
Abstract
The UN Sustainable Development Goal (SDG) Target 2.5 explicitly calls for the maintenance of genetic diversity of seeds through soundly managed and diversified seed and plant banks at national, regional, and international levels as an essential undertaking to end global hunger (SDG Goal 2). This special issue results from a renewed call to demonstrate the value-in-use of conserving and supplying plant genetic resources conserved in genebanks to researchers, plant breeders, and farmers. We present these studies as a collective contribution to a relatively small body of literature that highlights not only the importance of crop plant diversity managed by genebanks but also the diversity of genebank functions and uses. In this overview, we begin by restating foundation concepts that economists have applied to study the value of crop genetic resources conserved as genebank accessions. We then provide a synthesis of previous research on genebank values from the late 1990s until the present. We summarize the main messages of the studies included in this special issue of Food Security and explain how they contribute to a better understanding of the role, function, and value of genebanks, particularly as we address food security challenges in a changing agricultural context. Finally, we draw implications for further applied research and policy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Aims of this special issue of Food Security on genebanks
Evidence regarding the economic returns of plant genetic resource use in breeding and crop improvement is very strong: the social value of new and improved crop varieties has been documented continuously for many decades. By contrast, applied research about the value of conserving the diversity of plant genetic resources outside their place of origin (ex situ, in genebanks) is scant.
Why? One explanation is that genebanks have long-term objectives and are physically remote from eventual development outcomes (Crop Trust 2015). In fact, the essential operations of genebanks and their social utility are poorly understood. Further, even when we are able to assess the impact of improved varieties grown by farming households on food security, it is not easy to disentangle the specific contribution of the genetic resources supplied by the genebanks in the ancestry of the varieties. Our analysis of the literature suggests that the economic value of genebanks is likely to be understated when derived only from the yield impacts of varieties on farms. Diverse crop varieties, and the germplasm they embody, contribute many other benefits to farmers and society as a whole, such as reducing the potential economic costs wrought by plant pests and diseases as well as climatic variability.
Some historical perspective helps to understand the economic importance of the genebanks and the diversity they conserve. During the Green Revolution, Harlan (1972) and others expressed fears that valuable genes would be lost as farmers replaced their landraces with more genetically uniform, modern varieties. The diffusion of modern varieties occurred on a massive scale and entire farming systems were reshaped with greater use of fertilizers and controlled irrigation. At that time, modern varieties were intentionally bred to be responsive to fertilizer under favorable moisture conditions. Many of these new varieties, such as early semi-dwarf wheat varieties bred by Norman Borlaug, also incorporated genetic resistance to major diseases, including stem rust.
The Southern Corn Leaf Blight epidemic that occurred in the United States in 1970 illustrates the crop devastation that may occur when large areas are planted to uniformly, genetically susceptible plants (Bruns 2017; National Research Council 1972). Then, as now, crop geneticists and breeders often hypothesized that rare, locally adapted plant genotypes might be found among the varieties maintained by farmers in isolated, extreme, or heterogeneous environments. They led extensive and intensive efforts to collect diverse seed samples of landraces and crop wild relatives, protecting them for future use as ‘accessions’ in cold storage units. Public organizations, such as the Food and Agriculture Organization of the United Nations, and private foundations such as the Ford and Rockefeller Foundations, contributed to these efforts.
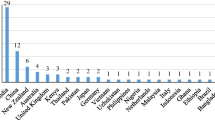
Over the last 50 years, the CGIAR (formerly the Consultative Group for International Agricultural Research) research centres have built up the largest and most frequently accessed network of global crop collections (Fig. 1). Coordinated by the CGIAR Genebank Platform and in partnership with the Global Crop Diversity Trust,Footnote 1 the 11 genebanks of the CGIAR currently manage more than 773,000 accessions of genetic materials of cultivated plants and crop wild relatives that are crucial to achieving worldwide food and nutrition security (Table 1). While this represents only 10% of the total accessions stored in about 1750 genebanks around the world, the CGIAR genebanks account for about 95% of the germplasm distributed within the guidelines of the International Treaty on Plant Genetic Resources for Food and Agriculture (ITPGRFA) (ITPGRFA 2020).

The CGIAR genebanks and where they are located. Source: CGIAR Genebank Platform (2019)
These efforts have not proceeded without controversy. During the late 1990s, Raeburn (1995) referred to genebanks as little more than ‘seed morgues’. Criticism mounted that materials stored in genebanks were rarely used because little was known about them (Plucknett et al. 1987; Wright 1997). With economic theory, Simpson et al. (1996) demonstrated that excess capacity, redundancy, or availability of close substitutes drives the marginal value of an individual genetic resource toward zero. While accepting this argument, several studies undertaken in the early 2000s contested the viewpoint that genebanks were ‘unused’ and developed some analytical approaches for assessing genebank values (Gollin et al. 2000; Johnson et al. 2003; Zohrabian et al. 2003; Koo et al. 2004; Day-Rubenstein et al. 2006). Researchers focused on the large collections housed in the CGIAR system and the US National Plant Germplasm System (US NPGS), both of which serve a global mandate, although a number of case studies of national genebanks were also conducted during that period (e.g. Virchow 2003; Kingwell et al. 2001).
Since then, we find only a few studies that explore the value of genebanks in the peer-reviewed literature, and these are not directly related to agricultural development in lower income countries. Xepapadeas et al. (2014) explored the insurance value of the Greek genebank, and an article by Tyack and Ščasný (2018) applied stated preference methods to value accessions in the Czech genebank. Meanwhile, the technical and operational management of genebanks progressed enormously. Tools were developed to provide geneticists with new means of searching for useful traits and new ways of using materials. McCouch et al. (2012) describe how genotyping and sequencing can enable scientists to document the genetic identity of their accessions, identify duplicates and optimize collections (also see article by Gollin, in this issue). The availability of genomics data will likely lead to innovative, international research and breeding partnerships (Halewood et al. 2018).
However, research on the value of international genebanks and the genetic diversity held in the collections has not kept pace. Schreinemachers et al. (2014) published a cost analysis conducted for the World Vegetable Center (AVRDC). Gotor and Caracciolo (2010) applied an econometric framework to data provided by the International Rice Research Institute (IRRI) to investigate the effects of the in-trust agreements on germplasm distributions. Their results were consistent with the hypothesis that there was a changing point in germplasm distributions before and after 1994, when the in-trust agreements (ITAs) were enacted. A significant drop in distributions was observed from 1989 to 1994, followed by a growing trend after ITAs were established and the policy environment became more stable. Given the sample data at hand, they were unable to draw conclusions regarding the effects of the policy change on the value of the material held in-trust. Galluzzi et al. (2016) concluded that countries could use a wide diversity of germplasm from many other countries as inputs into their crop improvement programmes if they are provided with the opportunity of facilitated access to genetic resources. Similarly, the authors were also unable to draw conclusions regarding the value of the international germplasm distributions with the type of data used. They did, however, highlight the importance of non-monetary benefits, such as information exchange, technology transfer, and capacity building, in the global movement of germplasm.
Concurrently, the geopolitical and environmental context in which genebanks conduct their work has evolved in new directions. We believe that public concern for the physical security of seed samples, for the rate of loss of remaining landraces and wild relatives due to threatened habitat, and for finding ways to address human needs caused by weather extremes are even greater today than they were two decades ago when earlier work was published. The UN Sustainable Development Goal (SDG) Target 2.5 explicitly calls for the maintenance of genetic diversity of seeds through soundly managed and diversified seed and plant banks at national, regional, and international levels as an essential undertaking to end global hunger (SDG Goal 2). SDG Goal 2 recognizes that genebanks are critical to the achievement of global food security. Target 2.5 further acknowledges that because food security depends on the food production adapted to local farming environments and suited to the needs of local societies, genebank systems must encompass a diversity of plants, crops, and commodities.
With this special issue in Food Security, our principal aim is to update the evidence and enhance the understanding of the role and value of genebanks in a changing agricultural context. The studies presented in this special issue were undertaken in partnership with genebank staff and scientists at the International Rice Research Institute (IRRI) in the Philippines, the International Center for Maize and Wheat Improvement (CIMMYT) in Mexico, the International Potato Center (CIP) in Peru, the International Center for Tropical Agriculture (CIAT) in Colombia, the International Livestock Research Center (ILRI) in Ethiopia, World Agroforestry (ICRAF) in Kenya, and the Pacific Community’s Centre for Pacific Crops and Trees (CePaCT) in Fiji. The analyses are interdisciplinary in nature, employing either quantitative or qualitative methods, or both. In addition, we invited lead articles from Gollin to provide a theoretical economics perspective on changing technologies and from Tyack and Ščasný to offer another example of a genebank valuation study based on stated preference approaches in a developed economy.
2 Concepts and definitions
Genebanks perform a broad range of functions, from collecting new plant materials to conserving them in medium- and long-term storage, testing seed health, conducting research to characterize and evaluate the resistance of plant materials to disease, and distributing plant materials to plant breeders, research scientists, and farmers (Popova 2018). Many have argued that genetic resources stored in genebanks are invaluable; seeking to value them in economist’s terms is therefore purposeless (see examples in Wilson 1988). We do not entirely agree with this view; indicators of value and measurements of the magnitude of value are crucial elements for decision making when resources are scarce and choices must be made. That said, valuing genetic resources is notoriously challenging.
Our definition of economic value is anthropocentric because plant domestication and agricultural production are human activities. When discussing the diversity of crop genetic resources, we refer to both phenotypic and genotypic variation within and among all agricultural crop species, including varieties recognized as agro-morphologically distinct by farmers and as genetically distinct by plant breeders. We include crop wild relatives (Aberkane et al.; Tyack and Ščasný in this issue) and any other plant material that is a basic unit of heredity (or ‘genetic material’), such as semi-natural accessions of forages (Kitonga et al. in this issue). The plant genetic resources that are our focus are conserved as germplasm or ‘accessions’ in genebanks are typically stored as seeds, seedlings, tissue, and other forms that contain genetic information and are being distributed and used by farmers, scientists, and breeders.
Several studies in this collection also refer to farmers’ varieties or landraces when constructing the genealogies of improved varieties (Bernal-Galeano et al.; Villanueva et al. in this issue), discussing crop genetic diversity (Alexandra et al.; Bernal-Galeano et al. in this issue), or presenting a dynamic model of conservation (Ocampo et al. in this issue). Typically, farmers’ varieties do not satisfy breeders’ or legal definitions of variety because these varieties are heterogeneous and segregate genetically. Farmers’ varieties are often called ‘landraces’, while those bred by professional plant breeders are termed ‘modern varieties’. Definitions for landraces are numerous in the crop science literature. Harlan (1992) defined them broadly as variants, varieties, or populations of crops as maintained by farmers, with plants that are often highly variable in appearance. The genetic structure of landraces is shaped by farmers’ seed selection practices and management as well as by natural selection processes over generations of cultivation. A plant phenotype is the observable manifestation of a genotype. A genotype is determined by its alleles, or types of genes. Morphology refers to physical characteristics or form.
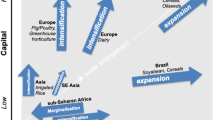
To understand the impacts of a genebank, we need first to understand the sources of value in plant genetic resources. Economists are often asked to value plant genetic resources because these resources are limited. When resources are limited, any choice is a value judgment. One way to conceptualize sources of value is the total economic value framework developed by resource economists (Fig. 2 is one version of the framework). Total economic value includes both use and non-use components. Use value can be direct, deriving from the food, fibre, and medicinal products to which plant genetic resources contribute; use value can also be indirect, relating to the habitat or ecosystem. Both direct and indirect values have current and future expected values. A third component of use value is option value. Option value derives from uncertainty and implies the flexibility to respond to unexpected future challenges. Depending on the articulation of the framework, cultural services may be included in use or option values.

Total economic value framework. Source: authors
Non-use values are typically composed of existence and bequest values. Existence value refers to the satisfaction felt simply from knowing that something exists, whether it is used or not. Bequest value is defined as the satisfaction gained by individuals from knowing that others will have the opportunity to enjoy an asset in the future. Endowing a genebank as a trust for future generations (e.g. The Crop Trust) might be viewed as an explicit recognition of bequest value. It has been argued, however, that the existence value of plant genetic resources is appreciated by relatively few individuals. Unlike ‘charismatic species’ such as polar bears (e.g. in Svalbard), plant genetic resources are conserved precisely because they are thought to embody genes and gene combinations of future use to human society through crop improvement.
Years ago, an eminent resource economist admitted that he was ‘awed by how little economics could contribute to valuation of genetic resources’ (Brown 1990). He explained that since most value components do not trade in markets, they have no market prices. Even when only the value components related to use are considered, few are measurable with quantities and observable prices. A genebank collection, as compared to a breeder’s working collection, is maintained largely to respond to future demand. Thus, the expected future use and option value components of use value are sizeable in a genebank collection. Option value is more difficult to assess empirically than expected future use, which can be calculated from historical records. We do know that option value cannot be negative in sign.
Second, the commercial value of plant genetic resources may represent a relatively small component of their total use value when markets function poorly. In many developing agricultural economies, farmers obtain seed outside of formal markets, and although there is considerable value in the plant genetic resources they use, they may not sell their harvest.
Third, each market-based analysis is generally fixed in time and projections are based on assumptions that might not bear out. Tastes and preferences are dynamic. Production shocks occur from which plant genetic resources that are rarely used in crop improvement derive substantial economic value, such as a genetic material that confers resistance to a new pathogen.
Finally, since the potential usefulness of any single genetic resource is often highly uncertain, and time horizons for developing products from genetic resources are long, economists generally contend that private investors under-invest in conserving them at the levels needed by society. As a consequence, the public sector has played a pivotal role in conserving these resources and will continue to do so in the foreseeable future.
Revisiting these concepts today, we recognize that genebanks have multiple functions that are not strictly limited to the conservation of plant genetic resources, including the documentation and provision of scientific information for further research. We have particular concern for endangered crop wild relatives found in natural habitats or interspersed in agricultural and peri-urban landscapes beset by weather extremes.
3 Literature review
Considering the breadth of sources of value that can be ascribed conceptually to plant genetic resources, the empirical documentation of these values is remarkably narrow. Below, we provide a synthesis of this literature here, by the general research question addressed. Details on theory and methods are found in the original sources.
3.1 What is the value of genebanks in crop improvement?
Economists have repeatedly demonstrated that the continuous release of improved varieties by plant breeding programmes has brought economic returns that far outweigh the costs of investment. Most of these net benefits have been generated by publicly funded institutions. Society and consumers have especially benefited from lower food prices, particularly in less advanced economies where consumers spend a larger share of their budget on food. Estimated rates of return to investment are high (often within the 40–60% range, e.g. Alston et al. 2000; Evenson 2001; Evenson and Gollin 2003; Raitzer and Kelley 2008; Renkow and Byerlee 2010). Research on farm-level adoption and impacts of improved varieties is also voluminous. A smaller set of studies explores the effects of certain categories of plant genetic resources (e.g. landraces) and indices of variety diversity or genealogical diversity on yield or yield risk (e.g. Widawsky and Rozelle 1998; Smale et al. 1998, 2008; Di Falco and Chavas 2009).
Among studies that assessed the economic impact of using plant genetic resources in crop improvement, we know of only a handful that sought to relate productivity changes in farmers’ fields to genebank accessions. Evenson and Gollin (1997) consulted the genealogies of rice varieties released by national programmes and IRRI from 1965 to 1990, correlating productivity changes with changes in IRRI programmes in an econometric model. They estimated that adding 1000 cataloged accessions was associated with the release of 5.8 additional varieties. Assuming a 10-year lag for variety development and a 10% discount rate, they calculated that these new accessions generated a present value (in 1990 dollars) of USD $325 million. A similar study by Gollin and Evenson (1998) focused on India.
Johnson et al. (2003) estimated that 49% of bean area in Latin America was planted to varieties associated with CIAT and genebank distributions in 1998, with an incremental value of production of USD $177 million. Robinson and Srinivasan (2013) estimated the benefits of a high-yielding cassava variety (Kasetsart 50) in Southeast Asia and a high-yielding potato variety (Cooperation 88) in China, linking these explicitly to genebank accessions. They found aggregate economic benefits nearing USD $100 million from the diffusion of Kasetsart 50, arguing that these would not have been achieved without the use of the genebank. Benefits accruing to Cooperation 88 in China were predicted to reach nearly USD $500 million per year, with poor people earning the largest share.
The methodological approach in most studies of economic benefits of farm productivity gains builds on the compendium by Alston et al. (1998), although methodological challenges continue to be debated and refined. Partial equilibrium analysis, economic surplus methods, and econometric approaches are commonly used. Econometric models of yield response and production functions may be estimated, incorporating indices of genetic diversity or ancestry as explanatory variables. Pedigree data and genealogies have been employed in combination with data on variety transfers or with farm survey data or secondary data measured at a district or regional scale. Forms of hedonic analysis were also applied to explore the value of genetic resource attributes.
There are several inherent limitations to this literature. First, the literature on crop productivity documents the value of plant genetic resources used in plant breeding only for commercial agriculture. For example, the value of crops not often used by breeding programmes, of which relatively little scientific research has been conducted (sometimes referred to as ‘neglected and underutilized crops’), is not included. Also, semi-subsistence farmers, or farmers in remote areas who do not sell their crops, are not represented. Crop wild relatives are also not represented, unless they were included as ancestors in the breeding programme.
Second, analysts face challenges when estimating the share of the productivity gain that is attributable to genetic advance. The genetic component is only one of many factors that affects yields in farmers’ fields.
Third, even when genetic gains can be estimated, apportioning these among the progenitors of the variety involves ‘rules of thumb’, such as Mendelian rules of inheritance. The Mendelian rules of inheritance assume that each parent in the pedigree of a variety contributes equally in each generation, ignoring both the effects of random genetic drift and the effect of selection by breeders from crosses for traits of economic interest. Both processes of attributing yield gains in farmers’ fields to their genetic component and apportioning the genetic component to individual ancestors are data-intensive. They require accurate genealogies, specialized trial data, or farm survey data with variety-specific information on areas and production. Studies of the value of genebanks in crop improvement need to able to link a genebank accession directly to an improved variety grown in farmer’s fields and the production outcome of research interest.
3.2 What is the value of a collection?
Spurred by debates over whether the expansion of genebanks was justified, Gollin et al. (2000) developed a model that portrays the relationship between genebank size and the search for new traits. Applying the model to data assembled at CIMMYT about search costs and areas planted to wheat that was potentially susceptible to the Russian wheat aphid, they simulated various scenarios to represent a range of adoption, cost, and benefit outcomes. Russian wheat aphid spread from its centre of origin in the Caucasus and Central Asia to numerous geographical areas, including the US, South Africa, parts of the Southern Cone of Latin America, and North and East Africa, but searches of advanced lines at USDA revealed the near absence of resistance in materials originating outside Central Asia. Across the scenarios, the range in discounted, expected net benefits was huge (from USD $1.2 to $165.8 million), depending mostly on the time lag from discovery of resistant material to adoption. When the time lag was as short as 7 years, the expected net benefits justified a search that was larger than the total number of wheat landraces in the CIMMYT genebank.
In a second example, a trait of value was found in a tiny subset of the world’s collection of genetic resources. Searching a tiny sample of Iranian landraces from the region of pest origin for resistance to Russian wheat aphid increased net benefits substantially, revealing the value of specialized knowledge and passport information. Although such sub-collections may be searched rarely, there are reasons for storing them ‘unused’ for years.
The third example (Septoria tritici, which causes leaf blotch) demonstrated that it may be economically optimal for plant breeders to search their own collections before they demand unimproved materials from genebanks—and this in no way implies that genebank accessions are less valuable. Despite the superior distribution of resistance among accessions of emmer wheat the relative costs of search and trait transfer from emmer were too high, considering the technology available at that time.
The examples presented by Gollin et al. (2000) revealed that much of the value of large collections in crop improvement is derived from rare traits of economic importance. Payoffs may be modest in the short-term but great over the longer term when both predictable and unpredictable challenges must be addressed. These results hint at insurance and option values.
The authors adapted the search theory framework used previously by Evenson and Kislev (1975) and Simpson et al. (1996), which relies on numerical methods and Monte Carlo simulations applied to data on probability distributions for traits, search costs, and benefits of successful search. The probability of finding a targeted trait is sensitive to the frequency distribution of the desired trait. This distribution depends on the size of the collection and the trait distribution in the underlying plant population. The discounted stream of future benefits depends on many factors, including the time required for plant breeders to effectively transfer the new source of resistance into the variety, the time needed for the improved variety to pass any regulatory hurdles before release, and the time lag between release and actual use by farmers. Costs depend very much on the tools used to find and confer sources of genetic resistance.
A limitation of the Gollin et al. (2000) study was that the authors were not able to examine the overall distributions of resistance across numerous traits, search costs, and benefits streams in order to draw more general conclusions about the optimal size of a genebank. Today, the model could be used to demonstrate the value of using more advanced genetic information, such as genotyping, in search processes. Gollin et al. (2000) did not estimate the expected value of an additional genebank accession. This question, discussed next, is relevant for the decision to expand a collection, discard an accession, or restrict the use of an accession through imposition of property rights.
3.3 What are the costs of genebank conservation?
The costs of conserving accessions in genebanks are relatively easy to tabulate compared to their expected benefits. The studies compiled by Koo et al. (2004) confirmed the relatively low costs of maintaining large collections of some major crops. Their analysis showed that the present value of conserving and distributing an accession into perpetuity varied significantly by crop, reflecting in part its reproductive biology. For example, the costs of conserving an existing accession of maize was estimated at USD $141 in 1996 with 6% initial regeneration, compared to only USD $10 for an accession of wheat. Comparable figures for wild groundnut at the International Crops Research Institute for the Semi-Arid Tropics (ICRISAT) were USD $19 for an existing accession, USD $28 for cultivated rice, and USD $49 for wild rice. At CIAT, conserving an accession of beans into perpetuity was estimated to cost USD $13 in 2000 with a 6% regeneration, compared to USD $23 for an accession of forages. Wages, plant maintenance, and operational costs also differ with the location of the physical plant.
Other cost studies of national genebanks have been implemented (see Virchow 2003; Kingwell et al. 2001) and reveal variation in conservation cost by method of conservation, crop, and genebank location. Horna et al. (2010) built on the work of Koo et al. (2004) by developing a decision-support tool aimed at enhancing the cost-effectiveness of collection management. Again, the authors found that the reproductive biology of the crop is a major determinant of the relative costs of operations, particularly the cost of regeneration. Characterizations and regeneration were often the most resource-intensive operations for materials that are seed-propagated. The average costs of some operations are similar across materials (e.g. distribution and seed health testing, which follows a formula). Distribution costs were higher than expected and import clearance was a large component of costs. The authors concluded that molecular characterization to prevent duplication or for pre-breeding purposes added value to the collection.
The theoretical approach of Koo et al. (2004) was cost minimization. They showed that if the costs of conserving an accession are lower than any sensible lower-bound estimate of benefits, undertaking the expensive and challenging exercise of benefits estimation is not necessary to justify its conservation. The methodology of Horna et al. (2010) followed the overall approach of Koo et al. (2004).
3.4 What is the value of an individual genebank accession?
There are many instances in which a single plant genetic resource has proved to have large commercial value by conferring a specific trait, but these cannot be generalized. Well-known examples include the Rht 1 and Rht 2 dwarfing genes, which were bred into modern wheats via Norin 10 and whose original source was named Daruma, a Japanese landrace. Sr2, the gene known as ‘Hope’, conferred resistance to devastating stem rust and was found in a genetic material named Yaroslav Emmer. Documented examples among crop wild relatives include the wild tomato and sunflower (see subsection 3.7). Even in these cases, since the genes interact with the genetic background of each line into which they are bred, estimating the value of the specific trait with accuracy would require carefully designed trials to compare the performance of lines that are otherwise identical.
Generally, in the process of plant breeding numerous genetic resources are continually shuffled and reshuffled in an uncertain search for traits that must be well expressed in a crop variety destined for highly differentiated conditions of production. Economically important traits are distributed statistically across plant genetic resources, with varying likelihood of attaining useful levels. The traits demanded by societies, such as resistance to plant pests and diseases, and quality attributes preferred by consumers also change frequently in response to environmental stress and economic changes. Breeding products (crop varieties) contain many ‘ingredients’ that are also genetic resources. These products are combined in turn with others to produce the next variety. The marginal contribution of the last resource used may be slight.
The uniqueness of an individual accession also matters for value. The same trait may be apparent to one degree or another in multiple plant genetic resources. Even when rare in a given collection, accessions carrying useful traits may be duplicated among seed samples (accessions) in multiple collections. Seed samples of the same genetic resource may also be found in more than one genebank collection and in more than one political jurisdiction.
To say that the expected value of an individual accession may not be ‘enormous’ does not imply that its expected benefits do not justify the costs of conserving it. Because many factors outside the control of genebank managers influence the magnitude of benefits from finding and transferring traits into crop varieties, predicting the benefits of conserving an individual accession, and deciding whether to keep or discard it based on economics principles (equating marginal expected benefits to marginal expected costs), is not easy.
To our knowledge, the work by Zohrabian et al. (2003) was the first and only published example of an attempt to estimate the marginal value of an accession. The authors found that the expected marginal benefit from exploring an additional, unimproved accession in breeding soybean varieties resistant to cyst nematode was small but more than covered its acquisition and conservation costs. They found an expected benefit-cost ratio in the range of 36–61 for investing in an additional accession to prevent losses from a single pest. Findings justified the expansion of the US soybean collection.
Zohrabian et al. (2003) also drew on the search theoretic framework but applied a maximum entropy approach to model the distribution of a quantitative trait when data are sparse. By applying the decision rule of Koo et al. (2004), they found that the lower-bound benefit from utilizing a marginal accession was higher than the upper-bound cost. A limitation of the analysis was that it involved only a single trait, although individual accessions may be useful in the search for more than one trait.
3.5 What is the value of accession information?
The lack of useful data about accessions was cited as an obstacle to greater utilization of accessions in plant breeding and scientific research when the valuation work on genetic resources began (Wright 1997). Evaluation data are of great value to plant breeders seeking to improve traits related to biotic or abiotic stress. Koo and Wright (2000) asked when genebank managers should decide to evaluate genetic materials and whether new technological tools might change this decision. Employing the example of plant disease, they showed that the benefit of ex ante evaluation is largest when the likelihood of infection is at an intermediate rather than minimum or maximum level. When a disease is rare, the cost of searching today is large relative to the expected present value of future benefits. When disease is expected to occur soon, host plant resistance will be evaluated in any case and the importance of timing is reduced. Examples of this situation include breeding for nonspecific resistance against the rusts of wheat. The marginal benefit of technology breakthroughs that reduce the time spent evaluating for resistance traits is larger when the development process begins earlier, favoring ex ante evaluation.
Based on a survey of requestors of genetic materials from the US NPGS, Day-Rubenstein et al. (2006) tested the effect of accompanying data on the share of seed samples reported to be ‘useful’. They found that accompanying data improved the chances that a sample received was used within a five-year period in a breeding programme, whether evaluated or used in other ways. A survey of users of the Musa International Transit Center (ITC) revealed that receipt of ITC materials allowed them to save time and resources by basing research design on previous results (Garming et al. 2010).
The theoretical model proposed by Koo and Wright (2000) is mathematical, with no empirical application. The main limitation of the model is that the authors consider only the evaluation of resistance based on single genes. The analyses of the US NPGS and the Musa ITC were each based on distribution data and user surveys. US NPGS data were sufficient to estimate an econometric model. The major constraints of the user surveys were low response rates, which were difficult to raise above 35–40%, regardless of the survey tool used.
3.6 What is the value of germplasm flows from collections?
Studies by Evenson and Gollin (1997) and Johnson et al. (2003) documented the reliance of individual countries on genetic materials obtained from numerous countries via genebanks and nurseries of the CGIAR. For example, of the 18 countries studied by Johnson et al. (2003) in Latin America, 11 received over 70% of the genetic material in their released varieties from other countries. Only 8.5% of 1709 rice varieties studied by Evenson and Gollin (1997) had been developed entirely from own-country progenitors.
Fowler et al. (2001) showed that developing countries were net recipients of germplasm samples from six of the CGIAR genebanks (CIAT, CIMMYT, ICARDA, ICRISAT, ILRI, and IRRI). More than 80% of the materials that had been distributed by that time, which totaled about one million samples, went to organizations in developing countries. Most of these were universities and national agricultural research systems. Galluzzi et al. (2016) analyzed 25 years of distribution data from the CGIAR. They found that developing and transition economies dominated as recipients, utilizing transferred germplasm within their public agricultural research systems, and development programmes.
User surveys generally contradict the notion that genebanks are rarely used. For example, the US NPGS survey indicated higher rates of direct utilization in plant breeding than had been suggested in earlier studies, secondary to use through sharing within and outside respondents’ institutions, and proportionately higher use rates among respondents in low- and middle-income countries (Smale and Day-Rubenstein 2002). Within the brief five-year period covered by respondents, 11% of germplasm samples received had already been incorporated into breeding programmes, another 43% were still being evaluated, and 19% were reported as useful in other ways, leaving 28% categorized as ‘not useful’. Garming et al. (2010) found a continuous increase in the number of accessions available for distribution and in the number of samples distributed by the Musa ITC. The ITC had distributed germplasm to over 100 different countries since its foundation. For a number of countries, the ITC is the only source of superior Musa germplasm. In countries with strict quarantine, survey respondents reported that they could not have conducted their research at all without the ITC, since it was the only legal source of germplasm.
Of the earlier studies of germplasm flows, only Johnson et al. (2003) and Evenson and Gollin (1997) reported economic values, and these were simply estimated. User surveys have documented use of resources conserved in genebanks, which has implications for use value, but have not estimated economic value per se with the exception of the study on willingness-to-pay by Tyack and Ščasný (2018). Tyack and Ščasný (2018) estimated the willingness-to-pay for conserving additional crop varieties in the genebank for 10 years by the Czech population and the population in the agricultural region of South Moravia. A major finding of their research is that Czechs who are provided with information about the meaning and importance of crop diversity would be willing to pay a total of USD $68 million crop conservation during the coming decade. This amount represents about 4.5 times more than current conservation costs.
3.7 What is the value of crop wild relatives?
Tyack and Dempewolf (2015) summarized the findings of a number of studies that estimated the economic value of crop wild relatives in crop improvement. Annual benefit estimates ranged from USD $8 million to USD $165 billion (2012), for activities ranging from providing genes from wild tomato to contributing to the world economy. However, Tyack and Dempewolf noted that the reported estimates were based, in most cases, on ‘back-of-the-envelope’ calculations. In a number of cases, the entire value of the increase in the yield or quality of the finished variety was attributed to the wild crop relative with the known trait. Tyack and Dempewolf cited Prescott-Allen and Prescott-Allen (1986) as seeking to apportion the value more carefully by source and recommended a study on wild coffee by Hein and Gatzweiler (2006), who took costs into consideration.
Tyack and Dempewolf (2015) also proposed a new conceptual framework for analysis. They argue that previous studies focused narrowly on production value resulting from the introgression of wild genetic material and suggested that researchers consider the cost reduction due to reduced use of pesticides and herbicides, or nutrients, such as phosphorus or nitrogen. They recommend efforts to measure positive externalities, such as reduced emissions of carbon dioxide from lower use of fertilizers, pesticides, herbicides, and irrigation. Carbon savings have a dollar value in carbon markets. Reduced threat of habitat loss because of less need for water might be valued. Health benefits from reducing toxic substances in runoff are measurable.
4 Contributions of this special issue
We consider that in terms of methods and scope, there are five essential contributions of the applied studies in this collection to earlier research on genebank valuation. The first is that several lead authors were able to trace the ancestry of modern varieties adopted by farmers directly to specific genebank accessions and apportion or measure indicators of benefits by drawing from extensive, digitized information on pedigrees (Villanueva et al.; Bernal Galeano et al.; Selitti et al.).
Villanueva et al. focus on the contribution of the International Rice Genebank (IRG) at IRRI to rice-farming households in India, using survey data from nearly 9000 farmers. Supplemented by extensive pedigree and evaluation data, the authors examine the genetic composition of improved rice varieties that are derived from the genes of IRG accessions and adopted by farmers. Villanueva et al. extend a classic econometric model of farm productivity by applying a framework that tests the relationship of genebank provenance to productivity changes while controlling for the effects of other farm inputs and environmental factors. The authors also take a step further by documenting the genetic contribution to the pedigrees of not just one but a total of 45 varieties adopted on over 95% of the study area in Eastern India. Alongside the positive productivity effect associated with genebank provenance, they are able to document a broadening in the latent diversity of leading varieties grown as measured by the coefficient of parentage.
Bernal-Galeano et al. combined analyses of biological, agricultural, and market-related data to measure the contribution of the genebank at the CIP to the development of improved potato varieties released in low-income countries. They developed a new algorithm, the Relative Contribution of Provenance (RCP), and apply it to pedigree data to apportion the contribution of CIP genebank to Pallay poncho and Victoria varieties. Pallay Poncho is used to validate and test the restrictiveness of RCP measurement because all the ancestors used for its development were provided by the CIP genebank collection; Victoria was selected due to the availability of pedigree information and data on adoption rates and other economic parameters in Uganda. The authors then use an economic surplus approach to attribute the yield gain from Victoria, value the economic benefits, fits, and quantify the genebank contribution. This study also highlights the global interdependence in germplasm use, since genetic materials originally from the Andes region and conserved at CIP in Peru generated economic benefits from improved potato varieties grown in Africa. Uganda is only one of several African countries that cultivates Victoria. Global interdependence as a source of key genetic features is likely to become even more important as a means of coping with climate extremes and weather variability in agriculture.
In the aftermath of the 1990s genocide in Rwanda and paucity of written records, Selitti et al. faced a more challenging task in constructing pedigrees. CIAT played a crucial role in restoring diversity lost in farms, and Rwanda was the first country recipient of iron-biofortified beans. The authors investigate the contribution of the genebank at CIAT to the development of seven iron-biofortified varieties of climbing beans and document the links to the genebank through pedigree analysis and interviews with bean breeders. Selitti et al. apply econometric approaches to test the impact of farm adoption on the yield, consumption, and purchase of beans using a farm-level dataset of nearly 1400 households in Rwanda. The results confirm the importance of genetic diversity conserved in the bean collection at CIAT for developing of beans with high amounts of micronutrients and provide evidence of the links of the genebank to nutritional outcomes.
The analysis by Aberkane et al. represents a second major contribution of the studies included in this collection: the analysis of wild, semi-natural, and landrace genetic materials. Like the studies on improved varieties mentioned above, Aberkane et al. tapped large-scale datasets and search pedigrees to trace accessions of ‘goat grass’, a wild relative of wheat, to modern varieties grown on farms through the development of synthetic hexaploid wheat (SHW). SHW, which resulted from the crossing of durum wheat and goat grass, bridges the transfer of desirable traits into improved varieties of bread wheat. Over 1500 SHWs were developed by wheat scientists and the derivatives represented 21% of the germplasm distributed between 2000 and 2018 by the genebank of CIMMYT. The authors also surveyed wheat breeders to confirm the adoption of 85 SHW-derived varieties in 13 countries. Their study illuminates the potential of pre-breeding to cope with the challenges of agricultural change. Because pre-breeding is a long-term process, it should be strategically planned and designed to ensure ready access of a set of targeted, adaptive, and economically important traits for plant breeders. The authors emphasize that the case of ‘goat grass’ shows the key role of genebank collections in conserving crop wild relatives for pre-breeding.
Kitonga et al. examine farmer-perceived benefits derived from the adoption of semi-natural tree species by livestock and maize farmers. Calliandra and Gliricidia are the two most popular fodder tree species in Kenya. The availability of high quality fodder is one of the major constraints in livestock production, while poor soil fertility is a serious limiting factor to maize productivity. According to the authors, these constraints could be addressed through the adoption of a tree-based production system that incorporates the use of high quality tree species. Through a survey of key informants and genebank users, the authors provide anecdotal evidence of improved food security, higher incomes, increased milk production, reduced vulnerability to drought, and enhanced soil fertility due to the adoption of Calliandra and Gliricidia. The study also validate the unique role of the genebank at ICRAF as the main source of tree germplasm for many Kenyan smallholders and points to the under-recognized potential of trees in ameliorating agricultural outcomes from policy interventions in developing countries.
The study by Ocampo-Giraldo et al. represents both the second and third contribution of this collection. The authors present a unique circular model of conservation that involves combining ex situ and in situ approaches. These are discussed in the specific context of Jala, a maize landrace of Mexico named after its geographical origin. Jala has the largest ear and tallest plant of all maize landraces in the world, but due to changes in local economic and environmental conditions, the genetic erosion of this ancestral landrace is now becoming evident. Without a conservation strategy, evolutionary processes and farmers’ selection practices over generations of cultivation have resulted in a landrace with shorter ears and plants. Together with genebank staff and scientists from CIMMYT, the authors interviewed various stakeholders, including maize experts, CIMMYT and national agricultural research systems (NARS) breeders, farmers, consumers, and local youth groups to elaborate on the cultural and economic significance of Jala. Dynamic conservation combines both in situ and ex situ conservation methods and supports exchange of germplasm where farmers’ decision-making process regarding germplasm conservation and use is considered and where crop diversity conserved by local communities is made available to other users through the international maize genebank at CIMMYT.
Alexandra et al. provides the fourth contribution of this collection. The authors narrated the formation of CePaCT in Fiji, underscoring the role of a global effort to collect, conserve, and breed taro in response to the Pacific’s version of the Irish potato famine, the Taro Leaf Blight (TLB) catastrophe, that occurred in Samoa in the 1990s. Their effort led to the development of improved, disease-resistant clones. The authors conducted ethnobotanical fieldwork in Fiji, Vanuatu, and the Cook Islands and interviewed more than 50 taro experts and farmers. The TLB story highlights the intricate inter-organizational cooperation and global commitment needed to safeguard a prestige crop of the Pacific and find solutions to agricultural challenges.
The unique valuation study by Tyack and Ščasný makes the fifth contribution. For the first time in this literature, the authors estimate the willingness of the Czech population to pay for the information services generated by the activity of pre-breeding accessions, including the wild relatives of wheat. Finally, Gollin questions the “value” of valuation exercises—instead, the economics of prioritization is the most important direction for future research on genetic resource conservation. He also reminds us that despite attractive claims that physical collections can be readily replaced but digital records of DNA sequence, the value of physical seed samples likely extends beyond the DNA code.
When the seed samples analyzed here were originally collected from the fields of local farmers who developed them and conserved as genebank accessions, they were ‘priceless’. Yet, each of the empirical studies presented here illustrates how, because of genebanks, these accessions can be made tremendously valuable in addressing the challenges of global food security.
5 Implications
This collection of studies attests to the value of genebanks in numerous ways. Several studies estimate genebank value in physical or monetary terms. For example, Villanueva et al. estimate that a 10% increase in the genetic contribution of IRG accessions to an improved rice variety grown by rice farmers in Eastern India is associated with a yield increase of 27%. At the same time, the average coefficient of parentage of all pairwise combinations among the 10 most adopted rice varieties is only 0.0973, reflecting crossing germplasm and multiple, functional trait combinations received from 19 countries of origin. Bernal-Galeano et al. estimate a gross benefit of the Victoria potato variety in Uganda at USD $1.04 billion dollars (2016), which exceeds the annual operational cost of the entire genebank many times over. Seventy-two percent corresponded to the CIP genebank contribution. Tyack and Ščasný estimate that the Czech population is willing to pay half of the current budget of the genebank on pre-breeding over 10 years.
Given the difficulty in measuring the monetary value of genebank accessions, it is significant that each of these studies reveals a point that is relevant to the 2020 SDGs and to science policy in a changing agricultural context. SDG Target 2.5 highlights the contribution of genetic diversity of seeds and cultivated plants and their wild relative species as well as the promotion of access and benefit-sharing arising from the use of genetic resources to achieving SDGs. Ocampo et al. document the role of the CIMMYT genebank in combatting genetic erosion in a Mexican landrace while seeking to empower local communities as custodians of the resource. The example of Victoria—a potato variety that connects the Andes to the East African Highlands—demonstrates that the availability of diverse germplasm, as is found at CIP, is perhaps the most important element in varietal development. The role of CIAT’s genebank in transferring germplasm for development of iron-biofortified beans in Rwanda confirms this point. Micronutrient deficiency is one of the three types of nutritional challenge we face today, along with persistent undernutrition and obesity (SDG Targets 2.1 and 2.2). The riveting story of how scientists overcame Taro Leaf Blight emphasizes, once again, that global cooperation is needed to protect farming systems against vulnerability from genetic uniformity (SDG Target 2.A). Kitonga et al. record farmer-perceived benefits and farmer reliance on the ICRAF collection for semi-natural fodder trees with multiple on-farm and environmental benefits (SDG Target 2.3). Aberkane et al. clearly show how pre-breeding techniques can be deployed to transfer genetic diversity from a wild crop relative and confer resistance to pathogens and pests, yield, and nutritional advantages (SDG Target 2.4). Villanueva et al. provide evidence that the genebank contributed to rice diversity on farms, which has positive ramifications for reducing genetic vulnerability in a rice-farming system based on improved varieties.
In which directions might applied research lead us from here? The most gnarly issue in our analyses—and a long-standing challenge—is how to apportion genetic contributions of genebank accessions to improved varieties. With improved data and information management tools, it should be much easier to track the supply and exchange of genetic materials, maintain accurate pedigrees, and compute genetic contributions using conventional Mendelian principles. Yet, we now know more about the multiple mechanisms through which genetic expression is conferred (Gollin, this issue) and are increasingly stymied by the assumptions behind conventional Mendelian principles. New approaches for apportioning genetic contributions are needed if we are to achieve more precise estimation.
Finding a better way to apportion genetic contribution is only one example of how genebank valuation is fundamentally multidisciplinary. From its conception to its completion, safeguarding crop diversity involves many disciplinary perspectives and approaches. In addition, the global genebank system is only a single player in the global system for crop conservation. This present set of studies indicates the involvement of international nurseries, plant breeding programmes, and genetics resource experts, partners in national research systems, community organizations, youth groups, individual consumers, and farmers. Genetic conservation involves many players in many jurisdictions, and this contributes to the complexity of the policies and decision making needed to promote it. In this collection, only one study by Ocampo et al. addresses in situ, on-farm conservation. Much more work is needed on the topic, including valuation studies building on those conducted years ago (e.g. Birol et al. 2006).
Progress has now been made in establishing the necessary standards and protocols to conserve genebank accessions. Genebank services that promote use are hypothesized to add value, such as supplying characterization and evaluation data, systematic tracking of users, and feedback from users that enables providers to better respond to the changing demand for accessions. Future applied research could seek to estimate value added by such services.
Alexandra et al. stress the importance of conserving and supplying clean and healthy germplasm to reduce the devastation of diseases carried in planting material. While conservation is a key function of genebanks, an underlying assumption is that the materials stored and distributed must be clean, viable, and health-tested. The reliance of farmers on seeds from ICRAF because they are assured of quality also supports this. There is a huge opportunity to raise global awareness on how protecting plant health can contribute to development goals by estimating the value of this genebank activity.
We hope that this special issue contributes to a better understanding of the role, function, and value of genebanks, as we address food security challenges in a changing agricultural landscape.
Change history
01 September 2020
The original version of this article unfortunately contained mistakes.
19 January 2021
The article “Valuing genebanks”, written by Melinda Smale and Nelissa Jamora, was originally published Online First without Open Access.
26 January 2021
A Correction to this paper has been published: <ExternalRef><RefSource>https://doi.org/</RefSource><RefTarget Address="https://doi.org/" TargetType="URL"/></ExternalRef> <ExternalRef><RefSource>https://doi.org/10.1007/s12571-021-01144-0</RefSource><RefTarget Address="10.1007/s12571-021-01144-0" TargetType="DOI"/></ExternalRef>
Notes
The Global Crop Diversity Trust (https://www.croptrust.org/) is an international organization based in Bonn, Germany. Established in 2004 by Bioversity International, on behalf of CGIAR and the UN Food and Agriculture Organization, the Crop Trust helps support the global conservation system by providing long-term grants through an endowment fund.
References
Alston, J. M., Norton, G. W., & Pardey, P. G. (1998). Science under scarcity: Principles and practice for agricultural research evaluation and priority setting. Wallingford: CAB International.
Alston, J. M., Chan-Kang, C., Marra, M. C., Pardey, P. G., & Wyatt, T. J. (2000). A meta-analysis of rates of return to agricultural R&D: Ex pede herculem? IFPRI research report no. 113. Washington, DC: International Food Policy Research Institute.
Birol, E., Smale, M., & Gyovai, Á. (2006). Using a choice experiment to estimate farmers' valuation of agrobiodiversity on Hungarian small farms. Environmental and Resource Economics, 34(4), 439–469.
Brown, G. M. (1990). ValuingGenetic Resources. In G. H. Orians, G. M. Brown, W. E. Kunn, & J. E. Swierzbinski (Eds.), The preservation and valuation of biological resources (pp. 203–226). Seattle: University of Washington Press.
Bruns, H. A. (2017). Southern corn leaf blight: A story worth retelling. Agronomy Journal, 109, 1–7.
CGIAR Genebank Platform. 2019. https://www.genebanks.org/genebanks/. Accessed 1 December 2019.
Crop Trust. (2015). Securing crop diversity for sustainable development. Bonn: Global Crop Diversity Trust.
Day-Rubenstein, K., Smale, M., & Widrlechner, M. (2006). Demand for genetic resources and the U.S. National Plant Germplasm System. Crop Science, 46, 1021–1031.
Di Falco, S., & Chavas, J.-P. (2009). On crop biodiversity, risk exposure, and food security in the highlands of Ethiopia. American Journal of Agricultural Economics, 91(3), 599–611.
Evenson, R. E. (2001). Economic impacts of agricultural research and extension. In B. Gardner & G. Rausser (Eds.), Handbook of agricultural economics (pp. 573–628). Amsterdam: Elsevier.
Evenson, R. E., & Gollin, D. (1997). Genetic resources, international organizations, and improvement in rice varieties. Economic Development and Cultural Change, 45(3), 471–500.
Evenson, R. E., & Gollin, D. (2003). Crop variety improvement and its effect on productivity: The impact of international agricultural research. Wallingford: CABI Publishing.
Evenson, R. E., & Kislev, Y. (1975). Agricultural research and productivity. New Haven: Yale University Press.
Fowler, C., Smale, M., & Gaiji, S. (2001). Unequal exchange? Recent transfers of agricultural resources. Development and Policy Review, 19(2), 181–204.
Galluzzi, G., Halewood, M., Noriega, I. L., & Vernooy, R. (2016). Twenty-five years of international exchanges of plant genetic resources facilitated by the CGIAR genebanks: A case study on global interdependence. Biodiversity and Conservation, 25, 1421–1446.
Garming, H., Van de Houwe, I., & Roux, N. (2010). The impact of the Musa international transit Centre – Review of its services and cost-effectiveness, and recommendations for rationalization of its operations. Montpellier: Bioversity International.
Gollin, D., & Evenson, R. E. (1998). An application of hedonic pricing methods to value rice genetic resources in India. In R. E. Evenson, D. Gollin, & V. Santaniello (Eds.), Agricultural values of plant genetic resources (pp. 139–150). Wallingford: CAB International.
Gollin, D., Smale, M., & Skovmand, B. (2000). Searching an ex situ collection of wheat genetic resources. American Journal of Agricultural Economics, 82(4), 812–827.
Gotor, E., & Caracciolo, F. (2010). An empirical assessment of the effects of the 1994 in trust agreements on IRRI germplasm acquisition and distribution. International Journal of the Commons, 4(1), 437–451.
Halewood, M., Chiurugwi, T., Hamilton, R. S., Kurtz, B., Marden, E., Welch, E., et al. (2018). Plant genetic resources for food and agriculture: Opportunities and challenges emerging from the science and information technology revolution. New Phytologist, 217, 1407–1419.
Harlan, J. R. (1972). Genetics of disaster. Journal of Environmental Quality, 1(3), 212–215.
Harlan, J. R. (1992). Crops and man. Madison: American Society of Agronomy, Inc. and Crop Science Society of America, Inc..
Hein, L., & Gatzweiler, H. (2006). The economic value of coffee genetic resources. Ecological Economics, 60, 176–185.
Horna, D., Debouck, D., Dumet, D., Hanson, J., Payne, T., Sackville-Hamilton, R., et al. (2010). Evaluating cost-effectiveness of collection management: Ex-situ conservation of plant genetic resources in the CG system. Final report. Collective action for rehabilitation of global public goods CGIAR genetic resources systems: Phase 2 (GPG2).
ITPGRFA. (2020). Germplasm flow. Rome: FAO https://mls.planttreaty.org/itt/index.php?r=stats/pubStats. Accessed 6 January 2020.
Johnson, N. L., Pachico, D., & Voysest, O. (2003). The distribution of benefits from public international germplasm banks: The case of beans in Latin America. Agricultural Economics, 29, 277–286.
Kingwell, R., Godden, D., Kambuou, R., & Jackson, G. (2001). Managing and funding germplasm preservation in Papua New Guinea — For improved indigenous foods. Food Policy, 26(3), 265–280.
Koo, B., & Wright, B. D. (2000). The optimal timing of evaluation of genebank accessions and the effects of biotechnology. American Journal of Agricultural Economics, 82(4), 797–811.
Koo, B., Pardey, P. G., & Wright, B. D. (2004). Saving seeds: The economics of conserving crop genetic resources ex situ in the future harvest Centres of the CGIAR. Wallingford: CABI Publishing.
McCouch, S., McNally, K. L., Wang, W., & Sackville-Hamilton, R. (2012). Genomics of gene banks: A case study of rice. American Journal of Botany, 99(2), 1–17.
National Research Council (NRC). (1972). Genetic vulnerability of major crops. Washington, DC: National Academy of Sciences.
Plucknett, D. L., Smith, N. J. H., Williams, J. T., & Anishetty, N. M. (1987). Gene banks and the world’s food. Princeton: Princeton University Press.
Popova, E. (2018). Special issue on agricultural genebanks. Biopreservation and Biobanking, 16, 325–326. https://doi.org/10.1089/bio.2018.29044.ejp.
Prescott-Allen, C., & Prescott-Allen, R. (1986). The first resource: Wild species in the north American economy. New Haven: Yale University Press.
Raeburn, P. (1995). The last harvest: The genetic gamble that threatens to destroy American agriculture. New York: Simon and Schuster.
Raitzer, D. A., & Kelley, T. G. (2008). Benefit-cost meta-analysis of investment in the international agricultural research centers of the CGIAR. Agricultural Systems, 96, 108–123.
Renkow, M., & Byerlee, D. (2010). The impacts of CGIAR research: A review of recent evidence. Food Policy, 35(5), 391–402.
Robinson, J. & Srinivasan, C. S. (2013). Case studies on the impact of germplasm collection, conservation, characterization and evaluation in the CGIAR. CGIAR Standing Panel on Impact Assessment.
Schreinemachers, P., Ebert, A. W., & Wu, M.-H. (2014). Costing the ex situ conservation of plant genetic resources at AVRDC-the world vegetable center. Genetic Resources and Crop Evolution, 61, 757–773.
Simpson, R. D., Sedjo, R. A., & Reid, J. W. (1996). Valuing biodiversity for use in pharmaceutical research. Journal of Political Economy, 104, 163–185.
Smale, M., & Day-Rubenstein, K. (2002). The demand for crop genetic resources: International use of the US National Plant Germplasm System. World Development, 30, 1639–1655.
Smale, M., Hartell, J., Heisey, P., & Senauer, B. (1998). The contribution of genetic resources and diversity to wheat productivity and stability in the Punjab of Pakistan. American Journal of Agricultural Economics, 80, 482–493.
Smale, M., Singh, J., Di Falco, S., & Zambrano, P. (2008). Wheat breeding, productivity and slow variety change: Evidence from the Punjab of India after the green revolution. Australian Journal of Agricultural and Resource Economics, 6(52), 419–432.
Tyack, N., & Dempewolf, H. (2015). The economics of crop wild relatives under climate change. In R. J. Redden, S. S. Yadav, N. Maxted, M. E. Dulloo, L. Guarino, & P. Smith (Eds.), Crop wild relatives and climate change (pp. 281–291). New Jersey: Wiley-Blackwell.
Tyack, N., & Ščasný, M. (2018). Social valuation of genebank activities: Assessing public demand for genetic resource conservation in the Czech Republic. Sustainability., 10. https://doi.org/10.3390/su10113997.
Virchow, D. (2003). Efficient conservation of crop genetic diversity. Theoretical approaches and empirical studies. Berlin: Springer Verlag.
Widawsky, D., & Rozelle, S. (1998). Varietal diversity and yield variability in Chinese rice production. In M. Smale (Ed.), Farmers, gene banks, and crop breeding (pp. 159–172). Boston: Kluwer.
Wilson, E. O. (1988). Biodiversity. Washington, DC: The National Academies Press. https://doi.org/10.17226/989. Accessed 1 December 2018.
Wright, B. D. (1997). Crop genetic resource policy: The role of ex situ genebanks. The Australian Journal of Agricultural Economics, 41(1), 81–115.
Xepapadeas, A., Ralli, P., Kougea, E., Spyrou, S., Stavropoulos, N., Tsiaousi, V., & Tsivelikas, A. (2014). Valuing insurance services emerging from a gene bank: The case of the Greek gene bank. Ecological Economics, 97, 140–149.
Zohrabian, A., Traxler, G., Caudill, S., & Smale, M. (2003). Valuing pre-commercial genetic resources: A maximum entropy approach. American Journal of Agricultural Economics, 85, 429–436.
Acknowledgements
Funding for this research was provided by the CGIAR Genebank Platform and the Crop Trust.
Authorship contribution
Both authors contributed to the research conceptualization and design, writing, and editing.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The second author is an agricultural economist at the Crop Trust. The first author declares no conflict of interest.
Additional information
The original online version of this article was revised due to a retrospective Open Access order.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Smale, M., Jamora, N. Valuing genebanks. Food Sec. 12, 905–918 (2020). https://doi.org/10.1007/s12571-020-01034-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12571-020-01034-x