
Abstract
Curcuma longa is well known for its use as spice and medicine. The remarkable feature of the plant is the presence of rhizome, which provides an interesting habitat for association by various groups of bacteria. Some of these associated endophytic bacteria can have growth-promoting effects. In the current study, two species of endophytic Paenibacillus has been identified from the rhizome as indole 3 acetic acid producers. These isolates can thus have potential growth-regulating effect in rhizomes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Endophytes are microorganisms that live in the internal tissues of plants without altering the normal functioning of the tissues (Stone et al. 2000). These microorganisms are unique in their adaptations to specific chemical environment of the host plant. Some of these microorganisms have the molecular machinery for the synthesis of host plant-specific compounds and are thus considered as an unexplored source of natural products (Strobel 2003). The host plants are also benefited by the endophyte-mediated production of plant growth regulators, resistance to diseases and also endophyte-assisted phytoremediation.
Curcuma longa is a rhizomatous plant of the family Zingiberaceae. Its rhizome is well known for its medicinal properties: anti-inflammatory, antiulcerogenic and antitumor activities (Miquel et al. 2002). The active constituents of turmeric rhizome include curcumin and various volatile oils, including tumerone, atlantone and zingiberone. In addition to its chemical complexity, rhizome is biologically interesting because of its ability to stay viable for a long time under adverse conditions (Ramirez-Ahumada Mdel et al. 2006). At the same time, the rhizome is least explored in terms of its biochemical processes at various stages of its development. The close contact of the rhizome with the soil largely favors the association by various communities of microorganisms as endophytes. These associated microbes can have a remarkable role in rhizome physiology and can have varying roles in regulating plant growth.
Microbial association as endophytes has been reported from a variety of plants. Variations in the endophytic community of plants are also known with regard to the plant source, age, type of tissue, season of sampling and environment. Usually, the concentration of the endophytic bacteria is higher at the root than at shoot tissue. A large variety of plant species are shown to be endophytically associated with bacteria such as Pseudomonas, Bacillus, Azospirillum, etc. (Chanway 1996). Several genera of endophytic bacteria have been isolated from different plants, including Aerobacter, Aeromonas, Agrobacterium, Chryseomonas, Curtobacterium, Enterobacter, Erwinia, Flavimonas and Sphingomonas (Sturz et al. 1997). Studies also showed the presence of endophytic diazotrophs, Acetobacter diazotrophicus, Herbaspirillum sp., Burkholderia sp., Enterobacter sp. and Klebsiella sp., associated with sugarcane plants (Boddey et al. 2003). So, by considering the remarkable features of turmeric rhizome, the presence of a diverse and even specific bacterial association can well be expected.
The functional roles of endophytes are studied only from limited cases, though their presence has been reported from a wide variety of plants. The production of plant growth-promoting molecules like indole 3 acetic acid (IAA) is an important contribution of endophytic microorganisms (Spaepen et al. 2007). IAA is synthesized mainly through a coupled transamination and decarboxylation of l-tryptophan (l-Trp) by microorganisms and plants (Bhavdish et al. 2003). IAA can stimulate both rapid responses such as cell elongation and long-term responses such as cell division and differentiation of plants tissues (Taghavi et al. 2009). Previous studies have shown that IAA plays a major role in controlling various physiological processes in plants including apical dominance, tropism, shoot elongation and root initiation. Due to its importance in plants, endophyte-mediated production of IAA has gained a great deal of attention.
In the case of Curcuma longa, the presence of rhizome is expected to provide a specialized habitat for the association of a diverse group of bacteria with potential impact on plant growth. This makes studies on isolation and characterization of endophytic bacteria from turmeric much more interesting and informative. In the current study, two endophytic Paenibacillus sp. were isolated from turmeric rhizome and both of them were found to have the ability to produce IAA as confirmed by HPLC analysis.
Rhizomes of turmeric (Curcuma longa) collected from Navajyothisree Karunakara Guru Research Centre for Ayurveda and Siddha, Uzhavoor, Kottayam were used as the source for isolating endophytic bacteria. The rhizomes were cleaned with tap water to remove soil and were made into 1–2 cm long pieces. These were further treated with Tween 80 for 10 min with vigorous shaking and followed by washing with distilled water several times to remove Tween 80. The samples were then dipped in 70 % ethanol for 1 min and further treated with 1 % sodium hypochlorite for 10 min. The samples were then washed several times with sterilized distilled water and the final wash was spread onto a nutrient agar plate (g/L; peptone 5, beef extract 2, yeast extract 3, sodium chloride 5 and agar 18, pH 7.0) as control. For the isolation of endophytic bacteria, the outer surface of the sterilized plant material was trimmed and placed onto nutrient agar plates. Nutrient agar is one of the commonly used media for the isolation of endophytic bacteria (Aravind et al. 2009). All plates including the control were incubated at room temperature for 5 days and observed periodically for bacterial growth. Those experiments in which bacterial growth was observed in the control plate were completely discarded. Morphologically distinct colonies as identified by colony characters were selected, purified and used for further studies.
For the molecular identification, genomic DNA was isolated from the bacterial isolates and used as template for PCR. Genomic DNA isolation of the obtained bacterial strains was conducted as per the method described by Ausubel et al. (1995). The isolates were first cultured overnight in Luria–Bertani broth and the cells were harvested from 1.5 mL culture by centrifugation. The harvested cells were resuspended in 567 μL of TE buffer and lysed using 30 μL of 10 % SDS and 3 μL of 20 mg/mL proteinase K. The mixture was then incubated for 1 h at 37 °C. The lysate was mixed thoroughly with 100 μL of 5 M sodium chloride and 700 μL chloroform:isoamyl alcohol (24:1) and centrifuged at 8,000 rpm for 10 min. After centrifugation, the aqueous layer was transferred without disturbing the interface to a fresh tube and equal volume of isopropanol was added. This was then inverted several times and centrifuged at 8,000 rpm for 10 min. The pellet was washed in 70 % ethanol (v/v) and air dried at room temperature. The dried DNA pellet was resuspended in 100 μL TE buffer and visualized in 0.8 % agarose gel (w/v). Primers used for the amplification of part of 16S rDNA was 16SF (5′-AgA gTT TgA TCM Tgg CTC-3′) and 16SR (5′-AAg gAg gTg WTC CAR CC-3′) and was selected based on a previous report of Chun and Goodfellow (1995). PCR was carried out in a 50 μL reaction volume containing 50 ng of genomic DNA, 20 pM of each primer, 1.25 units of Taq DNA polymerase (Bangalore Genei), 200 μM of each dNTPs and 1× PCR buffer. PCR was carried out for 35 cycles in a Mycycler™ (Bio-Rad, USA) with an initial denaturation at 94 °C for 3 min and cyclic denaturation at 94 °C for 30 s, annealing at 58 °C for 30 s and extension at 72 °C for 2 min with a final extension of 7 min at 72 °C. The PCR product was checked by agarose gel electrophoresis, purified and further subjected to sequencing. The sequence data was checked for similarity analysis with BLAST program (Zhang et al. 2000). The phylogenetic analysis of the 16S rDNA sequence of the isolates obtained was carried out with MEGA 5 using neighbor-joining method with 1,000 bootstrap replicates (Tamura et al. 2011).
For screening IAA production, the bacterial isolates were inoculated into 20 mL of nutrient broth supplemented with 0.2 % (v/v) of l-tryptophan and incubated for 10 days at 28 °C. After incubation, the culture was centrifuged at 3,000 rpm for 20 min to collect the supernatant. Initially, 1 mL of supernatant was mixed with 2 mL of Salkowski reagent and the tubes were incubated in dark as described earlier (Rahman et al. 2010). The development of red color was observed after 1 h of incubation. Uninoculated growth medium was used as negative control. The isolates positive in the color reaction were further inoculated into 200 mL of nutrient broth supplemented with 0.2 % (v/v) of l-tryptophan and incubated for 10 days at 28 °C. After incubation, the supernatant was collected by centrifugation at 3,000 rpm for 20 min. The supernatant was then acidified to pH 2.5–3.0 with 1 N HCl and extracted twice with equal volume of ethyl acetate. The extracted ethyl acetate fraction was vacuum dried in a rotary evaporator at 40 °C. The dried powder was dissolved in 1 mL of methanol (MeOH) and stored at −20 °C. Crude extracts made from the isolates were spotted on a thin layer chromatographic (TLC) plate (Silica gel Gf 254, thickness 0.25 mm) along with standard IAA as control. The solvent system n-butanol:ethyl acetate:ethanol:water (3:5:1:1) was used as the mobile phase. The plates were developed by spraying paradimethyl aminobenzaldehyde reagent and incubated at 50 °C for 1 h. The Rf values of the extracts were compared with those of the standard for identification as the methods explained by Chaiharn and Lumyong (2011). Further confirmation of the presence of IAA in the methanolic extract of culture supernatant was conducted by reverse-phase high pressure liquid chromatography (HPLC) analysis on a Supelcosil LC-18 column with a flow rate of 1 mL min−1 as described by Jensen et al. (1995). Elution was performed with a mixture of H2O and MeOH (60:40), both containing 0.5 % acetic acid. Elution was monitored at 280 nm by a Shimadzu UV–Vis detector model SPD 10A.
The surface sterilization procedure was quite satisfactory for isolation of endophytic bacteria as no growth appeared on the control plate. Also, bacterial growth was obtained in the nutrient agar plates which were inoculated with the rhizome piece. Based on the distinct colony characteristics, two isolates were obtained from the rhizome of turmeric and named as ClB1 and ClB2. The colony of ClB1 was small, pinpoint and yellow pigmented, and that of ClB2 was small, lobate margined and cream colored.
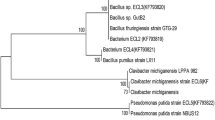
Molecular identification of the isolates was done by the sequencing of 16S rDNA gene. The presence of 1,500 bp product as observed by agarose gel electrophoresis confirmed the amplification of 16S rDNA gene. The PCR product was gel eluted, sequenced and the sequence data were submitted to NCBI under the accession numbers (JN835217 and JN835218). As the 16S rDNA gene sequence provides accurate grouping of organism even at subspecies level, it is considered as a powerful tool for the rapid identification of bacterial species (Clarridge 2004). The sequence data thus obtained was subjected to BLAST analysis. The sequence similarity analysis of 16S rDNA sequence of ClB1 and ClB2 showed its maximum identity of 99 % to Paenibacillus favisporus and Paenibacillus sp., respectively. So the isolates ClB1 and ClB2 can be considered as strains coming under the Paenibacillus genus. The phylogenetic analysis of 16S rDNA sequence of the isolates along with the sequences retrieved from the NCBI was carried out with MEGA 5 using the neighbor-joining method with 1,000 bootstrap replicates. The result of phylogenetic analysis showed distinct clustering of the isolates and confirms the results of the sequence similarity analysis (Fig. 1).

Phylogenetic analysis of 16S rDNA sequences of the bacterial isolates (ClB1 and ClB2) from Curcuma longa along with the sequences from NCBI. The analysis was conducted with MEGA5 using neighbor-joining method
Endophytic Paenibacillus sp. have been found associated with various plants like Pinus sp. and Coffea arabica (Bent and Chanway 2002; Vega et al. 2005). The observations of Scherling et al. (2009) confirmed experimentally the association of Paenibacillus sp. with the root of in vitro grown poplar plants. However, the information on the presence of two species of Paenibacillus associated with Curcuma longa as endophyte is novel. This highlights the significance of the isolates obtained in the study.
The culture supernatant of CB1 and CB2 were found to have the presence of IAA by conducting the preliminary screening using Salkowski reagent and this was further confirmed by TLC and HPLC. In the TLC analysis, the pure IAA and the extracts were found to have Rf value of around 0.62. The result was finally confirmed by HPLC where the pure IAA produced a peak at 9.3 min retention time (Fig. 2) and the crude extract had a predominant peak at comparable retention time (Figs. 3, 4). Thus, the result of HPLC confirmed the production of indole 3 acetic acid by the bacterial isolates ClB1 and ClB2 (Paenibacillus sp.) from turmeric.

HPLC chromatogram for IAA standard carried out using water and methanol, both containing 0.5 % acetic acid in the ratio 60:40 on reversed phase C18 column with a flow rate 1 mL min−1 and detected under UV at 280 nm

HPLC chromatogram for the extract from isolate ClB1 carried out using water and methanol, both containing 0.5 % acetic acid in the ratio 60:40 on reversed phase C18 column with a flow rate 1 mL min−1 and detected under UV at 280 nm

HPLC chromatogram for the extract from isolate ClB2 carried out using water and methanol, both containing 0.5 % acetic acid in the ratio 60:40 on reversed phase C18 column with a flow rate 1 mL min−1 and detected under UV at 280 nm
Some endophytic microorganisms have the potential to synthesize IAA. For the microbial synthesis of IAA in tryptophan-dependent route, tryptophan is used as the precursor. There are different pathways which can lead to tryptophan-dependent microbial production of IAA. The various pathways for IAA biosynthesis include tryptophol, tryptamine, indole-3-pyruvic acid and indole-3-acetamide pathways (Gravel et al. 2007). The presence of Paenibacillus sp. as endophyte and its ability to produce IAA have already been reported from diverse plants (Lebuhn et al. 1997; Timmusk and Wagner 1999). Acuña et al. (2011) reported the increase in the production of IAA by tenfold in the low-nutrient (diluted) LB medium than in nutrient-rich medium by Paenibacillus sp. However, the presence of Paenibacillus sp. as endophytic bacteria from turmeric and their ability to synthesize IAA have not yet been reported. This makes the present finding much more interesting. Thus, two strains of Paenibacillus sp. identified in the study can have important growth-regulating role in the rhizome of turmeric. The strains may also have unique features to survive in the specific chemical environment of the rhizome.
In conclusion, the results of this study demonstrated that, (1) two different strains of Paenibacillus sp. are endophytically associated with the rhizome of Curcuma longa and (2) both the endophytic Paenibacillus sp. strains (ClB1 and ClB2) have the ability to form IAA as confirmed by HPLC analysis. Hence this isolate can be considered as having possible growth-promoting effect in Curcuma longa.
References
Acuña JJ, Jorquera MA, Martínez OA, Menezes-Blackburn D, Fernández MT, Marschner P, Greiner R, Mora ML (2011) Indole acetic acid and phytase activity produced by rhizosphere bacilli as affected by pH and metals. J Soil Sci Plant Nutr 11(3):1–12
Aravind R, Dinu A, Eapen SJ et al (2009) Isolation and evaluation of endophytic bacteria against plant parasitic nematodes infesting black pepper (Piper nigrum L.). Indian J Nematol 39(2):211–217
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K (1995) Short protocols in molecular biology, Chap 2.4. Wiley, New York
Bent E, Chanway CP (2002) Potential for misidentification of a spore-forming Paenibacillus polymyxa isolate as an endophyte by using culture-based methods. Appl Environ Microbiol 68:4650–4652
Bhavdish N, Johri A, Sharma J, Virdi S (2003) Rhizobacterial diversity in India and its influence on soil and plant health. Adv Biochem Eng Biotechnol 84:49–89
Boddey RM, Urquiaga S, Alves BJR, Reis V (2003) Endophytic nitrogen fixation in sugarcane: present knowledge and future applications. Plant Soil 252:139–149
Chaiharn M, Lumyong S (2011) Screening and optimization of indole-3-acetic acid production and phosphate solubilization from Rhizobacteria aimed at improving plant growth. Curr Microbiol 62:173–181
Chanway CP (1996) Endophytes: they’re not just fungi! Can J Bot 74:321–322
Chun J, Goodfellow M (1995) A phylogenetic analysis of the Genus Nocardia with 16s rRNA Gene Sequences. Int J Syst Bacteriol 45:240–245
Clarridge JE (2004) Impact of 16S rRNA gene sequence analysis for identification of bacteria on clinical microbiology and infectious diseases. Clin Microbiol Rev 17:840–862
Gravel V, Antoun H, Tweddell RJ (2007) Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: possible role of indole acetic acid (IAA). Soil Biol Biochem 39:1968–1977
Jensen JB, Egsgaard H, Van Onckelen H, Jochimsen BU (1995) Catabolism of Indole-3-Acetic Acid and 4- and 5-Chloroindole-3-Acetic Acid in Bradyrhizobium japonicum. J Bacteriol 177:5762–5766
Lebuhn M, Heulin T, Hartmann A (1997) Production of auxin and other indolic and phenolic compounds by Paenibacillus polymyxa strains isolated from different proximity to plant roots. FEMS Microbiol Ecol 22:325–334
Miquel J, Bernd A, Sempere JM, Diaz-Alperi J, Ramirez A (2002) The curcuma antioxidants: pharmacological effects and prospects for future clinical use. A review. Arch Gerontol Geriat 34:37–46
Rahman A, Sitepu IR, Tang S-Y, Hashidoko Y (2010) Salkowski’s reagent test as a primary screening index for functionalities of Rhizobacteria isolated from wild dipterocarp saplings growing naturally on medium-strongly acidic tropical peat soil. Biosci Biotechnol Biochem 74:2202–2208
Ramirez-Ahumada Mdel C, Timmermann BN, Gang DR (2006) Biosynthesis of curcuminoids and gingerols in turmeric (Curcuma longa) and ginger (Zingiber officinale): identification of curcuminoid synthase and hydroxycinnamoyl-CoA thioesterases. Phytochemistry 67:2017–2029
Scherling C, Ulrich K, Ewald D, Weckwerth W (2009) A metabolic signature of the beneficial interaction of the endophyte Paenibacillus sp. isolate and in vitro-grown poplar plants revealed by metabolomics. Mol Plant Microbe Interact 22:1032–1037
Spaepen S, Vanderleyden J, Remans R (2007) Indole-3-acetic acid in microbial and microorganism–plant signaling. FEMS Microbiol Rev 31:1–24
Stone JK, Bacon CW, White JF (2000) An overview of endophytic microbes: endophytism. In: Bacon CW, White JF (eds) Microbial endophytes. Marcel Dekker, New York, pp 3–30
Strobel G (2003) Endophytes as sources of bioactive products. Microbes Infect 5:535–544
Sturz AV, Christie BR, Matheson BG, Nowak J (1997) Biodiversity of endophytic bacteria which colonize red clover nodules, roots, stems and foliage and their influence on host growth. Biol Fertil Soils 25:13–19
Taghavi S, Garafola C, Monchy S, Newman L, Hoffman A, Weyens N, Barac T, Vangronsveld J, van der Lelie D (2009) Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees. Appl Environ Microbiol 75:748–757
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28:2731–2739
Timmusk S, Wagner EG (1999) The plant-growth-promoting rhizobacterium Paenibacillus polymyxa induces changes in Arabidopsis thaliana gene expression: a possible connection between biotic and abiotic stress responses. Mol Plant Microbe Interact 12:951–959
Vega FE, Pava-Ripoll M, Posada F, Buyer JS (2005) Endophytic bacteria in Coffea arabica L. J Basic Microbiol 45:371–380
Zhang Z, Schwartz S, Wagner L, Miller W (2000) A greedy algorithm for aligning DNA sequences. J Comput Biol 7:203–214
Acknowledgments
This study was supported by the Department of Biotechnology (DBT), Government of India under the DBT-RGYI support scheme.
Conflict of interest
The authors declare that they have no conflict of interest in the publication.
Author information
Authors and Affiliations
Corresponding author
Additional information
A. J. Aswathy and B. Jasim contributed equally to the work.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Aswathy, A.J., Jasim, B., Jyothis, M. et al. Identification of two strains of Paenibacillus sp. as indole 3 acetic acid-producing rhizome-associated endophytic bacteria from Curcuma longa. 3 Biotech 3, 219–224 (2013). https://doi.org/10.1007/s13205-012-0086-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13205-012-0086-0