
Abstract
The Hagen-Balve Reef Complex of the northern Rhenish Massif, Germany, is one of the best examples for the globally widespread Givetian to Frasnian reefs. At its eastern end, in the Hönne Valley, it reaches up to 1000 metres in thickness. In general, there is limited knowledge concerning the faunas and palaeoecology of Rhenish initial reef phases. A unique section at Binolen exposes the sharp boundary between the underlying, siliciclastic Upper Honsel Formation and the lower part of the reefal Hagen-Balve Formation. A ca. 14 m thick succession is assigned to the new Binolen Member and investigated in terms of carbonate microfacies, micro-, and macrofaunas. Seven microfacies types (MF) characterise a crinoidal coral-stromatoporoid garden setting (MF-2 to MF-6) or allobiostrome of an initial open platform turning upsection into a coral parabiostrome (MF-7/8). The base of the Asbeck Member is locally defined by the appearance of back-reef facies types, such as fenestral limestones (MF-9), which indicate the transformation into a bioherm. Low-diversity conodont faunas prove an influx of open shelf organisms for the Binolen Member and suggest a lower Givetian age (probably Polygnathus timorensis Zone). There are two new Givetian conodont biofacies types characteristic for open biostrome settings, the Bipennatus Biofacies and a Simple Cone Biofacies. Thin sections reveal over 40 different reef builders (rugose and tabulate corals, stromatoporoids, and chaetetid sponges). Around 4200 microfossils from conodont residues consist of agglutinating foraminifers, calcareous sponge spicules (e.g. Heteractinida), scolecodonts, echinoderms (mostly crinoid ossicles and holothurian sclerites), pyritised benthic ostracods, assumed calcified moulds of chitinozoans, subordinate microvertebrates, and other groups. They were analysed quantitatively in order to document changing distribution patterns and environments through time. We distinguish six microfossil-based biofacies types (BF), namely the Ostracoda Biofacies (BF-O), Foraminifera Biofacies (BF-F), Scolecodont Biofacies (BF-S), Chitinozoa Biofacies (BF-C), Porifera-Scolecodont Biofacies (BF-PS), and Porifera-Echinodermata Biofacies (BF-PE). Microfacies and microfossil assemblages led to a reconstruction of the local facies development, consisting of seven successive depophases with further subdivisions. At the base, the sudden termination of siliciclastic influx enabled a proliferation of reef builders and microfaunas (Depophase I), followed by a longer deepening phase characterised by exclusive bioclastic wackestones (Depophase II). Depophase III is a short regressive phase with crinoidal coral-stromatoporoid floatstones. Depophase IV is characterised by fluctuations in facies and relative sea-level, which results in three subphases. Depophase V resembles Depophase III, but with argillaceous interbeds and more abundant scolecodonts. It is followed by an interval with coral-stromatoporoid rudstones, which grade into coral-dominated bafflestones of an (auto)parabiostrome (both Depophase VI). Depophase VII represent the back-reef facies, including fenestral pack-bindstones of the basal Asbeck Member. The systematic analysis of reefal microfaunas represents pioneer work that should be tested and expanded to other reefs of the Rhenish Massif and beyond.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The Devonian, especially the Givetian and Frasnian, was a phase of extensive reef growth on a global scale, only rivalled in terms of abundance by the Late Jurassic and early Neogene (Kiessling et al. 1999; Flügel and Kiessling 2002; Copper and Scotese 2003; Kiessling 2008). The Hönne Valley Reef, the eastern part of the Hagen-Balve Reef Complex in the northern Rhenish Massif, is an outstanding example in terms of thickness (up to 1000 metres), economic significance, and exposure in partly huge active or abandoned quarries. Pioneering studies go back to the early 20th century (see publication history in Becker et al. 2016).
To understand the evolutionary history of reef carbonate platforms, all phases need to be studied, from the initial stadium to the period of main reef growth and final extinction. Fortunately, all phases are observable in the Hönne Valley region. Numerous publications deal with the main reef (e.g. Jux 1960; May 1988, 1991, 1997; Koch-Früchtl and Früchtl 1993; Schudack 1993; May and Marks 2013), not listing the many unpublished Diploma, mapping, and B.Sc. theses, and the eventual drowning and extinction in the Frasnian (e.g. Stichling 2011; Stichling et al. 2022, this issue). However, except for a preliminary description by Becker et al. (2016), no previous study focused on the initial phase of the reef, which is only exposed near Binolen. In general, there is still limited knowledge of Rhenish initial reef phases with respect to different ages and general facies settings (e.g. intra-shelf versus outer shelf edge versus seamount). This publication aims to narrow this knowledge gap, also as a base for future comparisons beyond the Rhenish Shelf. It includes pioneer work since, apart from occasional records (e.g. Flügel and Hötzl 1971; Langer 1979, 1997; Malmsheimer et al. 1996), only Koch-Früchtl and Früchtl (1993) made a limited previous attempt to obtain microfauna successions from conodont sample residues of Rhenish reef facies.
The paper is primarily based on the unpublished B.Sc. thesis of the first author (Löw 2020), who focused on the microfaunas from the initial reef stadium of Binolen, a conference abstract (Löw et al. 2021), and on data from the Ph.D. cooperation project of S. Stichling and R. T. Becker with the Lhoist Germany Rheinkalk GmbH. The major focus lies on:
-
A bed-by-bed bio- and lithostratigraphic analysis
-
Carbonate microfacies analysis and local reef palaeoecology
-
Description of micro- and macrofaunas
-
Quantitative micropalaeontological analysis
-
Biofacies analysis based on reef microfaunas and in relation to microfacies types
Abbreviations
Alv. = Alveolites, Bi. = Bipennatus, C. = Carpinaria, Cl. = Clathrocoilona, Dendro. = Dendrostella, Dv. = Dvorakia, Eo. = Eocaudina, G. = Gondekia, I. = Icriodus, Neop. = Neopanderodus, P. = Pachyfavosites, Po. = Polygnathus, S. = Stromatopora, Sc. = Scoliopora, Sp. = Spinocyrtia, St. = Stachyodes, T. = Temnophyllum, Th. = Thamnopora, Thur. = Thurammina, To. = Tolypammina.
Material and methods
Our study is based on a bed-by-bed logging of the Binolen outcrop for lithology, microfacies, micro-, and macrofaunas. The section had to be cleared from vegetation; it tends to be overgrown strongly in wet summers. A total of 72 beds were examined, with four minor outcrop gaps, 39 micropalaeontological samples taken, and 75 thin sections prepared. For the sedimentological description, we follow the Dunham classification for carbonate rocks (Dunham 1962; Embry and Klovan 1971). Additionally, we included carbonate microfacies terms coined by Tsien (1982, 1984). The characterisation of microfacies types (MF) is based on the Facies Zones (FZ) after Wilson (1975) and on the Standard Microfacies Types (SMF) sensu Flügel (1978, 2010). For the micropalaeontological analysis (conodonts and BF types), 2–3 kg limestone samples were dissolved in 10% formic acid solution. Residues were washed and separated into 0.1–0.315, 0.315–0.63, and >0.63 mm fractions. Weak acid leaching enables the full recovery of phosphatic (conodonts), organic (scolecodonts), siliceous (locally not present), pyritic (secondarily goethitic: ostracods), or agglutinated microfossils (foraminifers). It is also well-known (e.g. Langer 1991) that by the preferential dissolution of the finest carbonate matrix, complete calcareous sklerites and shells can be recovered if samples are not acidized for too long. This applies to echinoderms, which consist of single Mg-calcite crystals, but also to delicate, calcitic sponge spicules and assumed calcified chitinozoans. For an even microfossil recovery, all samples were uniformly washed and sieved after two days, and residues were completely picked. The presence of similarly preserved calcareous skeletal elements in most samples suggests that the processing did not introduce a significant bias. This is supported by the fact that we observed no increased corrosion of specimens in samples with a low content of calcareous microfossils. The slender and corrosion-prone calcitic monaxone spicules or delicate holothurian sklerites occur throughout the section, the first even in beds without any echinoderm remains. Ostracods are often found in limestone residues due to diagenetic silicification, but we obtained only primarily pyritised specimens that were oxidised in the weathering zone. The lack of silicified microfossils reflects the absence of primarily siliceous skeletal elements (e.g. hexactinellid spicules, radiolaria) that commonly become the source of diagenetically mobilised SiO2.
A total of ca. 4200 microfossils were collected during this study. Representative specimens are pictured using either a Keyence VHX digital microscope or a SEM. Calcareous sponges, annelids, the various echinoderm groups, and ostracods produced different numbers of skeletal elements/shells during their life and after death, in contrast to foraminifers or chitinozoans, which represent individuals. Therefore, the counted microfossil numbers do not reflect proportional abundances of protists/animals in the palaeoecosystem. The arrival of a few sponges creates a stronger signal in the raw data than the invasion of the same number of foraminifers, assuming a similar preservation potential. This does not diminuish the suitability of abundance data to characterise empirical assemblage types and to interpret their fluctuations in time in terms of relative shifts of palaeoecology and taphonomy.
The presence of differently sized microfossils in all samples suggests that there was no clear size fractionation by the variable water agitation, which can be inferred from the microfacies analysis. The lack of tempestites or turbidites suggests that there was no admixture of material with different provenance during sudden sedimentary events. The Binolen microfossil assemblages represent organisms that partly lived close-by or that were washed in from neighbouring areas of the initial carbonate platform. Therefore, observed fluctuations of microfossil assemblages do not necessarily record ecological changes at the collected spot but in the overall regional biostrome. However, the interpretation of assemblage changes has to take transport variation into account.
All micropalaeontological samples and thin sections are housed in the Geomuseum of the Westfälische Wilhelms University, Münster (GMM) under the numbers GMM B1B.2-1–14 (foraminifers and chitinozoans), B2B.9-1–6 (porifers), B3A.8-1–6 (holothuroids and echinoids), B3A.2-1–6 (crinoids), B3C.1-1 (ophiuroid), B4A.8-1–6 (scolecodonts), B7B.6-1–3 (ostracods), B5A.5-1 (bryozoan), B6B.12-1 (gastropod), B9A.15-1–32 (conodonts), B5B.18-1–2 (brachiopods), B2C.58-1 (coral), and A1C.6-1–3 (fish remains). Supplementary material (thin sections and macrofossils) is stored by the co-author S. F. Zoppe (SFZ-4613-01, -02, -03, -07, -10, -11).
Geological setting, litho-, and biostratigraphy
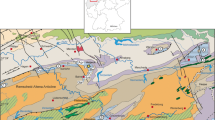
The Binolen section is situated in the northern part of the Rhenish Massif (Hönne Valley, northern Sauerland; Fig. 1), at the eastern end of the Remscheid-Altena Anticline. The locality consists of several small outcrops along a forest track running on the western slope in parallel to the Hönne river (Fig. 2). Access is from the parking area of “Haus Recke”, following the small road towards the river and after crossing the Hönne via a small bridge. The forest track branches just after the bridge towards the north, at GPS 51°22’09.6” N, 7°51’31.1” E (topographic map 1:25000, 4613 Balve; Fig. 3). The measured strata are dipping eastwards, due to their position on the eastern axial ramp of the Remscheid-Altena Anticline. Below and at the base of our measured section, middle-grey, micaceous, cross-bedded, and poorly fossiliferous siltstones of the Upper Honsel Formation occur (Figs. 2a, 4). These rocks are folded, with the top of the formation being exposed on the northern limb of a 4th order (decametre-scale) anticline. The siltstones represent distal siliciclastic deposits of pro-deltaic, shallow water environments deposited along the southeastern shelf of Laurussia (Langenstrassen 1983; Hartkopf-Fröder and Weber 2016; Fig. 5). They yielded locally the bivalve Ptychopteria and fragmentary spiriferids (Becker et al. 2016). Jux (1960) mentioned Spinocyrtia (Carpinaria) ascendens (possibly Sp. (C.) beckmanni) from west of Binolen, which provided a lower Givetian age (Becker et al. 2016). This corresponds well to the records of May (1991), who found at the top of the Upper Honsel Formation at Bäingsen (west of Binolen) the marker brachiopods Schnurella custos, Dicamara prunulum, Undispirifer undiferus, Spinatrypina girzenensis, and Desquamatia (Variatrypa) ajugata. Especially the latter two are useful to correlate the Upper Honsel Formation with the lower Givetian Dreimühlen Formation of the Eifel Mountains. The Dreimühlen Formation was placed by Weddige (1988) in the middle part of the Po. hemiansatus Zone (MD II-A).

Location of the Binolen section and Steltenberg Quarry within the northern Rhenish Massif. Thin lines show the margins of the surface Carboniferous and of the Lower Devonian in the core of major anticlines

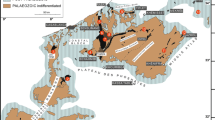
Field photos (taken by SFZ) from the outcrop of Binolen. Hammer (b, c) and backpack (a, d) for scale. a Upper Honsel Formation underneath the investigated section. b Lower boundary of the new Binolen Member of the Hagen-Balve Formation at the base of Bed 4 (first limestone bed), overlying the uppermost siliciclastic strata of the Upper Honsel Formation. c Section of locally folded, well-bedded allo- to parabiostromes. d Cliff of the Binolen section, yielding (auto)parabiostromal beds with predominance of baffling-types of reef builders

Locality map at Binolen in the Hönne Valley

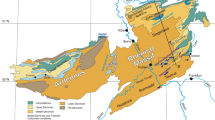
Section log of Binolen showing the lithology, depophases, reconstructed sea-level fluctuations, lithostratigraphic boundaries, microfacies and biofacies succession as well as the position of microfossil samples

Palaeogeograpic location of Binolen, modified after Langenstrassen (1983)
The occurrence of Stringocephalus at the base and within the overlying biostromal limestones of our section supports an overall Givetian age (Fig. 6a). This terebratulid brachiopod is a classical index fossil for the Givetian stage within the Rhenish Massif and elsewhere (e.g. Beyrich 1837; Struve 1982; Sun and Boucot 1999). Above the siltstones of the Upper Honsel Formation, our bed-by-bed survey started with the sharp onset of dark- to middle-grey, calcareous, coarse crinoid limestones and first substrate-stabilising organisms (e.g. sponges, tabulate corals, and stromatoporoids). The abrupt decrease in siliciclastic discharge from Laurussia enabled the development of a biostromal succession. Since there was no evidence for initial deepening (see microfacies analysis), this probably reflects climatic changes, such as increased aridity in the Laurussian source region (Becker et al. 2016). A change in continental drainage directions as a reason for the decrease of siliciclastic discharge is unlikely, because wedges with coarse siliclastics disappeared everywhere in the northen Rhenish Massif. We place the base of the reefal Hagen-Balve Formation at the sharp onset of limestones (base of Bed 4; Figs. 2b, 4) although a typical, poorly-bedded “Massenkalk” facies is not yet developed. The here described biostromal initial reef succession is assigned to the new Binolen Member of the Hagen-Balve Formation, with the slope of the forest track as the type-section and a total thickness of ca. 14 m. It is notable, that already the Upper Honsel Formation includes several biostromes with abundant reef builders, but these are always small, isolated local structures, which were quickly buried by renewed siliciclastic influx. The Hagen-Balve Formation, therefore, represents a time interval when the arrival of silt and fine sand stopped for a long time, lasting in the Hönne Valley region until well after the reef extinction, to the top of the Famennian (e.g. Becker et al. 2021a).

Givetian macro- and microfossils from Binolen. a Stringocephalus sp., loose, GMM B5B.18-1. b-c Mesophyllum (Mesophyllum) vesiculosum (Goldfuss, 1826), Bed 12, GMM B2C.58-1, transverse (b) and longitudinal (c) section. d Tolypammina cf. continuus Gutschick, 1962, Bed 6, GMM B1B.2-1. e To. tortuosa Dunn, 1942, Bed 5, GMM B1B.2-2. f To. aff. tortuosa, Bed 6, GMM B1B.2-3. g Psammosphaera fusca Schulze, 1875, Bed 5, GMM B1B.2-4. h Thurammina cf. congesta Gutschick, Weiner and Young, 1961, Bed 6, GMM B1B.2-5. i Thur. cf. sphaerica Ireland, 1939, Bed 44, GMM B1B.2-6. j-k Webbinelloidea hemisphaerica Stewart and Lampe, 1947, double-chambered specimen, Bed 5, GMM B1B.2-7, single-chambered specimen, Bed 6, GMM B1B.2-8. l Rhizammina cf. algaeformis Brady, 1879, Bed 5, GMM B1B.2-9. m Ensiferites langeri Kozur, Mostler and Repetski, 2008, Bed 5, GMM B2B.9-1. n Gondekia sp., Bed 5, GMM B2B.9-2. o Praephobetractinia eocambrica Kozur, 1991, Bed 5, GMM B2B.9-6. p Dvorcia cf. mira Nekvasilova and Stemprokova, 1960, Bed 8, GMM B2B.9-5. q straight monaxon, Bed 5, GMM B2B.9-3. r curved monaxon, Bed 24, GMM B2B.9-4
Numerous conodont samples were taken in the hope for a more precise age determination or even a local succession. Although rare conodonts were obtained from 15 beds, this attempt failed since mostly long-ranging, zonally non-diagnostic taxa were found. There are two different assemblages. The first limestone (Bed 4) as well as beds 18a, 18b, 23, and 29 yielded Pa elements of Bi. bipennatus, a species that was first described as a species of Spathognathodus by Bischoff and Ziegler (1957). It is the characteristic conodont of the lower Givetian, biostromal Sparganophyllum Limestone of the Sauerland (Bischoff and Ziegler 1957). It was later also found in the lower Givetian of the Eifel Mountains (Weddige 1977) and then in many other regions. Bultynck (1987) distinguished two morphotypes; our material clearly belongs to the narrow Morphotype α (Fig. 7l), which was shown to have a lower range than Morphotype ß. Mawson (1993) introduced the genus Bipennatus, summarised the published evidence, and suggested that both morphotypes range down into the lower/middle Eifelian. However, it is now known that Eifelian Bipennatus includes several morphologically similar species that have previously been included in wide taxonomic concepts of Bi. bipennatus (see Narkiewicz 2015; Lu and Königshof 2019). As a consequence, Eifelian records of Bi. bipennatus require a critical re-evaluation. The stratigraphically youngest records are from the top of the middle Givetian of Queensland (Mawson and Talent 1989) and southern Morocco (Aboussalam and Becker 2011), but such specimens are very rare. At Binolen, Bipennatus is associated with rare single, smooth cones (Drepanodus sp., Bed 4) and polygnathid ramiforms (beds 4, 18a, 23). A single, incomplete icriodid from Bed 4 resembles I. latecarinatus, which enters in the lower Givetian Po. timorensis Zone (Narkiewicz and Bultynck 2007).

Assumed chitinozoan moulds, scolecodonts and conodonts from Binolen. a Angochitina pseudoavelinoi Gaugris and Grahn, 2006, Bed 31, GMM B1B.2-10. b Angochitina pseudoavelinoi Gaugris and Grahn, 2006, Bed 31, GMM B1B.2-11. c Angochitina pseudoavelinoi Gaugris and Grahn, 2006, Bed 27 GMM, B1B.2-12. d Lagenochitina avelinoi Lange, 1952, Bed 27, GMM B1B.2-13. e Lagenochitina avelinoi Lange, 1952, Bed 23, GMM B1B.2-14. f Oenonites sp., right MI in dorsal view, Bed 35, GMM B4A.8-1. g Oenonites sp., right MI in dorsal view, Bed 50, GMM B4A.8-2. h Oenonites sp. incomplete right MI in dorsal view, Bed 33, GMM B4A.8-3. i Oenonites sp., right MI in right lateral view, Bed 50, GMM B4A.8-4. j Oenonites sp., right MI in dorsal view, Bed 33, GMM B4A.8-5. k Skalenoprion sp., right MI in ventral view, Bed 50, GMM B4A.8-6. l Bipennatus bipennatus (Bischoff and Ziegler, 1957), Morphotype α sensu Bultynck (1987), Bed 23, GMM B9A.15-28. m Dvorakia chattertoni Klapper and Barrick, 1983, Bed 6, GMM B9A.15-8. n Belodella resima (Philip, 1965), Bed 6, GMM B9A.15-10. o Neopanderodus perlineatus Ziegler and Lindström, 1971, Bed 6, GMM B9A.15-11
The second conodont assemblage consists mostly of simple cone genera. We recognised Belodella resima (Fig. 7n, Bed 16 and Bed 20b), ?Neopanderodus sp. (Bed 5, a form with lenticular cross-section, only few costellae, and a short furrow on the obverse side), Neop. perlineatus (Fig. 7o, Bed 6, ?juvenile in Bed 21), Dv. chattertoni (Fig. 7m, beds 6, 20b, 22, and 25), Dv. klapperi (Bed 50, with more rotund cross-section than Dv. chattertoni), and Panderodus cf. unicostatus (?Bed 16, Bed 21). Prioniodinid elements occur in beds 6 and 9. The upper part of the section yielded no conodonts (after Bed 51; Fig. 4), which indicates a strong reduction of faunal influx from the open shelf.
If the long-lasting change from the siliciclastics of the Upper Honsel Formation to the first reefal limestones was mostly controlled by climatic change in the hinterland, not by local factors, it is likely that the base of the Hagen-Balve Formation was roughly isochronous along the lateral extent of the Hagen-Balve reef. Therefore, a comparison with conodont data from the Hagen-Hohenlimburg region in the west is indicated (Koch-Früchtl and Früchtl 1993). Samples from the Steltenberg Quarry yielded more icriodids and polygnathids than at Binolen; the first require partly revision (K. Narkiewicz, oral comm.). The local basal reef limestone (beds 40–23, ca. 10 m) was devoid of conodonts. In Bed 22, the entry of I. brevis suggests a position high in the lower Givetian, within the higher Po. timorensis Zone (previous lower part of the Lower Po. varcus Zone). The first I. difficilis from Bed 11, from ca. 4.5 m higher, may even indicate a basal middle Givetian age (see icriodid zonation of Bultynck 1987 and the review by Liao and Valenzuela-Ríos 2017). However, there is increasing evidence from Morocco, that close relatives of I. difficilis have a slightly lower, top lower Givetian range (see Becker et al. 2020, 2021c). The combined Binolen-Steltenberg data suggest to place the base of the Hagen-Balve Formation within the Po. timorensis Zone.
Near the top of our section, starting after Bed 58, there is an outcrop gap of a few metres due to extensive vegetation. This gap ends with a small cliff (“C-layers”, Figs. 2d, 4, beds 59–65), which contains the well-bedded succession of a first true biostrome (predominantly (auto)parabiostrome sensu Kershaw 1994). Apart from bulbous tabulate corals and stromatoporoids, some levels yielded dominant, in-situ, baffling reef-builders. The rugose coral Dendro. trigemme is very abundant, as typical for initial biostromes of the region (see Birenheide 1978; Koch-Früchtl and Früchtl 1993; Schröder 2005; Becker et al. 2016). Together with co-occurring specimens of Spinophyllum ex gr. spongiosum and Temnophyllum specimens (T. latum, T. astrictum), these corals are index species for the Grypophyllum-Spinophyllum-Dendrostella assemblage zone proposed by Schröder (2005) for the eastern Rhenish Massif. Based on conodonts, it was calibrated with the lower to middle Givetian Po. hemiansatus to Po. ansatus zones. Based on the enormous thickness (up to 1000 metres) of the subsequent, mostly lagoonal Asbeck Member, which includes middle Givetian stringocephalid beds throughout its main part (Schudack 1993), it is likely that the top of our section falls either still in the top lower Givetian or in the basal middle Givetian.
The top of the biostromal Binolen Member is not exposed but, after a short outcrop gap, typical lagoonal facies (see microfacies chapter) of a bioherm characterise beds 66–73; these are placed in the basal Asbeck Member of the Hagen-Balve Formation (Fig. 4).
Binolen micro- and macrofauna (Tab. 1)
The conodont fauna has been discussed above. Nearly all identified corals, stromatoporoids, chaetetids, calcimicrobes/calcareous algae, and bryozoans are known from Givetian sedimentary rocks of other parts of the Rhenish Massif and the Ardennes. The majority of them has been described by May (1992b, 1993a, 1993b, 1993c) from two slightly older, lower Givetian coral limestones, the Grünewiese Member of the Ihmert Formation and the Bredenbruch Member of the Lower Honsel Formation.
Calcimicrobes
The calcimicrobes, which include calcareous algae and forms of controversial systematic affinity, were only preliminarily identified but confirm an euphotic setting. There are similarities with taxa described by May (1992b) from the lower Givetian of the northwestern Sauerland. The fragments determined as Litanaia sp. possibly belong to Litanaia (Paralitanaia) baileuxensis (Fig. 8a). The occurrence of the genera Halysis (= Flabellia in May 1993b, described from the Grünewiese Member and Bredenbruch Member), Givetianella tsienii (Fig. 8b), Kamaena, and Resteignella suggests a maximum water depth of 90 m and, with high probability, a water depth of less than 50 m (compare Roux 1985 and May 1992a).

Givetian microfossils of the thin sections from Binolen. Scale bar: 1 mm except for c with 100 μm. a Litanaia sp., Bed 15. b Givetianella tsienii Mamet and Préat, 1982, Bed 14. c Calcisphere, Bed 15. d Cross-section of a juvenile euomphalid, loose at cliff
Foraminifers
Among the foraminifers, we were able to distinguish nine species among 791 specimens (Fig. 6d–l). Even though they are all agglutinated forms, five families are present in the residues at Binolen (systematics after Kaminski 2014), the Ammodiscidae (most common Tolypammina, Fig. 6d–f), Saccaminidae (very common Thurammina, Fig. 6h–i), Psammosphaeridae (moderately common Psammosphaera fusca, Fig. 6g), Lacustrinellidae (rare Webbinelloidea hemisphaerica, Fig. 6j–k), and Rhizamminidae (very rare, only two individuals of Rhizammina, Fig. 6l). All taxa belong to the morphogroups ED2 (ED2a and ED2b) and ED4 sensu Holcová and Slavík (2013). There are ten individuals, which are assigned to Tolypammina aff. tortuosa (Fig. 6f). These differ from typical specimens (Fig. 6e) by the agglutination of larger grains, being more branching, with small appendages, and by more angular edges than in the typical form. Notably, in two thin sections from the cliff, cross-sections of supposed individuals of the Eonodosarioidea were identified, but a detailed taxonomical assignment is not possible.
“Calcispheres”
Microfossils of unknown phylogenetic position are represented in thin sections by “calcispheres” (Calcitarcha of Versteegh et al. 2009), which are locally widespread (Fig. 8c).
Assumed chitinozoans
There are 108 microfossils that, in the absence of any other known similar Devonian microfossils, are interpreted as calcified chitinozoans, probably only their moulds. Their morphologies resemble two described species, Angochitina pseudoavelinoi and Lagenochitina avelinoi. The difference between both is that the first has extra spines on the body chamber (Gaugris and Grahn 2006). Unfortunately, the calcareous preservation at Binolen does not preserve such characteristics (compare Fig. 7a–e). We exclude affinities of our specimens with agglutinated foraminifer genera, such as Hyperammina, because they are clearly recrystallised and internally solid calcite, unlike as in the associated true agglutinated foraminifers, which are hollow and consist of glued silt grains. One wonders if it is possible to preserve only calcitic moulds of chitinozoans, and not their original organic tests, especially since chitinous scolecodonts are present in the same samples. The ambiguous nature of our microfossils is considered during interpretations, but the term Chitinozoa is used in the figures and tables.
Sponges (including stromatoporoids and chaetetids)
Representative macrofauna can be seen in the thin sections (Figs. 9, 10 and 11) or was collected from limestone slabs. For the stromatoporoids, the publications of Lecompte (1951, 1952), May (1993c, 2005), and Salerno (2008) are most important. Among them, three morphological groups can be distinguished: the thin-layered encrusting Clathrocoilona (e.g. Cl. (Cl.) solidula spissa, Figs. 9c, 10f, 11b, d), the thick-layered to bulbous Hermatostroma (Fig. 11e), Stromatoporella (Figs. 9c, e, 10f), Stromatopora (e.g. S. huepschii, Fig. 9c), and Gerronostromaria (Fig. 10e), and the branching Stachyodes (Fig. 11c, h) and Amphipora. The latter are typical fossils of the Hagen-Balve Reef Complex, especially within the lagoonal and back-reef deposits of the Asbeck Member (May 1987, 1988; Schudack 1993).

Microfacies at Binolen. Scale bar: 10 mm. a MF-1 Unfossiliferous siltstone, Bed 3, showing bioturbation overprinting small-scale cross-bedding and with local pyrite nests. b MF-5a Crinoidal coral-stromatoporoid float-rudstone, Bed 4, with a large Mesophyllum (Mesophyllum) vesiculosum (Goldfuss, 1826) (1), which is encrusted by an intergrowth of Clathrocoilona (Cl.) obliterata (Lecompte, 1951), Fistulipora sp. (2), and Thamnopora angusta Lecompte, 1939 (3) inside a crinoidal packstone matrix. c MF-5a Crinoidal coral-stromatoporoid float-rudstone, Bed 5, with Stromatoporella sp. (1), tubes of Syringopora expansa Maurer, 1885 (2), overgrown by Stromatopora huepschii Bargatzky, 1881 (3), which yields tubes of Syringopora hanshanensis Chow, 1980 (4), and thin layers of Clathrocoilona (Cl.) solidula spissa (Lecompte, 1951) (5) on top. d MF-3 Crinoidal and bioclastic packstone, Bed 9, a crinoid-dominated packstone with rare fragments of brachiopods and perhaps small debris of auloporid tabulate corals. e MF-5b Crinoidal coral-stromatoporoid float-rudstone, Bed 10, with a large Stromatoporella sp. (1), which includes tubes of Syringopora expansa Maurer, 1885 (2). Above ?Disphyllum sp. (3), with Aulopora (Mastopora) sp. (4) on top. f MF-4a Peloidal and bioclastic grain-pack-rudstone, Bed 11, large crinoid, coral, chaetetid, and gastropod debris, often with micrite envelopes, and clast size varies across the thin section. g MF-6 Coverstone/Bindstone, Bed 13, wackestone-packstone matrix with a sediment-binding layer of Pachytheca stellimicans Schlüter, 1885 (1). h MF-5b Crinoidal coral-stromatoporoid float-rudstone, Bed 15, large bioclasts in packstone matrix dominated by crinoids and additional small fragments of Thamnopora sp., Stachyodes sp., and brachiopods

Microfacies at Binolen. Scale bar: 10 mm except for c and h with 1 mm. a MF-2 Bioclastic wacke-packstone, Bed 17, micritic matrix with shell fragments from bivalves and crinoids. b MF-2 Bioclastic wacke-packstone, Bed 20a, detailed view of micritic matrix with shell fragments from bivalves and crinoids. c MF-2 Bioclastic wacke-packstone and MF-4b Peloidal and bioclastic grain-pack-rudstone, Bed 22, micritic matrix with large extraclast of MF-4b. d MF-5b Crinoidal coral-stromatoporoid float-rudstone, Bed 37, floatstone with Platyaxum (Egosiella) ex gr. clathratum (Steininger, 1831) (1), isolated fragments of tabulate corals, crinoids, brachiopods, and isolated pyrite cubes. e MF-7 Coral-stromatoporoid frame-rudstone, Bed 56, with Alveolites (Alv.) suborbicularis pseudorbicularis Iven, 1980 (1) dominating the thin section, the tabulate coral Platyaxum sp. (2) on the right, and Gerronostromaria sp. (3) between both. f MF-7 Coral-stromatoporoid frame-rudstone, Bed 57, with Alveolites (Alveolites) sp. (1) in the centre overgrown by both Stromatoporella sp. (2), with tubes of Syringopora expansa Maurer, 1885 (3), and Clathrocoilona (Cl.) solidula spissa (Lecompte, 1951) (4). g MF-8c Coral-stromatoporoid baffle-floatstone, loose near the cliff, with a colony of Scoliopora insueta Janet, 1972 (1) occuring together with Clathrocoilona (Cl.) sp. (2). h MF-8 Coral-stromatoporoid baffle-floatstone, loose at cliff, detailed view of Murchisonia sp. with two prominent spiral ribs

Microfacies at Binolen. Scale bar: 10 mm. a MF-8a Coral-stromatoporoid baffle-floatstone, base of cliff, with Dendrostella trigemme (Quenstedt, 1879) (1) and associated Stachyodes (St.) ex. gr. radiata Lecompte, 1952 (2) and Remesia sp. (3). b MF-7 Coral-stromatoporoid frame-rudstone, Bed 59, framestone with large rugose coral Dohmophyllum difficile (Wedekind, 1925) (1), incrusted by a partially untypical Clathrocoilona (Cl.) solidula spissa (Lecompte, 1951) (2); on the right side branches of Trachypora circulipora Kayser, 1879 (3). c MF-8d Coral-stromatoporoid baffle-floatstone, Bed 63, Alveolitella (1)-Stachyodes (2) floatstone including brachiopod remains and Remesia crispa (Schlüter, 1885) (3). d MF-4c Peloidal and bioclastic grain-pack-rudstone, Bed 64, with remains of indeterminate molluscs, auloporids, gastropods (1), as well as Stachyodes (St.) sp. (2) and Pachyfavosites polymorphus (Goldfuss, 1829) (3), which is incrusted by Clathrocoilona (Cl.) solidula spissa (Lecompte, 1951) (4). e MF-7 Coral-stromatoporoid frame-rudstone, Bed 65, stromatoporoid Hermatostroma sp. (1) growing on tabulate coral Pachyfavosites polymorphus (Goldfuss, 1829) (2). Dense growth-interruption bandings occur together with a ragged margin (3) and sediment inclusions (4), which are overgrown by enveloping margins. f MF-9 Fenestral, peloidal pack-bindstone, Bed 69, fenestral bindstone with peloids and Stromatactis structures. g MF-9 Fenestral, peloidal pack-bindstone, Bed 71, fenestral peloid packstone with rugose coral (1), St. (St.) ex gr. radiata Lecompte, 1952 (2), and Thamnopora sp. (3). h MF-8b Coral-stromatoporoid baffle-floatstone, Bed 72, Stachyodes (1) bafflestone with Dendrostella sp. (2), peloids, and small Stromatactis-structures
Next to stromatoporoids, hypercalcified sponges are also represented by one species of the chaetetid Pachytheca (Pachytheca stellimicans, Fig. 9g), which enters in the Sauerland succession in basal Eifelian biostrome facies (Avlar and May 1997).
Based on numerous isolated calcareous spicules, Heteractinida, a low diverse, distinctive class of Palaeozoic sponges, were common at Binolen. The samples are dominated by representatives of the order Octactinellida, which is known from Rhenish biostromes (e.g. Langer 1991; Nose et al. 2017) but still poorly documented. We have spicules that are six-rayed and planar (evenly spaced), without any distal or proximal rays (Fig. 6n). Such spicules were described by Kučera (1993) from the Lower Devonian of the Barrandian area, Czech Republic, as Octactine spicules type A (see pl. 1, fig. 5 in Kučera 1993). The spicules refered to Gondekia sp. resemble the two described Gondekia species, which, however, were previously not known from the Devonian of Europe (Fig. 6n, compare Rigby 1991). The Binolen specimens differ from G. hastula and G. lancifer by being distinctively smaller. Another representative of the Octactinellida is Ensiferites langeri Kozur, Mostler and Repetski, 2008. Its spicules show six planar rays, one big and long proximal ray, as well as trichotomously forked distal rays (Fig. 6m). Associated are polyactinellid sponges (possible Calcarea, see Mostler 1996). There are two taxa, Praephobetractinia eocambrica, with primary three-rayed spicules, two of which are parallel to each other, and Dvorcia cf. mira. Our representatives of the latter are assigned with a cf. because their characteristic lower rays converge but do not connect at the end, probably due to fragmentary preservation (Fig. 6p).
Calcareous monaxones occur in two different shapes: straight oxes (Fig. 6q) and curved amphioxes (Fig. 6r). They are rarely described as isolated spicules from the Devonian (e.g. Langer 1991). Rigby et al. (1979) noted that simplifed spicules may occur in the outer part of Ensiferites. However, since the abundances of our comparably too slender monaxones do not correlate well with those of the typical heteractinid spicules, we consider that they do not belong to the same original sponges. Hence, they were counted separately in Tab. 2.
Corals
Locally, rugose corals contribute to the reef construction. However, their importance and frequency are subordinate compared to the dominant tabulate corals and stromatoporoids. The determination of the rugose corals is based on Birenheide (1978), May (1993b), Coen-Aubert (2002), and Schröder (2005). Solitary rugose corals occur occasionally at Binolen and are represented by Dohmophyllum difficile (Fig. 11b), T. latum, T. astrictum, and Spinophyllum ex gr. spongiosum. More important as bafflers are the dendroid or phaceloid colonial corals Mesophyllum (Mesophyllum) vesiculosum (Figs. 6b–c, 9b; a rather large-sized species), Dendro. trigemme (Fig. 11a), ?Disphyllum sp. (Fig. 9e), and Spongophyllum immensum pusillum.
The determination of the tabulate corals is based on Lecompte (1939), Iven (1980), Birenheide (1985), and May (1993a, 2005). Tabulate corals as well as stromatoporoids are common and diverse in the section and of more or less comparable importance as frame builders. This is remarkable, because in the younger, lagoonal Asbeck Member, stromatoporoids dominate clearly and tabulate corals account for only about 20% of the reef-builders (compare May 1988).
Both, the rugose as well as the favositid tabulate corals Pachyfavosites (e.g. P. polymorphus, Fig. 11d, e), Thamnopora (e.g. Th. angusta, Fig. 9b), and Caliapora are clear indicators of fully marine, stenohaline conditions (see May 1988). The tabulate corals Thamnopora, Scoliopora, and partially also Roemerolites were able to trap sediment due to their upright and ramose branching colonies (‘medial growth strategy’ Scrutton 1998). However, Roemerolites brevis rhiphaeus followed mainly the ‘peripheral growth strategy’ of Scrutton (1998) by irregular and partially anastomosing branches growing along the sediment surface or by encrusting on reef builders (see also May 1993a). The corallites of Syringopora expansa lived as commensals in different stromatoporoids (May 1993a, 2005); in Binolen in Stromatoporella sp. (Figs. 9c, e, 10f).
Only one thin section shows the first proof for Sc. insueta in Europe (Fig. 10g). The species was so far only known from the Givetian (beds with Stringocephalus) of the Ural Mountains (Janet 1972). On the first glimpse, the colony resembles a Thamnopora species. However, many corallites have a more or less alveolitid shape, even in the central part of the branch. The calices are meandroid, the pores are frequent and well-developed. These characteristics indicate a species of Scoliopora. However, the more common species Sc. denticulata (Milne-Edwards and Haime, 1851) can be excluded due to a different shape of the corallites in the peripheral part and due to significantly smaller as well as more pronouncedly meandroid corallites.
Molluscs
Complete gastropods, bivalves, and cephalopods are rare in the Binolen section. Gastropods can be a common faunal element in the lagoonal main reef facies of the Hönne Valley Reef (Schudack 1993), but they occur only as juveniles at Binolen. Following the descriptions and discussions in Frýda and Manda (1997), Bandel and Frýda (1998), Nützel (2002), and Heidelberger and Koch (2005), two gastropod taxa could be classified in thin sections. One trochspiral specimen is assigned to Murchisonia due to the prominent two spiral ribs, which form the periphery for the concave-shaped selenizone (Fig. 10h). A cross-section through a planispirally coiled shell belongs probably to a juvenile euomphalid (Fig. 8d); it consists of the openly coiled whorls of the protoconch and initial teleoconch. A complete specimen of the megalodontid bivalve Eomegalodus was pictured from Binolen by Polenz (2008) as “Megalodon”; the precise location is not known. The new discosorid cephalopod genus Binoleniceras is described by Afhüppe and Becker (2022, this issue). Despite many years of fossil collecting by student groups, only the holotype is available.
Annelids
Scolecodonts from Binolen are mostly placed in the family Polychaetaspidae, a group with various lapidognath jaw elements. The small number of recognised species is caused by the fact that most jaw elements are only small fragments without any characteristics. The genus Oenonites was distinguishable based on the first maxilla (Fig. 7f–j). It is elongated, with a row of 10–15 denticles, which become less pronounced towards the posterior end. With the taxonomic assignment to Oenenites sp. we follow Eriksson (1997) and Tonarová et al. (2016). However, it should be noted that some authors regard Oenenites as a nomen dubium and assign such scolecodonts to Polychaetaspis (compare discussion in Szaniawski and Drygant 2014). In addition, some individuals can be assigned to Skalenoprion sp., a prionognathid representative with a long and bent hook (Fig. 7k). It is likely that other taxa occur at Binolen since many scolecodonts are only preserved as fragments, such as single denticles or anterior hooks with no identifiable characteristics. In total, there are 1110 specimens from Binolen.
Brachiopods
Brachiopods are often preserved as disarticulated and fragmentary remains, whereas articulated specimens are rare. As noted above, some specimens of the Givetian index brachiopod Stringocephalus were found (Fig. 6a). One specimen is assigned to Stringo. ex gr. burtini (Defrance, 1825; Fig. 12a–d), based on comparisons with Struve (1992) and Thormann and Weddige (2001). Besides signs of microborings, this specimen preserved concentric patterns of likely colour bandings (Fig. 12d), which are comparable to previously documented colour patterns in Stringocephalus (e.g. Blodgett et al. 1988). Fragmentary atrypids and other brachiopods have been noted in the field.

Small-sized Stringocephalus ex. gr. burtini (Defrance, 1825) with partially preserved concentric colour banding (indicated by black arrows), loose, GMM B5B.18-2. Scale bar: 20 mm. a ventral view. b dorsal view. c lateral view. d detailed view of concentric colour banding and microborings on dorsal site
Bryozoans
Remains of bryozoans are rare both in the thin sections and microfossil residues. There are a few incrusting colonies of Fistulipora sp. (Fig. 9b), some fragments of fenestellids, and some branching bryozoans in thin sections. In sample residues, there are ?Spinofenestella sp. (Fig. 13r) and rhabdomesid cryptostomatids (det. A. Ernst).

Givetian ostracods, echinoderms, bryozoans, and other microfossils from Binolen. a ?Evlanella sp., Bed 4, GMM B7B.6-1. b ?Hollinella sp., Bed 9, GMM B7B.6-2. c ?Podocopida, Bed 4, GMM B7B.6-3. d Glyphidocrinus sp., Bed 5, GMM B3A.2-1. e indeterminate crinoid columnal Morphotype 1, Bed 5, GMM B3A.2-2. f cupressocrinitid Morphotype A, Bed 8, GMM B3A.2-3. g indeterminate crinoid columnal Morphotype 2, Bed 9, GMM B3A.2-4. h indeterminate crinoid columnal Morphotype 3, Bed 12, GMM B3A.2-5. i cupressocrinitid Morphotype B, Bed 9, GMM B3A.2-6. j Eocaudina cf. patella Boczarowski, 2001, Bed 33, GMM B3A.8-1. k Eo. cf. patella Boczarowski, 2001, Bed 33, GMM B3A.8-2. l echinoid spine fragment, Bed 52, GMM B3A.8-3. m Eo. ovalis Matyja, Matyja, and Szulczewski, 1973, Bed 22, GMM B3A.8-4. n Eo. ovalis Matyja, Matyja and Szulczewski, 1973, Bed 24, GMM B3A.8-5. o Staurocaudina cf. dombrowiana Boczarowski, 2001, Bed 22, GMM B3A.8-6. p Ophiuroidea inc. sed., Bed 8, GMM B3C.1-1. q ?toothplate, Bed 4, GMM A1C.6-1. r ?Spinofenestella sp., Bed 9, GMM B5A.5-1. s juvenile gastropod, Bed 6, GMM B6B.12-1. t ?Onychodus sp., Bed 20b, GMM A1C.6-2. u indeterminate fish scale, Bed 22, A1C.6-3
Ostracods
The ostracods are preserved as internal, originally pyritic moulds. Therefore, only vague determinations were made. For example, the elongated shape with fine papillae on the ventral edge and a bump-like elevation on the straight side suggest an affiliation with the order Palaeocopida. Specimen GMM B7B.6-2 shows the typical nodes and wide flare of Hollinella (Fig. 13b). Other forms are compact and kidney-shaped, with a distinct pit for the closing muscle. These belong to the order Platycopida and are classified, with reservation, as ?Evlanella sp. (Fig. 13a, det. H. M. Weber). In yet other forms no clear characteristics can be discerned due to the poor preservation. However, their bulky, oval shape, and the differently-sized valves indicate an assignment to the Podocopida (Fig. 13c). A total amount of 64 ostracods was examined.
Echinoderms
The echinoderms from Binolen consist of members of the Asterozoa (ophiuroids), Echinozoa (holothuroids, echinoids), and Crinozoa (crinoids), with a total of 397 isolated sclerites found in the residues. Their increased numbers in individual samples correlate well with the recognition of echinoderm debris in thin sections (e.g. of MF-3 and MF-5).
The crinoids are represented by isolated stem elements (columnals). They are partly recrystallised by dolomitisation, which limits accurate assignments. Occassionally, columnals show the relatively well-preserved, porous stereom microstructure. Columnals of cupressocrinitid crinoids are common but their preservation is moderate in comparison with specimens from other Rhenish strata (e.g. Ebert 1994). They still can be easily recognised, based on their characteristic articular facet with rounded to slightly tetragonal shape and the axial canal (lumen), which is surrounded by four smaller canals (Fig. 13f, i). With regard to cupressocrinitid revisions based on calyces (e.g. Bohatý 2005), it is preferred to assign our specimens solely at the family level. Our specimens are assigned in open nomenclature as Morphotype A (rounded shape of articular facet) and Morphotype B (slightly tetragonal shape of articular facet). In comparison with similar columnals described by Gluchowski (1993) from the Holy Cross Mountains in Poland, some Binolen specimens are assigned to Glyphidocrinus (Fig. 13d). They show a relatively wide and convex epifacet, as well as the concave articular facet with a small lumen and short crenulae. Due to an overall poor preservation, the majority of columnals are referred to as indeterminate crinoid columnal morphotypes 1–3 (Fig. 13e, g–h).
Holothuroid sclerites are assigned to Eo. ovalis (Fig. 13m–n), Eo. cf. patella (Fig. 13j–k), and Staurocaudina cf. dombrowiana (Fig. 13o). The sclecrites of the latter are oval, with 3–4 central pores and probably 10 marginal pores. Finer pores at the marginal pores are not observed due to the bad preservation; therefore, a cf. identification is given (compare Boczarowski 2001). It has to be mentioned that Eo. cf. patella resembles the ophiocistioid genus Rotasaccus described from the “Schlade Event Layer” of the middle Büchel Formation at Bergisch Gladbach (compare Bohatý and Herbig 2010, Fig. 7a–f). In the residues, there is also an ophiuroid sclerite (Fig. 13p) and one echinoid spine fragment (Fig. 13l). A second small echinoid spine fragment was identified as a cross-section in a thin section from the cliff.
Microvertebrates
There is one microfossil that we regard as a small tooth plate (Fig. 13q). There are no shark teeth. Bony fishes are represented by rare teeth of probably juvenile sarcopterygians (?Onychodus sp., Fig. 13t) and an isolated fish scale (possibly Actinopterygii, compare Choo 2015; Fig. 13u). In general, Devonian bony fishes are wide-spread in neritic facies and known from the initial reef phase at Hohenlimburg (Koch-Früchtl and Früchtl 1993) but are not known from bioherm reef cores or lagoons.
Microfacies (MF)
Apart from MF-1 of the Upper Honsel Formation, all facies types are limestones or at least marlstones (Figs. 9, 10 and 11). MF-2–8 are mostly part of the Binolen Member of the Hagen-Balve Formation and represent two types of biostromal sets. The first set (MF-2 to MF-6) is represented by well-bedded shallow-water carbonates with some reef builders showing pronounced features of re-sedimentation processes (allo- to parabiostrome sensu Kershaw 1994). The second set (MF-7 and most of MF-8), developed in the cliff at the top of our section, represents an (auto)parabiostrome sensu Kershaw (1994) with both in situ and reworked reef organisms. Overlying thin-bedded peloidal, bioclastic, and fenestral floatstones (parts of MF-8b, MF-9) represent backreef facies of the basal Asbeck Member (compare Fig. 4).
Unfossiliferous siltstone (MF-1)
Examples: Beds 0 and 3 (Fig. 9a)
Description: Fine, middle brownish-grey, flaser-bedded, micaceous siltstones occur only at the base of the section (beds 0–3). They host small pyrite cubes/nests. Macroscopically and in thin sections, there are no body fossils, but intense bioturbation that overprinted small-scale cross-bedding. Fine, subhorizontal to oblique calcite veins result from late diagenesis.
Discussion: The fine grain size and traces of small-scale cross-bedding of the Upper Honsel Formation suggest deposition in a moderately deep, subtidal prodelta lobe rather far away from the coast. Larger cross-bedding sets can be observed in the folded area just south of the section base. The local absence of shelly neritic fauna suggests a muddy substrate that was rather hostile apart for the unknown burrowers. In his facies analysis of the Honsel Formation, Çinar (1978) described a similar lithology for his facies type F and explained the lack of fauna by strongly unstable depositional conditions, which are difficult to adapt to.
Adjacent localities of the formation yielded the byssate bivalve Ptychopteria, the spiriferid brachiopod Spinocyrtia (C.) sp. (possibly beckmanni), and other brachiopods (Jux 1960; May 1991). These are characteristic for the Spinocyrtia-Ptychopteria-community, which was contradictory regarded as an outer shelf and very shallow-water environment (Goldring and Langenstrassen 1979; Langenstrassen 1983). As shown by these authors, it is likely that the fluctuating deltaic system produced shallow shoals within the wide outer shelf.
The small-scaled nests with idiomorphic pyrite cubes were formed during early diagenesis (Fig. 9a). In marine (non-euxinic) sedimentary rocks, pyrite formation is limited mainly by the available organic matter (Berner 1984, 1985). The endobenthic substrate conditions were clearly oxygenated, due to the presence of intense bioturbation (Fig. 9a). The endobenthos may have enabled the incorporation of organic matter from the sediment-water interface into the substrate initiating micromilieus with reduced conditions for the pyrite formation (compare Berner 1985).
Bioclastic wacke-packstone (MF-2)
Examples: Beds 8 (partly), 16, 17 (Fig. 10a), 18a, 18b, 19, 20a (Fig. 10b), 21, 22 (Fig. 10c), 24, 25, 26, 27, 28, 29, 31, 33, 35 (partly), 39 (partly), 40 (partly), 41 (partly), 42 (partly), 45, 47 (partly), 49 (main part), 50, 51 (upper part), and 53
Description: The middle part of the section consists mostly of thin-bedded, middle- to dark-grey limestones characterised by small-sized bioclastic debris and micritic matrix. Bed 50 is especially rich in Corg. The fauna of thin sections comprises crinoid debris, shell fragments (mostly thick-shelled brachiopods, some thin-shelled bivalve fragments), gastropods, small fragments of tabulate corals (e.g. Bed 25) and stromatoporoids (e.g. Clathrocoilona), ostracods (e.g. in beds 18 and 19), and occasional sponge spicules. There are fluctuations between matrix- or clast-supported lithology, sometimes within beds (Bed 22, Fig. 10c). Bioturbation occurs frequently, sometimes resulting in a cloudy appearance. In other beds, the primary layering is partly still visible. In a small amount of reworked bioclasts, the margins were micritized before re-deposition (cortoids). As a special feature, only the middle of Bed 53 displays thin-shelled circular cross-sections, which potentially represent orthoconic cephalopods, partly with inclined geopetal filling. There is no evidence for graded bedding or cross-bedding. MF-2 grades into MF-3 (parts of beds 39–42), and by the increase of larger bioclasts (e.g. of tabulate and rugose corals of Bed 35, of stromatoporoids in Bed 47), into MF-5b.
Discussion: The relatively high content of fine bioclastic debris speaks for allochthonous or parautochthonous deposits (an allobiostrome sensu Kershaw 1994), but there is no evidence for turbidite transport. Pelagic fauna is missing, except for the potential orthoconic cephalopods. The micrite has not been washed out, indicating weak or a lack of bottom currents, but the subordinate larger bioclasts must have been washed in by episodic slightly stronger currents. The setting of MF-2 was subtidal and well below the fair-weather wave base. In the contemporaneous initial reef phase of Hagen-Hohenlimburg, Koch-Früchtl and Früchtl (1993) noted a much higher siliciclastic content in their MF-Typ 1. This suggests a more gradual transition from the Upper Honsel to the basal Hagen-Balve Formation in the west.
Crinoidal and bioclastic packstone (MF-3)
Examples: Beds 9 (Fig. 9d), 10 (partly), 12, 14 (partly), 15 (partly), 38 (partly), 39 (partly), 40 (partly), 41 (partly), and 42 (partly)
Description: MF-3 consists of middle-grey limestones that differ from MF-2 by a higher amount of larger bioclasts, especially by dominant and partly larger-sized crinoid debris, associated with variable amounts of angular to subrounded bioclasts consisting of fragmentary brachiopods, bryozoans, calcareous algae (Litanaia, ?Girvanella, Givetianella, ?Resteignella, Halysis), stromatoporoids (Clathrocoilona), and tabulate corals (Scoliopora), giving a heterogeneous packstone fabric (Fig. 9d). Large gastropods and rugose corals (Mesophyllum, Bed 12, Fig. 6b–c) occur in transition towards floatstones of MF-5b (e.g. beds 14, 38). The matrix is made of fine debris and micrite, but the latter has partly been washed out in small pockets. There is no sorting or grading of bioclasts. Peloids occur in a few beds (e.g. Bed 49).
Discussion: The lack of grading and cross-bedding rule out short-time deposition on a marginal reef slope, as typical for the Garbeck Member of the Balve region (Eder 1971) or in other Rhenish fore-reef settings (e.g. Stritzke 1990; May 1997). Crinoids of MF-3 are solely preserved as isolated columnals or their fragments, which is a result of postmortem disarticulation under agitated water conditions. The porous stereom microstructure of the columnals enhances transportation due to the reduced skeletal density. Nevertheless, the timing of postmortem decay of organic matter (mesodermal stroma tissue) and possible secondary filling of pore spaces by early diagenetic sparite limited the transport distance (see Savarese et al. 1997). MF-3 is seen as remains of crinoid meadows (see also May 1997) growing on the initial, shallow-water carbonate platforms (e.g. Préat and Mamet 1989; Préat et al. 2007) in the neighbourhood of reef builders under a moderate influence of waves and currents. In the biostromes of the Eifel Mountains, Faber (1980) described similar “arenitic crinoid debris” and “Crinoiden-Rasenfazies”, also pointing out a moderate water agitation in shallow-water.
Peloidal and bioclastic grain-pack-rudstone (MF-4)
Examples: MF-4a: Bed 11 (Fig. 9f); MF-4b: within Bed 22 (Fig. 10c) and 38; MF-4c: Bed 64 (Fig. 11d)
Description: There are three different types of peloidal microfacies without microbial features.
MF-4a, peloidal grain-rudstone, is similar to MF-3 but lacks a micritic matrix, which is replaced by small peloids and orthosparite. Common bioclasts are fragmentary crinoids, brachiopod debris, gastropods (including possible murchisoniids), corals (Caliapora), stromatoporoids (Stachyodes), and chaetetids (Pachytheca). Their size ranges between 0.5 mm and 8 mm, resulting in a variation between grain- and rudstone; the latter gives a transition towards MF-5. Many of the bioclasts represent cortoids with thin micrite envelopes (Fig. 9f). There is gradation towards MF-3.
MF-4b, poorly fossiliferous peloidal grainstone, occurs as an exception within MF-2 of Bed 22 (Fig. 10c) and is represented by a ca. 3.5 cm extraclast. It contains some gastropods and parathuramminids. Thin layers of poorly fossiliferous peloidal grainstones are also intercalated in Bed 38.
MF-4c, peloidal and bioclastic pack-grainstone, is restricted to a thin interbed of the upper biostrome (Bed 64, Fig. 11d). Apart from the dark, rather small-sized peloids, it is characterised by abundant fragmentary molluscs (probably gastropod remains), brachiopods, Stachyodes branches, only small amount of crinoid debris, parathuramminids, and some isolated larger tabulate corals (Pachyfavosites) encrusted by Clathrocoilona.
Discussion: The bioclasts in MF-4a are not well-rounded ruling out a long transport. However, their coating implies a long interval of exposure to endolithic microborers and marginal micritization before final re-sedimentation in a moderately agitated environment, which reworked or washed out the micrite. The lack of sorting and grading excludes a single storm event. The similar SMF Type 11 of Flügel (2010) formed under recurrent wave activity within reef platforms or winnowed fore-reef areas. At Binolen, such a setting is supported by the abundant reef debris and high content of isolated crinoid columnals in MF-4a.
MF-4b represents a less turbulent variant, where bottom currents were still sufficient to rework micrite as pseudopeloids (compare SMF 16 of Flügel 2010).
MF-4c resembles the reef debris facies described by Faber (1980) from fore-reef settings of the Eifel Mountains, in agreement with the intercalation at Binolen into the biostrome at the top.
In the western part of the Hagen-Balve Reef, Koch-Früchtl and Früchtl (1993) included locally common MF-4 variants in their MF-types 3 (ca. MF-4a) and 4 (ca. MF-4b). Since both yielded more varied conodont and microvertebrate faunas in several beds, their setting was probably slightly deeper or more open to the outer shelf than at Binolen.
Crinoidal coral-stromatoporoid float-rudstone (MF-5)
Examples: MF-5a: Beds 4 (Fig. 9b), 5 (Fig. 9c), 6, 7, and 43 (partly); MF-5b: Beds 8 (partly), 10 (partly), Fig. 9e), 14 (partly), 15 (partly, Fig. 9h), 35 (partly), 37 (Fig. 10d), 38 (partly), 43 (partly), 47 (partly), 49 (top), and 51 (lower part)
Description: There are two subtypes.
MF-5a, rudstones with angular fragments and subordinate complete specimens of variably sized reef builders, such as medium to large-sized rugose corals (Mesophyllum, Dohmophyllum, Spinophyllum), branching (e.g. Thamnopora), bulbous (Pachyfavosites), and laminar to cylindrical (Platyaxum (Roseoporella), Squameoalveolites, Alveolites (Alveolitella), and Alv. (Tetralites)) tabulate corals, chaetetids (Pachytheca), incrusting bryozoans (Fistulipora), calcareous algae (Rothpletzella), or stromatoporoids (Stromatopora, Stromatoporella, Clathrocoilona, Stachyodes), associated brachiopods (stringocephalids), and small-sized gastropods. The matrix represents crinoidal and bioclastic packstones grading laterally into crinoidal grainstone (MF-4), locally with intercalations of dark, micritic mud-wackestone. There is no sorting or grading. The crinoid and fine coral debris is mostly angular. Subordinate micrite envelopes occur.
MF-5b, middle-grey floatstones with reefal debris, including fragmentary rugose (?Disphyllum) and tabulate corals (Thamnopora, Platyaxum (Egosiella)), other favositids, Roemerolites, encrusting auloporids), stromatoporoids (Clathrocoilona, Stachyodes, Idiostroma, rare Amphipora), bryozoans, calcareous algae, as well as associated gastropods (Bed 8), embedded in a crinoidal and bioclastic wackestone-packstone matrix. MF-5b grades laterally and vertically into MF-5a (Bed 43), MF-3 and MF-4 (e.g. in beds 14 and 38). Abrupt facies boundaries within some beds (e.g. Bed 15) were caused by strong bioturbation.
Discussion: The fragmentation of reef builders and dwellers was caused by wave/storm action on a shallow platform but there is no evidence for long transport or deposition by episodic tempestites. Water agitation must have been rather variable to explain the close intercalation of coarse and fine debris, with MF-5b having been deposited under slightly calmer conditions. This is also supported by a more subordinate occurrence of thin micrite envelopes on a few bioclasts in MF-5a, resulting from just marginal micritization prior to reworking. Debris flow deposition could be considered but is unlikely in the absence of a proven slope. Equivalents of MF-5, with a rather diverse, mixed assemblage of reef builders and neritic elements, were assigned in the western part of the Hagen-Balve Reef by Koch-Früchtl and Früchtl (1993) to their MF-Typ 2. Krebs (1974) and Machel and Hunter (1994) recognised a widespread, detrital stromatoporoid-crinoid facies at the transition from the reef core to the fore-reef of bioherms. A higher abundance of corals, as at Binolen, is known from Givetian biostromes of the Moroccan Meseta (Eichholt and Becker 2016).
Coverstone/Bindstone (MF-6)
Examples: Bed 13 (Fig. 9g)
Description: In one thin section, crinoidal debris of MF-3 was covered by thin layers of laminar growing reef builders, such as chaetetids (Pachytheca), tabulate corals (Platyaxum (Roseoporella)), or stromatoporoids (Clathrocoilona), which partly grew on each other.
Discussion: The term “coverstone” was introduced by Tsien (1982, 1984) for carbonate rocks, which are dominated by in situ tabular or lamellar organisms that cover and stabilise broken debris. Comparable microfacies were described by Hubmann (1993) and Préat et al. (2007). MF-6 is known from the lower Givetian of the northwestern Sauerland (May 1992a, 1993c), where it plays a significant role in the substrate colonisation of reefal pionier stages. The growth of coverstones requires a localised, episodic interruption of debris shedding within the otherwise agitated setting of MF-2 and MF-3.
Coral-stromatoporoid frame-rudstone (MF-7)
Examples: Beds 56 (Fig. 10e), 57 (Fig. 10f), 59 (Fig. 11b), and 65 (Fig. 11e); loose samples from below the cliff
Description: MF-7 occurs in the cliff at the top of the section and is characterised by a variably dense packing of reef builders, which are at least partly preserved in situ. In comparison to MF-5, there is a lack or rarity of crinoid remains. Faunal elements are varied and include stromatoporoids (Gerronostromaria, Stromatoporella, Clathrocoilona, Trachypora, Hermatostroma, Stachyodes (Stachyodes)), rugose corals (Spongophyllum, Dohmophyllum), tabulate corals (Alv. (Alveolites), Alv. (Alveolitella), Platyaxum (Egosiella), Pachyfavosites), bryozoans, and chaetetids. At the top of the cliff (Bed 65), stringocephalids re-appear. Micrite envelopes are common. The matrix consists variably of dark marl and micrite, bioclastic wackestone, or organic-rich, dark, peloidal pack-grainstone (coral rudstone of Bed 59, Fig. 11b).
Discussion: MF-7 represents an (auto)parabiostrome (Kershaw 1994) that grew on a shallow subtidal, wave and current influenced platform, where occasional storms contributed to the destruction of frame builders. The variable preservation of fine detritus documents fluctuating turbulence. Supporting evidence is given by a specimen of the bulbous stromatoporoid Hermatostroma, which shows, next to a single ragged margin, occasional sediment inclusions (Fig. 11e). These patterns are mostly connected to growth-interruption bandings and speak for incipient growth cessation due to episodic changes in water turbulence with higher sediment accumulation rates or re-sedimentation processes (e.g. Kershaw 1984; Young and Kershaw 2005; Webby and Kershaw 2011). However, this specimen shows an upright growth strategy with enveloping margins formed by new latilaminae (Webby and Kershaw 2011), and, therefore, show an overgrowth of previously accumulated sediment particles.
The diversity of reef builders is typical for an undifferentiated biostrome where the palaeobathymetry has not yet caused a strong palaeoecological zonation. Faber (1980) recognised a corresponding “initial facies” in Eifel reefs, which is also known from the Ardennes (Lecompte 1970). Further equivalents were observed in the Givetian of the Moroccan Meseta as MF C2c (Eichholt and Becker 2016).
Coral-stromatoporoid baffle-floatstone (MF-1)
Examples: MF-8a: sample “base of cliff” (Fig. 11a), beds 66, 67; MF-8b: Beds 60, 61, 62, and 72 (Fig. 11h); MF-8c: loose sample from near the cliff (Fig. 10g); MF-8d: Bed 63 (Fig. 11c)
Description: Macroscopically, MF-8 occurs as middle- to dark-grey, argillaceous limestones with abundant branching reef builders that are either still in situ or broken but not aligned by currents, therefore, (par)autochthonous. Based on the dominant taxa, several subfacies can be separated, which may grade into each other:
MF-8a, Dendrostella baffle-floatstones, consist of the fasciculate (loosely dendroid to phaceloid) branches of the rugose coral Dendrostella (see Becker et al. 2016), partly associated with Stachyodes and the auloporid coral Remesia (sample “base of cliff”). The matrix is marly limestone or bioclastic wackestone. As a variant of MF-8a, beds 66 and 67 are characterised by a flaser-bedded peloidal pack-grainstone matrix, in which Dendrostella, Thamnopora, and Stachyodes branches, as well as stromatoporoid fragments (Hermatostroma), are floating. It is intermediate towards MF-4c.
MF-8b, Stachyodes bafflestones, are dark, argillaceous, and/or organic-rich and characterised by Stachyodes branches that are partly encrusted by Clathroilona and associated with Dendrostella and Remesia. The matrix is mud-wackestone with some ostracods and a small amount of fine crinoid debris. Fenestral fabrics (e.g. Stromatactis structures) are common in MF-8b (Fig. 11h) and, therefore, show transitions towards MF-9 (see below).
MF-8c, Scoliopora bafflestones, are characterised by the branching tabulate coral Scoliopora, which is partly overgrown by Clathrocoilona, and associated with algal crusts and the tabulate coral Platyaxum (Egosiella). The matrix consists of mostly dark, organic-rich mud-wackestones with some gastropods, brachiopod fragments, and a minor amount of fine crinoid debris.
MF-8d, Alveolitella-Stachyodes baffle-floatstones, show branches of Stachyodes, Alv. (Alveolitella), and Remesia jointly with stringocephalid fragments in a moderately organic-rich wackestone matrix. The slightly higher amount of crinoid debris indicates a transition towards MF-5b, but the faunal elements are different.
Discussion: MF-8 alternate with MF-7 within the autoparabiostromal section at the top cliff of Binolen. It reflects phases of calmer condition, when more delicate, branching reef builders baffled fine, partly argillaceous and organic-rich detritus. Occasional storms broke many branches but there was no wide transport so that it is partly possible to assign branches to an original colony. Dendrostella is one of the few genera of loosely dendroid to phaceloid rugose corals that may form distinctive assemblages of Givetian reefs on a global scale and that may contribute to carbonate accumulation as a baffler (e.g. Pedder 1964; Oliver et al. 1975; Zhen 1996; Schröder 2005; Becker et al. 2016).
According to Krebs (1974), Stachyodes preferred niches between more robust reef builders and extended from the inner reef core into the back-reef of bioherms (May 1987; Malmsheimer et al. 1991; Schudack 1993; Machel and Hunter 1994: Zone IVb), where it tends to be associated with thamnoporids (Eichholt and Becker 2016). Stachyodes formed also distinctive meadows in reefs of the Eifel Mountains (Faber 1980, facies 3.13) and the Ardennes (Lecompte 1970). Turbulence was moderate. In the Binolen context, the increasing abundance of Stachyodes in MF-8b and in intermediate facies towards MF-4c and MF-9 may be seen as an indicator for the beginning morphological differentiation of the carbonate platform.
Scoliopora is widely distributed but normally not a dominant element of branching coral assemblages in the Rhenish Massif (e.g. Stadelmaier et al. 2005). However, in the restricted, lagoonal parts of the Rhenish reefs, Scoliopora can be common and be the dominant tabulate coral (May 1987, 1988).
The sectioned loose slab probably represents a very localised feature within the biostrome. The same may apply to MF-8d but a local dominance of Alv. (Alveolitella) was described by Stadelmaier et al. (2005) from Sötenich in the Eifel Mountains, with associated subordinate Scoliopora.
Fenestral, peloidal pack-bindstone (MF-1)
Examples: Beds 68, 69 (Fig. 11f), 70, 71 (Fig. 11g), and 73
Description: In the cliff above the main section, several beds of middle-grey limestones are characterised by variably sized and abundant, irregular sparitic fenestrae surrounded by peloidal pack- or grainstones. The bioclastic content is low to moderate, with some floating branches of Stachyodes, Amphipora (only Bed 70), and Thamnopora, fragmentary rugose corals (Bed 71, Fig. 11g), or thick-shelled brachiopods. The size of the fenestrae or birdseyes (Ham 1952) varies roughly between 0.5 and 2.5 mm. Their shape is irregular to elongate and agrees with the LF-B type of Mestermann (1995). Only in the upper part of Bed 69, they are aligned horizontally, forming LF-A type patterns (Mestermann 1995) or small-sized Stromatactis layers with planar base, giving a bindstone fabric (see Read 1975).
Discussion: The formation of peloids in MF-9 is most likely a result of micrite reworking due to permanent low-velocity bottom currents (Fåhræus et al. 1974; Flügel 2010). A biotic origin of peloids (e.g. gastropod fecal pellets) is unlikely or at least of subordinate significance, due to the lack of known gastropod remains in MF-9. The combination of peloids and fenestral fabrics is common in inter- to supratidal settings of back-reef environments, where early cementation, meteoric water influx, and shrinking processes affected bacterial mats (e.g. Shinn 1983). However, a higher amount of bioclasts, especially of corals and stromatoporoids, as at Binolen, resembles the “fenestrae-like features in subtidal grainstones” of Shinn (1983) and supports a shallow subtidal origin, as suggested by Hladil (2005). In any case, the onset of MF-9 shows that the Hönne Valley Reef has left the open biostromal phase and that lagoonal conditions, implying a protecting lateral reef core, were established.
Related facies have been described from Hagen-Hohenlimburg by Koch-Früchtl and Früchtl (1993) and from the Brilon Reef (May 1987; Mestermann 1995). They fall in SMF 21 sensu Flügel (2010). Krebs (1974) described a fenestral facies from the Langenaubach Reef Complex, representing an intertidal palaeoenvironment with slightly increased salinity. The latter aspect is unlikely at Binolen due to the presence of stenohaline reef builders. Read (1975) identified in Devonian tidal-flat carbonates a sequence of different “cryptalgal” and fenestral fabrics depending on the position within the tidal-flat.
Conodont Biofacies
The Middle Devonian conodont biofacies model is still incomplete and based on only a few studies. Following the separation of more shallow, turbulent, and peri-reefal Icriodus Biofacies from the deeper, more offshore and calmer Polygnathus (s.l.) Biofacies by Weddige and Ziegler (1976), only two studies dealt with more detailed conodont distributions in relation to facies and palaeobathymetry. Sparling (1984) studied Eifelian assemblages of Ohio and recognised faunas with abundant Prioniodina and co-occurrences of icriodids and single cone genera, such as Belodella, Coelocerodontus, and Panderodus. Narkiewicz et al. (2016) reconstructed species-level distributions in the top middle Givetian of Poland and noted important differences in the Polygnathus ansatus and Linguipolygnathus facies ranges. Based on our data and by comparison with previous records from the literature, we propose two new Givetian conodont biofacies types for biostromal settings, where conodont abundances are notoriously very low. Since only specialised, mostly long-ranging taxa lived in small populations in reefal settings, it is not possible to require the usual (> 20, better > 50) specimen numbers to identify biofacies types. Therefore, definitions have to be robust and simple; transitions are common. Due to the low specimen numbers, conodont biofacies assignments of beds at Binolen are statistically very weak.
Bipennatus Biofacies
Examples: Beds 4, 18, 23, and 29
Definition: Conodont faunas consisting only of rare Bipennatus specimens or the genus makes up more than 50% of specimen-poor to median rich assemblages.
Discussion: The new biofacies is introduced for biostromal, coral- or stromatoporoid-bearing strata characterised by the name-giving index genus, which is locally either the only conodont present or associated with rare icriodids or simple cone forms. In principle, it was recognised by Bischoff and Ziegler (1957), who found that Bi. bipennatus was restricted in the Givetian of the Sauerland to the biostromal Sparganophyllum Limestone. Co-occurrences with Linguipolygnathus and Icriodus (Ziegler 1965) suggest gradual transitions towards polygnathid and icriodid biofacies. For example, both pure Bipennatus and mixed Bipennatus-Polygnathus-Icriodus faunas are known from biostromes intercalated within the Lower Honsel Formation of the Hohenlimburg region (Ziegler 1965; v. Kamp 1972). The Bipennatus Biofacies is also known from the shallow-water Eifelian Wojciechowice Formation of Poland (Narkiewicz and Narkiewicz 2010: with 91 % Bipennatus of 33 Pa elements), from the biostromal Abîme Member of the Couvin Formation in the Ardennes (Bultynck in Narkiewicz and Narkiewicz 2010), from the Eifelian Hobräck Formation of the Sauerland (“Basiskalk”) and the lower Givetian of the Eifel Mountains (see charts in Weddige 1977 and Weddige 1988, p. 103: occurrences of Eognathodus = Bipennatus alone). In South China, a sample from the neritic Beiliu Formation of the Nalai section (Lu and Königshof 2019) showed that Bipennatus can be the far dominant genus (almost 90 % of all Pa elements) in a moderately rich assemblage with 75 specimens.
The genus Bipennatus is characterised by a differentiated apparatus with massive P1 (= Pa) elements that served for food processing.
Simple Cone Biofacies
Examples: Beds 5–6, 9, 16, 20–22, and 25
Definition: Combined simple cone specimens outnumber the sum of all associated genera, such as polygnathids, icriodids, spathognathodids, or prioniodinids.
Discussion: The new biofacies is characterised at Binolen by the almost exclusive occurrence of low numbers of coniform genera, notably of Dvorakia, Panderodus, and Neopanderodus, and with subordinate Belodella. Such faunas are also known from reefal facies of the Eifel Mountains (Weddige 1988, p. 103) and shallow-water settings of Queensland (Mawson and Talent 1989). Since Belodella may be more common than other simple cones in some carbonate platforms (e.g. initial reef phase at Hagen-Hohenlimburg, Koch-Früchtl and Früchtl 1993; Oued Cherrat region of Morocco, Becker et al. 2020), extending to non-reefal, open settings, it may become useful to establish subfacies types in future. For example, in the Lower Devonian, Belodella can be far dominant in assemblages with hundreds of specimens (e.g. Aboussalam et al. 2015). Givetian faunas dominated either by Belodella or Neopanderodus (e.g. from the Po. timorensis Zone of the Moroccan Meseta, Becker et al. 2021b, his tab. 2) tend to be specimen-poor. At Binolen, Belodella, Dvorakia, or Neopanderodus have not been found in association with Bipennatus but there are co-occurrences in Australia (Mawson and Talent 1989). Therefore, an intergradation of the Bipennatus and Simple Cone biofacies is expected and probably a matter of further sampling.
Simple cone genera are characterised by relative uniform apparatuses that are suitable for grasping and active hunting of larger prey (Murdock and Smith 2021), possibly with the help of venom in the case of taxa (Panderodus, Neopanderodus) with a distinctive longitudinal furrow on the elongate teeth (Szaniawski 2009).
Other microfossil biofacies (BF)
Palaeozoic microfossils were often taken as a biofacies proxy in order to evaluate patterns of palaeobathymetry or palaeoecology. Biofacies analysis has been successfully used in foraminifers (e.g. Gutschick and Sandberg 1983) and conodonts (e.g. Clark 1984; Lüddecke et al. 2017; Söte et al. 2017; see above). However, most approaches were restricted to just one fossil group. A holistic approach to successive microfossil assemblages from a Devonian reef succession has not been attempted before. It has to consider aspects of palaeoecology, sample processing (see methods), taphonomy, diagenesis, and possibly variable transport. The lack of sorting and bioclasts rounding at Binolen suggest parautochthonous deposition throughout the section.
Apart from the new conodont biofacies, we distinguish empirically six local microfossil assemblages or biofacies types (BF types). All are based on a quantitative approach by counting the total abundance of each individual fossil group per bed and by calculating their relative proportions (Figs. 14, 15). More detailed data are given in Tab. 2. We considered beds with less than 30 individuals to be representative in a limited way. This occurs five times, in beds 11, 18b, 19, 46 and 49. In all fossil poor beds, the first group that disappears are the echinoderms, but Bed 19 is a sample where calcareous spicules are more common than non-calcareous groups, which speaks against a simple processing bias. Only the scolecodonts are present in all five beds while the microfacies varies strongly.

Vertical fluctuations of absolute abundances of microfossil groups at Binolen, excluding the rare conodonts, showing significant changes in assemblages

Vertical changes in microfossil assemblages at Binolen, enabling the distinction of six microfossil facies types: O Ostracoda Biofacies, F Foraminifera Biofacies, PE Porifera-Echinodermata Biofacies, PS Porifera-Scolecodont Biofacies, C Chitinozoa Biofacies, S Scolecodont Biofacies. Shaded intervals, with bed numbers in brackets, indicate levels with less than 30 individuals, where the biofacies assignment becomes less reliable
Ostracoda Biofacies (MF-1)
Examples: Beds 4, 19, and 20a
Definition: Peak abundances of small-sized, originally pyritic ostracod moulds, recorded by microfossil assemblages with >15% ostracods, variably in faunas with dominant foraminifers (Bed 4) or sponge spicules (beds 19–20).
Discussion: Benthic ostracods occur in small numbers throughout Middle Devonian reef complexes but are much more diverse and abundant in the neritic, subphotic “Eifel Ecotype”. This was confirmed by quantitative diversity analyses of a transect from lagoonal to brachiopod-rich basal marginal slope settings by G. Becker (1969, 1971). A strong restriction of ostracods in Givetian reef platform facies is also known from the Ardennes (e.g. Maillet et al. 2016). Therefore, the BF-O biofacies could indicate an open, moderately deep neritic setting but the taphonomy indicates in addition poor ventilation within the sediment. Partial oxygen deficiency was probably caused by the bacterial degradation of the strongly increased influx of organic matter. In one case (Bed 19) an increased content of ostracods is visible in the thin section.
Foraminifera Biofacies (MF-1)
Examples: Beds 5–6, 59, possibly beds 11 and 46 with only small microfaunas
Definition: Microfaunas with an amount of >50% of agglutinating foraminifers. Associated are either sponge spicules (beds 5–6) and echinoderms, or scolecodonts (Bed 59).
Discussion: Apart from some potential specimens of the Eonodosarioidea in two thin sections, multi-chambered Givetian calcareous forms, such as Nanicella, have not been found; they occur frequently in the final reef stage (Stichling et al., 2022, this issue). The locally dominant agglutinated forms are part of the Saccaminid Biofacies sensu Gutschick and Sandberg (1983), which was thought to characterise the “upper foreslope” at the shelf margin. This is certainly not the facies setting at Binolen. In general, there is a very restricted knowledge of benthic, non-calcareous foraminifers from Devonian reef facies, which glued exclusively non-calcareous particles during life. Langer (in Koch-Früchtl and Früchtl 1993) reported only Kettnerammina and Saccorhina from the initial reef facies of Hohenlimburg. Both genera do not occur at Binolen. In this respect, our assemblages from Binolen appear to be rather exceptional. Tolypammina, Thurammina, and Psammosphaera are well-known from Lower to Upper Devonian subphotic, pelagic ramp and seamount facies with low sedimentation rates (e.g. Eickhoff 1973; Holcová 2004; Saupe 2018). The latter was probably a decisive palaeoecological factor. Aboussalam et al. (2021) mentioned a small assemblage of Psammosphaera and Tolypammina from condensed, cherty, deep neritic facies of the southwestern Moroccan Meseta. Webbinelloidea had its palaeoecological maximum in neritic platform facies (Gutschick and Sandberg 1983), which is supported by records from the Eifel Mountains (Langer 1991). However, it ranged also into condensed cephalopod facies (Eickhoff 1973: Famennian; Aboussalam and Becker 2001: Givetian). The locally rare genus Rhizammina has been described together with Psammosphaera and Sorosphaera from Lower Devonian neritic crinoidal limestones of the Carnic Alps (Bandel 1969; Schönlaub and Flajs 1975). In modern oceans, Rhizammina ranges into deep oceanic facies (Kaminski et al. 1988).
In summary, BF-F indicates a strong open shelf influence and lowered sedimentation rates. If there is no fine matrix in thin sections, faunas may have been transported.
Scolecodont Biofacies (MF-1)
Examples: Beds 18b, 54, 56, and small sample from Bed 49
Definition: Microfaunas with >70% scolecodonts.
Discussion: Microfossil assemblages dominated very strongly by scolecodonts are rare at Binolen even though they are abundant in general.
The locally dominant genus Oenonites was described by Hinde (1879) from Upper Ordovician shales and intervening limestone beds of Toronto, Silurian shales and sandstones of Ontario, and the neritic, argillaceous Middle Devonian Hamilton Group (compare Eller 1941). Further occurrences were recorded from bluish-grey calcareous shales or mudstones between beds of limestones in the Silurian of west England (Hinde 1880). This suggests a preference to muddy mixed siliciclastic-calcareous shelf settings, which is supported by faunas from the upper Ludlow of Bohemia (Kopanina Formation, Tonarová et al. 2012), and, as for Polychaetaspis, from the Upper Ordovician of Gotland (Van Hinte et al. 1995). In the latter, Oenonites increases both in terms of abundance and species diversity with the decrease of water energy. It was also noticed that “there seems to be a close relationship regarding the abundances of polychaetes and chitinozoans” (Eriksson 1997). Hints and Eriksson (2007) described Oenonites as a widespread and long-ranging genus since the Middle Ordovician and Skalenoprion as less common, predominantly known from Baltica occurring only since the Upper Ordovician; both genera occur preferentially in shallow shelf settings.
In eastern North America, polychaetaspids are widely distributed in Middle Devonian neritic limestones of Iowa, Ohio, Kentucky, Michigan, and New York State (e.g. Eller 1938, 1941, 1963, 1964). However, more recent Devonian data are restricted. Szaniawski and Drygant (2014) described Oenonites and Skalenoprion from conodont-rich lower Lochkovian deposits of Podolia (Ukraine). For the Middle Devonian of the Eifel Mountains, Tonarová et al. (2016) showed that Oenonites was much more common than Skalenoprion in an upper ramp position with “moderate water depth just below the fair-weather wave base” (Königshof et al. 2015). The fragmentation and abundance of scolecodonts corresponds with the water-energy level. Strongly bioturbated bioclastic wackestones show a reduced number of more fragmented specimens (Tonarová et al. 2016). In the top-Emsian to lower Eifelian Plabutsch Formation of the Graz Palaeozoic, a small scolecodont assemblage was found in conodont-bearing peri-reefal limestones (Suttner and Hints 2010) but it differs strongly from the Binolen faunas by an extreme rarity of Oenonites-type forms.
In summary, rich scolecodont occurrences, especially Oenonites faunas, are not normal in reefal facies but more typical for calmer neritic settings. Therefore, BF-S indicates a strong influx of open shelf faunas. Since only three sampled beds are completely without scolecodonts, even when the absolute number of microfossils is rather low, we conclude that the main part of the Binolen Member lacked a seaward barrier.
Chitinozoa Biofacies (MF-1)
Examples: Beds 31 and 33
Definition: Microfaunas with ca. 15–35% assumed calcified moulds of chitinozoans.
Discussion: At Binolen, the abundance of calcified chitinozoans is strongly correlated with a high abundance of scolecodonts, a co-occurrence noted previously in an Ordovician example by Eriksson (1997). Laufeld (1974) suggested an inverse relationship between chitinozoan abundance and carbonate content, which is strange with respect to their assumed planktonic lifestyle, for which the sea-floor substrate should be of little significance. Among the few forms with similar shape, Gaugris and Grahn (2006) described Angochitina pseudoavelinoi from Frasnian siltstones of the São Domingos Formation of the Paraná Basin (Brazil), while Lagenochitina avelinoi was described by Grahn and de Melo (2002) from the Frasnian lower Barreirihna Formation of the Amazonas Basin, which comprises well-laminated dark grey to black shales. They are also present in Frasnian strata of the Parnaíba Basin (Brazil, Grahn and de Melo 2005) and Bolivia (Iquiri Formation, Grahn 2002). Since our material is older and from completely different lithofacies, the taxonomy remains arbitrary, and no biofacies conclusions can be drawn from the South American occurrences. A range of non-calcified chitinozoans, including Angochitina, occurs in Givetian open neritic limestones with brachiopods, trilobites, conodonts, crinoids, and rugose corals of Iowa (e.g. Urban 1972). This is the closest known equivalent to the Binolen faunas.
Porifera-Scolecodont Biofacies (MF-1)
Examples: Beds 15–16, 18a, 20b–29, 35, 44, 48, 50, and 52 (Fig. 15)
Definition: Microfaunas with >20% sponge spicules, >10% scolecodonts, and >70% of both groups combined. Dominant are the various types of heteractinid spicules, represented by Ensiferites langeri and Gondekia sp.
Discussion: For the scolecodont facies distributions, see BF-S. There is intergradation from BF-S towards BF-PS and BF-PE. Hereractinid sponge spicule occurrences are known from Devonian reefal facies of several regions. As pioneer work, Bartenstein (1935) described them from the Middle Devonian of the Eifel Mountains (Fleringen, Prüm Anticline). Much later, Langer (1991) added material from the Eifelian Lower Nohn Formation of the Hillesheim Anticline (Eifel Mountains). Additionally, he gave examples of both species of polyactinellid sponges found at Binolen from the Hustley-Barley and Wotan Members (Loogh Formation) at Üxheim, naming them “Phobetractin A and B”. Kozur et al. (2008) described Ensiferites langeri from the the Hustly-Barley Member of Gerolstein. East of the Rhine, Hartkopf-Fröder and Weber (2016) found octactinellid spicules in the biostromal, middle Frasnian Refrath Formation of Bergisch Gladbach-Refrath.
From the Holy Cross Mountains (Gałęzice Syncline) of Poland, Hurcewicz (1993) mentioned Ensiferites spicules from upper Givetian reef facies. She also described calcareous monaxones from Laskowa Góra, with the same combination of straight and curved types as at Binolen (Fig. 6q–r). However, she suggested that they were originally siliceous and calficied during diagenesis, which is not likely. Another comparable assemblage of spicules was mentioned by Kučera (1993) from the Pragian shallow marine Dvorce-Prokop Limestones of the Barrandian, Czech Republic (Rietschel 1968). Gondekia and Ensiferites spicules were described as “Octactines type A and B”, while the polyactinellid sponges Praephobetractinia and Dvorcia were illustrated as “Dichodiaenes type A and B”. Gondekia was first described by Rigby (1991) from the Givetian Hungry Hollow Formation of southwestern Ontario. The local facies represent a coral-dominated biostrome overlying bedded limestones with interbedded shales. Van Hinte et al. (1995) documented Ensiferites spicules from two pebbles dredged from an offshore crustal segment named as Orphan Knoll, northeast of Newfoundland, Canada. They were associated with simple cone conodonts, suggesting a similar setting as at Binolen, but their age is not clear.
Both the Rhenish heteractinid and polyactinellid sponges obviously preferred subtidal, neritic to biostromal limestone facies but seem to have avoided bioherms, their slopes and lagoons. They do not co-occur with hexactinellids; instead they built their own habitat in combination with echinoderms (Mostler 1996). This is corroborated by observations at Binolen, where heteractinid spicules are associated both with scolecodonts and echinoderms, and only with small amounts of foraminifers and chitinozoans.
Porifera-Echinodermata Biofacies (MF-1)
Examples: Beds 8–10, 12, 39–42, 47, and 51
Definition: Assemblages with >20% of sponge spicules, >10% echinoderm sclerites, and with >50% of both combined.
Discussion: This biofacies differs from BF-PS by a larger number of echinoderms and a reduced amount of scolecodonts. In general, there is a good correlation of sponge spicule and echinoderm abundances at Binolen (Fig. 15), with a dominance of the first as skeletal elements in residues. Scolecodonts, however, are still present and occur together with some foraminifers and other microfauna. The calcified chitinozoans are missing or occur only in very minor quantities.
The echinoderms are mostly represented by crinoid ossicles and holothuroid sclerites. In Middle Devonian reefs, the predominant position of crinoid meadows is on the outer slopes of bioherms and along the margin of biostromes (e.g. Burchette 1981; Machel and Hunter 1994). They may extend on carbonate ramps into shallow pelagic settings (e.g. Hartenfels 2011) but are also common on shallow silisiclastic shelfs (e.g. Çinar 1978). Disarticulation and current transport are the rule. At Binolen, high numbers of ossicles in the residues correlate with crinoid-rich microfacies types.
The holuthuroids are more characteristic for BF-PE. Eocaudina ovalis was described previously from Frasnian deposits (Matyja et al. 1973; Boczarowski 2001). It occurs in biostromal limestones (e.g, Korczmin, Poland) as well as in cephalopod limestones (Manticoceras limestone of Kadzielnia, Poland), resulting in a broad facies spectrum (Matyja et al. 1973). Other Givetian species of Eocaudina have been described from various biostromal units of the Eifel Mountains and of the Steltenberg Reef of Hohenlimburg (Langer 1991; Koch-Früchtl and Früchtl 1993), a reef drowning episode (“Schlade Event”) in the Büchel Formation (Bohatý and Herbig 2010), and middle Givetian shallow pelagic limestones of Pic de Bissous in the Montagne Noire, southern France (Aboussalam 2003). In summary, Eocaudina preferred the open shelf side in reef complexes; it is not known from reef core and backreef settings. In the Devonian, Staurocaudina is known from neritic brachiopod facies (Boczarowski 2001) but otherwise has rarely been mentioned. The combined heteractinid-crinoid-holothuroid, very poor conodont, and microfacies evidence suggests that BF-PE represents a moderately open, agitated, shallow-water carbonate platform. In the initial reef phase of Hagen-Hohenlimburg (Koch-Früchtl and Früchtl 1993), calcareous sponge spicules and holothurian sclerites occur abundantly either in somewhat deeper neritic, peloidal wackestones with various icriodids and Belodella (their MF-Typ 4) or in coral-stromatoporoid floatstones (their MF-Typ 2).
Facies development at Binolen and comparisons
The parallel plotting of BF and MF in the section log (Fig. 4) gives a moderate correlation between both. Microfossils were found in each sampled bed but in different absolute abundances (Fig. 14, Tab. 2). Beds with low abundances are assigned to BF types with reservation (see shaded samples in Fig. 15). Due to laboratory constraints on sample numbers, combined BF- and MF-assignments are not available for all beds.
Based on the combined microfacies and microfauna analyses, the Binolen succession, excluding the Upper Honsel Formation at the base, consists of seven depositional phases, termed I to VII (Figs. 4, 15). There are minor subdivisions (e.g. of Depophases I and IV). Depophases I to VI represent the Binolen Member, Depophase VII the basal Asbeck Member (Fig. 4). Despite the moderate MF-BF correlation, it is remarkable that all microfacies transitions can also be recognised by changes in the microfossil assemblages (Fig. 15). This suggests that these reflect palaeoecological changes in the overall biostrome part, not the random arrival of allochthonous skeletal material.
Upper Honsel Formation
The three lowermost layers at Binolen are siltstones at the top of the Upper Honsel Formation, which represents an offshore prodeltaic sedimentary system with very variable current regimes. For a more detailed analysis see Çinar (1978). One of the largest and youngest “intra-Honsel” biostromes occurs just 750 m to the SE of Binolen, at Volkringhausen-Ruthmecke (old quarry north of the railway station), but has never been studied in terms of faunas or microfacies. Two conodont samples were barren. The Volkringhausen Reef shows that parts of the Hönne Valley region were episodically free from high siliciclastic influx before the prodeltaic system came to a final halt.
The turn from the Upper Honsel to the basal Hagen-Balve Formation was somewhat more gradual in the west, at the Steltenberg Quarry of Hagen-Hohenlimburg, where there are silty, bioclastic (crinoidal) wacke-packstones with corals at the reef base, grading upwards into peloidal facies (Koch-Früchtl and Früchtl 1993).
Depophase I
At Binolen, a sharp facies break occurs at the base of Bed 4. The final end of significant siliciclastic discharge into the region was sudden. The lithology turns without transition into organic-rich, greyish limestones with rather varied reefal fauna, such as stromatoporoids, rugose and tabulate corals. Depophase Ia includes beds 4–7 and is characterised by predominant rudstones with significant crinoidal debris, an allobiostrome after Kershaw (1994). The weak rounding of bioclasts in MF-3 and MF-5a suggests short transport distances of reef builders and dwellers and the existance of adjacent crinoid meadows destructed by storm and wave activity. There are no facies characteristics of tempestites and turbidites, as they are known from the Givetian Garbeck Limestone at the steep southern end of the Hönne Valley Reef (Eder 1971). A bathymetric position within the realm of fair-weather waves contradicts assumptions that the initiation of the Hagen-Balve reef belt was caused by a significant transgression. Decisive, as in the case of the older “intra-Honsel” reef bodies, was the sudden termination of siliciclastic influx, which resumed only briefly and very moderately in subsequent depophases (see marlstone beds in Fig. 4). Since the southern coastline of Laurussia was far away (e.g. Langenstrassen 1983), we assume that the Hagen-Balve Reef belt settled on a structural high, which became later the core of the eastern Remscheid-Altena Anticline. Laterally, to the east, there was a contemporaneous basin characterised by deposition of organic-rich shales intercalated with turbiditic limestones (e.g. Ziegler 1965).
The peak occurrence of benthic, (orginally) pyritised ostracods (BF-O) and the incursion of Bipennatus at the base of the Binolen Member indicate shallow open shelf conditions and reduced ventilation inside Bed 4. This short phase was followed by a burst of agglutinating foraminifer population (BF-F) while calcareous sponges and scolecodonts appeared more gradually and first in small numbers. All groups benefitted at the seafloor from the termination of high clastic influx. The occurrence of ostracods ended in Bed 5. The coincident change among the conodonts implies a trend from chewing taxa with molar-type P1 elements (Bipennatus) to macrophagous predators (e.g. Murdock and Smith 2021) that hunted in an increasingly differentiated environment (Szaniawski 2009). Holothuroid sclerites increase in abundance but are always less common than crinoid columnals. At the top, we found the locally highest number of fish remains.
Depophase Ib (beds 8–15) is characterised by floatstones with reef builders of MF-5b that grade within beds into the crinoidal wacke-packstones of MF-3 and, only in Bed 8, of the less bioclastic MF-2. There is a variety of reef builders that characterise an agitated coral-stromatoporoid garden, which housed the only known bryozoans of the local succession (Tab. 2). There were interspersed crinoid meadows, which is also expressed by the abundant echinoderms of BF-PE. Complete, not fragmented crinoid ossicles suggest relative proximity to their original habitat. At the same time, bottom conditions worsened dramatically for the agglutinating foraminifers in Bed 8 (Figs. 14–15) while the marked proliferation of heteractinids suggests that the biostromal platform was still open seawards. However, the gradual disapperance of conodonts suggests shallowing upwards. Conditions resemble MF-Typ 2 developed in the lower part (2–5.5 m) of the Steltenberg Reef in Hagen-Hohenlimburg (Koch-Früchtl and Früchtl 1993).
The small BF-F fauna of Bed 11, combined with the unique MF-4a, indicates a short interruption of the facies trend. This is the only peloidal level, while such facies dominate the initial reef at Hagen-Hohenlimburg (Koch-Früchtl and Früchtl 1993). This indicates a distinctive difference at the beginning of reef growth. In the western region, micrite reworking bottom currents were much more common and continued throughout the local development of the Binolen Member.
At the top of Depophase Ib (Bed 15, BF-PS), scolecodonts show a first increase of abundance, as a transition towards the subsequent interval.
Depophase II
Depophase II occupies the lower middle part of the section from Bed 16 to the lower part of Bed 35, an interval with increasing intercalation of thin argillaceous beds and monotonous microfacies; only MF-2 occurs, characterised by short episodes of moderate bottom turbulence. The interval represents a deepening phase with some fluctuations of the microfossil assemblages, but BF-PS dominates. The heteractinids continue but are accompanied by increasing absolute numbers of monaxones (Tab. 2). Between beds 20b to 35, the heteractinids are almost exclusively Gondekia sp. and there are hardly any Ensiferites langeri. Polyactinellid sponges flourished and holothuroid sclerites became more frequent than crinoid osscles. Microvertebrates occur consistently in small numbers. The gradual rise of scolecodonts, with peak absolute abundances in beds 25, 29, 33, and lower 35 (Fig. 14, Tab. 2), is distinctive. The relative peak in Bed 18a (“BF-S”) is based on the potential bias of another very small microfossil collection. Transgression is supported by the re-appearance of conodonts, followed by a short recurrence of ostracods (BF-O) in beds 19 and 20a, suggesting episodically hypoxic conditions below the sediment surface. The influx of skeletal debris, and episodically also of fine siliciclastics (Fig. 4), remained too high for agglutinating foraminifers. At the maximum of deepening, in the argillaceous Bed 26, a large-sized, poorly stream-lined (breviconic) discosorid cephalopod entered the platform (Binoleniceras stichlingi, Afhüppe and Becker 2022, this issue).
A peculiar change of the microfossil assemblages occurred subsequently near the top, in beds 31–33, with the sudden onset of abundant, calcified chitinozoans, giving the only BF-C interval. The microfacies provides no evidence that the provenance of the fine fraction had changed. The largest density of chitinozoan tests ever observed was from pelagic marly limestones right at the Frasnian-Famennian boundary of the Montagne Noire (Paris et al. 1996). This bloom was based on a different taxon but it shows that the palaeoecology of the planctonic group was complex and is not fully understood. The combined peak abundance of calcified chitinozoans and scolecodonts near the top of Depophase II is unknown from any other beds. It may reflect a phase of increased surface water productivity. Together with the fine siliciclastics of intercalated marlstones, land-derived nutrients may have arrived. The highest absolute number of scolecodonts is from Bed 35 (Tab. 2, Fig. 14) but they are mostly so poorly preserved that they cannot be identified.
Depophase III
The next part of the section, from the top of Bed 35 to Bed 38, is a short regressive phase marked by thicker beds and the re-onset of MF-5b, floatstones with reef builders, notably with abundant Alv. (Alveolitella).
Depophase IV
Depophase IV is characterised by a sequence of facies fluctuations. The lower part (IVa) begins with a minor and short deepening trend characterised by a marly limestone (Bed 39), where the highest absolute number of sponge spicules was found. However, the trend reverted in beds 40–42 that are characterised by bioclastic wackestones of MF-2 grading into crinoidal packstones of MF-3. The enrichment of echinoderm debris is found in the microfossil assemblages of BF-PE, where the increase of heteractinid spicules, with a peak of absolute abundance in Bed 39 (Fig. 14), gives similarities with Depophase Ib lower down. The different MF/BF correlation is notable but a restricted openness of the platform is supported by the lack of conodonts. Depophase IVb includes beds 43–44, where floatstones with reef builders (MF-5b) correlate with a return of scolecodonts (BF-PS). This is a repetition of the environmental conditions of Depophase III, interpretated as a shallowing phase. Depophase IVc, beds 45–46, includes the next minor deepening phase with MF-2, with limited support by a peak of relative foraminifer abundance (BF-F) as at the section base, however in a small fauna with restricted value (Figs. 14–15).
Depophase V
The next interval, represented by beds 47–53, is characterised by alternating and intergrading of MF-2 and MF-5b coinciding with alternating BF-PE and BF-PS, which gives 1 ½ cycle. Following a more restricted platform setting with abundant reef builders and crinoid ossicles at the base (Bed 47: MF-5b, BF-PE), a minor deepening trend is indicated by subsequent thin bedding and three marly intercalations (beds 48, 50, 52). This is supported by peaks of scolecodont abundance, as within Depophase II, and by abundant monaxones (Tab. 2). In contrast, simple cone conodonts re-appear only in a single sample (Bed 50). The microfossil assemblage from Bed 49, consisting only of scolecodonts (BF-S), is embraced by richer samples with more abundant (in absolute numbers, Fig. 14) specimens of the group. The first deepening cycle ends in Bed 51 with a return to the microfauna assemblage similar as in Bed 47, characterised by frequent crinoid ossicles. A small foraminifer peak (only of Tolypammina) suggests a reduced sedimentation rate. It is followed by a second bloom of scolecodonts and decline of crinoids starting in Bed 52, a new onset of BF-PS, as in the deeper/more open middle part of Depophase V.
Depophase VI
This interval comprises beds 54–58 and the subsequent cliff (beds 59–65). It differs from the units below by MF-7, coral-stromatoporoid frame-rudstones, and MF-8, coral-dominated bafflestones, of an (auto)parabiostrome. As expected from numerous failed attempts to date such facies in the past, there are no conodonts. The microfaunas of the lower part, with MF-7, are characterised by decreasing absolute numbers of microfossils (Fig. 14) but first by very high proportions of scolecodonts (BF-S, beds 54, 56), then of foraminifers (BF-F, Bed 59). This gives a contradiction to the preference of both groups for open neritic platforms and to the palaeoecological interpretations below. It seems that the biostrome frame provided suitable niches for the vagile annelids, unlike the crinoid-rich beds of BF-PE. Bed 59 differs from all others by its peloidal pack-grainstone matrix, which implies winnowing of fine detritus at the seafloor. In this case, an allochthonous influx of the agglutinating foraminifers from an adjacent more open setting with lower sedimentation rate has to be considered.
Depophase VII
Following a small outcrop gap, beds 66–73 are dominated by MF-9, fenestral (microbial) and peloidal pack- and bindstones as they are widely known from lagoonal settings of bioherms with protecting reef core, especially of the main Asbeck Member of the Hönne Valley region. Since our work was focused on the Binolen Member, and because of complete failure of more than ten samples from the nearby Asbeck Quarry (see section description in Schudack 1993), no microfossil samples were taken.
Conclusions
-
1.
The Hagen-Balve Reef, especially the Hönne Valley region, is both an outstanding and typical example for Givetian coral-stromatoporoid reefs of the Rhenish Massif. The unique outcrop at Binolen enabled a representative case study to better understand a Rhenish initial reef phase, which is locally assigned to the new Binolen Member of the Hagen-Balve Formation. Results enable comparisons with previous data from the Steltenberg Quarry of Hagen-Hohenlimburg to the west.
-
2.
Macrofaunas identified from thin sections give insights into the local diversity and composition of early-stage reef builders. There are more than 40 species of stromatoporoids, chaetetids, rugose and tabulate corals, bryozoans, and calcimicrobes. These are mostly known from other Rhenish reefs, including the small-sized, older coral limestones intercalated within the Ihmert and Honsel formations of the Sauerland. There are some rugose corals, such as Mesophyllum and Dendrostella, which are not known from reef core or lagoonal settings of bioherms and that suggest open shelf conditions for the Binolen Member.
-
3.
A late early Givetian age of the Binolen Member is indicated from the combined evidence of Stringocephalus, some corals, and a rare Icriodus ?latecarinatus. Assuming a roughly contemporaneous onset of the Hagen-Balve Reef along its W-E extent, this is supported by conodont data from the Steltenberg Quarry at Hagen-Hohenlimburg (Koch-Früchtl and Früchtl 1993).
-
4.
The microfacies analysis led to the distinction of nine MF types. MF-1 comprises the prodeltaic and locally poorly fossiliferous siltstones of the Upper Honsel Formation. The Binolen Member consists of MF-2 to MF-7, with several subtypes. The bioclastic and variably crinoidal MF-2/3 include poorly sorted and non-graded wacke- to packstones with some reef builders, characteristic for an open, shallow neritic platform or allobiostrome sensu Kershaw (1994). In the peloidal facies of MF-4, recurrent wave/current action reworked or washed out the micrite, suggesting slightly shallower condition. The float-rudstones of MF-5 represent an agitated “stromatoporoid-coral-crinoid garden” facies, which alternates in the upper part of the section with thin marly intervals, suggesting short-term fluctuations of the palaeoenvironment. Allobiostrome debris of MF-2/3 was partly stabilised by coverstones of biotic origin (MF-6). The coral-stromatoporoid frame-rud- and bafflestones of MF-7/8 belong near the top of the member to a morphologically more prominent (auto)parabiostrome sensu Kershaw (1994). Fenestral limestones (MF-9), a shallow lagoonal and microbial facies type, appear at the base of the Asbeck Member.
-
5.
Two new conodont biofacies types are introduced for notoriously specimen-poor open biostromal settings. The Bipennatus Biofacies was first recognised in the Sparganophyllum Limestone east of the Hönne Valley. The Simple Cone Biofacies is characterised by a strong dominance of predatory, grasping or even venomous simple cone genera, such as Panderodus, Neopanderodus, Dvorakia, and Belodella. As in all conodont biofacies types, transitions towards other neritic biofacies (e.g. Icriodid and Polygnathid biofacies) are known from the literature.
-
6.
More than 4200 calcareous, agglutinating, (originally) pyritic, and organic microfossils picked from conodont sample residues belong to fluctuating assemblages with variably common ostracods, foraminifers, sponge spicules, echinoderm sclerites, scolecodonts, and assumed calcified chitinozoans. Since there is no microfacies evidence for major, short-termed sedimentary events (tempestites, turbidites), changes of microfossil compositions reflect partly palaeoecological fluctuations on the overall platform, due to water agitation and transport not necessarily at the collected site. Taphonomy and diagenesis affected the preservation of ostracods and chitinozoans but we observed no bias by the uniform sample processing.
-
7.
The systematic quantitative analysis of reefal microfossil assemblages represents pioneer work. Empirically, we separate six microfossil biofacies types. The Ostracoda Biofacies (BF-O) is characterised by increases of originally pyritic, secondarily oxidised ostracod moulds, which suggests episodic hypoxia within the sediment. The Foraminifera Biofacies (BF-F) is defined by assemblages with >50% of benthic agglutinating forms, which thrive in times of low sedimentation rates. The most common taxa (Tolypammina, Thurammina, Psammosphaera) are more typical for open outer shelf to pelagic platform/ramp settings. The Scolecodont Biofacies (BF-S) includes assemblages with abundant Oenonites, which is also an open neritic form; it was previously not known to have thrived in biostromal facies. The Chitinozoa Biofacies (BF-C) is difficult to interpret with respect to the uncertain interpretation of the microfossils as calcified chitinozoan moulds. The mixed Porifera-Scolecodont Biofacies (BF-PS) includes abundant calcitic monaxones and spicules of Heteractinida and Polyactinellida. They lived on moderately open, shallow neritic platforms, clearly separate from the mostly deeper realm of hexactinellids. The Porifera-Echinodermata Biofacies (BF-PE) is characterised by assemblages with >50% crinoid ossicles, holothuroid sclerites, and calcitic sponge spicules. The palaeoenvironment is interpreted as agitated, moderately open and shallow platform.
-
8.
The combined data from microfacies, conodonts, and microfossil assemblages is used to reconstruct the depositional history at Binolen, separating seven depophases, with subdivisions. The sharp facies break at the top of the Upper Honsel Formation shows that the siliciclastic influx from the distant Laurussia stopped suddenly. Since there is no microfacies evidence for deepening, we support the hypothesis that it was the combination of climate change (aridification), decreasing erosion and clastic discharge, not sea-level, which controlled the beginning of the main Middle Devonian reefal stage in the Rhenish Massif. The global eustatic curve (last update in Becker et al. 2020) shows no transgression in the higher part of the lower Givetian.
-
9.
Depophase I is characterised by the flourishing of crinoidal meadows, early reef builders, and of benthic foraminifers (BF-F) when the influx of silt and fine sand ended. Other faunal groups, such as brachiopods (stringocephalids and others), conodonts, scolecodonts, and calcareous sponges spread gradually in the basal allobiostrome. The increased content of organic matter led to a short initial phase with hypoxic conditions within the sediment and ostracod pyritisation (BF-O of Bed 4). There is a slight shallowing upwards trend, with higher sedimentation rate delimiting the foraminifer population, manifested in BF-PE. Depophase II is a deepening phase with monotonous bioclastic wackestones (MF-2), flourishing scolecodonts, holothurians (mostly Eocaudina), and sponges (dominant BF-PS). In the upper part, perhaps connected with marly interbeds, peak abundances of assumed, planktonic chitinozoans (BF-C) may reflect increased surface water productivity. Depophase III is a short regressive phase, followed by strongly fluctuating BF- and MF-types in Depophase IV, reflecting two minor sea-level cycles, with a small bloom of foraminifers at the top. In Depophase V, short-term facies oszillations continue but with more marl interbeds that bracket a scolecodont bloom (BF-S). Depophase VI is characterised by frame-rud- and bafflestones of MF-7/8 of an (auto)parabiostrome, culminating in the coral-rich cliff at the top of the Binolen Member. In the absence of micritic matrix, we consider that a small foraminifer peak (of BF-F) was caused by specimens washed in from an adjacent calmer region. Depophase VII includes the fenestral, microbial facies (MF-9) of the lagoonal, basal Asbeck Member, which implies that a protecting reef core had been established.
-
10.
There are both similarities (abundance of calcareous sponge spicules and holothurian sclerites, scolecodonts) and differences (completely different foraminifer assemblages, conodont and microvertebrate diversity, microfacies) between the contemporaneous initial phases of the Hagen-Balve Reef in the west (Steltenberg, Koch-Früchtl and Früchtl 1993) and east (Binolen). Especially microfossil successions should be studied in more and different reef complexes, in order to test our results and to improve their value as palaeoenvironmental indicators.
References
Aboussalam, Z. S. (2003). Das "Taghanic-Event" im höheren Mittel-Devon von West-Europa und Marokko. Münstersche Forschung zur Geologie und Paläontologie, 97, 1–332.
Aboussalam, Z. S., & Becker, R. T. (2001). Prospects for an upper Givetian substage. Mitteilungen aus dem Museum für Naturkunde Berlin, Geowissenschaftliche Reihe, 4, 83–99.
Aboussalam, Z. S., & Becker, R. T. (2011). The global Taghanic Biocrisis (Givetian) in the eastern Anti-Atlas, Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 136–164.
Aboussalam, Z. S., Becker, R. T., & Bultynck, P. (2015). Emsian (Lower Devonian) conodont stratigraphy and correlation of the Anti-Atlas (Southern Morocco). Bulletin of Geosciences, 90(4), 893–980.
Aboussalam, Z. S., Becker, R. T., Eichholt, S., El Hassani, A., Bouari, A., Mottequin, B., & Baidder, L. (2021). The Devonian of Jebel Ardouz (Mzoudia region, SW Moroccan Meseta) – new data on stratigraphy, facies, and palaeogeography. Frontiers in Science and Engineering, Earth, Water and Oceans, Environmental Sciences, 10(2), 225–249.
Afhüppe, L., & Becker, R. T. (2022). A new discosorid and some other nautiloids from the Givetian of the Rhenish Massif, Germany. In S. Hartenfels, C. Hartkopf-Fröder, & P. Königshof (Eds.), The Rhenish Massif: More than 150 years of research in a Variscan mountain chain. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-022-00541-3. [this issue]
Avlar, H., & May, A. (1997). Zur Fauna und Stratigraphie der cultrijugatus-Schichten (Wende Unter-/Mittel-Devon) im West-Sauerland (Rheinisches Schiefergebirge). Coral Research Bulletin, 5, 103–119, pls. 1–4.
Bandel, K. (1969). Feinstratigraphische und biofazielle Untersuchungen unterdevonischer Kalke am Fuße der Seewarte (Wolayer See: zentrale Karnische Alpen). Jahrbuch der Geologischen Bundesanstalt, 112, 197–234.
Bandel, K., & Frýda, J. (1998). The systematic position of the Euomphalidae (Gastropoda). Senckenbergiana lethaea, 78(1/2), 103–131.
Bargatzky, A. (1881). Die Stromatoporen des Rheinischen Devons. Verhandlungen des Naturhistorischen Vereins der Preußischen Rheinlande und Westfalens, 38, 233–304.
Bartenstein, H. (1935). Schwamm-Nadeln im Mittel-Devon der Eifel und Ober-Devon des Ost-Thüringischen Schiefergebirges. Senckenbergiana, 17(5), 238–241.
Becker, G. (1969). Ostracoda aus dem Mitteldevon der Sötenicher Mulde (N-Eifel). Biostratigraphie, Paläoökologie und taxonomische Bedingungen. Senckenbergiana lethaea, 50(2/3), 239–271.
Becker, G. (1971). Paleoecology of Middle Devonian ostracods from the Eifel region, Germany. Bulletin du Centre des recherches de Pau – SNPA, 5(supplement), 801–816.
Becker, R. T., Aboussalam, Z. S., Stichling, S., May, A., & Eichholt, S. (2016). The Givetian-Frasnian Hönne Valley Reef Complex (northern Sauerland) – an outline of stratigraphy and facies development. Münstersche Forschungen zur Geologie und Paläontologie, 108, 126–140.
Becker, R. T., Aboussalam, Z. S., El Hassani, A., Eichholt, S., & Helling, S. (2020). The Devonian of the Oued Cherrat Zone (Western Meseta) – review and new data. Frontiers in Science and Engineering, Earth, Water and Oceans, Environmental Sciences, 10(1), 27–85.
Becker, R. T., Hartenfels, S., & Kaiser, S. I. (2021a). Review of Devonian-Carboniferous Boundary sections in the Rhenish Slate Mountains (Germany). In M. Aretz, & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2), 357–420.
Becker, R. T., Aboussalam, Z. S., El Hassani, A., Baidder, L., Weyer, D., & Jansen, U. (2021b). Devonian of the Benahmed region, western Moroccan Meseta. Frontiers in Science and Engineering, Earth, Water and Oceans, Environmental Sciences, 10(2), 13–73.
Becker, R. T., Aboussalam, Z. S., El Hassani, A., Baidder, L., Hüneke, H., Mayer, O., Cózar, P., Helling, S., Seyffert, K., Afhüppe, L., & May, A. (2021c). Devonian and the Carboniferous transgression in the Skoura region, Sub-Meseta Zone, Morocco. Frontiers in Science and Engineering, Earth, Water and Oceans, Environmental Sciences, 10(2), 251–333.
Berner, R. A. (1984). Sedimentary pyrite formation: An update. Geochimica et Cosmochimica Acta, 48, 605–615.
Berner, R. A. (1985). Sulphate reduction, organic matter decomposition and pyrite formation. Philosophical Transactions of the Royal Society London, A315, 25–38.
Beyrich, E. (1837). Beiträge zur Kenntniss der Versteinerungen des Rheinischen Übergangsgebirges. Erstes Heft. Berlin: Dümmler.
Birenheide, R. (1978). Rugose Korallen des Devon. Leitfossilien, 2, 1–265.
Birenheide, R. (1985). Chaetetida und tabulate Korallen des Devon. Leitfossilien, 3, 1–249.
Bischoff, G., & Ziegler, W. (1957). Die Conodontenchronologie des Mitteldevons und des tiefsten Oberdevons. Abhandlungen des Hessischen Landesamtes für Bodenforschung, 22, 1–136.
Blodgett, R. B., Boucot, A. J., & Koch, W. F. II (1988). New occurrences of color patterns in Devonian articulate brachiopods. Journal of Paleontology, 62(1), 46–51.
Boczarowski, A. (2001). Isolated sclerites of Devonian non-pelmatozoan echinoderms. Palaeontologica Polonica, 59, 1–219.
Bohatý, J. (2005). Doppellagige Kronenplatten: Ein neues anatomisches Merkmal paläozoischer Crinoiden und Revision der Familie Cupressocrinitidae (Devon). Paläontologische Zeitschrift, 79(2), 201–225.
Bohatý, J. & Herbig, H. G. (2010). Middle Givetian echinoderms from the Schlade Valley (Rhenish Massif, Germany): habitats, taxonomy and ecostratigraphy. Paläontologische Zeitschrift, 80, 365–85.
Brady, H. B. (1879). Notes on some of the reticularian Rhizopoda of the “Challenger” Expedition; Part I. On new or little known arenaceous types. Quarterly Journal of Microscopical Sciences, 19, 20–67.
Branson, E. B., & Mehl, M. G. (1933). Conodont studies. University of Missouri Studies, 8(1/2), 1–167, pls. 1–12.
Bultynck, P. (1974). Conodontes de la Formation de Fromelennes du Givetien de l’Ardenne Franco-Belge. Bulletin de l´Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 50(10), 1–30.
Bultynck, P. (1987). Pelagic and neritic conodont successions from the Givetian of pre-Sahara Morocco and the Ardennes. Bulletin de l´Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 57, 149–181.
Burchette, T. P. (1981). European Devonian reefs: A review of current concepts and models. SEPM Special Publication, 30, 85–142.
Chatterton, B. D. E. (1974). Middle Devonian conodonts from the Harrogate Formation, southeastern British Columbia. Canadian Journal of Earth Sciences, 11, 1461–1484.
Choo, B. (2015). A new species of the Devonian actinopterygian Moythomasia from Bergisch Gladbach, Germany, and fresh observations on M. durgaringa from the Gogo Formation of Western Australia. Journal of Vertebrate Paleontology, 35(4), 1–21.
Çinar, C. (1978). Marine Flachwasserfazies in den Honseler Schichten (Givet-Stufe, Lüdenscheider Mulde, Rechtsrheinisches Schiefergebirge). Göttinger Arbeiten zur Geologie und Paläontologie, 20, 1–77.
Clark, D. L. (1984). Conodont Biofacies and Provincialism. The Geological Society of America, Special Paper, 196, 1–340.
Coen-Aubert, M. (2002). Temnophyllids and Spinophyllids (Rugosa) from the Givetian Mont d’Haurs Formation in Belgium. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 72, 5–24.
Copper, P., & Scotese, C. R. (2003). Megareefs in Middle Devonian supergreenhouse climates. Geological Society of America, Special Paper, 370, 209–230.
Defrance, M. (1825). p. 511. In H. M. D. Blainville (Ed.), Manuel de Malacologie et de Conchyliologie. Paris.
Dunham, R. J. (1962). Classification of carbonate rocks according to depositional texture. American Association of Petroleum Geologists, Memoir, 1, 108–121.
Dunn, P. H. (1942). Silurian foraminifera of the Mississippi Basin. Journal of Paleontology, 16(3), 317–342.
Ebert, J. (1994). Crinoiden-Stielglieder aus der Ems- und Eifel-Stufe des Sauerlandes (Rheinisches Schiefergebirge). Göttinger Arbeiten zur Geologie und Paläontologie, 64, 1–85.
Eder, F. W. (1971). Riff-nahe detritische Kalke bei Balve im Rheinischen Schiefergebirge (Mittel-Devon, Garbecker Kalk). Göttinger Arbeiten zur Geologie und Paläontologie, 10, 1–66.
Eichholt, S., & Becker, R. T. (2016). Middle Devonian reef facies and development in the Oued Cherrat Zone and adjacent regions (Moroccan Meseta). Facies, 62(7), 29 pp. https://doi.org/10.1007/s10347-015-0459-z.
Eickhoff, G. (1973). Das hohe Oberdevon und tiefe Unterkarbon im Bahneinschnitt Oberrödinghausen bei Menden (Rheinisches Schiefergebirge). Compte Rendu Septième Congrès International de Stratigraphie et de Géologie du Carbonifère, Krefeld 23.-28. August 1971, 2, 417–434.
Eller, E. R. (1938). Scolecodonts from Potter Farm Formation of the Devonian of Michigan. Annals of the Carnegie Museum, 27, 175–186.
Eller, E. R. (1941). Scolecodonts from the Windom, Middle Devonian, of western New York. Annals of the Carnegie Museum, 28, 323–440.
Eller, E. R. (1963). Scolecodonts from the Dundee, Devonian of Michigan. Annals of the Carnegie Museum, 36, 173–180.
Eller, E. R. (1964). Scolecodonts of the Delaware Limestone, Devonian of Ohio and Ontario. Annals of the Carnegie Museum, 36, 229–275.
Embry, A. F., & Klovan, E. J. (1971). A late Devonian reef tract on Northeastern Banks Island, NWT. Canadian Society of Petroleum Geologists, 19, 730–781.
Eriksson, M. (1997). Lower Silurian polychaetaspid polychaetes from Gotland, Sweden. GFF, 119(3), 213–230.
Faber, P. (1980). Fazies-Gliederung und Entwicklung im Mittel-Devon der Eifel. Mainzer geowissenschaftliche Mitteilungen, 8, 83–149.
Fåhræus, L. E., Slatt, R. M., & Nowlan, G. S. (1974). Origin of carbonate pseudopellets. Journal of Sedimentary Petrology, 44(1), 27–29.
Flügel, E. (1978). Mikrofazielle Untersuchungsmethoden von Kalken. Berlin: Springer.
Flügel, E. (2010). Microfacies of carbonate rocks. Analysis, interpretation and application. Second Edition, Berlin: Springer.
Flügel, E., & Hötzl, H. (1971). Foraminiferen, Calcisphaeren und Kalkalgen aus dem Schwelmer Kalk (Givet) von Letmathe im Sauerland. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 137(3), 358–395.
Flügel, E., & Kiessling, W. (2002). Patterns of Phanerozoic reef crises. Society for Sedimentary Geology, Special Publication, 72, 691–733.
Frýda, J., & Manda, S. (1997). A gastropod faunule from the Monograptus uniformis graptolite Biozone (Early Lochkovian, Early Devonian) in Bohemia. Mitteilungen des Geologisch-Paläontologischen Instituts der Universität Hamburg, 80, 59–121.
Gaugris, K. A. & Grahn, Y. (2006). New chitinozoans from the Devonian of the Paraná Basin, south Brazil, and their biostratigraphic significance. Ameghiniana, 34(2), 1–18.
Gluchowski, E. (1993). Crinoid assemblages in the Polish Givetian and Frasnian. Acta Palaeontologica Polonica, 38(1), 35–92.
Goldfuss, G. A. (1826–1833). Petrefacta Germaniae. Erster Theil. Düsseldorf: Arnz & Comp.
Goldring, R., & Langenstrassen, F. (1979). Open shelf and near-shore clastic facies in the Devonian. In M. R., House, C. T. Scrutton, & M. G. Bassett (Eds.), The Devonian System – A Palaeontological Association International Symposium. Special Papers in Palaeontology, 23, 81–97.
Grahn, Y. (2002). Upper Silurian and Devonian chitinozoans from central and southern Bolivia, central Andes. Journal of South American Earth Sciences, 15, 315–326.
Grahn, Y. & de Melo, J. H. G. (2002). Chitinozoan biostratigraphy of the Late Devonian formations in well Caima PH-2, Tapajós River area, Amazonas Basin, northern Brazil. Review of Palaeobotany and Palynology, 108, 115–139.
Grahn, Y. & de Melo, J. H. G. (2005). Middle and Late Devonian Chitinozoa and biostratigraphy of the Parnaíba and Jatobá Basins, northeastern Brazil. Palaeontographica Abteilung B, 272, 1–50.
Gutschick, R. C. (1962). Arenaceous foraminifera from the oncolites in the Mississippian Sappington Formation of Montana. Journal of Paleontology, 36(6), 1291–1304.
Gutschick, R. C., & Sandberg, C. A. (1983). Mississippian continental margins of the conterminous United States. Society of Economic Paleontologists and Mineralogists, Special Publication, 33, 79–96.
Gutschick, R. C., Weiner, J. L., & Young, L. (1961). Lower Mississippian arenaceous Foraminifera from Oklahoma, Texas, and Montana. Journal of Paleontology, 35(6), 1193–1221.
Ham, W. E. (1952). Algal origin of the “Birdseyes” Limestone in the McLish Formation. Oklahoma Academy of Sciences, Proceedings, 33, 200–203.
Hartenfels, S. (2011). Die globalen Annulata-Events und die Dasberg-Krise (Famennium, Oberdevon) in Europa und Nord-Afrika – hochauflösende Conodonten-Stratigraphie, Karbonat-Mikrofazies, Paläoökologie und Paläodiversität. Münstersche Forschung zur Geologie und Paläontologie, 105, 17–527.
Hartkopf-Fröder, C., & Weber, H. M. (2016). From Emsian coastal to Famennian marine environments: palaeogeographic evolution and biofacies in the Bergisch Gladbach-Paffrath Syncline area (Rhenish Massif, Germany). In R. T. Becker, S. Hartenfels, P. Königshof, & S. Helling (Eds.), Middle Devonian to Lower Carboniferous stratigraphy, facies, and bioevents in the Rhenish Massif, Germany – an IGCP 596 Guidebook. Münstersche Forschungen zur Geologie und Paläontologie, 108, 46–75.
Heidelberger, D., & Koch, L. (2005). Gastropoda from the Givetian “Massenkalk” of Schwelm and Hohenlimburg (Sauerland, Rheinisches Schiefergebirge, Germany). Geologica et Palaeontologica, Sonderband 4, 1–107.
Hinde, G. J. (1879). On annelid jaws from the Cambro-Silurian, Silurian and Devonian Formations in Canada and from the Lower Carboniferous in Scotland. The Quarterly Journal of the Geological Society of London, 35, 370–389.
Hinde, G. J. (1880). On annelid jaws from the Wenlock and Ludlow Formations of the West of England. The Quarterly Journal of the Geological Society of London, 36, 368–378.
Hints, O., & Eriksson, M. E. (2007). Diversification and biostratigraphy of scolecodont-bearing polychaetes in the Ordovician. Palaeogeography, Palaeoclimatology, Palaeoecology, 245, 95–114.
Hladil, J. (2005). The formation of stromatactis-type fenestral structures during the sedimentation of experimental slurries – a possible clue to a 120-year-old puzzle about stromatactis. Bulletin of Geosciences, 80(3), 193–211.
Høeg, O. A. (1932). Ordovician algae from the Trondheim area. In J. Kiaer (Ed.), The Hovin Group in the Trondheim area, II, Palaeontological part. Skrivter utgift av Det Norske Videnskaps-Akademi i Oslo, Matematisk-Naturvidenskapelig Klasse, 1927(4), 63–96.
Holcová, K. (2004). Foraminifers from the Lower/Middle Devonian boundary beds of the Barrandian area, Czech Republic, and their paleoecology. The Journal of Foraminiferal Research, 34(3), 214–231.
Holcová, K., & Slavík, L. (2013). The morphogroups of small agglutinated foraminifera from the Devonian carbonate complex of the Prague Synform, (Barrandian area, Czech Republic). Palaeogeography, Palaeoclimatology, Palaeoecology, 386, 210–224.
Hubmann, B. (1993). Ablagerungsraum, Mikrofazies und Paläoökologie der Barrandeikalk-Formation (Eifelium) des Grazer Paläozoikums. Jahrbuch der Geologischen Bundesanstalt, 136(2), 393–461.
Hurcewicz, H. (1993). Middle and Late Devonian sponge spicules of the Holy Cross Mountains and Silesian Upland. Acta Palaeontologica Polonica, 37(2–4), 291–296.
Ireland, H. A. (1939). Devonian and Silurian foraminifera from Oklahoma. Journal of Paleontology, 30, 831–864.
Iven, C. (1980). Alveolitiden und Heliolitiden aus dem Mittel- und Oberdevon des Bergischen Landes (Rheinisches Schiefergebirge). Palaeontographica, Abteilung A, 167(4–6), 121–179.
Janet, F. E. (1972). Gruppa Chaetetida, Podklass Tabulata. In A. N. Chodalevič (Ed.), Kišečnopolostnye i brachiopody živetskich otloženij vostočnogo sklona Urala (pp. 43–98). Moskva: Ministerstvo geologii SSSR, Ministerstvo geologii RSFSR, Ural’skoe territorial’noe geologičeskoe upravlenie.
Jux, U. (1960). Die devonischen Riffe im Rheinischen Schiefergebirge, Teil I. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 110(2), 186–258.
Kaminski, M. A. (2014). The year 2010 classification of the agglutinated foraminifera. Micropaleontology, 60(1), 89–108.
Kaminski, M. A., Grassle, J. F., & Whitlatch, R. B. (1988). Life history and recolonization among agglutinated Foraminifera in the Panama Basin. Jahrbuch der Geologischen Bundesanstalt, 41, 229–243.
Kamp, H. von (1972). Erläuterungen zu Blatt 4611 Hohenlimburg, 2. Völlig neu bearbeitete Auflage. Geologische Karte von Nordrhein-Westfalen 1:25000 (pp. 1–182). Krefeld: Geologisches Landesamt Nordrhein-Westfalen.
Kayser, E. (1879). Über einige Versteinerungen aus dem Kalk der Eifel. Zeitschrift der Deutschen geologischen Gesellschaft, 31(2), 301–307.
Kershaw, S. (1984). Patterns of stromatoporoid growth in level-bottom environments. Palaeontology, 27(1), 113–130.
Kershaw, S. (1994). Classification and geological significance of biostromes. Facies, 31, 81–92.
Kiessling, W. (2008). Sampling-standardized expansion and collapse of reef building in the Phanerozoic. Fossil Record, 11(1), 7–18.
Kiessling, W., Flügel, E., & Golonka, J. (1999). Paleoreef maps: Evaluation of a comprehensive database on Phanerozoic reefs. American Association of Petroleum Geologists, Bulletin, 83(1), 1552–1587.
Klapper, G., & Barrick, J. E. (1983). Middle Devonian (Eifelian) conodonts from the Spillville Formation in northern Iowa and southern Minnesota. Journal of Paleontology, 57(6), 1212–1243.
Koch-Früchtl, U., & Früchtl, M. (1993). Stratigraphie und Faziesanalyse einer mitteldevonischen Karbonatabfolge im Remscheid-Altenaer Sattel (Sauerland). Geologie und Paläontologie in Westfalen, 26, 47–75.
Königshof, P., Da Silva, A. C., Suttner, T. J., Kido, E., Water, J., Carmichael, S., Jansen, U., Pas, D., & Spassov, S. (2015). Shallow-water facies setting around the Kačák Event: a multidisciplinary approach. In R. T. Becker, P. Königshof, & C. E. Brett (Eds.), Devonian climate, sea level and evolutionary events, Geological Society London, Special Publications, 423, 171–199.
Kozur, H. (1991). Erster Nachweis von Polyactinellidae (Calcispongea, Porifera) im Perm und Revision der Gattung Phobetractinia. Zeitschrift für Geologische Wissenschaften, 19, 585–591.
Kozur, H., Mostler, H., & Repetski, J. (2008). A new Heteractinellid sponge from the lowermost Ordovician of Nevada and a discussion of the Suborder Heteractinellidae. Geo.Alp, 5, 53–67.
Krebs, W. (1974). Devonian carbonate complexes of Central Europe. In L. F. Laporte (Ed.), Reefs in time and space. Society of Economical Paleontologists and Mineralogists, Special Publication, 18, 155–208.
Kučera, M. (1993). Sponge Spicules fom the Lower Devonian (Pragian) of the Barrandian. Journal of Czech Geological Society, 38(3–4), 193–200.
Lange, F. W., (1952). Quitinozoários do Folhelho Barreirinha, Devoniano do Pará. Dusenia, 3, 373–386.
Langenstrassen, F. (1983). Neritic sedimentation of the Lower and Middle Devonian in the Rheinische Schiefergebirge east of the River Rhine. In H. Martin, & F. W. Eder (Eds.), Intracontinental Fold Belts (pp. 43–76). Berlin, Heidelberg, New York, Tokyo: Springer.
Langer, W. (1979). Neue karbonatische Microproblematica aus dem westdeutschen Devon. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 1979(12), 723–733.
Langer, W. (1991). Beiträge zur Mikropaläontologie des Devons im Rheinischen Schiefergebirge. Geologisches Jahrbuch, A128, 35–65.
Langer, W. (1997). Neue oder wenig bekannte Mikrofossilien aus dem europäischen Devon. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 205(1), 129–144.
Laufeld, S. (1974). Silurian Chitinozoa from Gotland. Fossils and Strata, 5, 1–130.
Lecompte, M. (1939). Les tabulés du Dévonien moyen et supérieur du bord du bassin de Dinant. Mémoires du Musée royal d’Histoire naturelle de Belgique, 90, 1–229.
Lecompte, M. (1951). Les stromatoporoides du Dévonien moyen et supérieur du bassin de Dinant. Première Partie. Mémoires de l'Institut Royal des Sciences Naturelles de Belgique, 116, 1–215.
Lecompte, M. (1952). Les stromatoporoides du Dévonien moyen et supérieur du bassin de Dinant. Deuxième Partie. Mémoires de l'Institut Royal des Sciences Naturelles de Belgique, 117, 217–359.
Lecompte, M. (1970). Die Riffe im Devon der Ardennen und ihre Bildungsbedingungen. Geologica et Palaeontologica, 4, 25–71.
Liao, J.-C., & Valenzuela-Ríos, J. I. (2017). Givetian (Middle Devonian) historical bio- and chronostratigraphical subdivision based on conodonts. Lethaia, 50, 440–446.
Löw, M. (2020). Mikrofaunen aus dem initialen Riffstadium von Binolen im Hönnetal (Sauerland, Givetium) (pp. 1–59). Unpublished B.Sc. Thesis, Münster: Westfälische Wilhelms-Universität Münster.
Löw, M., Söte, T., Becker, R. T., May, A., & Stichling, S. (2021). Microfauna and Microfacies from the initial reef stadium of Binolen in the Hönne Valley (Sauerland, Middle Devonian). In S. Stumpf, J. Türtscher, C. Pfaff, P. L. Jambura, & J. Kriwet (Eds.), Abstracts of the 92nd Annual Meeting of the Paläontologische Gesellschaft – Berichte der Geologischen Bundesanstalt, 142, p. 54. Wien.
Lu, J., & Königshof, P. (2019). Eifelian (Middle Devonian) species of Bipennatus from the Beiliu Formation at Nalai, South China. Palaeoworld, 29(4), 682–694.
Lüddecke, F., Hartenfels, S., & Becker, R. T. (2017). Conodont biofacies of a monotonous middle Famennian pelagic carbonate succession (Upper Ballberg Quarry, northern Rhenish Massif). In B. Mottequin, L. Slavík, & P. Königshof (Eds.) Climate change and biodiversity patterns in the mid-Palaeozoic. Palaeobiodiversity and Palaeoenviroments, 97(3), 591–613.
Machel, H. G., & Hunter, G. (1994). Facies models for Middle to Late Devonian shallow-marine carbonates, with comparisons to modern reefs: a guide for facies analysis. Facies, 30, 155–176.
Maillet, S., Danelian, T., & Casier, J.-G. (2016). Middle/Late Givetian ostracod assemblages from the Aisne quarry (Durbuy area, Ardenne, Belgium). Biostratigraphic and palaeoecological implications. Annales de Paléontologie, 102, 11–29.
Malmsheimer, K. W., Mensink, H., & Stritzke, R. (1991). Gesteinsvielfalt im Riffgebiet um Brilon. Geologie und Paläontologie in Westfalen, 18, 67–83.
Malmsheimer, K. W., Flajs, G., & Koch-Früchtl, U. (1996). Middle Devonian Initial Reef-Facies from the Rhenish Schiefergebirge (Sauerland and Eifel), Western Germany. In J. Reitner, F. Neuweiler, & F. Gunkel (Eds.), Global and Regional Controls on Biogenic Sedimentation. I. Reef Evolution. Research Reports. Göttinger Arbeiten zur Geologie und Paläontologie, Sb2, 371–375.
Mamet, B., & Préat, A. (1982). Givetianella tsienii, une Dasycladacée nouvelle du Givétien de la Belgique. Bulletin de la Société belge de Géologie, 91(4), 209–216.
Matyja, B. A., Matyja, H., & Szulczewski, M. (1973). The genus Eocaudina Martin (Holothuroidea) from the Devonian of Poland. Acta Geologica Polonica, 23, 135–147.
Maurer, F. (1885). Die Fauna der Kalke von Waldgirmes bei Gießen. Abhandlungen der Geologischen Landesanstalt, 1, 67–340.
Mawson, R. (1993). Bipennatus, a new genus of Devonian conodonts. Memoirs of the Australasian Association of Palaeontologists, 15, 137–140.
Mawson, R., & Talent, J. (1989). Late Emsian-Givetian stratigraphy and conodont biofacies – Carbonate slope and offshore shoal to sheltered lagoon and nearshore carbonate ramp – Broken River, North Queensland, Australia. Courier Forschungsinstuitut Senckenberg, 117, 205–259.
May, A. (1987). Der Massenkalk (Devon) nördlich von Brilon (Sauerland). Geologie und Paläontologie in Westfalen, 10, 51–84.
May, A. (1988). Fossilführung und Palökologie des lagunären Massenkalks (Devon) im Sauerland (Rheinisches Schiefergebirge). Paläontologische Zeitschrift, 62, 175–192.
May, A. (1991). Die Fossilführung des westsauerländischen Givetiums (Devon; Rheinisches Schiefergebirge) in der Sammlung des Städtischen Museums Menden. Geologie und Paläontologie in Westfalen, 17, 7–42.
May, A. (1992a). Paleoecology of Upper Eifelian and Lower Givetian Coral Limestones in the Northwestern Sauerland (Devonian, Rhenish Massif). Facies, 26, 103–116.
May, A. (1992b). Die Kalkalgen-Flora des Ober-Eifeliums und Unter-Givetiums (Devon) des nordwestlichen Sauerlandes (Rheinisches Schiefergebirge). Palaeontographica, Abteilung B, 228, 1–28.
May, A. (1993a). Korallen aus dem höheren Eifelium und unteren Givetium (Devon) des nordwestlichen Sauerlandes (Rheinisches Schiefergebirge). Teil I. Tabulate Korallen. Palaeontographica, Abteilung A, 227, 87–224.
May, A. (1993b). Korallen aus dem höheren Eifelium und unteren Givetium (Devon) des nordwestlichen Sauerlandes (Rheinisches Schiefergebirge). Teil II. Rugose Korallen, Chaetetiden und spezielle Themen. Palaeontographica, Abteilung A, 228(1/3), 7–42.
May, A. (1993c). Stratigraphie, Stromatoporen-Fauna und Palökologie von Korallenkalken aus dem Ober-Eifelium und Unter-Givetium (Devon) des nordwestlichen Sauerlandes (Rheinisches Schiefergebirge). Geologie und Paläontologie in Westfalen, 24, 5–93.
May, A. (1997). Sind die devonischen Riffe des Sauerlandes heutigen Korallenriffen vergleichbar? Dortmunder Beiträge zur Landeskunde, naturwissenschaftliche Mitteilungen, 31, 127–135.
May, A. (2005). Die Stromatoporen des Devons und Silurs von Zentral-Böhmen (Tschechische Republik) und ihre Kommensalen. Zitteliana, B25, 117–250.
May, A., & Marks, J. (2013). Eine Korallen-Fauna aus der Oberhonsel-Formation (Givetium; Devon) von Garbeck (West-Sauerland). Dortmunder Beiträge zur Landeskunde, naturwissenschaftliche Mitteilungen, 45, 69–80.
Mestermann, B. (1995). Fenstergefüge im südlichen Briloner Massenkalk. Geologie und Paläontologie in Westfalen, 41, 55–67.
Milne-Edwards, H., & Haime, J. (1851). Monographie des polypiers fossiles des terrains palæozoïques, précédée d’un tableau général de la classification des polypes. Muséum d’Histoire Naturelle, Archives, 5, 1–502.
Mostler, H. (1996). Polyactinellide Schwämme, eine auf das Paläozoikum beschränkte Calcispongien-Gruppe. Geologisch-Paläontologische Mitteilungen Innsbruck, 21, 223–243.
Murdock, D. J. E., & Smith, M. P. (2021). Panderodus from the Waukesha Lagerstätte of Wisconsin, USA: a primitive macrophagous vertebrate predator. Papers in Palaeontology, 7(4), 1997–1993.
Narkiewicz, K. (2015). Taxonomic revision and phylogenetic affinities of the conodont Bipennatus montensis (Weddige, 1977) from the Eifelian (Middle Devonian) of Poland. Paläontologische Zeitschrift, 89, 125–132.
Narkiewicz, K. & Bultynck, P. (2007). Conodont biostratigraphy of shallow marine Givetian deposits from the Radom-Lublin area, SE Poland. Geological Quarterly, 51(4), 419–442.
Narkiewicz, K., & Narkiewicz, M. (2010). Mid Devonian carbonate platform development in the Holy Cross Mts. area (central Poland): new constraints from the conodont Bipennatus fauna. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 255(3), 287–300.
Narkiewicz, K., Narkiewicz, M., & Bultynck, P. (2016). Conodont biofacies of the Taghanic transgressive interval (middle Givetian): Polish record and global comparisons. In R. T. Becker, P. Königshof, & C. E. Brett (Eds.), Devonian Climate Sea Level and Evolutionary Events. Geological Society, London, Special Publications, 423, 201–222.
Nekvasilova, O., & Stemprokova, D. (1960). Jehlice hub z branickych vrstev Barrandienu. Časpois pro mineralogii a geologii, 5(4), 400–405.
Nose, M., Schröder, S., & Fischer, U. (2017). Paläoökologie einer mergeligen Korallen-Assoziation aus dem Mitteldevon der Eifel (Hillesheimer Mulde), Rheinisches Schiefergebirge. Zitteliana, 89, 113–131.
Nützel, A. (2002). An evaluation of the recently proposed Palaeozoic gastropod subclass Euomphalomorpha. Palaeontology, 45(2), 259–266.
Oliver, W. A., Merriam, C. W., & Churkin, M. (1975). Ordovician, Silurian, and Devonian corals of Alaska. Geological Survey Professional Paper, 823B, 13–44.
Orbigny, A. d‘ (1850). Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés faisant suite au cours élémentaire de paléontologie et de géologie stratigraphiques. I. Paris: Masson.
Paris, F., Girard, C., Feist, R., & Winchester-Seeto, T. (1996). Chitinzoan bio-event in the Frasnian-Famennian boundary beds at La Serre (Montagne Noire, Southern France). Palaeogeography, Palaeoclimatology, Palaeoecology, 121, 131–145.
Pedder, A. E. H. (1964). Correlation of the Canadian Middle Devonian Hume and Nahanni formations by tetracorals. Palaeontology, 7(3), 430–451.
Philip, G. M. (1965). Lower Devonian Conodonts from the Tyers Area, Gippsland, Victoria. Proceedings of the Royal Society of Victoria, 79(1), 95–115, pls. 8–10.
Préat, A., & Mamet, B. (1989). Sédimentation de la plate-forme carbonatée givétienne franco-belge. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, 13(1), 47–86.
Préat, A., Blockmans, S., Capette, L., Dumoulin, V., & Mamet, B. (2007). Microfaciès d'une lentille biohermale à la limite Eifélien-Givétien ('Fondry des Chiens', Nismes, bord sud du Synclinorium de Dinant). Geologica Belgica, 10(1-2), 3–25.
Polenz, H. (2008). Eulenköpfe, Seelilien, und Flugsaurier. Ein Geo-Reiseführer zum Kalkstein, Kalk-Abbau, und zu den Fossilien des Hönnetals. Essen: Klartext Verlag.
Quenstedt, F. A. (1878–1881). Petrefactenkunde Deutschlands: 6. Die Röhren- und Sternkorallen. Leipzig: Fues’s Verlag.
Read, J. F. (1975). Tidal-Flat Facies in Carbonate-Cycles, Pillara Formation (Devonian), Canning Basin, Western Australia. In R. N. Ginsburg (Ed.), Tidal Deposits (pp. 251–256). Berlin, Heidelberg, New York: Springer.
Rietschel, S. (1968). Die Octactinellida und ihnen verwandte paläozoische Kalkschwämme (Porifera, Calcarea). Paläontologische Zeitschrift, 42(1), 13–32.
Rigby, J. K. (1991). The new Devonian (Givetian) heteractinid sponge Gondekia from Ontario, Canada, and evolution of the astraeospongiids and eiffeliids. Journal of Paleontology, 65(1), 38–44.
Rigby, J. K., Schumacher, D., & Meader, S. J. (1979). The genus Ensiferites, a Devonian astraeosponge of North America. Journal of Paleontology, 53(2), 475–493.
Roux, A. (1985). Introduction à l’étude des Algues fossiles paléozoïques (de la Bactérie à la tectonique des plaques). Bulletin des Centres Recherches Exploration – Production Elf-Aquitaine, 9/2,465–699.
Salerno, C. (2008). Stromatoporen-Fauna, Fazies und Paläoökologie von Plattformkarbonaten aus dem Unter-Givetium der Eifel (Devon, Rheinisches Schiefergebirge). Zitteliana, B27, 1–129.
Saupe, A. (2018). Biofazies-Analyse agglutinierender Foraminiferen entlang eines Transsekts von Mitteleuropa bis Nordafrika im mittleren bis oberen Famennium (Oberdevon). Unpublished M.Sc. Thesis, Westfälische Wilhelms-Universität Münster, 1–113.
Savarese, M., Dodd, J. R., & Lane, N. G. (1997). Taphonomic and sedimentologic implications of crinoid intraskeletal porosity. Lethaia, 29, 141–156.
Schlüter, C. (1885). Über einige neue Anthozoen aus dem Devon. Verhandlungen des Naturhistorischen Vereins der Preussischen Rheinlande und Westfalens, Sitzungsberichte, 42, 144–152.
Schlüter, C. (1889). Anthozoen des rheinischen Mitteldevon. Abhandlungen zur geologischen Special-Karte von Preussen und den thüringischen Staaten, 8(4), 259–465.
Schönlaub, H.-P., & Flajs, G. (1975). Die Schichtfolge der Nordwand der Hohen Warte (Mt. Coglians) in den Karnischen Alpen (Österreich). Carinthia II, 165/85, 83–96.
Schröder, S. (2005). Stratigraphie und Systematik rugoser Korallen aus dem Givetium und Unter-Frasnium des Rheinischen Schiefergebirges (Sauerland/Bergisches Land). Zitteliana, B25, 39–116.
Schudack, M. E. (1993). Karbonatzyklen in Riff- und Lagunenbereichen des devonischen Massenkalkkomplexes von Asbeck (Hönnetal, Rheinisches Schiefergebirge). Geologie und Paläontologie in Westfalen, 26, 77–106.
Schulze, F. E. (1875). Rhizopodenstudien 3. Archiv für Mikroskopische Anatomie, 11, 94–139.
Scrutton, C. T. (1998). The Palaeozoic corals, II: structure, variation and palaeoecology. Proceedings of the Yorkshire Geological Society, 52(1), 1–57.
Shinn, E. A. (1983). Birdseyes, fenestrae, shrinkage pores and loferites: a reevaluation. Journal of Sedimentary Petrology, 53(2), 619–628.
Söte, T., Hartenfels, S., & Becker, R. T. (2017). Uppermost Famennian stratigraphy and facies development of the Reigern Quarry near Hachen (northern Rhenish Massif, Germany). In B. Mottequin, L. Slavík, & P. Königshof (Eds.) Climate change and biodiversity patterns in the mid-Palaeozoic. Palaeobiodiversity and Palaeoenviroments, 97(3), 633–654.
Sparling, D. (1984). Paleoecologic and paleogeographic factors in the distribution of lower Middle Devonian conodonts from north-central Ohio. Geological Society of America, Special Paper, 196, 113–125.
Stadelmaier, M., Nose, M., May, A., Salerno, C., Schröder, S., & Leinfelder, R. R. (2005). Ästige tabulate Korallen-Gemeinschaft aus dem Mitteldevon der Sötenicher Mulde (Eifel): Faunenzusammensetzung und fazielles Umfeld. Zitteliana, B25, 5–38.
Steininger, J. (1831). Bemerkungen über die Versteinerungen, welche in dem Uebergangs-Kalkgebirge der Eifel gefunden werden. Trier: Gymnasium zu Trier.
Stewart, G. A., & Lampe, L. (1947). Foraminifera from the middle Devonian bone beds of Ohio. Journal of Paleontology, 21, 529–536.
Stichling, S. (2011). Stratigraphie und Fazies der Bohrung B102 im Raum Hönnetal (Nord-Sauerland). Unpublished B.Sc. Thesis, Westfälische Wilhelms-Universität Münster, 1–67.
Stichling, S., Becker, R. T., Hartenfels, S., & Aboussalam, Z. S. (2022). Drowning, extinction, and subsequent facies development of the Devonian Hönne Valley Reef (northern Rhenish Massif, Germany). In S. Hartenfels, C. Hartkopf-Fröder, & P. Königshof (Eds.), The Rhenish Massif: More than 150 years of research in a Variscan mountain chain. Palaeobiodiversity and Palaeoenvironments.https://doi.org/10.1007/s12549-022-00539-x. [this issue]
Stritzke, R. (1990). Die Karbonatsedimentation im Briloner Vorriffbereich. Geologisches Jahrbuch, D95, 253–315.
Struve, W. (1982). The Eifelian within the Devonian frame, history, boundaries, definitions. Courier Forschungsinstitut Senckenberg, 55, 401–432.
Struve, W. (1992). Neues zur Stratigraphie und Fauna des rhenotypen Mittel-Devon. Senckenbergiana lethaea, 71(5/6), 503–624.
Sun, Y., & Boucot, A. J. (1999). Ontogeny of Stringocephalus gubiensis and the origin of Stringocephalus. Journal of Palaeontology, 73(5), 860–871.
Suttner, T., & Hints, O. (2010). Devonian scolecodonts from the Tyrnaueralm, Graz Palaeozoic, Austria. Memoirs of the Australasian Palaeontologists, 39, 139–145.
Szaniawski, H. (2009). The earliest known venomonous animals recognized among conodonts. Acta Palaeontologica Polonica, 54(4), 669–676.
Szaniawski, H. & Drygant, D. (2014). Early Devonian Scolecodonts from Podolia, Ukraine. Acta Palaeontologica Polonica, 59(4), 967–983.
Thormann, F., & Weddige, K. (2001). Addendum zu Struve, W. (1992), Neues zur Stratigraphie und Fauna des rhenotypen Mittel-Devon: Abbildungen der Holotypen. Senckenbergiana lethaea, 81(2), 307–327.
Tonarová. P., Eriksson, M. E., & Hints, O. (2012). A jawed polychaete fauna from the late Ludlow Kozlowskii event interval in the Prague Basin (Czech Republic). Bulletin of Geosciences, 87, 713–732.
Tonarová, P., Hints, O., Königshof, P., Suttner, T. J., Kido, E., da Silva, A.-C., & Pas, D. (2016). Middle Devonian jawed polychaete fauna from the type Eifel area, western Germany, and its biogeographical and evolutionary affinities. Papers in Palaeontology, 2(2), 295–310.
Tsien, H. H. (1974). Espèces du genre Stringophyllum (Rugosa) dans le Dévonien moyen de la Belgique. Annales de la Société géologique de Belgique, 97, 257–271.
Tsien, H. H. (1982). Ancient reefs and reef carbonates. Proceedings of the Fourth International Coral Reef Symposium, Manila, 1981, Volume 1, 601–609.
Tsien, H. H. (1984). Organisms: Their ecology and function in carbonate construction. Palaeontographica Americana, 54, 415–420.
Urban, J. B. (1972). A reexamination of Chitinozoa from the Cedar Valley Formation of Iowa with observations on their morphology and distribution. Bulletins of American Paleontology, 63(275), 5–43.
Van Hinte, J. E., Ruffmann, A., Boogard, M., Jansonius. J., Kempen, T. M. G., Melchin, M. J., & Miller, T. H. (1995). Paleozoic microfossils from Orphan Knoll, NW Atlantic Ocean. Scripta Geologica 109, 1–63.
Versteegh, G. J. M., Servais, T., Streng, M., Munnecke, A., & Vachard, D. (2009). A discussion and proposal concerning the use of the term calcispheres. Palaeontology, 52(2), 343–348.
Walther, C. (1928). Untersuchungen über die Mitteldevon-Oberdevongrenze. Zeitschrift der deutschen geologischen Gesellschaft, A 80(2), 97–151.
Weddige, K. (1977). Die Conodonten der Eifel-Stufe im Typusgebiet und in benachbarten Faziesgebieten. Senckenbergiana lethaea, 58, 271–419.
Weddige, K. (1988). Eifel Conodonts. In W. Ziegler (Ed.), 1st International Senckenberg Conference and 5th European Conodont Symposium (ECOS V), Part 1: Guide to Field Trips. Field Trip A: Eifel Hills. Courier Forschungsinstitut Senckenberg, 102, 103–150.
Weddige, K., & Ziegler, W. (1976). The significance of Icriodus: Polygnathus ratios in limestones from the type Eifelian, Germany. In C. R. Barnes (Ed.), Conodont Palaeoecology. Geological Association of Canada, Special Papers, 15, 187–199.
Wedekind, R. (1925). Das Mitteldevon der Eifel. Eine biostratigraphische Studie. II. Teil. Materialien zur Kenntnis des mittleren Mitteldevons. Schriften der Gesellschaft zur Beförderung der Gesamten Naturwissenschaften, Marburg, 14(4), 1–85.
Webby, B. D., & Kershaw, S. (2011). Part E, Revised, Volume 4, Chapter 9B: External Morphology of the Paleozoic Stromatoporoidea: Shapes and Growth Habits. Treatise Online, 25, 1–73.
Wilson, J. L. (1975). Carbonate facies in geologic history (pp. 1–471). New York: Springer.
Young, G. A., & Kershaw, S. (2005). Classification and controls of internal banding in Palaeozoic stromatoporoids and colonial corals. Palaeontology, 48(3), 623–651.
Zhen, Y.-Y. (1996). Succession of coral associations during a Givetian transgressive-regressive cycle in Queensland. Acta Palaeontologica Polonica, 41(1), 59–88.
Zhou, X. (1980): Tabulata. In X. Siyuan, S. Wang, X. Zhou, J. Xiong, & T. Zhou (Eds.), Nandan typical stratigraphy and paleontology of Devonian, in South China (pp. 117–135, 158–161). Guizhou: Peoples Press [= Chow, 1980].
Ziegler, W. (1965). Zum höchsten Mitteldevon an der Nordflanke des Ebbesattels. Fortschritte in der Geologie von Rheinland und Westfalen, 9, 519–538.
Ziegler, W., & Lindström, M. (1971). Über Panderodus Ethington, 1959, und Neopanderodus n. g. (Conodonta) aus dem Devon. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 1971, 628–640.
Acknowledgements
We would like to thank the team of Rike Zimmerman (Münster), who helped by producing most of the thin sections. Furthermore, we like to thank Axel Munnecke (Erlangen) and Stephen Kershaw (London) for their very helpful reviews as well as Sven Hartenfels (Krefeld), Christoph Hartkopf-Fröder (Köln), and Sinje Weber (Frankfurt) for additional editorial comments. This publication is a contribution to IGCP 596 on “Climate change and biodiversity patterns in the Mid-Palaeozoic (Early Devonian to Early Carboniferous)”. We also like to thank Hans Martin Weber (Essen) for helping with the ostracods, Andrej Ernst (Hamburg) for helping with the bryozoans, and Petra Tonarová (Czech Geological Survey) for helping with the scolecodonts. Lastly, we would like to thank Lhoist Germany Rheinkalk GmbH who partially funded the research.
Data Availability Statement
All data generated during or analysed during the current study are included in this published article.
Funding
Open Access funding enabled and organised by Projekt DEAL. This paper was partially funded by the Lhoist Germany Rheinkalk GmbH, who financed the Ph.D. project of Sören Stichling.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue “The Rhenish Massif: More than 150 years of research in a Variscan mountain chain”
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Löw, M., Söte, T., Becker, R.T. et al. The initial phase of the Hönne Valley Reef at Binolen (northern Rhenish Massif, Middle Devonian). Palaeobio Palaeoenv 102, 573–612 (2022). https://doi.org/10.1007/s12549-022-00540-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-022-00540-4