portions of food, i.e. walking or running between valleys/ridges) and (4) social behaviour (interactions, play and communal behaviour, such as animals chasing one another within the herd, jumping up and down, animals sliding on their hind-quarters over patches of snow, mutual sniffing). The ages of the chamois (young, including kids and yearlings, or adult), their sex and herd size were also recorded during each observation bout. The age and sex identification was based on the morphology, i.e. the body size and silhouette, the head and horns size and shape and the coat (fur) colouration (Jamrozy et al. 2007). The positions of the herds observed were marked on a map. The 144 sampling bouts performed during 232 h yielded 16,025 scan samples (records of behaviour).
The weather conditions during each observation bout were recorded, the following categories being applied: cloudiness: 1—cloud cover 0–33%, 2—33–66%, 3—66–100%; rainfall: 1—none, 2—drizzle, 3—a heavy downpour (thunderstorm) or continuous rain; wind: 1—no wind or a gentle breeze, 2—a moderate wind, sufficient to raise dust or small fragments of plants, 3—a strong wind: its whistling was clearly audible and the observer had to make a physical effort to move against it.
Data handling and analyses
Observations in which animals were observed for less than 10 min were discarded. The mean sampling bout duration was 96.7 min (SD = 64.3, n = 144). For each sampling bout, the scan samples of all individuals from a given herd referring to particular categories of behaviour were summed. In order to eliminate the effect of different bout durations of observations of particular herds, the proportion of behaviours in the daytime budget is expressed as a percentage (the sum of a herd’s behaviours was 100%). Percentages were rounded to the nearest integers, in turn allowing us to analyse them through generalised linear models with Poisson errors (link function: log) (Bolker et al. 2009). As 0 and positive integers between 0 and 100 may be ascribed to count data, we feel confident that such an approximation did not alter the biological meaning of our results. The percentage (integer) of each behavioural category, i.e. (1) foraging, (2) resting, (3) moving and (4) social behaviour, was thus treated as a response variable. The explanatory variables were time of day, weather conditions (i.e. cloudiness, rainfall and wind), herd size (the number of animals in a herd) and distance from the nearest hiking trail, which was treated as an index of human pressure. The distance of a herd from the nearest hiking trail was calculated using ArcGIS software (ESRI 2005) in the flat system without regard for relative height, assuming that this is the distance of the centre of the herd at the start of observations from the nearest hiking trail. The age-sex composition of a herd was used as an explanatory variable. Based on the collected data, two age-sex categories were distinguished: (1) groupings consisted of males only or (2) groupings consisted of females with young (kids and/or yearlings). Groupings consisted of females only were not recorded during the study. In model building females with young category was used as a reference level.
Time of day was expressed as the hour in the middle of the observation bout of a particular herd. Principal component analysis (PCA) was used to define the index characterising the weather conditions within the sampling scan. From the variables cloudiness, rainfall and wind (each variable was expressed on a three-point scale), two components were obtained, which were used as new variables: Weather PCA1 and Weather PCA2 (Table 1). Both of these variables explained 52.9% of the total variance. Weather PCA1 was a predictor of high cloud coverage and intense rain, while Weather PCA2 indicated windy conditions with low cloud coverage and less intense rain.
Sex-age herd composition was used as a discrete predictor variable, while time of day, weather conditions, herd size and distance from the nearest trail were used as continuous predictor variables. Distance from the nearest trail considered as an estimate of human disturbance was used as a main effect, and its interaction with herd size and time of the day was tested in the model for each of the behavioural categories. To control multicollinearity between variables, Pearson’s correlation coefficients matrix was calculated (McGarigal et al. 2000). None of the variables correlated (r ≥ 0.5) to one another. Since the response variables were expressed as a percentage of the total daytime budget, three separate models were run for each of the behavioural categories (except for social behaviours, which were excluded from the model building, since this category was recorded in only a small number of observation bouts). As response variables are not independent (the sum of three main activities is approaching 100%), the presented models are considered separately.
Generalised linear models imply linear relationships between any dependent variable and its predictors. However, daily activity budget of chamois may also change in a non-linear manner. Thus, as a further step, we also explored whether distinguished behavioural categories showed potential, non-linear variations by using generalised additive models (Hastie & Tibshirani 1990, Schimek 2000). A smoothing spline was applied to visualise the relationship between the chamois’ behaviour and (1) time of day; (2) herd size; and (3) distance from a trail. The statistical procedures were performed using Statistica software (StatSoft 2011).
Results
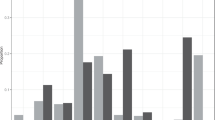
Tatra chamois spent an average of 45.7% of their daytime budget foraging (Fig. 2). Another 40.0% of the time was spent resting and 12.8% on moving around; social behaviours accounted for only 1.5% of the daytime budget. Mean and median herd size was 8.8 individuals (± 8.4 SD) and 5 individuals (quartile range 2–16), respectively. The largest herd consisted of 29 individuals.

Daytime budget of the Tatra chamois Rupicapra rupicapra tatrica in the summer season (Tatra National Park, Carpathians)
Time of day, distance from the nearest trail, herd size and the sex structure of a herd all influenced the proportion of time the chamois spent foraging (Table 2). This proportion increased during the day and with greater distance from the nearest trail (Fig. 3). The proportion of time foraging (ca 50%) was high and relatively constant in herds of more than ten animals (Fig. 3) and at distances between 150 and 400 m from the nearest hiking trail (Fig. 3). At distances greater than 400 m from the nearest trail, the proportion of time spent foraging dropped. Interaction between distance to the nearest trail and time of the day had the highest effect on the proportion of foraging in the daytime budget (Table 2), indicating the effect of trails is influenced by the time of the day. Weather conditions had no effect on the proportion of time the chamois spent foraging (Table 2). Males spent less time foraging than females (Table 2, Fig. 4).

Proportion of the main categories of behaviour (foraging, resting, moving) in the daytime budget of the Tatra chamois Rupicapra rupicapra tatrica depending on the time of day (hour), distance to the nearest hiking trail and herd size during the summer season (Tatra National Park, Carpathians)

Sex-related differences in the daytime budget of the Tatra chamois Rupicapra rupicapra tatrica during the summer season (Tatra National Park, Carpathians)
The amount of time spent resting was affected by time of day, distance from the nearest trail, herd size and the sex structure of the herd (Table 3). In particular, it increased during the day, culminating in the hours after noon (Fig. 3). Proportion of time resting increased along with the distance from the nearest trail. The proportion of the daytime budget devoted to resting was relatively constant and high (ca. 40%) at distances greater than 150 m from the nearest trails (Fig. 3). Interaction between distance to the nearest trail and herd size had significant effect on the proportion of resting in the daytime budget (Table 2). Weather conditions had no effect on the proportion of time the chamois spent resting (Table 3). Males devoted more time to resting than females (Table 3, Fig. 4).
Time of day, distance from the nearest hiking trail, herd size and weather conditions affected the proportion of time the chamois spent moving around (Table 4). The proportion of this behaviour in the daytime budget declined until the afternoon hours, but rose again in the evening (Fig. 3). The larger the herd (Fig. 3) and the greater the distance from the nearest trail (Fig. 3), the less time spent in moving around. The proportion of time spent moving around was at a relatively low but constant level in herds consisting of more than 15 animals and at distances greater than 100 m from hiking trails. Interaction between distance to the trail and time of the day has the highest effect on the proportion of movement in the daytime budget (Table 2), indicating that the effect of trails is strongly influenced by the time of the day. Intense rain and high cloud coverage decreased the proportion of time spent moving (Table 4). Males spent more time moving around (Table 2, Fig. 4).
Discussion
The results of this work show that the diurnal activity of the Tatra chamois is influenced by a series of natural factors, i.e. weather conditions, herd size, time of day, sex structure, and also human-induced ones like distance from the nearest hiking trail. Nearly half of the Tatra chamois’ daily time budget is taken up by foraging. This activity intensifies in the afternoon and evening hours—this is a well-known phenomenon among ungulates (Cederlund 1989; Shi et al. 2003). The studies carried out in the 1960s on Tatra chamois (Briedermann 1967) and recent reports on Alpine chamois Rupicapra r. rupicapra (Brivio et al. 2016) showed that these ungulates exhibited an additional early-morning foraging peak: the present work has not confirmed this. The lack of such a morning foraging peak could be due to disturbance on the part of hikers walking the trails already from the early hours of the morning (Frid & Dill 2002). Nocturnal foraging was thought to be rather rare and occur only where the chamois are subject to additional stress (Briedermann & Štill 1976). In the Western Caucasus, chamois were found to be foraging actively at night, but this was the result of serious disturbance (hunting) during the daytime (Nassimowitch 1949). However, recent data on the activity of Alpine chamois in areas where large terrestrial predators are absent (assuming low level of natural disturbance) indicates that nocturnal activity was detected throughout the year, increasing during autumn (Brivio et al. 2016, Carnevali et al. 2016). Tourist pressure in the Tatras has an adverse effect on the chamois population (Zwijacz-Kozica et al. 2013, Pęksa & Ciach 2015), which could have led to permanent changes in the animals’ behaviour and to a considerable increase in foraging in the evening and presumably also at night. However, further research should be conducted to assess how human disturbance might affect spatio-temporal activity of a diurnal ungulate.
Disturbance can cause a significant decline in the amount of time spent foraging (Gill et al. 2001, Namgail et al. 2007). The proportion of the Tatra chamois’ daytime budget taken up by foraging was nearly 50% in the case of herds consisted of females with young and nearly 30% in the case of groupings consisted of males. These figures are somewhat lower than those recorded for the Cantabrian chamois Rupicapra pyrenaica parva (53% for females and 37% for males) (Perez-Barberia et al. 1997) and for Apennine chamois Rupicapra pyrenaica ornata (60% for females and 63% for males) (Ferretti et al. 2014). This lower proportion of foraging in the daytime budget of the Tatra chamois may be due to the density of hiking trails in the Tatras (2 km of trails/1 km2 area), which means that the animals are very frequently disturbed (Zwijacz-Kozica et al. 2013). This is especially true in the middle of the day, when numerous tourists reach the refuge of the Tatra chamois. It was confirmed by our study, in which the effect of the distance to the nearest trail on daytime activity budget was influenced by the time of the day (a strong interaction between the distance to the nearest hiking trail and time of day). However, the foraging proportion can also drop when temperatures and solar radiation increase during the hot summer months (Bourgoin et al. 2008, Mason et al. 2014). This is particularly visible in adult males which, because of their larger body sizes, are more sensitive to high temperatures and solar radiation levels. As a result, the males move to higher-lying locations in the mountains or seek out shady spots for resting (Aublet et al. 2009).
The weather, especially heavy rain, decreased the amount of time the Tatra chamois spent foraging, and increased the time spent resting. Rainfall wets the coats of ungulates, so in poor weather, they have to expend nearly 30% more energy in order to maintain their normal body temperature (Parker 1988). The animals’ natural reaction is therefore to reduce activity in order to save energy (Loe et al. 2007). The chamois applied a similar effort-avoiding strategy on cloud-free and windless days, when the temperature rose. The rate of metabolism in animals adapted to low temperatures begins to increase at temperatures above 14 °C, and above 20 °C, they begin to pant visibly (Renecker & Hudson 1986). It has also been observed in the Tatras that when low cloud impairs visibility, the chamois foraged very close to the trails without being noticed by hikers (Jamrozy & Pęksa 2004).
The sex structure of herds is important for the allocation of activities within the time budget (Ruckstuhl & Neuhaus 2002, Yearsley & Pérez-Barbería 2005), as our study has confirmed. In accordance with the task budget for gregarious animals (Ruckstuhl & Kokko 2002), males display a greater tendency to rest than the more active females. In contrast, once the males are on the move, they exhibit greater activity than the family herds. At the same time, the probability to move depends on the size of the herd. A smaller herd has to devote more time to locate scattered food resources (Beauchamp 2008), whereas a larger number of foraging animals has the additional advantage of protection from predators as a result of the dilution effect (Shorrocks & Cokayne 2005). Likewise, our results reflect this dependence: the larger the herd, the less time spent in moving around and the more time spent foraging.
A larger herd means better predator detection (Childress & Lung 2003), which reduces the amount of time necessary for scanning the landscape and leaves more time for other activities (Lima 1995, Ferretti et al. 2014). Predator detection can increase when the animals avail themselves of warning signals emitted by another species in the vicinity (Kitchen et al. 2010), e.g. the mutual warnings by marmots Marmota marmota and chamois in the Tatras. However, this advantage can be cancelled out, since large groups of potential prey are more readily discovered (Hebblewhite & Pletscher 2002). Moreover, if the herd is too large, the animals’ level of vigilance may drop as a result of responsibility being delegated to the many members of the herd (Beauchamp 2008), or because of excessive competition for food (Moncorps et al. 1997). Among the Tatra chamois, groups of 2–15 animals were most often seen (Jamrozy et al. 2007), which may either indicate that this is the optimal herd size for functioning in the conditions prevailing in the Tatras or resulted from limited number of individuals present in the population.
The proportion of time spent foraging and resting increased up to 100 m from a hiking trails; this distance can be regarded as a buffer zone beyond which the animals do not feel threatened by people. Beyond 100 m, the portions of time spent foraging and resting were stable at 50 and 45%, respectively. By comparison, the flight distance from humans in the Alpine chamois was found to range from 103 to 180 m (Gander & Ingold 1997). The Tatra chamois’ shorter flight distances may thus be an evidence for its partial adaptation to the almost constant presence of people in its habitats. Study on Apennine chamois indicates that flight distance in the places visited frequently by people was shorter than in the places where animals were less accustomed to people (Patterson 1988). But the very existence of a buffer zone, mentioned above, means that habitat use is limited there, since the animals cannot utilise some part of the existing habitat to its maximum potential. This is particularly important in areas with a high density of hiking trails, as the Tatras, where the trail density of 2 km trail/1 km2 area can lead to limitation of the proportion of the foraging over the area of 40% of the existing habitats.
The behaviour of alpine ungulates is the result of their adaptation to the environmental conditions prevailing in high mountains. The results of this study have shown that the daytime budget of the Tatra chamois is influenced by a set of environmental and social factors. The existence of tourist infrastructure and the consequent human penetration of chamois habitats additionally burdens the animals’ time budget. Any increase in the intensity of human-induced factors is of particular importance for a population inhabiting a small, isolated area, as is the case with strictly high-mountain species.
References
Abramsky Z, Rosenzweig ML, Subach A (2002) The costs of apprehensive foraging. Ecology 83:1330–1340
Aublet J-F, Festa-Bianchet M, Bergero D, Bassano B (2009) Temperature constraints on foraging behaviour of male Alpine ibex (Capra ibex) in summer. Oecologia 159:237–247
Baruzzi C, Lovari S, Fattorini N (2017) Catch me if you can: antipredatory behaviour of chamois to the wolf. Ethol Ecol Evol 29:589–598
Beauchamp G (2008) What is the magnitude of the group-size effect on vigilance? Behav Ecol 19:1361–1368
Beauchamp G (2014) Social predation. How group living benefits predators and prey. Academic Press, Amsterdam and Boston, MA
Belovski GE, Slade JB (1986) Time budgets of grassland herbivores: body size similarities. Oecologia 70:53–62
Bleich VC, Bowyer RT, Wehausen JD (1997) Sexual segregation in mountain sheep: resources or predation. Wildlife Monogr 134:1–50
Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, Stevens MH, White J-SS (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol Evol 24:127–135
Bon R, Gonzalez G, Estevez I, Recarte JM (1992) Comparison of social patterns during the rut within three European populations of Corsican mouflons (Ovis gmelini). Etologia 2:9–20
Bourgoin G, Garel M, Van Moorter B, Dubray D, Maillard D, Marty E, Gaillard J-M (2008) Determinants of seasonal variation in activity patterns of mouflon. Can J Zool 86:1410–1418
Bourgoin G, Garel M, Blanchard P, Dubray D, Maillard D, Gaillard J-M (2011) Daily responses of mouflon (Ovis gmelini musimon × Ovis sp.) activity to summer climatic conditions. Can J Zool 89:765–773
Briedermann L (1967) Zum Ablauf der sommerlichen Aktivitäts-periodik des Gamswildes in freier Wildbahn. Der Zool Garten 33:279–305
Briedermann L, Štill V (1976) Die Gemse des Elbsandsteingebietes. Ziemsen Verlag, Die Neue Brehm-Bücherei 493:1–136
Brivio F, Bertolucci C, Tettamanti F, Filli F, Apollonio M, Grignolio S (2016) The weather dictates the rhythms: Alpine chamois activity is well adapted to ecological conditions. Behav Ecol Sociobiol 70:1291–1304
Buchwał A, Fidelus J (2010) Monitoring ruchu turystycznego przy użyciu czujników ruchu na przykładzie Tatrzańskiego i Babiogórskiego Parku Narodowego. In: Krzan Z (ed) Nauka a zarządzanie obszarem Tatr i ich otoczeniem. Wydawnictwo TPN, Zakopane, pp 45–54
Carnevali L, Lovari S, Monaco A, Mori E (2016) Nocturnal activity of a diurnal species, the northern chamois, in a predator-free alpine area. Behav Process 126:101–107
Cederlund G (1989) Activity patterns in moose and roe deer in a north boreal forest. Holarct Ecol 12:39–45
Childress MJ, Lung MA (2003) Predation risk, gender and the group size effect: does elk vigilance depend upon the behaviour of conspecifics? Anim Behav 66:389–398
Clarke CMH (1986) Chamois movements and habitat use in the Avoca River area, Canterbury, New Zealand. New Zeal J Zool 13:175–198
Clutton-Brock TH, Iason GR, Guinness FE (1987) Sexual segregation and density related changes in habitat use in male and female red deer. J Zool 211:275–289
Côté SD, Schaefer JA, Messier F (1997) Time budgets and synchrony of activities in muskoxen: the influence of sex, age, and season. Can J Zool 75:1628–1635
Czochański JT (2002) Ruch turystyczny w Tatrzańskim Parku Narodowym. In: Partyka J (ed) Użytkowanie turystyczne parków narodowych. Ruch turystyczny – zagospodarowanie – konflikty – zagrożenia. Ojcowski Park Narodowy, Ojców, pp 385–404
Dehn MM (1990) Vigilance for predators: detection and dilution effects. Behav Ecol Sociobiol 26:337–342
Elgar MA (1989) Predator vigilance and group size in mammals and birds: a critical review of the empirical evidence. Biol Rev 64:13–33
ESRI Inc (2005) ArcGIS. Ver. 9.1. Redlands, CA
Ferretti F, Costa A, Corazza M, Pietrocini V, Cesaretti G, Lovari S (2014) Males are faster foragers than females: intersexual differences of foraging behaviour in the Apennine chamois. Behav Ecol Sociobiol 68:1335–1344
Frid A, Dill LM (2002) Human-caused disturbance stimuli as a form of predation risk. Conserv Ecol 6:11 http://www.consecol.org/vol6/iss1/art11
Gander H, Ingold P (1997) Reactions of male alpine chamois Rupicapra r. rupicapra to hikers, joggers and mountainbikers. Biol Conserv 79:107–109
Gill JA, Norris K, Sutherland WJ (2001) Why behavioural responses may not reflect the population consequences of human disturbance. Biol Conserv 97:265–268
Grignolio S, Rossi I, Bassano B, Apollonio M (2007) Predation risk as a factor affecting sexual segregation in Alpine Ibex. J Mammal 88:1488–1497
Hastie TJ, Tibshirani RJ (1990) Generalized additive models. Chapman & Hall, NY
Hebblewhite M, Pletscher DH (2002) Effects of elk group size on predation by wolves. Can J Zool 80:800–809
Illius AW, Tolkamp BJ, Yearsley J (2002) The evolution of the control of food intake. Proc Nutr Soc 61:465–472
Jamrozy G, Pęksa Ł (2004) Numbers, distribution and population changes of the Tatra chamois Rupicapra rupicapra tatrica Blahout, 1971. Nat Conserv 60:63–73
Jamrozy G, Pęksa Ł, Urbanik Z, Gąsienica-Byrcyn W (2007) The Tatra chamois Rupicapra rupicapra tatrica. Tatra National Park, Zakopane
Jennings T, Evans SM (1980) Influence of position in the flock and flock size on vigilance in the starling, Sturnus vulgaris. Anim Behav 28:634–635
Kitchen DM, Bergman TJ, Cheney DL, Nicholson JR, Seyfarth RM (2010) Comparing responses of four ungulate species to playbacks of baboon alarm calls. Anim Cogn 13:861–870
Knaus W, Schröder W (1975) Das Gamswild – Naturgeschichte, Verhalten, Ökologie, Hege und Jagd, Krankheiten. Paul Parey V, Hamburg und Berlin
Lima SL (1995) Back to the basics of anti-predatory vigilance: the group-size effect. Anim Behav 49:11–20
Limanówka D, Cebulak E, Cichocki J, Kilar P, Pyrc R (2008) Informator klimatyczny. 70 lat Wysokogórskiego Obserwatorium Meteorologicznego na Kasprowym Wierchu. IMGW, Kraków
Lind J, Cresswell W (2005) Determining the fitness consequences of antipredation behavior. Behav Ecol 16:945–956
Loe LE, Bonenfant C, Mysterud A, Severinsen T, Øritsland NA, Langvatn R, Stien A, Irvine RJ, Stenseth NC (2007) Activity pattern of arctic reindeer in a predator-free environment: no need to keep a daily rhythm. Oecologia 152:617–624
Main MB, Coblentz BE (1990) Sexual segregation among ungulates: a critique. Wildlife Soc Bull 18:204–210
Main MB, Weckerly FW, Bleich VC (1996) Sexual segregation in ungulates: new directions for research. J Mammal 77:449–461
Martin P, Bateson P (2007) Measuring behaviour: an introductory guide, 3rd edn. Cambridge University Press, Cambridge
Mason TH, Stephens PA, Apollonio M, Willis SG (2014) Predicting potential responses to future climate in an alpine ungulate: interspecific interactions exceed climate effects. Glob Chang Biol 20:3872–3882
McGarigal K, Cushman SA, Stafford S (2000) Multivariate statistics for wildlife and ecology research. Springer-Verlag, NY
Messier F, Barrette C (1985) The efficiency of yarding behaviour by white-tailed deer as an antipredator strategy. Can J Zool 63:785–789
Miquelle DG, Peek JM, Van Ballenberghe V (1992) Sexual segregation in Alaskan moose. Wildlife Monogr 122:1–57
Moncorps S, Boussès P, Réale D, Chapuis J-L (1997) Diurnal time budget of the mouflon (Ovis musimon) on the Kerguelen archipelago: influence of food resources, age, and sex. Can J Zool 75:1828–1834
Namgail T, Fox JL, Bhatnagar YV (2007) Habitat shift and time budget of the Tibetan argali: the influence of livestock grazing. Ecol Res 22:25–31
Nassimowitch AA (1949) Neue Angaben über die Biologie der Gemse im westlichen Kaukasus. Proc Caucasus Nat Reserve 3:51–64
Parker KL (1988) Effects of heat, cold, and rain on coastal black-tailed deer. Can J Zool 66:2475–2483
Patterson IJ (1988) Responses of Apennine chamois to human disturbance. Z Säugetierk 53:245–252
Pęksa Ł, Ciach M (2015) Negative effects of mass tourism on high mountain fauna: the case of the Tatra chamois Rupicapra rupicapra tatrica. Oryx 49:500–505
Pelletier F (2005) Foraging time of rutting bighorn rams varies with individual behavior, not mating tactic. Behav Ecol 16:280–285
Perez-Barberia FJ, Olivan M, Osoro K, Nores C (1997) Sex, seasonal and spatial differences in the diet of Cantabrian chamois Rupicapra pyrenaica parva. Acta Theriol 42:37–46
Pociask-Karteczka J, Baścik M, Czubernat S (2007) Ruch turystyczny w Tatrzańskim Parku Narodowym w latach 1993-2005. In: Kurek W, Mika M (eds) Studia nad turystyką. Tradycje, stan obecny i perspektywy badawcze. IGiGP UJ, Kraków, pp 271–279
Podgórski T, Baś G, Jędrzejewska B, Sönnichsen L, Śnieżko S, Jędrzejewski W, Okarma H (2013) Spatiotemporal behavioral plasticity of wild boar (Sus scrofa) under contrasting conditions of human pressure: primeval forest and metropolitan area. J Mammal 94:109–119
Post E, Stenseth NC, Langvatn R, Fromentin J-M (1997) Global climate change and phenotypic variation among red deer cohorts. Proc R Soc B 264:1317–1324
Renecker LA, Hudson RJ (1986) Seasonal energy expenditures and thermoregulatory responses of moose. Can J Zool 64:322–327
Ruckstuhl KE, Kokko H (2002) Modelling sexual segregation in ungulates: effects of group size, activity budgets and synchrony. Anim Behav 64:909–914
Ruckstuhl KE, Neuhaus P (2002) Sexual segregation in ungulates: a comparative test of three hypotheses. Biol Rev 77:77–96
Sansom A, Lind J, Cresswell W (2009) Individual behavior and survival: the roles of predator avoidance, foraging success, and vigilance. Behav Ecol 20:1168–1174
Schimek MG (2000) Smoothing and regression: approaches, computations, and application. Wiley, NY
Shackleton DM (ed.) & IUCN/SSC Caprinae Specialist Group (1997) Wild sheep and goats and their relatives. Status survey and conservation action plan for caprinae. IUCN, Gland and Cambridge
Shi J, Dunbar RIM, Buckland D, Miler D (2003) Daytime activity budgets of feral goats (Capra hircus) on the Isle of Rum: influence of season, age and sex. Can J Zool 81:803–815
Shorrocks B, Cokayne A (2005) Vigilance and group size in impala (Aepyceros melampus Lichtenstein): a study in Nairobi National Park, Kenya. Afr J Ecol 43:91–96
Singh NJ, Bonenfant C, Yoccoz NG, Côté SD (2010) Sexual segregation in Eurasian wild sheep. Behav Ecol 21:410–418
StatSoft Inc (2011) Statistica (data analysis software system). Version 10.0. Tulsa, OK
Willmer P, Stone G, Johnston I (2005) Environmental physiology of animals, 2nd edn. Wiley-Blackwell, Oxford
Yearsley JM, Pérez-Barbería FJ (2005) Does the activity budget hypothesis explain sexual segregation in ungulates? Anim Behav 69:257–267
Young TP, Isbell LA (1991) Sex differences in giraffe feeding ecology: energetic and social constraints. Ethology 87:79–89
Zwijacz-Kozica T, Selva N, Barja I, Silván G, Martínez-Fernández L, Illera JC, Jodłowski M (2013) Concentration of fecal cortisol metabolites in chamois in relation to tourists pressure in Tatra National Park (South Poland). Acta Theriol 58:215–222
Acknowledgements
We are grateful to Zuzanna Halat, who participated in the fieldwork. We wish to sincerely thank Tomasz S. Osiejuk, Marco Festa-Bianchet, Marco Apollonio and anonymous reviewers for valuable comments on a draft of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The study was carried out in accordance with the approved guidelines and Polish national law. This study was entirely based on observations of animals made from a distance, and additional approval by the local ethical committee was not required.
Additional information
Communicated by: Magdalena Niedziałkowska
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Pęksa, Ł., Ciach, M. Daytime activity budget of an alpine ungulate (Tatra chamois Rupicapra rupicapra tatrica): influence of herd size, sex, weather and human disturbance. Mamm Res 63, 443–453 (2018). https://doi.org/10.1007/s13364-018-0376-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-018-0376-y