
Abstract
We report the first records of the leptosomiform taxon Plesiocathartes from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK). Two species could be distinguished, one of which is described as Plesiocathartes insolitipes, sp. nov. (the other is classified as Plesiocathartes sp.). We furthermore describe two small leptosomiform-like birds from Walton-on-the-Naze, Waltonavis paraleptosomus, gen. et sp. nov. and W. danielsi, gen. et sp. nov., which are represented by partial skeletons of multiple individuals. In plesiomorphic characters of the coracoid and furcula, the new taxon Waltonavis is distinguished from Plesiocathartes and the extant Leptosomus; the skull of Waltonavis furthermore exhibits basipterygoid processes, which are absent in Leptosomus. Another new species, Lutavis platypelvis, gen. et sp. nov., is based on a partial skeleton from Walton-on-the-Naze and closely resembles Plesiocathartes in the overall shape of the tarsometatarsus. However, L. platypelvis is clearly distinguished from leptosomiform birds in the morphology of the hypotarsus, the more massive coracoid, and other skeletal features. Although we consider close affinities to Plesiocathartes and, hence, the Leptosomiformes to be possible, we could not identify derived characters that enable an unambiguous phylogenetic placement of L. platypelvis. In any case, the new species document an unexpected diversity of Plesiocathartes-like fossil in the London Clay and may open a new perspective for a reconstruction of the ancestral morphology of the avian clade Afroaves, which includes the Leptosomiformes and other arboreal landbirds.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Telluraves is the clade that includes most of the small arboreal neornithine birds (Sangster et al. 2022). This species-rich group comprises morphologically and ecologically disparate species, from large raptors (many Accipitridae) and long-legged ground-dwellers (Cariamidae, Sagittariidae) to small perching birds. Molecular analyses support a division of Telluraves into two clades termed Australaves and Afroaves (Sangster et al. 2022). The former includes seriemas (Cariamiformes), falcons (Falconiformes), parrots (Psittaciformes), and passerines (Passeriformes), whereas the latter comprises accipitriform diurnal birds of prey, owls (Strigiformes), coraciiform and piciform birds (rollers, woodpeckers, and allies), as well as the enigmatic courol or cuckoo-roller, Leptosomus discolor (Leptosomiformes). This latter species was long allied with true rollers (Coraciiformes), but in current sequence-based analyses L. discolor is the sister group of a clade including trogoniform, coraciiform, and piciform birds (Ericson et al. 2006; Hackett et al. 2008; Prum et al. 2015; Kuhl et al. 2021), and close affinities to coraciiform birds were also rejected on the basis of morphological data (Mayr 1998).
The distribution of L. discolor is confined to Madagascar and the Comoro Islands, but there exist various fossils of stem group Leptosomiformes from Eocene sites in Europe and North America, which show that the extant species is a relict of a once much more widely distributed group of birds (Mayr 2017, 2022a). All named species of stem group Leptosomiformes were assigned to the taxon Plesiocathartes, which was initially erected for P. europaeus, a species based on a tarsometatarsus from an unknown stratigraphic level of the middle Eocene to late Oligocene Quercy fissure fillings in France (Gaillard 1908; Mourer-Chauviré 2002). Before leptosomiform affinities of Plesiocathartes were proposed (Mayr 2002a), P. europaeus was long considered to be an Old World representative of the Cathartidae (New World vultures; Gaillard 1908; Cracraft and Rich 1972).
Except for a tentative record of an unnamed species from the late Paleocene of France (Mayr and Smith 2019a), all stratigraphically well-constrained Plesiocathartes fossils are from early or middle Eocene deposits. Four species have been named: P. kelleri from the latest early or earliest middle Eocene of the German fossil site Messel (Mayr 2002a), P. geiselensis from the middle Eocene of the Geisel Valley in Germany (Mayr 2002b, 2020), as well as P. wyomingensis and P. major from the early Eocene North American Green River Formation (Weidig 2006). Plesiocathartes-like tarsometatarsi also exist from the early Eocene of Belgium and India (Mayr and Smith 2019b; Mayr et al. 2010), and a Plesiocathartes-like distal tarsometatarsus was furthermore reported from the London Clay of the Isle of Sheppey (Essex, UK; Mayr 2002b).
The species of Plesiocathartes so closely resemble the extant courol in their skeletal morphology that it was concluded that the extant species qualifies as a “living fossil” (Mayr 2008). However, most fossil specimens of Plesiocathartes are either compressed skeletons, which allow the recognition of only a few osteological details, or they consist of a few isolated bones, mainly tarsometatarsi.
Here we describe fossils of Plesiocathartes from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK), which were collected by the late Michael Daniels and belong to two species, at least one of which is new (these fossils were listed as New World vultures by Feduccia 1999: tab. 4.1). In addition, we report two new small species from Walton-on-the-Naze, which show a close resemblance to the Leptosomiformes and are represented by three-dimensionally preserved bones of all major skeletal elements from multiple individuals. These new species are distinct from Plesiocathartes and the extant courol in some skeletal features and, compared with other early Eocene birds, they show the closest resemblance to Lapillavis incubarens from Messel, for which possible affinities to the Leptosomiformes were considered in the original description (Mayr 2016). We furthermore report an osteologically more aberrant taxon from Walton-on-the-Naze, which resembles Plesiocathartes in tarsometatarsus morphology and may be closely related to leptosomiform birds.
Material and methods
The fossils are deposited in the Geiseltalsammlung, Martin-Luther Universität of Halle-Wittenberg, Germany (GMH), the Royal Belgian Institute of Natural Sciences, Brussels, Belgium (IRSNB), the Natural History Museum, London, UK (NHMUK), the National Museums Scotland, Edinburgh, UK (NMS), the Senckenberg Research Institute Frankfurt, Germany (SMF), and the Wyoming Dinosaur Center, Thermopolis, USA (WDC).
Ancestral character states were reconstructed under the parsimony criterion with Mesquite 2.71 (Maddison and Maddison 2009), based on a tree topology resulting from recent analyses of molecular data (Prum et al. 2015, Kuhl et al. 2021).
Systematic palaeontology
Aves Linnaeus, 1758
Afroaves Ericson, 2012
Leptosomiformes Sharpe, 1891
Plesiocathartes Gaillard, 1908
Plesiocathartes sp.
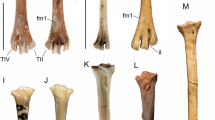
Referred specimens: NMS.Z.2021.40.34 (Fig. 1a–f; both tarsometatarsi and several pedal phalanges), collected in 1998 by Michael Daniels (original collector’s number WN 98977). NMS.Z.2021.40.35 (right tarsometatarsus), collected in 1982 by M. Daniels (Fig. 1g–j; original collector’s number WN 82409).

Plesiocathartes sp. from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK. a–f Specimen NMS.Z.2021.40.34: a–c Right tarsometatarsus in dorsal (a), plantar (b), and distal (c) view. d, e Proximal end of left tarsometatarsus in plantar (d) and proximal (e) view. f Pedal phalanges; the toes are numbered (of two phalanges, left and right ones are preserved). g–j Specimen NMS.Z.2021.40.35: right tarsometatarsus in dorsal (g), plantar (h), distal (i), and proximal (j) view. The scale bars equal 10 mm
Locality and horizon: Walton-on-the-Naze, Essex, United Kingdom; Walton Member of the London Clay Formation (previously Division A2; Rayner et al. 2009, Aldiss 2012); early Eocene (early Ypresian, 54.6–55 million years ago; Collinson et al. 2016).
Measurements (maximum length, in mm): NMS.Z.2021.40.34: Tarsometatarsus, 25.6 (left), 26.8 (right). NMS.Z.2021.40.35: Tarsometatarsus, 27.0 (right).
Comments: In size and morphology, the above tarsometatarsi from Walton-on-the-Naze correspond well to Plesiocathartes wyomingensis from the early Eocene North American Green River Formation (Weidig 2006). The exact stratigraphic age of the P. wyomingensis holotype is uncertain, but the fossil is likely to be a few million years younger than the specimens from Walton-on-the-Naze. The fossils furthermore are from different continents, so that it is unlikely that they belong to the same species.
NMS.Z.2021.40.34 is associated with a very poorly preserved partial skeleton that may belong to the same species (in this fossil, another left tarsometatarsus is preserved, which is broken into two pieces). As in P. wyomingensis the carpometacarpus of this latter fossil is longer than the tarsometatarsus, whereas both bones are of subequal length in P. kelleri (the carpometacarpus of other Plesiocathartes species is unknown).
As detailed in the descriptions below, the Plesiocathartes fossils from Walton-on-the-Naze are distinguished in their tarsometatarsus morphology from a specimen of an unnamed Plesiocathartes species from the London Clay of the Isle of Sheppey, which was described by Mayr (2002b). With the strata exposed at Walton-on-the Naze being from division A2 of the London Clay and the Plesiocathartes specimen from the Isle of Sheppey stemming from the stratigraphically younger divisions D–E (Rayner et al. 2009; Collinson et al. 2016), these differences possibly indicate temporal changes in the composition of the London Clay faunas.
Description and comparisons: In its size, proportions, and overall morphology, the tarsometatarsus corresponds to that of P. wyomingensis, but close comparisons are impeded by the poor preservation of this bone in the holotype and only known specimen of this species (Weidig 2006). The bone is also similar to a tarsometatarsus from the early Eocene of Egem in Belgium, which was referred to Plesiocathartes sp. by Mayr and Smith (2019a). By contrast, it is distinguished from the tarsometatarsus of P. europaeus in that the shaft is proportionally wider and the trochlea metatarsi IV reaches less far distally (only to the middle of the trochlea metatarsi III, whereas it extends beyond the middle of the tr. mt. III in P. europaeus). The tarsometatarsus of P. geiselensis has a narrower shaft, the trochlea metatarsi II is proportionally longer, and the proximal margin of the plantar articular surface of the trochlea metatarsi III is not tapering. The hypotarsus resembles that of P. geiselensis and, as in this species, it lacks bony canals and exhibits two very shallow sulci for the tendons of musculus flexor hallucis longus and m. flexor digitorum longus. Compared to a tarsometatarsus from the Isle of Sheppey, which was referred to Plesiocathartes by Mayr (2002b), the trochlea metatarsi III has a less pronounced trochlear furrow and the tr. mt. IV likewise lacks a pronounced plantar furrow; in distal view the trochlea metatarsi IV has a straight lateral margin, whereas this margin is slightly convex in the fossil from the Isle of Sheppey (Fig. 2n–p). The plantar articular surface of the trochlea metatarsi III tapers to a point. The trochlea metatarsi IV forms a narrow, rim-like plantar flange.

Comparison of the coracoid (a–f) and tarsometatarsus (g–kk) of Plesiocathartes and other fossil and extant taxa. a, b Right coracoid of Plesiocathartes insolitipes, sp. nov. from Walton-on-the-Naze in ventral (a) and dorsal (b) view (holotype, NMS.Z.2021.40.36). c Left coracoid of Plesiocathartes major from the North American Green River Formation in dorsal view (holotype, WDC 2001-CGR-022; coated with ammonium chloride). d Lutavis platypelvis, gen. et sp. nov. (holotype, NMS.Z.2021.40.37) reconstruction of coracoid (left side, dorsal view) by superimposing mirrored sternal portion of right coracoid on left coracoid; the dotted line indicates the reconstructed shape of the processus acrocoracoideus. e Left coracoid of Waltonavis paraleptosomus, gen. et sp. nov. in dorsal view (NMS.Z.2021.40.17). f Left coracoid of Leptosomus discolor (Leptosomiformes; SMF 17388) in dorsal view. g–j Right tarsometatarsus of Plesiocathartes sp. (NMS.Z.2021.40.35) in dorsal (g), plantar (h), distal (i), and proximal (j) view. k–m Distal end of right tarsometatarsus of P. insolitipes, sp. nov. (holotype, NMS.Z.2021.40.36) in dorsal (k), plantar (l), and distal (m) view. n–p Distal end of left tarsometatarsus of Plesiocathartes sp. from the early Eocene of the Isle of Sheppey, UK (NHMUK A 6178) in dorsal (n), plantar (o), and distal (p) view. q–t Right tarsometatarsus of L. platypelvis (holotype, NMS.Z.2021.40.37) in dorsal (q), plantar (r), distal (s), and proximal (t) view. u–x Left tarsometatarsus of W. paraleptosomus (NMS.Z.2021.40.17) in dorsal (u), plantar (v), distal (w), and proximal (x) view. y, z Right tarsometatarsus of Plesiocathartes geiselensis from the middle Eocene of the Geisel Valley, Germany (holotype, GMH XXXV- 559) in dorsal (y) and plantar (z) view. aa Left tarsometatarsus of P. major in plantar view (holotype, WDC 2001-CGR-022; coated with ammonium chloride). bb, cc Left tarsometatarsus of Plesiocathartes sp. from the early Eocene of Egem in Belgium (IRSNB Av 185) in dorsal (bb) and plantar (cc) view (coated with ammonium chloride). dd–gg Right tarsometatarsus of L. discolor (SMF 5438) in dorsal (dd), plantar (ee), distal (ff), and proximal (gg) view (dd and ee are mirrored images of the left tarsometatarsus). hh–kk Right tarsometatarsus of Cathartes aura (Cathartidae; SMF 5708) in dorsal (hh), plantar (ii), distal (jj), and proximal (kk) view. cpr crista procoracoidei, fdl hypotarsal sulcus/canal for tendon of musculus flexor digitorum longus, fhl hypotarsal sulcus/canal for tendon of musculus flexor hallucis longus, flg wing-like plantar flange. The scale bars equal 10 mm
NMS.Z.2021.40.34 includes eleven non-ungual pedal phalanges (Fig. 1f), which are of similar proportions to those of P. wyomingensis and P. kelleri. Three further phalanges associated with the fossil are too small to belong to the same species and must therefore be from a different bird (an association of remains from different species is not uncommon in the fossiliferous strata of Walton-on-the-Naze).
Plesiocathartes insolitipes, sp. nov.
Holotype: NMS.Z.2021.40.36 (Fig. 3a; partial skeleton, including right quadrate, several vertebrae, incomplete right coracoid, cranial extremity of left scapula, partial furcula, cranial portion of sternum, proximal end of right carpometacarpus, distal right tarsometatarsus and pedal phalanges of right foot), collected in 1993 by Michael Daniels (original collector’s number WN 93783).

Plesiocathartes insolitipes, sp. nov. from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK, overview of the two specimens. a Holotype, NMS.Z.2021.40.36 (the toes are numbered). b Referred specimen SMF Av 656. acr aromion, cdd condylus dorsalis, fns foramen nervi supracoracoidei, tbc tuberculum carpale, pii processus internus indicis. The scale bars equal 10 mm
Differential diagnosis: Distinguished from:
-
Plesiocathartes kelleri, P. major, and P. wyomingensis in the markedly widened distal ends of the phalanges of the third and fourth digits. Differs from Plesiocathartes sp. from Walton-on-the-Naze in its larger size (distal width of tarsometatarsus 6.2 mm in NMS.Z.2021.40.34 versus 7.2 mm in NMS.Z.2021.40.36) and in that the trochlea metatarsi IV has a better developed and medially inclined plantar flange; trochlea metatarsi II not forming a plantar projection. Also differs from P. kelleri in its larger size (Tab. 1) and from P. major in that the coracoid has the sternal margin oriented more obliquely to the longitudinal axis of the bone and the first phalanx of the hallux is proportionally longer (slightly longer than the first phalanx of the third toe, whereas it is shorter in P. major).
-
P. geiselensis in its smaller size (mediolateral width of trochlea metatarsi III 2.5 mm vs. 3.1 mm) and in that the tarsometatarsus shaft is proportionally wider and the dorsal articular surface of the trochlea metatarsi III is proportionally longer (reaching proximally to the level of the distal rim of the foramen vasculare distale).
-
P. europaeus in that the plantar articular surface of the trochlea metatarsi III is proportionally shorter and wider, with a straight proximal margin and a shallower trochlear furrow.
Etymology: From insolitus (Lat.): unusual and pes (Lat.): foot, in reference to the widened distal ends of some of the pedal phalanges.
Type locality and horizon: Walton-on-the-Naze, Essex, United Kingdom; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Referred specimens: SMF Av 656 (Fig. 3b; partial skeleton, including several vertebrae, right coracoid, partial right scapula, partial furcula, distal portion of right ulna, proximal end of right carpometacarpus, right os carpi radiale, partial phalanx digiti majoris), collected in 1987 by Paul Bergdahl (original collector’s number BC 8712).
Measurements (maximum length, in mm): NMS.Z.2021.40.36: Coracoid, maximum length as preserved, 30.7. SMF Av 656: Coracoid, maximum length as preserved, 30.3.
Description and comparisons: The quadrate (Fig. 4a–e) resembles that of Leptosomus discolor (Fig. 4f–j) in its shape and most morphological details. The condylus medialis is ventrally prominent. The processus oticus has a broad tip with widely separated capitula, and there is a tuberculum subcapitulare ventral to the capitulum squamosum. As in Leptosomus there is a large pneumatic foramen on the medial surface at the base of the processus orbitalis, which is situated more ventrally in the fossil. The condylus caudalis is somewhat wider mediolaterally than in Leptosomus.

Plesiocathartes insolitipes, sp. nov. from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK, anatomical details and comparison with the extant Leptosomus discolor. a–e Right quadrate of P. insolitipes (holotype, NMS.Z.2021.40.36) in lateral (a), medial (b), caudal (c), rostral (d), and ventral (e) view. f–j Quadrate of L. discolor (SMF 5438; left side, mirrored) in lateral (f), medial (g), caudal (h), rostral (i), and ventral (j) view. k ?Left pterygoid P. insolitipes (holotype, NMS.Z.2021.40.36). l Left pterygoid of L. discolor in ventral view (SMF 5438). m Partial atlas of P. insolitipes (holotype, NMS.Z.2021.40.36). n Atlas of L. discolor (SMF 17388). o, p Axis of P. insolitipes (holotype, NMS.Z.2021.40.36, left side) in dorsal (o) and lateral (p) view. q, r Axis of L. discolor (SMF 17388) in dorsal (q) and lateral (r) view. s Series of four thoracic vertebrae of P. insolitipes (holotype, NMS.Z.2021.40.36); the arrow indicates an enlarged detail of the corpus of one of the thoracic vertebrae. t Series of four thoracic vertebrae of L. discolor (SMF 17388; right side, mirrored). u, v Furcula of P. insolitipes (u; holotype, NMS.Z.2021.40.36) and L. discolor (v; SMF 17388) in caudal view. w–y Right coracoid of P. insolitipes (holotype, NMS.Z.2021.40.36) in ventral (w), medial (x), and dorsal (y) view. z–bb Right coracoid of P. insolitipes (SMF Av 656) in ventral (z), medial (aa), and dorsal (bb) view. cc–ee Cranial portion of sternum of P. insolitipes (holotype, NMS.Z.2021.40.36) in ventral (cc), dorsal (dd), and cranial (ee) view. ff, gg Left os carpi radiale of P. insolitipes (ff; holotype, NMS.Z.2021.40.36) and L. discolor (gg; SMF 5438). hh, ii Proximal portion of right carpometacarpus of P. insolitipes (holotype, NMS.Z.2021.40.36) in ventral (hh) and dorsal (ii) view. jj Proximal portion of right carpometacarpus of L. discolor (SMF 5438) in ventral view. kk P. insolitipes (holotype, NMS.Z.2021.40.36), first phalanx of third toe in plantar view. ll P. insolitipes (holotype, NMS.Z.2021.40.36), first phalanx of fourth toe in plantar view. apf apophysis furculae, blg bulges on distal end of pedal phalanx, cdc condylus caudalis, cdm condylus medialis, cpo capitulum oticum, cps capitulum squamosum, faa facies articularis acrocoracoidea, ftv foramen transversarium (incompletely closed), icf incisura fossae, pex processus extensorius, pis processus pisiformis, pnf pneumatic foramen, prj proximal projection, psp processus spinosus, sac sulcus articularis coracoideus, tbs tuberculum subcapitulare, zcd zygapophysis caudalis. The scale bars equal 10 mm
The pterygoid is shorter and stouter than that of Leptosomus discolor (Fig. 4k, l). As in the latter it lacks a facies articularis basipterygoidea.
The atlas (Fig. 4m) exhibits an incisura fossae. Unlike in Leptosomus discolor (Fig. 4n) it lacks (incompletely closed) foramina transversaria. The axis (Fig. 4o, p) is rostrocaudally longer than in Leptosomus; the processus spinosus is less dorsally protruding, the zygapophyses caudales are shorter and more caudally projecting (Fig. 4q, r). The holotype includes only one thoracic vertebra, but a sequence of four consecutive thoracic vertebrae is preserved in the referred specimen SMF Av 656 (Fig. 4s). Unlike in Leptosomus, but as in the Cathartidae (Mayr 2021), these thoracic vertebrae are pneumatized and exhibit pneumatic openings on the lateral surfaces of the corpus.
The coracoid (Fig. 4w-bb) differs from that of Leptosomus discolor (Fig. 2f) in that the crista procoracoidei is less pronounced, the processus acrocoracoideus is more medially extensive (the medial portion is flattened by the large facies articularis clavicularis in Leptosomus), and the facies articularis sternalis is proportionally longer.
The scapula is more complete in the referred specimen SMF Av 656 (Fig. 3b). The acromion is short and tapers into a small, pointed projection. The tuberculum coracoideum is only weakly developed.
The furcula (Fig. 4u) is broadly U-shaped, the scapi clavicularum widen towards the extremitas omalis. Unlike in Leptosomus (Fig. 4v) the sternal extremity is not reduced and bears a small apophysis furculae. The extremitas omalis is furthermore narrower than in Leptosomus and lacks a well-delimited facies articularis acrocoracoidea.
Only the cranial portion of the sternum is preserved in the holotype (Fig. 4cc–ee). As in Leptosomus a spina externa is absent. The sulci articulares coracoidei overlap in the midline of the bone.
Except for the proximodistally somewhat shorter condylus dorsalis, the distal end of the ulna (SMF Av 656) resembles that of L. discolor.
On the carpometacarpus (Fig. 4hh, ii), the proximal margin of the processus extensorius is concave, as it is in most birds, whereas it is straight in Leptosomus (Fig. 4jj). As in other Plesiocathartes species, of which the carpometacarpus is known, but unlike in Leptosomus, the os metacarpale minus does not exceed the os metacarpale majus in distal extent. The processus pisiformis is centrally positioned.
The phalanx proximalis digiti majoris bears a small processus internus indicis (Fig. 3a). The os carpi ulnare resembles that of Leptosomus in its shape, with the crus longum being longer than the crus breve. The os carpi radiale is proximodistally narrower than in Leptosomus (Fig. 4ff, gg); as in the latter, but unlike in coraciiform birds, it lacks a sulcus for the tendon of musculus extensor longus alulae (Mayr 2014).
The foramen vasculare distale appears to have been large, but only its distal rim is preserved in the fossil. The proximal margin of the plantar articular surface of the trochlea metatarsi III is broadly rounded. The lateral surface of the trochlea metatarsi IV forms a distinct bulge; the plantar articular surface of this trochlea exhibits a narrow, rim-like projection (in Leptosomus there is a marked flange; Fig. 2ff). The distal end of the bone differs from a Plesiocathartes-like tarsometatarsus from the Isle of Sheppey (Fig. 2n–p; Mayr 2002b) in that the trochlea metatarsi II is more medially protruding and does not form a plantar projection, the tr. mt. III is mediolaterally shorter and its plantar articular surface has a straight proximal margin, and the tr. mt. IV does not bear a marked plantar furrow.
The holotype includes four unguals and ten non-ungual pedal phalanges (Fig. 3a). All but two pairs of the non-ungual phalanges are of different sizes and likely to be from one foot (the right one). The first phalanx of the hallux is long and slender. Four phalanges are abbreviated and two of these have a similar shape and size. Because in other species of Plesiocathartes only the proximal three phalanges are abbreviated (Mayr 2002b; Weidig 2006), one of the duplicate phalanges probably belongs to the left foot. The morphology of the proximal articular surfaces also suggests that the first phalanx of the second toe is missing in the fossil, so that the set of non-ungual phalanges (of the right foot) is not completely preserved in the fossil. The distal portions of six of the phalanges (here identified as the proximal phalanges of the fourth and third digits) are strongly widened and exhibit marked bulges just proximal to the articular end (Fig. 4kk, ll), which give these phalanges an unusual, almost club-shaped outline. The medial portion of the proximal end of the first phalanx of the fourth toe forms a proximal projection (Fig. 4ll). The ungual phalanges have a laterally open sulcus neurovascularis and a weakly developed tuberculum extensorium.
?Leptosomiformes Sharpe, 1891
Waltonavis, gen. nov.
Type species: Waltonavis paraleptosomus, sp. nov.
Diagnosis: Characterised by (1) narrow and pointed upper beak; (2) pterygoid with prominent facies articularis basipterygoidea; (3) humerus with sharply delimited fossa musculi brachialis; (4) tarsometatarsus with very large medial foramen vasculare proximale (Fig. 5c); (5) hypotarsus enclosing two canals; (6) trochlea metatarsi II reaching much farther distally than trochlea metatarsi IV; (7) trochlea metatarsi IV mediolaterally narrow and forming a plantar rim; (8) distal ends of first and second phalanges of third toe bearing distinct lateral bulges. Characters (3), (4), and (8) are here considered to be autapomorphies of the new taxon.

Overview of selected skeletal elements of Waltonavis specimens from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK; all fossils shown to scale. a Holotype of Waltonavis paraleptosomus, gen. et sp. nov. (NMS.Z.2021.40.16; pterygoid, quadrate, coracoid, scapula, sternum, and tarsometatarsi). b Specimen tentatively referred to W. paraleptosomus (NMS.Z.2021.40.19; quadrate, pygostyle, scapula, coracoids, humerus, carpometacarpus, tarsometatarsus, and pedal phalanges). c W. paraleptosomus (NMS.Z.2021.40.17; coracoid, proximal carpometacarpus, tarsometatarsus, and pedal phalanges). d W. paraleptosomus (NMS.Z.2021.40.18; right tarsometatarsus of a juvenile individual). e Holotype of Waltonavis danielsi, gen. et sp. nov. (NMS.Z.2021.40.20; atlas, pygostyle, coracoid, scapula, humeri, radius, right ulna, proximal and distal left ulna, carpometacarpus, os carpi ulnare, proximal tibiotarsus, and tarsometatarsus). f–h Specimens identified as Waltonavis sp. (f NMS.Z.2021.40.21; partial, beak, quadrate, both humeri, coracoid, furcula, os carpi ulnare, partial ulna, tibiotarsus, and tarsometatarsus; g NMS.Z.2021.40.22; both coracoids, cranial end of right scapula, proximal end of left humerus; h NMS.Z.2021.40.23; proximal and distal ends of right tarsometatarsus, atlas, basiurohyal, and some pedal phalanges). mfv foramen vasculare proximale. Scale bars: 10 mm
Differential diagnosis: Distinguished from the late early/early middle Eocene Lapillavis from the Messel fossil site (Mayr 2016) in that the furcula has much narrower shafts (scapi clavicularum) and the foramen nervi supracoracoidei is situated closer to omal extremity of coracoid (Fig. 6). Apart from its much smaller size (Tab. 1), the new taxon differs from Plesiocathartes in that the coracoid has a narrower shaft (Fig. 2c, e), the medial foramen vasculare proximale of the tarsometatarsus is larger, the trochlea metatarsi II reaches much farther distally than the trochlea metatarsi IV and forms a more medially directed and much bulkier plantar projection, the trochlea metatarsi IV is proportionally narrower, and the hallux is proportionally longer (distinctly exceeding the other phalanges in length and measuring almost half the length of tarsometatarsus).

Comparison of the coracoid (a–c) and furcula (d, e) of Waltonavis, gen. nov. from the early Eocene London Clay of Walton-on-the-Naze, Essex (UK) and Lapillavis incubarens from the late early or earliest middle Eocene of Messel, Germany. a Left coracoid of W. paraleptosomus, gen. et sp. nov. (NMS.Z.2021.40.17). b, c Left coracoid of L. incubarens (holotype, SMF-ME 1066; coated with ammonium chloride); in c, the surrounding matrix was digitally brightened. d Furcula of Waltonavis sp. (NMS.Z.2021.40.21) in caudal view. e Furcula of L. incubarens (holotype, SMF-ME 1066; coated with ammonium chloride) in caudal view. apf apophysis furculae, fns foramen nervi supracoracoidei. Scale bars: 10 mm
Etymology: The name is derived from the type locality of the new taxon and avis (Lat.): bird.
Description and comparison: Because the two species assigned to Waltonavis mainly differ in the length proportions of some pectoral girdle and limb bones, a description is provided for the taxon Waltonavis rather than for each species.
Judging from the portion preserved in NMS.Z.2021.40.21 (Fig. 7a), the beak of Waltonavis had thrush-like proportions and was narrower than that of Leptosomus. The nostrils are long and take up about half of the beak length. The pterygoid (Fig. 7i, j) exhibits a distinct facies articularis basipterygoidea, which indicates the presence of basipterygoid processes in the basicranial area of the skull (absent in crown group Leptosomiformes).

Cranial elements and vertebrae of Waltonavis, gen. nov. from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK. a Upper beak in dorsal view (Waltonavis sp.; NMS.Z.2021.40.21). b, c Right quadrate (Waltonavis sp.; NMS.Z.2021.40.21) in lateral (b) and ventral (c) view. d Right quadrate (Waltonavis paraleptosomus, gen. et sp. nov.; NMS.Z.2021.40.19) in caudolateral view. e, f Left quadrate (W. paraleptosomus; NMS.Z.2021.40.16) in medial (e) and lateral (f) view; the dotted lines indicate the reconstructed shape of the damaged processus orbitalis. g, h Left quadrate of the extant Leptosomus discolor (SMF 5438) in lateral (g) and caudal (h) view. i, j Left pterygoid (W. paraleptosomus; NMS.Z.2021.40.16) in medial (i) and dorsal (j) view. k Basiurohyal bone of the hyoid apparatus (W. paraleptosomus; NMS.Z.2021.40.36). cdm condylus medialis, cdp condylus pterygoideus, cpo capitulum oticum, cps capitulum squamosum, cqj cotyla quadratojugalis, fab facies articularis basipterygoidea, orb processus orbitalis, tsc tuberculum subcapitulare, vrm ventral rim of condylus lateralis. Scale bars: 5 mm
The quadrate (Fig. 7b–f) is preserved in three specimens. The processus oticus has a mediolaterally broad tip, with the capitulum oticum and the capitulum squamosum being separated by a shallow incisura intercapitularis; the bone lacks pneumatic openings in the dorsal portion of its caudal surface. Situated ventral to the capitulum squamosum, there is a small tuberculum subcapitulare sensu Elzanowski and Stidham (2010). The short condylus lateralis lacks the broad ventral rim found in Leptosomus (Fig. 7g). The condylus medialis is smaller and more caudally located than in Leptosomus and most other extant birds, with most of this condyle being situated caudal rather than ventral to the processus oticus. The processus orbitalis is moderately long and has a fairly broad tip. In NMS.Z.2021.40.23, the basiurohyal of the hyoid apparatus is preserved, which is a rod-shaped element (Fig. 7k).
Two specimens, NMS.Z.2021.40.20 and NMS.Z.2021.40.21, include a larger number of vertebrae (Fig. 8). As in Leptosomus, the atlas (Fig. 8n) has an incisura fossae. The axis (Fig. 8a) is craniocaudally longer than the axis of Leptosomus, with a long dens and a prominent processus spinosus. The third vertebra exhibits a pair of small foramina (Fig. 8b, j); these foramina are much larger in Leptosomus. The other cervical vertebrae (Fig. 8c–h, k) have a rather unspecific morphology. The corpus of the thoracic vertebrae exhibits shallow fossae (pleurocoels; Fig. 8i, l), which are absent in Leptosomus; the occurrence of pleurocoels in early Paleogene stem group representatives is a recurrent theme in the evolution of neornithine birds and is functionally correlated with the absence of pneumatic foramina in the fossa pneumotricipitalis of the humerus (Mayr 2021). The pygostyle (Fig. 5b, e) is small.

Vertebrae of Waltonavis, gen. nov. from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK. a Axis in dorsal, cranial, and lateral view (Waltonavis sp.; NMS.Z.2021.40.21). b Third cervical vertebra in dorsal and lateral view (Waltonavis sp.; NMS.Z.2021.40.21). c–h Six cervical vertebrae in dorsal and ventral or ventrolateral view (Waltonavis sp.; NMS.Z.2021.40.21). i Thoracic vertebra in lateral view (Waltonavis sp.; NMS.Z.2021.40.21). j Third cervical vertebra in dorsal view (Waltonavis danielsi, gen. et sp. nov.; NMS.Z.2021.40.20). k Four cervical vertebrae in different views (Waltonavis danielsi; NMS.Z.2021.40.20). l Two thoracic vertebrae in lateral view (Waltonavis danielsi; NMS.Z.2021.40.20). m Five caudal vertebrae in different views (Waltonavis danielsi; NMS.Z.2021.40.20). n Atlas (Waltonavis sp.; NMS.Z.2021.40.23). dns dens, for foramen, fos fossa (pleurocoel), psp processus spinosus. Scale bar: 5 mm
The omal extremity of the coracoid (Fig. 9a–c) is smaller than in Leptosomus, and in its proportions the coracoid shows a resemblance to that of Lapillavis incubarens from Messel (Fig. 6b, c). The facies articularis clavicularis forms a lip that overhangs the sulcus supracoracoideus, so that the processus acrocoracoideus has a hook-like shape. The bone has a narrower shaft than the coracoids of Plesiocathartes and Leptosomus, but as in the latter two taxa (and Lapillavis) a foramen nervi supracoracoidei is present. The cotyla scapularis forms a concave articular surface. The extremitas sternalis exhibits a medial projection, which is absent in Leptosomus, and the processus lateralis is deeper in sterno-omal direction

Major pectoral girdle and wing bones of Waltonavis, gen. nov. from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK. a Left coracoid (Waltonavis sp.; NMS.Z.2021.40.21) in dorsal view. b, c Left coracoid (Waltonavis paraleptosomus, gen. et sp. nov.; NMS.Z.2021.40.17) in dorsal (b) and ventral (c) view. d Partial right scapula (W. paraleptosomus; NMS.Z.2021.40.19) in lateral view. e Furcula (Waltonavis sp.; NMS.Z.2021.40.21) in cranial view. f Partial sternum in cranial view (W. paraleptosomus; NMS.Z.2021.40.16). g, h Right humerus (Waltonavis sp.; NMS.Z.2021.40.21) in cranial (g) and caudal (h) view; the arrow denotes an enlarged detail of the distal end of the bone. i Proximal end of left ulna (NMS.Z.2021.40.20) in cranioventral view. j Distal end of left ulna (Waltonavis sp.; NMS.Z.2021.40.21) in ventral view. k Right os carpi ulnare (Waltonavis sp.; NMS.Z.2021.40.21). l Right os carpi radiale (Waltonavis sp.; NMS.Z.2021.40.21); the arrow denotes an enlarged view of the bone. m Proximal end of left carpometacarpus (W. paraleptosomus; NMS.Z.2021.40.17) in ventral view. n Right carpometacarpus (W. paraleptosomus; NMS.Z.2021.40.19) in cranioventral view. o, p Left carpometacarpus (NMS.Z.2021.40.20) in dorsal (o) and ventral (p) view; the arrow denotes an enlarged detail of the proximal end of the bone. q Left phalanx proximalis digiti majoris (NMS.Z.2021.40.20) in ventral view. apf apophysis furculae, cdd condylus dorsalis, cdv condylus ventralis, ctd cotyla dorsalis, ctv cotyla ventralis, cvx convexity caudal to processus pisiformis, fmb fossa musculi brachialis, fns foramen nervi supracoracoidei, mpr medial projection, pis processus pisiformis, spe spina externa, tbc tuberculum carpale, tbd tuberculum dorsale, tsd tuberculum supracondylare dorsale, vpr ventral projection on proximal end of os metacarpale minus. The scale bars equal 10 mm
Except for the slightly bifurcated tip of the acromion, the preserved portions of the scapula (Fig. 9d) do not offer traits of potential taxonomic or phylogenetic significance.
The furcula (Fig. 6d, 9e) is U-shaped and has much narrower shafts (scapi clavicularum) than the furculae of Plesiocathartes and Leptosomus, in which the dorsal portion of the shafts are broadened and strap-like (Fig. 4u, v). The furcular shafts are likewise much wider in Lapillavis (Fig. 6e). The sternal extremity bears a small apophysis furculae. Unlike in Leptosomus the omal extremity does not exhibits a well-developed facies articularis acrocoracoidea.
Only the cranial portion of the sternum is preserved in specimen NMS.Z.2021.40.16 (Fig. 9f). The spina externa is poorly developed.
The humerus (Fig. 9g, h) resembles that of Lapillavis in its shape and has a proportionally larger proximal end than the humerus of Leptosomus. As in Lapillavis and Leptosomus the tuberculum dorsale is small. The crista deltopectoralis is short, but dorsally prominent, and has a convex margin. The fossa pneumotricipitalis lacks pneumatic openings. The distal end of the bone corresponds to the distal humerus of Leptosomus. As in the latter, the fossa musculi brachialis is sharply delimited. Also as in Leptosomus there is a small tuberculum supracondylare dorsale. The configuration of the condyli is similar to Leptosomus, but the condylus ventralis is less globular and has a flattened cranial surface.
The ulna (Fig. 9i, j) exceeds the humerus in length. The proximal end resembles the proximal ulna of Plesiocathartes geiselensis (Mayr 2002a, 2020) and, unlike in the extant Leptosomus, the cotyla dorsalis reaches much farther distally than the cotyla ventralis. The tuberculum carpale, on the distal end of the bone, is small.
The carpometacarpus (Fig. 9m–p) corresponds to that of Plesiocathartes in its proportions. On the proximal end of the bone there is a pronounced convexity immediately caudal to the processus pisiformis. The processus extensorius is less pronounced than in Leptosomus. The intermetacarpal space is fairly narrow. The proximal portion of the os metacarpale minus exhibits a ventral projection, which is less pronounced than in Leptosomus. Furthermore, unlike in Leptosomus, the os metacarpale minus does not project much farther distally than the os metacarpale majus.
The os carpi ulnare (Fig. 9k) has a similar shape to that of Leptosomus. As in the latter, the crus longum and the crus breve are of similar length, whereas the crus longum is proportionally longer in Lapillavis. The os carpi radiale (Fig. 9l) also resembles the corresponding ossicle of Leptosomus and does not exhibit the derived morphology found in the Bucerotes, Coraciiformes, and Piciformes, in which there is a marked sulcus for the tendon of the musculus extensor longus alulae (Mayr 2014). The phalanx proximalis digiti majoris (Fig. 9q) lacks a processus internus indicis.
Only fragments of the femur are preserved. The distal end of the bone (Fig. 10a) differs from Leptosomus in that the caudal articular surfaces of the condyles are proximodistally longer and the condylus lateralis is mediolaterally wider.

Leg bones of Waltonavis, gen. nov. from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK. a Distal end of right femur (Waltonavis danielsi, gen. et sp. nov.; NMS.Z.2021.40.20) in caudal view. b Fragmentary distal end of right tibiotarsus (W. danielsi; NMS.Z.2021.40.20) in cranial view. c Proximal end of right tibiotarsus (W. danielsi; NMS.Z.2021.40.20) in cranial view. d Partial right tibiotarsus (Waltonavis sp.; NMS.Z.2021.40.21) in cranial view; the arrow denotes an enlarged detail of the distal end of the bone. e, f Distal end of left tarsometatarsus (Waltonavis sp.; NMS.Z.2021.40.21) in dorsal (e) and plantar (f) view. g, h Left tarsometatarsus (W. paraleptosomus, gen. et sp. nov.; NMS.Z.2021.40.17) in plantar (g) and dorsal (h) view; the arrow denotes an enlarged detail of the proximal end of the bone. i, j Left tarsometatarsus (W. paraleptosomus; NMS.Z.2021.40.16) in dorsal (i) and plantar (j) view; the arrow denotes an enlarged detail of the distal end of the bone (the trochleae are numbered). k–m Right tarsometatarsus (W. paraleptosomus; NMS.Z.2021.40.19) in plantar (k), lateroplantar (l), and dorsal (m) view. n Left tarsometatarsus (W. danielsi; NMS.Z.2021.40.20) in dorsal view. o–t Distal (o, q, s) and proximal ends (p, r, t) of the tarsometatarsus (o, p W. paraleptosomus, NMS.Z.2021.40.16, left [distal] and right [proximal] side; q, r W. paraleptosomus, NMS.Z.2021.40.17, left side; s, t Waltonavis sp., NMS.Z.2021.40.23, right side); the hypotarsus in p is damaged, the tarsometatarsal trochleae are numbered. cdl condylus lateralis, cdm condylus medialis, cmd crista medialis hypotarsi, fdl hypotarsal canal for musculus flexor digitorum longus, fhl hypotarsal canal for musculus flexor hallucis longus, fmt fossa metatarsi I, fvd foramen vasculare distale, lfv lateral foramen vasculare proximale, mfv medial foramen vasculare proximale, pst pons supratendineus, sul hypotarsal sulcus for a superficial flexor tendon. The scale bars equal 10 mm
The tibiotarsus (Fig. 10b–d) is likewise not completely preserved in any of the specimens. The cristae cnemialis cranialis is not very prominent and conjoins the crista cnemialis lateralis to form a crista patellaris in the cranioproximal portion of the bone. The distal end resembles the distal tibiotarsus of Leptosomus. It has widely spaced condyli of similar height. The pons supratendineus is situated in the medial portion of the bone.
The tarsometatarsus (Fig. 10e–t) is a comparatively short bone, which is subequal to the carpometacarpus in length. The proximal end is mediolaterally wide and the shaft narrower than in Leptosomus. The hypotarsus encloses two canals, for the tendons of musculus flexor digitorum longus and m. flexor hallucis longus (Fig. 10r, t); the plantar surface bears a shallow sulcus for one of the superficial flexor tendons (Fig. 10g). There is a well-developed crista medialis hypotarsi. By contrast, in Plesiocathartes geiselensis there are no closed hypotarsal canals, but two distinct furrows (Mayr 2002a, 2020). The medial foramen vasculare proximale is very large (Fig. 10h). The tuberositas musculi tibialis cranialis is medially situated and has an ovate outline. The fossa metatarsi I is well developed. The trochlea metatarsi II forms a medioplantarly-directed projection; the trochlea reaches much farther distally than the trochlea metatarsi IV, but does not exceed the trochlea metatarsi III in length. The incisura intertrochlearis lateralis is wide. The trochlea metatarsi IV bears a weakly developed plantar rim, whereas this trochlea exhibits a well-developed wing-like flange in the semi-zygodactyl Leptosomus. The trochlea metatarsi IV is proportionally shorter and narrower than in Plesiocathartes and Leptosomus; unlike in the latter two taxa it does not bear a well-developed trochlear furrow. The tarsometatarsi are poorly preserved in the holotype and only known specimen of Lapillavis incubarens, but appear to have been stouter than in Waltonavis.
The hallux is long and has a wide proximal end. The phalanges of the third toe have an unusual morphology in that the distal ends of the first and second phalanges form distinct lateral bulges (Fig. 11a–c). In the extant L. discolor similar bulges are also present, but occur on both sides of the phalanges and are less pronounced (Fig. 11d, e). As in Plesiocathartes (Mayr 2002b, Weidig 2006) the proximal three phalanges of the fourth toes are shortened, whereas in Leptosomus only the first and second phalanges are abbreviated (Fig. 11d; in the Lapillavis incubarens holotype the pedal phalanges are very poorly preserved). The ungual phalanges are only moderately curved and have rather weakly developed flexor tubercles; their surfaces bear marked vascular furrows and the neurovascular sulcus is laterally open.

Waltonavis, gen. nov. from the early Eocene London Clay of Walton-on-the-Naze, Essex (UK), tarsometatarsus and pedal phalanges (the latter shown in different orientations) in comparison to the extant Leptosomus discolor. a Left foot of NMS.Z.2021.40.21 (Waltonavis sp.). b Right foot of NMS.Z.2021.40.19 (Waltonavis paraleptosomus, gen. et sp. nov.). c Left foot of NMS.Z.2021.40.17 (W. paraleptosomus). d, e Left foot of L. discolor (d SMF 5438, e SMF 17457). The toes are numbered; the arrows indicate the bulges on the proximal two phalanges of the third toe. The scale bars equal 10 mm
Waltonavis paraleptosomus, sp. nov.
Holotype: NMS.Z.2021.40.16 (Fig. 5a; partial skeleton, including left quadrate and pterygoid, partial left scapula, left coracoid, cranial portion of sternum, left and proximal right tarsometatarsus), collected in 1988 by M. Daniels (original collector’s number WN 88599).
Diagnosis: As for genus; the coracoid is about as long as the tarsometatarsus (Fig. 12).

Coracoids and tarsometatarsi of Waltonavis specimens from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK, to illustrate the different length proportions of the bones in W. paraleptosomus, gen. et sp. nov. and W. danielsi, gen. et sp. nov.; all specimens shown to scale. a Holotype of Waltonavis paraleptosomus, gen. et sp. nov. (NMS.Z.2021.40.16). b, c Specimens referred to W. paraleptosomus (b NMS.Z.2021.40.19; c NMS.Z.2021.40.17). d Holotype of W. danielsi, gen. et sp. nov. (NMS.Z.2021.40.20). The scale bars equal 10 mm
Etymology: The species epithet refers to the osteological similarity of the new species and the extant taxon Leptosomus.
Type locality and horizon: Walton-on-the-Naze, Essex, United Kingdom; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Referred specimens: NMS.Z.2021.40.17 (Fig. 5c; partial skeleton, including left coracoid, proximal end of left carpometacarpus, left tarsometatarsus, and several pedal phalanges), collected in 1996 by M. Daniels (original collector’s number WN 96909). NMS.Z.2021.40.18 (Fig. 5d; right tarsometatarsus of a juvenile individual), collected in 1983 by M. Daniels (original collector’s number WN 83432). NMS.Z.2021.40.19 (Fig. 5b; partial skeleton, including right quadrate, partial mandible in a block of matrix, pygostyle, both coracoids, partial right scapula, partial left humerus, fragments of a radius, right carpometacarpus, partial right femur in a block of matrix, right tarsometatarsus, and several pedal phalanges), collected in 1988 by M. Daniels (original collector’s number WN 88585).
Measurements (maximum length, in mm): NMS.Z.2021.40.16: coracoid (left), 21.6; tarsometatarsus (left), 21.3. NMS.Z.2021.40.17: coracoid (left), 20.0; tarsometatarsus (left), 21.7. NMS.Z.2021.40.19: coracoid (left), 21.7; carpometacarpus (right), 20.1; tarsometatarsus (right), >19.1.
Waltonavis danielsi, sp. nov.
Holotype: NMS.Z.2021.40.20 (Fig. 5e; partial skeleton, including several vertebrae, right coracoid, partial right scapula, right humerus, proximal left humerus, right ulna, proximal and distal ends of left ulna, left carpometacarpus, both phalanges proximales digitorum majores, distal end of right femur, proximal end and fragmentary distal end of right tibiotarsus, partial left tarsometatarsus), collected in 1985 by M. Daniels (original collector’s number WN 85511).
Diagnosis: As for genus, the coracoid is distinctly shorter than the tarsometatarsus, which is more elongate and with a proportionally narrower proximal end than the tarsometatarsus of W. paraleptosomus (Fig. 12).
Etymology: The species is named in honour of Michael Daniels, who collected the fossils described in the present study.
Type locality and horizon: Walton-on-the-Naze, Essex, United Kingdom; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm): NMS.Z.2021.40.20: coracoid (right), 19.4; humerus (right), 35.0; ulna (right), 42.0; carpometacarpus (left), 19.3; tarsometatarsus (left), 22.6.
Waltonavis sp.
Referred specimens: NMS.Z.2021.40.21 (Fig. 5f; partial skeleton, including partial upper beak, right quadrate, several vertebrae, left coracoid, furcula, cranial extremities of both scapulae, right humerus, partial left humerus, right ulna lacking proximal end, fragmentary distal portions of both carpometacarpi, fragmentary right femur, distal end of left tarsometatarsus, and several pedal phalanges), collected in 1981 by M. Daniels (original collector’s number WN 81372). NMS.Z.2021.40.22 (Fig. 5g; both coracoids, cranial end of right scapula, proximal end of left humerus), collected in 1983 by M. Daniels (original collector’s number WN 83434). NMS.Z.2021.40.23 (Fig. 5h; proximal and distal ends of right tarsometatarsus, atlas, basiurohyal, and some pedal phalanges), collected in 1980 by M. Daniels (original collector’s number WN 80364).
Locality and horizon: Walton-on-the-Naze, Essex, United Kingdom; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (maximum length, in mm): NMS.Z.2021.40.21: coracoid (left), 20.9; humerus (right), 35.8; tibiotarsus (right) length as preserved, 29.0. NMS.Z.2021.40.22: coracoid (left), 20.3, coracoid (right), 19.6.
Remarks: Because either the coracoid or the tarsometatarsus are missing or incomplete, these specimens cannot be assigned to one of the two species above.
?Afroaves Ericson, 2012
Lutavis, gen. nov.
Type species: Lutavis platypelvis, sp. nov.
Diagnosis: Characterised by a coracoid with a large extremitas omalis (measuring about one third of the entire length of the bone); pelvis wide and dorsoventrally shallow, with small tubercula praeacetabularia; tarsometatarsus with very large medial foramen vasculare proximale and hypotarsus forming two crests that delimit a distinct sulcus.
Differential diagnosis: Differs from Plesiocathartes europaeus, the type species of the taxon Plesiocathartes, in that the trochlea metatarsi IV forms a more pronounced plantar flange. Furthermore differs from Plesiocathartes spp. in the large medial foramen vasculare proximale (tarsometatarsus) and the morphology of the hypotarsus, which exhibits a distinct sulcus. Distinguished from Waltonavis, gen. nov. in that the coracoid is much more massive, the humerus has a concave proximodorsal margin and a dorsally prominent crista deltopectoralis, and the hypotarsus lacks tendinal canals (Fig. 2).
Etymology: The name is derived from lutum (Lat.): mud or clay and avis (Lat.): bird, in reference to the fact that the fossils were found in the London Clay
Lutavis platypelvis, gen. et sp. nov.
Holotype: NMS.Z.2021.40.37 (partial skeleton, including a few thoracic vertebrae and the pygostyle, substantial portions of both scapulae and coracoids, proximal and fragmentary distal end of the right humerus, proximal end of the left ulna, distal ends of both ulnae, partial carpometacarpi, carpal bones and wing phalanges, partial pelvis, right tarsometatarsus, and several pedal phalanges), collected in 1995 by Michael Daniels (original collector’s number WN 95848A).
Diagnosis: As for genus.
Etymology: From πλᾰτῠ́ς (platús; Gr.): wide, in reference to the wide and shallow pelvis of the new species.
Type locality and horizon: Walton-on-the-Naze, Essex, United Kingdom; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (in mm): Left coracoid, length as preserved, 23.9; right humerus, proximal width, 18.0; pelvis, length as preserved, 29.2; right carpometacarpus, length, 32.3; right tarsometatarsus, length, 29.8.
Description and comparisons: The thoracic vertebrae (Fig. 13b, c) exhibit small pneumatic foramina on the lateral surfaces of the corpus, which are less distinct than those in Plesiocathartes insolitipes (Fig. 4s). The pygostyle (Fig. 13a) is fairly large with a well-developed lamina pygostyli.

Lutavis platypelvis, gen. et sp. nov. from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK (holotype, NMS.Z.2021.40.37). a Pygostyle in lateral view. b, c Thoracic vertebrae. d, e Left coracoid in dorsal (d) and ventral (e) view. f, g Partial right coracoid in ventral (f) and dorsal (g) view. h, i Partial left scapula in lateral (h) and medial (i) view. j Partial right scapula in medial view. k, l Proximal end of right humerus in cranial (k) and caudal (l) view. m Fragmentary distal end of left humerus in cranial view. n, o Distal portion of right ulna in caudal (n) and distal (o) view. p Proximal end of left ulna in cranial view. q Distal end of left ulna in ventral view. r, s Proximal portions of both radii. t, u Partial right carpometacarpus in ventral (t) and dorsal (u) view. v Proximal and distal ends of left carpometacarpus in ventral view. w Both ossa carpalia ulnaria. x Phalanges proximales digiti majoris (one fragmentary). y Phalanx distalis digiti majoris. z, aa Pelvis in dorsal (z) and lateral (aa) view. bb–ee Right tarsometatarsus in dorsal (bb), plantar (cc), distal (dd), and proximal (ee) view. ff Pedal phalanx with fossilised integument of the foot pad; the arrow indicates an enlarged detail. gg Pedal phalanges. acr acromion, cdd condylus dorsalis, cdp crista deltopectoralis, cdv condylus ventralis, css crista spinosa synsacri, fac foramen acetabuli, fii foramen ilioischiadicum, flg plantar flange, fns foramen nervi supracoracoidei, fvp foramen vasculare proximale, itg fossilised integument, pex processus extensorius, pha pedal phalanx, pnf pneumatic foramen, ppc processus procoracoideus, prj ventrally-directed projection of caudal portion of facies articularis humeralis, sul hypotarsal sulcus, tbc, tuberculum carpale, tbd tuberculum dorsale, tpa tuberculum praeacetabulare. The scale bar equals 10 mm
The coracoid (Fig. 13d–g) has a large omal extremity, which measures about one third of the entire length of the bone, and a massive processus acrocoracoideus. The foramen nervi supracoracoidei is situated close to the cotyla scapularis, with the latter being cup-like and having a drop-shaped outline. The processus procoracoideus is well-developed. The sternal extremity of the bone exhibits a pointed and protruding angulus medialis. The long processus lateralis is narrow in the sterno-omal direction.
The shaft of the scapula (Fig. 13h–j) is slender. The acromion is short and tapers into a small, pointed projection. The caudal portion of the facies articularis humeralis forms a ventrally-directed projection.
The proximal end of the humerus (Fig. 13k, l) shows a distinctive morphology. The crista deltopectoralis is dorsally prominent and has a triangular shape with a concave proximal section; it is proximodistally shorter and more dorsally protruding than in Leptosomus discolor, Plesiocathartes kelleri, P. wyomingensis, and P. major (the humeri of other Plesiocathartes species are unknown). The crista deltopectoralis of Waltonavis is equally prominent, but lacks a concave proximal section. The tuberculum dorsale is small. The crista bicipitalis is poorly developed and does not form a proximodistally extensive crest as it does in extant Leptosomus. The fragmentary distal end of the humerus (Fig. 13m) has an undiagnostic morphology and resembles the corresponding part of the humerus of Leptosomus, Waltonavis and various other birds.
The partial radii do not exhibit diagnostic features. This is also true for the distal end of the ulna, the preserved portions of which (Fig. 13n, o, q) resemble the distal ulna of Leptosomus, Plesiocathartes, and Waltonavis. The fragment of the proximal ulna (Fig. 13p) has a distinct olecranon and a fairly large cotyla dorsalis.
The carpometacarpus (Fig. 13t–v) is less elongated than that of Waltonavis and resembles the carpometacarpus of the putative cathartid Diatropornis ellioti from the Quercy fissure fillings in France (Mourer-Chauviré 2002). The processus extensorius is cranially prominent, but less so than in Leptosomus discolor, and is separated by a marked concavity from the facies articularis alularis. The processus pisiformis is small and centrally situated. Except for the shallow fossa infratrochlearis, the ventral surface of the carpal trochlea around the pisiform process is poorly differentiated. Caudal to the processus pisiformis there is a longitudinal ridge, which probably extended onto the proximal portion of the os metacarpale minus. As in Plesiocathartes and Waltonavis (but in contrast to Leptosomus), the os metacarpale minus does not protrude beyond the os metacarpale majus on the distal end of the bone. The phalanx proximalis digiti majoris (Fig. 13x) is moderately long; whether the fenestra present in one of the phalanges is an artefact of preservation or a true feature is uncertain. The crus longum of the os carpi ulnare (Fig. 13w) is proportionally longer than in Waltonavis.
The pelvis (Figs. 13z, aa, 14a, b) has distinctive proportions in being very wide and dorsoventrally shallow. It is proportionally wider than the pelvis of Leptosomus discolor (Fig. 14c, d), to which it otherwise shows a general resemblance. The cristae iliacae dorsales are low and do not reach the crista spinosa synsacri. The preacetabular portion is comparatively long and exceeds the postacetabular portion in length. Unlike in L. discolor there is a small tuberculum praeacetabulare. The foramen ilioischiadicum is proportionally longer than in Leptosomus. Unlike in L. discolor and Plesiocathartes major there are no large foramina intertransversaria.

Comparison of some skeletal elements of Lutavis platypelvis, gen. et sp. nov. from the early Eocene London Clay of Walton-on-the-Naze, Essex, UK (holotype, NMS.Z.2021.40.37) and extant Leptosomus discolor (Leptosomidae). a–d Pelvis of L. platypelvis (a, b) and L. discolor (c, d; SMF 17388) in lateral (a, c) and dorsal (b, d) view. e, f Proximal end of the right humerus (caudal view) of L. platypelvis (e) and L. discolor (f; SMF 5438). g, h Right carpometacarpus of L. platypelvis (g) and L. discolor (h; SMF 5438) in ventral view. cbp crista bicipitalis, fac foramen acetabuli, fii foramen ilioischiadicum, fit foramina intertransversaria. The scale bar measures 10 mm
The tarsometatarsus (Fig. 13bb–ee) has similar overall proportions to the tarsometatarsus of Plesiocathartes, extant Leptosomiformes and the accipitriform Cathartidae. However, unlike in these taxa the hypotarsus forms two crests, which delimit a deeply concave sulcus, presumably for the tendon of the musculus flexor digitorum longus; a narrow and shallow sulcus is situated on the medial surface of the lateral hypotarsal crest. The medial foramen vasculare proximale is very large, as it is in Waltonavis. The distal end of the bone shows a close resemblance to that of Plesiocathartes. The fossa metatarsi I is only weakly developed. The trochlea metatarsi II reaches slightly farther distally than the trochlea metatarsi IV. The caudal margin of the plantar articular surface of the trochlea metatarsi III is broadly rounded. The trochlea metatarsi IV forms a distinct, rim-like plantar flange, which in distal view is slightly medially inflected.
The holotype includes several pedal phalanges, which lack the distal widening found in Plesiocathartes, Waltonavis, and Leptosomus. One of the pedal phalanges is preserved in a piece of clay matrix and is associated with a dark mineralized layer that appears to represent fossilised integument of the toe pads (Fig. 13ff). A similar preservation of the toe pads was also reported for a strigiform bird from Walton-on-the-Naze (Mayr and Kitchener in press).
Discussion
Plesiocathartes: Although a fair number of specimens were assigned to Plesiocathartes in the past two decades, all consist of compression fossils (P. kelleri, P. wyomingensis, P. major) or are represented by a few isolated bones, mainly partial tarsometatarsi (P. europaeus, P. geiselensis). As a consequence many details of the skeletal morphology of the taxon are not known. The Plesiocathartes fossils from Walton-on-the-Naze are likewise fragmentary, but they provide new data on major postcranial bones of the taxon and on various skeletal elements that are rarely preserved in Paleogene fossil birds.
Plesiocathartes and Leptosomus share a characteristic morphology of the coracoid (which has a crista procoracoidei and a foramen nervi supracoracoidei), a tibiotarsus with low and widely spaced condyli, and a “cathartid-like” tarsometatarsus with only weakly plantarly deflected trochleae metatarsorum II et IV. The new fossils show that Plesiocathartes also shares with the extant courol a widening of the distal ends of the pedal phalanges, which is particularly pronounced in P. insolitipes. A similar widening of the pedal phalanges occurs in Waltonavis, gen. nov. and some coraciiform taxa with syndactyl feet, in which the basal phalanges are united by connective tissue (Mayr 2022b). Plesiocathartes kelleri furthermore exhibits a long postorbital process (Mayr 2009: fig. 16.1), which is a derived characteristic of the Leptosomiformes.
The Plesiocathartes skeletons from Messel and the Green River Formation resemble Leptosomus discolor so closely in overall morphology that Mayr (2008) considered the extant courol to be a “living fossil”. However, there are also some differences between these fossils and the courol, and the new fossils add several distinguishing features. A notable disparity between P. insolitipes and crown group Leptosomiformes concerns the presence of pneumatic openings on the corpus of the thoracic vertebrae of the fossil species (these openings appear to be absent in P. major, whereas the morphologies of the corresponding vertebrae cannot be discerned in the P. kelleri and P. wyomingensis fossils). As detailed in the description, the furcula of Plesiocathartes has a different morphology to that of Leptosomus, which indicates divergent ecologies of the fossil and extant species (the shape of the furcula is related to various factors, including the size of ingested food and the morphology of the sternum, as well as the size and position of the attached flight muscles; Stegmann 1964).
The fossils described in the present study, as well as others that were assigned to Plesiocathartes (Mayr 2002a, b, Weidig 2006), are furthermore distinguished from extant Leptosomiformes in some features of the tarsometatarsus. The most notable difference concerns the hypotarsus on the proximal end of the bone. In the extant Leptosomus discolor the hypotarsus encloses two canals for the tendons of musculus flexor digitorum longus and m. flexor hallucis longus (Fig. 2gg), as does the hypotarsus of Waltonavis, gen. nov. (Fig. 2x). By contrast, in most fossils assigned to Plesiocathartes, the hypotarsus is block-like and lacks distinct sulci or canals (Fig. 2j). This hypotarsus type was designated as asulcate by Mayr (2015) and otherwise only occurs in the Cariamiformes, the accipitriform Cathartidae (Fig. 2kk) and Sagittariidae, and in the Sphenisciformes.
Hypotarsal canals are present in almost all representatives of the Eucavitaves, the sister taxon of the Leptosomiformes, which includes the Trogoniformes, Coraciiformes, and Piciformes (the lack of canals in the piciform Galbulae is likely to be a derived feature of these birds, which are deeply nested within Eucavitaves). Their absence in Plesiocathartes is here considered to be plesiomorphic and indicates that these canals evolved independently in the Leptosomiformes and Eucavitaves, presumably through ossification of cartilaginous structures.
Extant Leptosomiformes have semi-zygodactyl feet with a laterally spread fourth toe. This derived position of the toes is facilitated by the fact that the trochlea metatarsi IV bears a large, wing-like flange (Fig. 2ff). The hypotarsus morphology of extant Leptosomiformes, with two closed canals, may be functionally related to this foot structure, because in all extant zygodactyl birds, except for the Piciformes, the tendons of musculus flexor digitorum longus and m. hallucis longus run through ossified hypotarsal canals (Mayr 2015).
As noted in the introduction, the taxon Plesiocathartes was established for P. europaeus from the Quercy fissure fillings in France, the holotype of which is a tarsometatarsus lacking the proximal end. Mourer-Chauviré (2002) referred a more complete tarsometatarsus to the species, in which a partial hypotarsus is preserved. The hypotarsus of this referred tarsometatarsus differs from that of other fossils assigned to Plesiocathartes in that it is not block-like, but exhibits a sulcus bordered by two crests (Mourer-Chauviré 2002: fig. 1T). If this tarsometatarsus is indeed from P. europaeus, the species with a block-like hypotarsus would have to be assigned to a new genus. However, and as also noted by Mourer-Chauviré (2002), the referred P. europaeus tarsometatarsus differs from the holotype in several features, including a narrower shaft. Mourer-Chauviré (2002) attributed these differences to individual variation, but given the diversity of Plesiocathartes-like birds in the London Clay, it appears more likely that they are of taxonomic significance and even more so, because the referred P. europaeus tarsometatarsus shows a resemblance to Lutavis platypelvis, gen. et sp. nov. concerning the shape of the hypotarsus.
Waltonavis: In size (Tab. 1) and – as far as comparisons are possible – morphology, the two species of Waltonavis resemble Lapillavis incubarens from the latest early or earliest middle Eocene of Messel (Mayr 2016). The holotype and only known specimen of this latter species is known from a rather poorly preserved partial skeleton, which does not allow the recognition of many osteological details. The phylogenetic affinities of L. incubarens were considered to be uncertain in the original description (Mayr 2016), but a possible relationship to the Leptosomiformes was noted. A similar coracoid to that of Waltonavis was reported by Zvonok and Gorobets (2016) from the middle Eocene of Ukraine, who also noted a resemblance between this bone and the coracoid of the Leptosomiformes.
Compared to extant birds, Waltonavis most closely resembles the Leptosomiformes in its overall skeletal morphology (that is, the shape and proportions of the major limb bones), but it is less straightforward to constrain the affinities of the new taxon in terms of derived characters. Waltonavis and crown group Leptosomiformes share a carpometacarpus with a ventral projection on the proximal portion of the os metacarpale minus (Fig. 9n), widely spaced condyli on the distal end of the tibiotarsus, a hypotarsus with two canals (Fig. 10r, t), a trochlea metatarsi II that reaches much farther distally than the trochlea metatarsi IV, and lateral bulges on the distal ends of the first two phalanges of the third toe. The similarities between Waltonavis and leptosomiform birds may be due to the retention of plesiomorphic resemblances, but no character evidence for an alternative placement of the new taxon could be identified. The presumably plesiomorphic presence of a foramen nervi supracoracoidei distinguishes the fossil from all extant coraciiform and piciform birds. In addition, the os carpi radiale does not show a marked sulcus for the tendon of musculus extensor longus alulae, the presence of which characterises extant Bucerotes, Coraciiformes, and Piciformes (Mayr 2014).
The two species of Waltonavis are smaller than Leptosomus discolor and the species of Plesiocathartes (Tab. 1), and they differ from Leptosomus and Plesiocathartes in skeletal features (see differential diagnosis). Judging from the shape of the omal portions of the furcular shafts, which are not widened and strap-like (compare Fig. 4u, v and Fig. 6d), and the morphology of the coracoid, which has a less pronounced crista procoracoidei (Fig. 2e, f), Waltonavis is outside a clade formed by Plesiocathartes and Leptosomus. However, unlike in Plesiocathartes, but as in the extant Leptosomus, the hypotarsus of Waltonavis encloses two canals for the deep flexor tendons (the hypotarsus morphology of Lapillavis is unknown).
The well-developed facies articularis basipterygoidea of the pterygoid suggests the presence of functional basipterygoid processes in Waltonavis, which may be another plesiomorphic feature of the new taxon (as detailed in the description, Plesiocathartes lacks basipterygoid processes). Amongst extant representatives of Telluraves, the clade that includes most of the arboreal land birds, basipterygoid processes only occur in the accipitriform Cathartidae and Sagittariidae as well as in the Strigiformes and Trogoniformes. An ancestral state reconstruction (Fig. 15) does not unambiguously resolve the condition in the stem species of Telluraves, but because basipterygoid processes occur in palaeognathous and galloanserine birds, which are successive sister groups of all other neornithine (crown group) birds, it is here considered likely that the presence of these processes also represents the ancestral condition for both Telluraves and the clade that includes the Accipitriformes, Strigiformes, Leptosomiformes, and Trogoniformes.

Ancestral state reconstruction of basipterygoid processes in taxa of the Telluraves; extinct taxa are indicated by a dagger (concerning the extant taxa, the tree topology is based on the analyses of Prum et al. 2015 and Kuhl et al. 2021). Black lines indicate the occurrence of basipterygoid processes, white lines their absence. Exclusion of Waltonavis did not alter the ancestral state reconstructions for the other taxa included in the analysis.
The extant Leptosomus discolor is a carnivorous bird and feeds on larger insects and small reptiles, which are usually caught by sallying flights from perches (Goodman 2001). The smaller size of Waltonavis, its narrower beak, the presence of basipterygoid processes, as well as some differences in the morphologies of the limb bones, suggest that the new taxon differed from Leptosomus in its feeding habits. However, without more data on the skull morphology of Waltonavis, conclusions on its feeding ecology remain speculative. The bulges on the distal ends of the first two phalanges of the third toe represent a distinctive feature of the new taxon. Similar bulges are also present in Plesiocathartes insolitipes, sp. nov. and the extant L. discolor, where they occur on both sides of the phalanges and are smaller. These bulges also occur in a less pronounced form in some coraciiform birds, where they are likely to be correlated with the presence of syndactyl feet (Mayr 2022b). However, Leptosomus lacks syndactyl feet and the functional significance of the bulges in Waltonavis therefore remains unknown.
Lutavis: Mainly owing to the similar morphology of the proximal and distal tarsometatarsus, and based on a brief examination of the specimen in 2008, the Lutavis platypelvis holotype was compared with the taxon Paracrax by Mayr (2022a). This putative stem group representative of the Cariamiformes occurred in the early Oligocene of North America, but apart from the temporal and geographical discrepancies in their occurrences, Lutavis and Paracrax are clearly distinguished from each other in osteological features. Most notably, the coracoid of Lutavis (Fig. 2e) is very dissimilar from that of Paracrax and this is also true for the pelvis, which shows little resemblance to the pelvis of cariamiform birds.
With regard to tarsometatarsus proportions and the morphology of the distal end of the bone, Lutavis closely resembles Plesiocathartes, but a similar tarsometatarsus morphology also occurs in the accipitriform Cathartidae (Fig. 2hh–kk). Cathartidae and Leptosomiformes are only separated by the Strigiformes and Coliiformes in current sequence-based phylogenies (Fig. 15; Prum et al. 2015, Kuhl et al. 2021), both of which exhibit autapomorphic tarsometatarsus and hypotarsus morphologies. Therefore, similarities in the tarsometatarsi of Plesiocathartes and the Cathartidae may well be plesiomorphic for the Afroaves.
Except for the lack of hypotarsal canals, Plesiocathartes is morphologically closer to crown group Leptosomiformes than is Waltonavis, gen. nov. Features shared by Plesiocathartes and Leptosomus that are absent in Waltonavis include a massive, roller-like beak, a large and ventrally prominent condylus medialis of the quadrate, a crista procoracoidei of the coracoid, and a furcula with shafts that are markedly wider towards the omal extremities of the bone.
The Lutavis platypelvis holotype does not exhibit derived features, which allow an unambiguous assignment to any of the avian higher-level clades, and its classification is therefore left open in the present study. L. platypelvis lacks derived characteristics shared by Plesiocathartes and Leptosomus (crista procoracoidei), and despite the similarity to Plesiocathartes in tarsometatarsus morphology, even a tentative assignment to the Leptosomiformes cannot be established with certainty and would be speculative.
L. platypelvis was a relatively short-legged bird. Unlike in birds that are capable of powerful sudden take-offs (e.g. Galliformes and Columbiformes), the tuberculum dorsale of the humerus is small, but the massive coracoid and dorsally prominent crista deltopectoralis indicate well-developed flight capabilities. L. platypelvis differs from the Leptosomiformes in that the proximodorsal portion of the humerus forms a concavity and in that the hypotarsus exhibits a deep sulcus bordered by two crests. The tarsometatarsus shows an overall resemblance to the Cathartidae (as does the tarsometatarsus of Plesiocathartes), whereas the hypotarsus is more similar to that of the Accipitridae. However, the shallow pelvis of Lutavis precludes a position of the taxon within crown group Accipitriformes and, hence, any closer affinities to the Cathartidae and Accipitridae.
Conclusions
The fossils described in this study increase the taxonomic diversity of leptosomiform and leptosomiform-like birds in the early Eocene of Europe. Waltonavis, gen. nov. and Lutavis, gen. nov. potentially also suggest a higher ecomorphological diversity of stem group Leptosomiformes in the early Eocene. However, the affinities of Lutavis are currently not well understood, and even though we consider the leptosomiform affinities of Waltonavis, gen. nov. to be best supported by the current character evidence, there remains a possibility that a Leptosomus-like skeletal morphology is plesiomorphic for the clade including the Leptosomiformes, Trogoniformes (trogons), Coraciiformes (true rollers), and Piciformes (woodpeckers and allies).
An improved assessment of the evolutionary significance of Waltonavis and Lutavis will probably be facilitated once the numerous coraciiform-like birds in the Daniels collection have been described and studied. When these studies have been performed, we aim at performing a comprehensive phylogenetic analysis that includes a broad array of stem group representatives of the arboreal taxa of Telluraves, and we hope that by then also more osteological data have been gathered for Lapillavis, which will enable us to assess the affinities between this taxon and leptosomiform birds.
References
Aldiss, D. T. (2012). The stratigraphical framework for the Palaeogene successions of the London Basin, UK. British Geological Survey Open Report, OR/12/004, 1–87.
Collinson, M. E., Adams, N. F., Manchester, S. R., Stull, G. W., Herrera, F., Smith, S. Y., Andrew, M. J., Kenrick, P., & Sykes, D. (2016). X-ray micro-computed tomography (micro-CT) of pyrite-permineralized fruits and seeds from the London Clay Formation (Ypresian) conserved in silicone oil: a critical evaluation. Botany, 94, 697–711.
Cracraft, J., & Rich, P. V. (1972). The systematics and evolution of the Cathartidae in the Old World Tertiary. The Condor, 74, 272–283.
Elzanowski, A., & Stidham, T.A. (2010). Morphology of the quadrate in the Eocene anseriform Presbyornis and extant galloanserine birds. Journal of Morphology, 271, 305–323.
Ericson, P. G. P., Anderson, C. L., Britton, T., Elzanowski, A., Johansson, U. S., Källersjö, M., Ohlson, J. I., Parsons, T. J., Zuccon, D., & Mayr, G. (2006). Diversification of Neoaves: integration of molecular sequence data and fossils. Biology Letters, 2, 543–547.
Feduccia, A. (1999). The Origin and Evolution of Birds. 2nd edition. New Haven: Yale University Press.
Gaillard, C. (1908). Les oiseaux des Phosphorites du Quercy. Annales de l’Université de Lyon (Nouvelle Série), 23, 1–178.
Goodman, S. M. (2001). Family Leptosomidae (Cuckoo-rollers). In J. del Hoyo, A. Elliott, & J. Sargatal, (Eds.), Handbook of the Birds of the World, volume 6: Mousebirds to Hornbills (pp. 390–395). Barcelona: Lynx Edicions.
Hackett, S. J., Kimball, R. T., Reddy, S., Bowie, R. C. K., Braun, E. L., Braun, M. J., Chojnowski, J. L., Cox, W. A., Han, K.-L., Harshman, J., Huddleston, C. J., Marks, B. D., Miglia, K. J., Moore, W. S., Sheldon, F. H., Steadman, D. W., Witt, C. C., Yuri, T. (2008). A phylogenomic study of birds reveals their evolutionary history. Science, 320, 1763–1767.
Kuhl, H., Frankl-Vilches, C., Bakker, A., Mayr, G., Nikolaus, G., Boerno, S. T., Klages, S., Timmermann, B., & Gahr, M. (2021). An unbiased molecular approach using 3’UTRs resolves the avian family-level tree of life. Molecular Biology and Evolution, 38, 108–127.
Maddison, W. P., & Maddison, D. R. (2009). Mesquite: a modular system for evolutionary analysis; version 2.71. http://mesquiteproject.org.
Mayr, G. (1998). “Coraciiforme” und “piciforme” Kleinvögel aus dem Mittel-Eozän der Grube Messel (Hessen, Deutschland). Courier Forschungsinstitut Senckenberg, 205, 1–101.
Mayr, G. (2002a). Avian remains from the Middle Eocene of the Geiseltal (Sachsen-Anhalt, Germany). In Z. Zhou, & F. Zhang, (Eds.), Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, Beijing, 1-4 June 2000 (pp. 77–96). Beijing: Science Press.
Mayr, G. (2002b). A new species of Plesiocathartes (Aves: ?Leptosomidae) from the Middle Eocene of Messel, Germany. PaleoBios, 22, 10–20.
Mayr, G. (2008). The Madagascan “cuckoo-roller” (Aves: Leptosomidae) is not a roller - notes on the phylogenetic affinities and evolutionary history of a “living fossil”. Acta Ornithologica, 43, 226–230.
Mayr, G. (2009). Paleogene fossil birds. Heidelberg: Springer.
Mayr, G. (2014). Comparative morphology of the radial carpal bone of birds and the phylogenetic significance of character variation. Zoomorphology, 133, 425–434.
Mayr, G. (2015). Variations in the hypotarsus morphology of birds and their evolutionary significance. Acta Zoologica, 97, 196–210.
Mayr, G. (2016). Fragmentary but distinctive: three new avian species from the early Eocene of Messel, with the earliest record of medullary bone in a Cenozoic bird. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 279, 273–286.
Mayr, G. (2017). Avian evolution: The fossil record of birds and its paleobiological significance. Chichester: Wiley-Blackwell.
Mayr, G. (2020). An updated review of the middle Eocene avifauna from the Geiseltal (Germany), with comments on the unusual taphonomy of some bird remains. Geobios, 62, 45–59.
Mayr, G. (2021). On the occurrence of lateral openings and fossae (pleurocoels) in the thoracic vertebrae of neornithine birds and their functional significance. Vertebrate Zoology, 71, 453–463.
Mayr, G. (2022a). Paleogene fossil birds, 2nd edition. Cham, Switzerland: Springer.
Mayr, G. (2022b). A partial skeleton of Septencoracias from the early Eocene London Clay reveals derived features of bee-eaters (Meropidae) in a putative stem group roller (Aves, Coracii). Palaeobiodiversity and Palaeoenvironments, 102(2), 449–463.
Mayr, G., & Kitchener, A.C. (in press). Early Eocene fossil illuminates the ancestral (diurnal) ecomorphology of owls and documents a mosaic evolution in the strigiform stem lineage. The Ibis.
Mayr, G., & Smith, T. (2019a). A diverse bird assemblage from the Ypresian of Belgium furthers knowledge of early Eocene avifaunas of the North Sea Basin. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 291, 253–281.
Mayr, G., & Smith, T. (2019b). New Paleocene bird fossils from the North Sea Basin in Belgium and France. Geologica Belgica, 22, 35–46.
Mayr, G., Rana, R. S., Rose, K. D., Sahni, A., Kumar, K., Singh, L., & Smith, T. (2010). Quercypsitta-like birds from the early Eocene of India (Aves, ?Psittaciformes). Journal of Vertebrate Paleontology, 30, 467–478.
Mourer-Chauviré, C. (2002). Revision of the Cathartidae (Aves, Ciconiiformes) from the Middle Eocene to the Upper Oligocene Phosphorites du Quercy, France. In Z. Zhou, & F. Zhang (Eds.), Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, Beijing, 1-4 June 2000 (pp. 97–111). Beijing: Science Press.
Prum, R. O., Berv, J. S., Dornburg, A., Field, D. J., Townsend, J. P., Lemmon, E. M., & Lemmon, A. R. (2015). A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature, 526, 569–573.
Rayner, D., Mitchell, T., Rayner, M., & Clouter, F. (2009). London Clay fossils of Kent and Essex. Rochester, Kent: Medway Fossil and Mineral Society.
Sangster, G., Braun, E. L., Johansson, U. S., Kimball, R. T., Mayr, G., & Suh, A. (2022). Phylogenetic definitions for 25 higher-level clade names of birds. Avian Research, 13, 100027.
Stegmann, B. (1964). Die funktionelle Bedeutung des Schlüsselbeines bei den Vögeln. Journal für Ornithologie, 105, 450–463.
Weidig, I. (2006). The first New World occurrence of the Eocene bird Plesiocathartes (Aves: ?Leptosomidae). Paläontologische Zeitschrift, 80, 230–237.
Zvonok, E., & Gorobets, L. (2016). A record of a landbird (Telluraves) from the Eocene Ikovo locality (East Ukraine). Acta Zoologica Cracoviensia, 59, 37–45.
Acknowledgements
We thank Sven Tränkner for taking most of the photographs (additional images were taken by GM). ACK thanks David Bain for his considerable assistance in the transfer of the Daniels collection to NMS. Georg Hantke helped with the packing and transport of the specimens. Vicen Carrió (NMS) carried out some preparation work on the Plesiocathartes skeleton associated with NMS.Z.2021.40.34. Comments from Ursula Göhlich and an anonymous reviewer improved the manuscript.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Data availability statement
All data generated or analysed during this study are included in this published article.
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is registered in Zoobank under: http://zoobank.org/urn:lsid:zoobank.org:pub:921446D9-F62B-4C74-8DFE-3FBF19CBEF38
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mayr, G., Kitchener, A.C. New species from the early Eocene London Clay suggest an undetected early Eocene diversity of the Leptosomiformes, an avian clade that includes a living fossil from Madagascar. Palaeobio Palaeoenv 103, 585–608 (2023). https://doi.org/10.1007/s12549-022-00560-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-022-00560-0