
Abstract
A systematic-stratigraphic revision of selected large Late Turonian–Early Coniacian inoceramids of the lamarcki group from Germany showed that both historical specimens of Goldfuss (1836), introduced as Inoceramus annulatus and later becoming the types of Late Turonian I. lamarcki stuemckei Heinz, 1928a and Early Coniacian I. annulatus Goldfuss, 1836, respectively, are of Late Turonian age. I. stuemckei Heinz, 1928a, as emended herein, is well represented by its lectotype. Bulky isolated hinges from Upper Turonian strata, used by Heinz (1932a) to erect Heroceramus hercules, are just fragments of contemporaneous large I. stuemckei; H. hercules is thus invalid. The common Late Turonian I. stuemckei occurs facies-independent from near- to offshore settings and its first common appearance is an excellent marker for the substage base. I. annulatus Goldfuss, 1836 was later applied to very large forms first appearing in the mid-Early Coniacian. Thus, I. annulatus sensu Walaszczyk and Wood (1998) is poorly represented by its medium-sized latest Turonian lectotype, causing some nomenclatorial issues. I. stuemckei (heightmax 500 mm) and I. annulatus (heightmax one metre) are characterized by a stratigraphically abrupt shell enlargement and hinge buttressing across the Middle/Upper Turonian boundary and within the Early Coniacian, respectively. This shift to larger sizes, a common passive defense strategy against predation, is accompanied by widespread evidence for increased Late Turonian predation pressure by marine durophages. We thus speculate that the size increase in Late Turonian–Early Coniacian inoceramid bivalves is part of an escalating arms race between prey and predators, ultimately triggering Late Cretaceous inoceramid gigantism.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Very large, thick-shelled inoceramid bivalves with heights > 250 mm and a conspicuous ornament of coarse concentric rugae are a characteristic feature of Upper Turonian and Lower Coniacian strata of Europe, northern America and Asia (e.g., Sowerby 1823; Woods 1912; Tröger 1984; Walaszczyk and Wood 1998, 2018; Walaszczyk and Cobban 2000). In addition, isolated prisms of inoceramid shells more than 5 mm in length and large, massive calcitic hinges with shell thicknesses of several centimeters were found in abundance, especially in the Upper Turonian. For both inoceramid shells and hinges of large to very large sizes three different names are still in use today: Inoceramus annulatus is used for Lower Coniacian representatives (cf. Walaszczyk and Wood 1998; Walaszczyk and Wood in Niebuhr et al. 1999), Inoceramus lamarcki stuemckei for forms predominantly occurring in the Upper Turonian (cf. Ernst et al. 1983; Wood et al. 1984), and Inoceramus hercules for time-equivalent, isolated massive hinges and shell fragments (cf. Heinz 1928a; Wiese et al. 2004; Tröger and Niebuhr 2014).
The first nomenclatural treatment of the large inoceramids and the introduction of the species name Inoceramus annulatus dates back to Goldfuss (1836) for specimens from the “grüne und weiße Kreide in Westfalen” (Münsterland Cretaceous Basin, northern Germany). Geinitz (1839, 1849) adopted this name for both large shells and hinges from Upper Turonian strata of Saxony (eastern Germany) and Czechia. In the Upper Cretaceous section of Lüneburg (Lower Saxony, northern Germany), Heinz (1926, 1928a, c) differentiates between “Inoceramus latus Mant., var. stüm-ckei n. var.”, represented by Goldfuss’ specimen 7b (1836, pl. 110, fig. 7b; see Fig. 1b), which appears in the “Lamarcki- und untere Scaphitenschichten”, while “Inoceramus annulatus Goldf., part. Heinz”, represented by Goldfuss’ specimen 7a (1836: pl. 110, fig. 7a; see Fig. 1a), is characteristic for the “mittlere und obere Scaphitenschichten”. Furthermore, a large specimen from the English chalk (Woods 1912: text-figs. 82, 83) was assigned to “Megaloceramus monstrum nov. spec.” by Heinz (1932b), a name which is no longer valid (Tröger 2009). However, despite the taxonomic muddle it became evident that M. monstrum Heinz, I. latus var. stümckei Heinz (I. stuemckei in our sense), I. hercules (Heinz) and I. annulatus Goldf., part. Heinz are very closely allied forms (cf. Walaszczyk 1992).

Inoceramus stuemckei Heinz, 1928a. Reproduction of the Goldfuss specimens (1836), photographs of the plaster casts in the collection of the BGR, Geozentrum Hannover. a Goldfuss (1836, pl. 110, fig. 7a): Inoceramus ex aff. annulatus Goldf. according to Heinz (1926); BGR: 6750b (Bonn); plaster cast of an internal mould; “mittlere und obere Scaphitenschichten” according to Heinz (1926, 1928a: pl. 3), upper Upper Turonian. b Goldfuss (1836: pl. 110, fig. 7b): Inoceramus latus Mant., var. stümckei n. var. according to Heinz (1928a, c); BGR: 6750c (Bonn); plaster cast of an internal mould, partly with shell preservation; “Lamarcki- und untere Scaphitenschichten” according to Heinz (1928a: p. 72, pl. 3), lower and middle Upper Turonian
In addition, isolated large inoceramid hinges were described and/or figured by Mantell (1822), Sowerby (1823), Geinitz (1839), Woods (1912), Heinz (1932a, b), Tröger (1984) and Tröger and Niebuhr (2014), but their relationship to the large contemporaneous inoceramid shells remained unclear. The designation “Inoceramus Lamarcki var. Cuvieri, Sow.” of Woods (1912) includes large shells and hinges. Exclusively for the large hinges Heinz (1932a) introduced the genus “Heroceramus” with the type species “H. hercules” for findings from the “obere Scaphitenschichten” of Lüneburg, northern Germany, and inferred that the hinges belong to different subgenera, species and subspecies [“H. hercules valida”, “H. hercules tubiformis” and “H. (Athletoceramus) atlas”; Heinz 1932b]. Tröger (1984) combined separate findings of large composite moulds and massive calcitic hinges from the mid-Upper Turonian Strehlen Limestone of Saxony under the name “Inoceramus (Heroceramus) hercules Heinz” and agreed with the observation of Heinz (1932b) that different hinge types occurred in this stratigraphic interval.
The aim of this paper is the evaluation of the taxonomic relationships of the large inoceramid bivalves and hinges found in Upper Turonian to Lower Coniacian strata of Germany, also with respect to the validity and independence of the three taxa (Inoceramus annulatus, I. stuemckei and I. hercules) still used until today and belonging to the heterogenous lamarcki group. Furthermore, their stratigraphic ranges, based on associated ammonite and inoceramid occurrences, are calibrated and the functional morphology of trends in shell size and shell stabilization are discussed in the light of possible evolutionary patterns. The Saxonian Cretaceous is particularly suitable for this approach because of the availability of large Upper Turonian populations of I. stuemckei with sizes up to 450 mm, representing all ontogenetic stages, from both marly offshore (Figs. 5a, 7, 8, 9d) and sandy nearshore facies (Figs. 10, 11). I. annulatus (cf. Walaszczyk and Wood 1998), however, has not yet been recorded from Saxony so that occurrences from northern Germany have been investigated (Figs. 2, 12, 13). However, based on the regional bias on Saxonian faunas and in order to keep the paper short, we will focus on the large Upper Turonian inoceramids and do not detail the taxonomy and its issues with respect to I. annulatus.

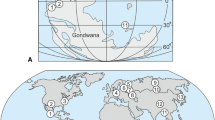
Palaeogeography of central Europe during the early Late Cretaceous (modified after Vejbæk et al. 2010, left) and simplified lithostratigraphy of the Plänerkalk and Elbtal groups (right). Sites with inoceramids studied herein are marked with green asterisks: 1. Münsterland Cretaceous Basin (incl. Goldfuss originals), 2. Staffhorst, 3. Helgoland, 4. Lüneburg, 5. Salzgitter, 6. Sachsen-Anhalt, 7. marly facies of the Elbtal Group (Dresden-Strehlen, Dresden-Räcknitz), 8. sandstone facies of the Elbtal Group (Reinhardtsdorf, Krippen, Königstein, Postelwitz), 9. Regensburg, Danubian Cretaceous Group. BCB Bohemian Cretaceous Basin, Döl./Pen. Fms Dölzschen and Pennrich formations, ESI East Sudetic Island, F./Fm Formation, Gr. Group, HTFm Hesseltal Formation, L.C. Lower Cretaceous, PKG Plänerkalk Group, SCB Saxonian Cretaceous Basin, WSI Westsudetic Island
Short geological overview
All studied sites are located in northern Germany and are part of the wide warm-temperate epicontinental Boreal shelf sea that covered large parts of western and central Europe north of the Mid-European Island during the Late Cretaceous (e.g., Vejbæk et al. 2010; Fig. 2). Additionally, inoceramid occurrences south of the Mid-European Island from the Danubian Cretaceous Group near Regensburg, Bavaria (site 9 in Fig. 2) have been integrated into the study. The roughly coast-parallel facies zonation on the low-angle, ramp like North German shelf comprises inner shelf sandstones and/or greensands, followed by mid-shelf marls and marl-limestone alternations, finally grading into outer shelf white limestones and chalks (Wilmsen et al. 2005; Niebuhr 2006). The Cenomanian to Lower Coniacian succession in the Münsterland, in Lower Saxony and the Subhercynian area (sites 1–6 in Fig. 2) are dominated by calcareous offshore sediments combined in the Plänerkalk Group (Wilmsen and Hiss in Niebuhr et al. 2007). In the Saxonian Cretaceous Basin, siliciclastic input from the eastern part of the Mid-European Island (Bohemian Massif) and the Westsudetic Island was higher and a sandy nearshore facies zone (today forming the Saxonian Switzerland) in the southeast grades into a marly–calcareous offshore facies zone in the northwest (Meißen–Dresden area; sites 7 and 8 in Fig. 2). The strata in Saxony are combined in the Elbtal Group (Voigt and Tröger in Niebuhr et al. 2007; Fig. 2), the marly offshore strata of which are now united in the Meißen-Dresden Subgroup (Wilmsen et al. 2019a). Via a former strait, the Saxonian Cretaceous Basin was directly connected to the Bohemian Cretaceous Basin in the southeast that opened towards the northern margin of the Tethyan Ocean (Fig. 2). For an overview of the current lithostratigraphy in the studied basins, see Hiss et al. (2018).
Material and methods
The material studied herein stems from different scientific collections that are identified by their representative acronyms: MMG: Museum für Mineralogie und Geologie of the Senckenberg Naturhistorische Sammlungen Dresden (SNSD, specimens MMG: Sak, MMG: TUD and MMG: CsK); BGR: Bundesanstalt für Geowissenschaften und Rohstoffe, Geozentrum Hannover; FG: Institut für Geologie, Technische Universität Bergakademie Freiberg (TU BAF); JME: Jura-Museum Eichstätt; pc: private collection.
Standard measurements for inoceramid bivalves, taken with a Vernier caliper, comprise height (h) and length (l) of the shell as well as observations on the sculpture (number, size and form of ribs) and the course of the growth axis (for terminology and measured parameters, see Fig. 3). For the maximum height of the shells, the following size classes are used: < 5 cm = small, 5–15 cm = medium-sized, 15–25 cm = large, 25–50 cm = very large, 50–100 cm = huge, and > 100 cm = gigantic (modified according to Tröger and Niebuhr 2014). For the higher rank systematics of the Bivalvia, we refer to Carter et al. (2011).

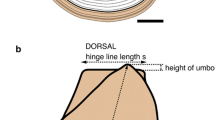
Terminology and biometric parameters of the inoceramid shell; AA = apical angle, * according to Heinz (1928b)
Systematic palaeontology
Family Inoceramidae Giebel, 1852
Genus Inoceramus Sowerby, 1814
Inoceramus stuemckei Heinz, 1928a
Figures 1, 4, 5, 6, 7, 8, 9, 10, 11
-
1822Inoceramus Cuvieri—Mantell: p. 213 (partim), pl. 27, fig. 4; pl. 28, fig. 1 [from the Terebratulina lata/Sternotaxis plana zonal boundary, Upper Turonian; Kelly and Wood (1996)].
-
? 1823Inoceramus Brongniarti—Sowerby: p. 60, pl. 441, figs. 2, 3.
-
1836Inoceramus annulatus nobis—Goldfuss: p. 114 (partim), pl. 110, fig. 7b [non pl. 110, fig. a].
-
1839Inoceramus annulatus Goldf. —Geinitz: p. 26.
-
non 1842bInoceramus annulatus Goldf.—von Hagenow: p. 558.
-
1849Inoceramus annulatus Goldf.—Geinitz: p. 174.
-
1870Inoceramus Brongniarti Sow.—Roemer: pl. 34, fig. 13.
-
1873Inoceramus Brongniarti Mant.—Geinitz: p. II.43 (partim), pl. II.11, figs. 7, 8.
-
? 1897Inoceramus annulatus Goldfuss—Leonhard: p. 48.
-
non 1912Inoceramus lamarcki var. Cuvieri, Sow.—Woods: p. 320 (partim), text-figs. 73–76, 79.
-
1912Inoceramus Lamarcki var. Cuvieri, Sow.—Woods: p. 320 (partim), text-figs. 77, 78, 80–84 [text-fig. 84 = Mantell (1822: pl. 28, fig. 1)].
-
1921Inoceramus Cuvieri var. annulata Goldf.—Seitz: p. 104.
-
non 1926Inoceramus ex aff. annulatus Goldf.—Heinz: p. 99 [= Goldfuss (1836, pl. 110, fig. 7a)].
-
* 1928aInoceramus latus Mant., var. stümckei n. var.—Heinz: p. 72, pl. 3.
-
non 1928aInoceramus annulatus Goldf., part. Heinz—Heinz: pl. 3.
-
1932aHeroceramus hercules n. g. n. sp.—Heinz: p. 34.
-
1932bMegaloceramus monstrum nov. spec.—Heinz: p. 7 [= Woods (1912: text-figs. 82, 83)].
-
1932bHeroceramus hercules Heinz—Heinz: p. 9.
-
1932bHeroceramus hercules Heinz var. valida nov. var.—Heinz: p. 9.
-
1932bHeroceramus hercules Heinz var. tubiformis nov. var.—Heinz: p. 9 [= Woods (1912: text-fig. 80)].
-
1932bHeroceramus (Athletoceramus) atlas nov. subgen. nov. spec.—Heinz: p. 9.
-
1932Inoceramus (Megaloceramus) ex. aff. monstrum Heinz—Wolansky: p. 27, pl. 4, fig. 1.
-
1932Inoceramus (Megaloceramus) aff. stümckei Heinz—Wolansky: p. 27, pl. 4, fig. 7.
-
1932Inoceramus (Boehmiceramus) sp.—Wolansky: p. 29, pl. 5, fig. 1.
-
1939Inoceramus (Orthoceramus) lamarcki Park.—Dacqué: p. 104 (partim), pl. 5, fig. 6.
-
1960Inoceramus cuvieri var. annulata Goldf.—Tröger and Wolf: p. 296, fig. 5.
-
1962Inoceramus annulatus Goldfuss part Heinz 1928—Bräutigam: p. 205, pl. 3, fig. 4.
-
1967Inoceramus lamarcki stümckei Heinz, 1928—Tröger: p. 67, pl. 5, fig. 9; pl. 6, fig. 6.
-
? 1982Inoceramus lamarcki stümckei Heinz, 1928—Keller: p. 82, pl. 6, fig. 1.
-
1984Inoceramus (Heroceramus) hercules Heinz—Tröger: p. 47, pl. 1, figs. 4–4b; pl. 2.
-
1984Inoceramus (Heroceramus) ?hercules Heinz—Tröger: p. 48, pl. 1, fig. 2.
-
1984Inoceramus sp. aff. annulatus Goldfuss—Tröger: p. 49, pl. 1, figs. 1, 1a.
-
1984Inoceramus sp. (n. sp.?) ex gr. Inoceramus lamarcki Parkinson—Tröger: p. 49, pl. 1, figs. 3, 3a.
-
1987Inoceramus (Heroceramus) hercules Heinz—Schloß—Tröger: p. 207, fig. 1.
-
1987Inoceramus (Heroceramus) ?hercules Heinz—Tröger: p. 207, fig. 1.
-
1987Inoceramus sp. (n. sp.?) ex gr. lamarcki Parkinson—Schloß—Tröger: p. 207, fig. 1.
-
1987Inoceramus lamarcki stuemckei Heinz—Cleevely and Morris: fig. 6.4a.
-
? 1988Inoceramus lamarcki stuemckei Heinz—Walaszczyk: pl. 4, fig. 1.
-
1988Inoceramus lusatiae Andert—Walaszczyk: pl. 5, fig. 8.
-
1991Inoceramus (Heroceramus) cf. hercules Heinz, 1932—Tröger and Christensen: p. 30, pl. 1, fig. 17; pl. 3, fig. 4.
-
1992Inoceramus lamarcki stuemckei Heinz—Walaszczyk: p. 31, pl. 9, fig. 6.
-
1996Inoceramus lamarcki stuemckei—Tarkowski: p. 490.
-
1999 large Inoceramus ex gr. lamarcki—Walaszczyk and Wood in Niebuhr et al.: p. 188, fig. 6.
-
2000Inoceramus cf. stuemckei Heinz, 1928—Walaszczyk and Cobban: p. 46, pl. 8, fig. 7; pl. 9, fig. 5; pl. 10, fig. 7.
-
2002Inoceramus lamarcki stumckei Heinz—Cleevely and Morris: text-fig. 7.2A
-
2004Inoceramus hercules Heinz—Wiese et al.: fig. 5d.
-
2009Inoceramus sp. aff. cuvieri Sowerby, 1814—Tröger et al.: p. 69, fig. 7C [= Dacqué (1939: pl. 5, fig. 6)].
-
2009Inoceramus lamarcki stuemckei Heinz, 1928—Tröger et al.: p. 71, figs. 10A, 10E.
-
2009Inoceramus lamarcki Parkinson, 1818 subsp. indet.—Tröger et al.: p. 73 (partim), fig. 7H.
-
2013Inoceramus lamarcki stuemckei Heinz, 1926—Walaszczyk et al.: pl. 2.
-
2014Inoceramus lamarcki stuemckei Heinz, 1928—Tröger and Niebuhr: p. 178, fig. 4c.
-
2014Inoceramus hercules (Heinz, 1932)—Tröger and Niebuhr: p. 179, fig. 4a.
-
2014 „Inoceramus gr. woodsi Jukes-Browne in Ivannikov, 1979”—Delvaque et al.: pl. 45, fig. 1.
-
2014Inoceramus sp.—Delvaque et al.: pl. 45, fig. 7.
-
2019bInoceramus lamarcki stuemkei—Wilmsen et al.: fig. 9.
Type. The lectotype of Inoceramus stuemckei by subsequent designation of Heinz (1928c: p. 75) is the original of Ino-ceramus annulatus of Goldfuss (1836: pl. 110, fig. 7b).


Inoceramus stuemckei Heinz, 1928a. a MMG: SaK 8768; left valve with parts of the wing preserved and only a few but strong adult rugae, 180 mm in height; Räcknitz Formation, (7) Dresden-Räcknitz, Saxony; I. inaequivalvis/I. stuemckei/I. perplexus Zone, lower Upper Turonian. b FG: 690–139/90; right valve with parts of the shell preserved, note conspicuous bend in growth axis, 130 mm in height, b1, outer view, b2 = anterior view; Salder Formation, (5) Salzgitter-Salder, Lower Saxony; M. striatoconcentricus Zone, mid-Upper Turonian; Hyphantoceras Event according to Tröger (in sched., leg. 1990)

Inoceramus stuemckei Heinz, 1928a. BGR: Ma 14106; type 2, most parts of the shell preserved; note the wide, regularly spaced, round-topped concentric rugae covered by fine, regularly spaced, raised growth lines (“Anwachsringreifen”), 240 mm in height; Salder Formation, (2) 572–575 m depth in the Staffhorst shaft, Lower Saxony; I. inaequivalvis/I. stuemckei/I. perplexus Zone, lower Upper Turonian

Inoceramus stuemckei Heinz, 1928a. FG: 346/1; type 2, left valve with parts of the shell preserved, the transition of the anterior margin to the beak shows a sharp kink, 480 mm in height; Strehlen Limestone of the lower Strehlen Formation, (7) former quarry in Dresden-Strehlen, Saxony; M. labiatoidiformis/M. striatoconcentricus Zone, mid-Upper Turonian; original and neotype of Inoceramus (Heroceramus) hercules Heinz of Tröger (1984: pl. 2)

Inoceramus stuemckei Heinz, 1928a. MMG: SaK 8417; left valve with parts of hinge and shell preserved, 165 mm in height; a1, outer view of the composite mould, a2, inner view with large parts of the massive calcitic hinge still in contact with the shell; Strehlen Limestone of the lower Strehlen Formation, (7) Weinböhla, Saxony; M. labiatoidiformis/M. striatoconcentricus Zone, mid-Upper Turonian

Massive calcitic hinges and shell preservation of Inoceramus stuemckei Heinz, 1928a. a–d Strehlen Limestone of the lower Strehlen Formation, (7) former quarry in Dresden-Strehlen, Saxony; M. labiatoidiformis/M. striatoconcentricus Zone, mid-Upper Turonian. a MMG: SaK 9609; large hinge with eight grooves on a length of 15 mm preserved; a1, large prisms up to 25 mm thick visible in cross-section, a2, lower side; original of Inoceramus (Heroceramus) hercules Heinz of Tröger (1981, pl. 1, fig. 4, 4a), Inoceramus sp. (n. sp.?) ex gr. Inoceramus lamarcki Parkinson of Tröger (1984: pl. 1, fig. 3, 3a) and Inoceramus hercules (Heinz, 1932a, b) of Tröger and Niebuhr (2014, fig. 4a). b FG: 346/2; massive hinge with 23 preserved grooves on a length of 42 mm, lower and upper side; original and neotype of Inoceramus (Heroceramus) hercules Heinz of Tröger (1984: pl. 1, fig. 4, 4a). c MMG: SaK 9640; c1, upper side of a hinge encrusted by small oysters; note growth lines of the outer shell, c2, large prisms of the hinge up to 14 mm thick and small prisms of the shell up to 2 mm thick visible in the cross-section. d MMG: SaK 9590; small hinge with 13 preserved grooves on a length of 25 mm; original of Inoceramus Brongniarti Sow. of Geinitz (1873: pl. II.11, fig. 7). e MMG: NwK 843; fragments of thick-shelled inoceramid valves partly in contact from the “grüne Kreide von Westphalen” of Goldfuss (1836); Soest Greensand Member of the Duisburg Formation, (1) Anröchte-Klieve, Münsterland Cretaceous Basin; M. labiatoidiformis/M. striatoconcentricus Zone, mid-Upper Turonian

Inoceramus stuemckei Heinz, 1928a. FG: 690-871/96; left valve with fine growth lines, parts of the wing and imprint of the hinge are preserved, 190 mm in height; Sandstone a3 of the Postelwitz Formation, (8) former quarries of Bad Schandau-Postelwitz, Saxony; I. inaequivalvis/I. stuemckei/I. perplexus Zone, lower Upper Turonian

Inoceramus stuemckei Heinz, 1928a. MMG: SaK 6620a + b; hypo- and epirelief of a shell bed with at least five juvenile specimens, specimens I and II are 110 mm in height; a1 (left), imprints of specimens I and II show nicely preserved “Anwachsringreifen” of the inner shell and the completely preserved wing (specimen I), a2 (right), on the composite moulds of specimens I and II only the strong rugae are visible; Sandstone a3 of the Postelwitz Formation, (8) former quarry Reinhardtsdorf 1 in Reinhardtsdorf-Kleingießhübel, Saxony; I. inaequivalvis/I. stuemckei/I. perplexus Zone, lower Upper Turonian
Stratum typicum. “Lamarcki- und untere Scaphitenschichten” according to Heinz (1928a: p. 72, pl. 3), which equals to the I. perplexus/I. stuemckei/I. inaequivalvis and M. striatoconcentricus/M. labiatoidiformis zones (lower and middle Upper Turonian).
Locus typicus. Exact place unknown, “grüne und weiße Kreide in Westfalen” according to Goldfuss (1836: p. 114), Münsterland Cretaceous Basin, northern Germany.
Material. Germany, Nordrhein-Westfalen—BGR: plaster casts of both Goldfuss specimens (1836: pl. 110, figs. 7a, b), no. 6750 (Bonn) and no. 6750b (Bonn); MMG: NwK 381 and NwK 843 from Salzkotten-Niederntudorf and Anröchte-Klieve. Lower Saxony—BGR: Ma 14105, Ma 14106 and further unregistered specimens from the Staffhorst shaft between 534 and 575 m, unregistered specimen from Salzgitter-Voßpaß; FG: 690-139/90. Sachsen-Anhalt—FG: 690-295/99 from Osterwiek-Hoppenstedt; MMG-TUD: Pz 1398 from the Spiegelsberge south of Halberstadt. Saxony—MMG: SaK 6513a–f, 6569, SaK 6600, SaK 6620a–c, SaK 8117, SaK 8412, SaK 8414, SaK 8417, SaK 8768, SaK 9000, SaK 9590, SaK 9609, SaK 9612, SaK 9639, SaK 9640, SaK 9642, SaK 12777, SaK 12779, SaK 12812, SaK 16109, and at least 25 specimens from the Reinhardtsdorf 2 quarry near Reinhardtsdorf-Schöna, for example MMG: SaK 12891a + b, SaK 12892, SaK 12892, SaK 12903, SaK 12905–12910, SaK 12916, SaK 13372, SaK 14911, SaK 15646, SaK 15647a + b, SaK 15171a + b, SaK 16795; FG: 346/1, 346/2, 690-754/67, 690-759/67, 690-764/67, 690-139/90, 690-871/96. Bavaria—JME: 91 1961 130, pc-Pförringer, pc-Niebuhr/Wilmsen.
Czechia—MMG: CsK 470, CsK 1036, CsK 1803, CsK 2445.
Description. Inoceramus stuemckei is a large to very large, relatively flat, distinctly ribbed, almost equivalved species with a maximum height and length of 480 mm and 360 mm (FG: 346/1 on Fig. 7), respectively. It is characterized by wide, more-or-less regularly spaced, rounded commarginal concentric rugae (“Anwachsreifen” according to Heinz 1928b: p. 18, 1928c: p. 75), running perpendicular to the straight to slightly convex growth axis. The rugae are asymmetric, i.e., the slopes directed to the umbo are shallower compared to the ventrally directed ones. The weakly prosogyre umbo is strongly vaulted and carries small, densely spaced rugae, the distance and height of which constantly increase during ontogeny. Rugae are 15 mm high and separated by inter-rugae spaces up to 40 mm wide on the adult part of the shell in specimens larger than 300 mm. The large, prolate wing has an almost straight hinge line with a broad hinge plate. The general outline is rounded-rectangular.
Two basic types can be differentiated among the studied population of Inoceramus stuemckei. Type 1 comprises forms with very strong, rounded to narrowly rounded concentric rugae. Juvenile stages are sturdily convex and thus well differentiated from slightly older I. lamarcki; adult shell parts are only weakly convex. Type 1 forms are characterized rather by box-shaped than circular ventral margins. In outline, they are thus almost symmetrically pentagonal or triangular-oval. Shells, external imprints and favourably preserved internal moulds have regular accretionary rings (growth rings) that also cross the entire wings. All specimens from the sandy nearshore facies in Saxony are members of type 1 and also the marly facies is predominated by this type.
Type 2 forms are very rare and relatively flat (i.e., weakly convex), characterized by only a few, rather weak, shallow, rounded concentric rugae that tend to be strongest in the juvenile and adult growth stages. Shells have regular growth rings while internal moulds are smooth. Their outline is elongated to asymmetric-triangular, resembling in form, but not in sculpture, the lectotype of Goldfuss (1836: pl. 110, fig. 7b). Examples of type 2 forms are BGR: Ma 14106 from the Staffhorst shaft section between 572 and 575 m (Fig. 6) and the specimen FG: 346/1 from the Strehlen Limestone of Saxony (Fig. 7).
The hinges of large specimens (Figs. 8, 9), introduced by Heinz (1932a: p. 34) as Heroceramus hercules, are of great thickness, and isolated segments of them are often found separately. The hinges are thickest near the umbo and become thinner towards the posterior end. The ligament pits are numerous, shallow, and several times higher than wide, reaching their maximum height not far from the umbo. Irrespective of the size of the hinge, they are always ~ 2 mm wide, resulting in eight grooves on a length of 15 mm (MMG: SaK 9609; Fig. 9a), 13 grooves on a length of 26 mm (MMG: SaK 9590; Fig. 9d), and 23 grooves on a length of 42 mm (FG: 690-S4/84; Fig. 9b).
The calcitic shell and the massive hinges are only preserved in specimens from calcareous and calcareous marl facies (e.g., Strehlen Limestone of the lower Strehlen Formation in Saxony, Tröger 1984; Hundorf Limestone of the lower Teplice Formation in Czechia, Wiese et al. 2004; Walliser et al. 2020) or calcareous glauconitic sandstones (e.g., Soest Greensand Member of the Duisburg Formation, Münsterland Cretaceous Basin; see MMG: NwK 843, Fig. 9e). The shell thickness increases with distance from the umbo from 0.5 to 3.5 mm. The shell shows fine, regularly spaced, slightly raised growth lines (“Anwachsringreifen”, cf. Heinz 1928b: p. 19) that are between ~ 2 and 5 mm wide. In the sandstone facies, these raised growth lines are usually preserved only in external imprints while moulds are characterized by simple rugae in most cases. Only in exceptional cases they are preserved on sandstone moulds (Fig. 10). Specimen MMG: SaK 6622a + b from the sandstone facies of the Saxonian Switzerland shows both preservation types: moulds with simple, regularly spaced, rounded concentric rugae (Fig. 11a2) and external imprints of the same specimens with rugae covered by fine growth lines (Fig. 11a1). This indicates that the occurrence of the raised growth lines in imprints and moulds is related to different preservation forms and not a characteristic feature for specific inoceramid taxa such as the Lower Coniacian species Inoceramus annulatus (see “Discussion” below).
Remarks. In contrast to former authors (e.g., Tröger 1967; Keller 1982) we regard Inoceramus stuemckei Heinz as an individual species, morphologically sufficiently distinct from Inoceramus lamarcki Parkinson (also Walaszczyk and Cobban 2000: p. 46 regard Heinz’s taxon as a species). Parkinson’s species is generally smaller than 150 mm in height (i.e., medium-sized), and the shells are more convex with strongly domed umbo and dorso-ventrally elongated, characterized by weaker sculpture of shallow, oval rugae. Furthermore, both taxa are stratigraphically largely discrete, I. lamarcki characterizing the (late) Middle Turonian and I. stuemckei, after its appearance in the late Middle Turonian, is dominating in the early to mid-Late Turonian. Nevertheless, both species are closely allied.
The species Heroceramus hercules was introduced by Heinz (1932a: p. 34) for large, massive calcitic inoceramid hinges from the “obere Scaphitenschichten” (upper Upper Turonian) in the Lüneburg section, northern Germany (Heinz 1932b). Additional findings of such large-scale hinges are known, e.g., from the mid-Upper Turonian Strehlen Limestone of the lower Strehlen Formation, Saxony (Figs. 9a–d; Geinitz 1839, 1873; Tröger 1984; Tröger and Niebuhr 2014) and the time-equivalent Hudcov (Hundorf) Limestone of the lower Teplice Formation, Czechia (e.g., Wiese et al. 2004). Tröger (1984: pl. 2) designated both an isolated hinge and a large composite mould from the Streh-len Limestone with parts of the shell preserved as neotypes for Inoceramus (Heroceramus) hercules (Figs. 7, 9b). However, we consider I. (H.) hercules Heinz, 1932a as a younger subjective synonym of Inoceramus stuemckei Heinz, 1928a. As can be shown by conjugate occurrences [e.g., specimen MMG: Sak 8417 from the mid-Upper Turonian Strehlen Limestone at Weinböhla (Fig. 8) in which the hinge is in contact with the composite mould, or the specimen illustrated by Tröger and Christensen (1991: pl. 3, fig. 4)], the massive hinges and hinge plates belong to shells of I. stuemckei and are not remains of another large species.
In Tröger and Wolf (1960) “Inoceramus cuvieri var. annulata Goldfuss” and “Inoceramus latus Man. var. stümckei Heinz” from the marls of the upper Räcknitz Formation a few meters below the Strehlen Limestone are described as synonyms. It should be noted that there is in fact some taxonomic confusion concerning the separation of the two species and that we regard the hitherto accepted lectotype of Inoceramus annulatus [Goldfuss 1836, pl. 110, fig 7a) from the “mittlere und obere Scaphitenschichten” (Heinz 1926, 1928a: pl. 3; middle to upper Upper Turonian) as a possible representative of I. lusatiae Andert, 1911 (see “Discussion” below). Also, the medium-sized Inoceramus annulatus Goldfuss part Heinz 1928 of Bräutigam (1962: pl. 3, fig. 4), from his lower vancouverensis Zone at Langelsheim, Harz foreland near Goslar, ca. 30 m above the first appearance of the zonal index Inoceramus perplexus and ca. 20 m below the FO of Mytiloides striatoconcentricus (Bräutigam 1962; app. 7), is questionable. This level equals to the I. perplexus/I. stuemckei/I. inaequivalvis Zone, far below the first appearance of Lower Coniacian I. annulatus in the sense of Walaszczyk and Wood (1998), and we thus include it in I. stuemckei. The specimen of Wolanski (1932: p. 29, pl. 5, fig. 1), however, described as Boehmiceramus sp./Inoceramus (Boehmiceramus) sp. from Upper Turonian limestones of Wusterhusen-Gustebin, Vorpommern [not “Mucronatensenon”, see Deecke (1908)], and labeled by von Hagenow as Inoceramus annulatus Goldf., is a typical I. stuemckei due to the presence of wide, regularly spaced, round-topped concentric rugae covered by conspicuous, regularly spaced, raised growth lines which are not developed in the “Senonian” subfamily Boehmiceraminae of Heinz (1932b: p. 18).
The large, allegedly Coniacian species Inoceramus sachsi Bodylevsky, 1958 (p. 79, pl. 30, fig. 1; pl. 32, figs. 1a, 6; pl. 33, fig. 1) is similar to I. stuemckei and, according to the stratigraphic revision of its stratum typicum in Taymyr (western Sibiria), of Late Turonian age (Walaszczyk and Cobban 2000: p. 50), too, such as the North American records reported therein. I. sachsi may well turn out to as a younger subjective synonym of Heinz’s species but a revision of Bodylevsky’s taxon is beyond the scope of this paper.
Stratigraphy. Inoceramus stuemckei (including I. hercules) is the oldest inoceramid species of Late Cretaceous age (early to mid-Late Turonian) with very large size (> 250 mm in height) and conspicuously thickened shells equipped with massive hinges. Following its first appearance in the late Middle Turonian, it ranges from the basal Upper Turonian I. perplexus/I. stuemckei/I. inaequivalvis assemblage zone in which very large forms dominate the inoceramid record (Walaszczyk and Cobban 2000; 1st acme, see Fig. 14) to the top of the Mytiloides striatoconcentricus/M. labiatoidiformis concurrent range zone (2nd acme, see Fig. 14). In the Staffhorst shaft section both the last occurrence of I. stuemckei and the first occurrence of M. scupini fall into a three-metre-thick interval (534–537 m depth, Walaszczyk and Wood in Niebuhr et al. 1999; see Fig. 12). Associated with the frequent occurrences of I. stuemckei in Saxony are typical Late Turonian inoceramid faunas such as Inoceramus perplexus Whitfield, I. inaequivalvis Schlüter, smooth and large representitives of the lamarcki group, Mytiloides mytiloidiformis (Tröger), M. dresdensis (Tröger), M. striatoconcentricus (Gümbel), M. incertus (Jimbo), M. turonicus Walaszczyk and M. labiatoidiformis (Tröger) (e.g., Tröger and Niebuhr 2014; Niebuhr and Wilmsen, in prep.) as well as the ammonites Lewesiceras mantelli Wright and Wright, Subprionocyclus neptuni (Geinitz), S. branneri (Anderson), Allocrioceras billinghursti Klinger, A. strangulatus Wright, Eubostrychoceras saxonicuum (Schlüter), Hyphantoceras reussianum (d´Orbigny), Sciponoceras bohemicum (Fritsch) and Scaphites geinitzi d´Orbigny (e.g., Wilmsen and Nagm 2014).

The three known localities in northern Germany where both very large Inoceramus stuemckei and huge Inoceramus annulatus were found in-situ (for locations see Fig. 2); note occurrences compared to the FOs and LOs of additional inoceramid bivalves. (2) Staffhorst shaft section (see Walaszczyk and Wood in Niebuhr et al. 1999; Walaszczyk and Wood 2018); (4) Lüneburg section (see Heinz 1928a); (5) Salzgitter-Salder section (see Wood and Ernst 1998; Walaszczyk and Wood in Niebuhr et al. 1999; Walaszczyk et al. 2021); Cww = C. waltersdorfensis waltersdorfensis, Ccc = C. crassus crassus, Cdd = C. deformis deformis
Palaeogeographic occurrence. Central and East Europe (very common), Middle East (Iran) to (Central) Asia, northern America. The following occurrences in Europe can be specified:
Germany. Sachsen: Königstein, Kleingießhübel, Postelwitz and Reinhardtsdorf-Schöna = Sandstone a3 of the Postelwitz Formation; Nikolsdorf = Sandstone c of the Postelwitz Formation; Dresden-Räcknitz = Räcknitz Formation; Weinböhla, Dresden-Strehlen and Dresden-Zschertnitz = lower and middle Strehlen Formation. Sachsen-Anhalt: Osterwiek-Hoppenstedt and Halberstadt = upper Söhlde and Salder Formation. Mecklenburg-Vorpommern: Wusterhusen-Gustebin = Salder Formation. Niedersachsen: Staffhorst, Lüneburg, Salzgitter-Salder, Salzgitter-Bad = Salder Formation. Nordrhein-Westfalen: Ahaus, Halle = Wüllen Formation; Salzkotten-Niederntudorf = Oerlinghausen Formation; Anröchte-Klieve = Duisburg Formation; Lengerich = Lengerich Formation. Bayern: Regensburg = Kagerhöh Formation.
Czechia. Volavec (central Bohemian Cretaceous Basin), Teplice, Litomĕřice, Úpohlavy (northwestern BCB) = lower Teplice Formation.
Poland. Lubin/Lebbin (island of Wolin, western Pomerania); Opole Cretaceous Basin.
France. Massif d’Uchaux.
England. Yorkshire to Kent = lower Upper Chalk.
Discussion
As outlined in the taxonomic part, there is only one very large-sized inoceramid species in Upper Turonian of northern Germany, i.e., Inoceramus stuemckei Heinz, 1928a. It is followed by another very large to huge Early Coniacian species, i.e., Inoceramus annulatus sensu Walaszczyk and Wood (1998). Their stratigraphic distribution and possible evolutionary ties as well as the functional morphology of shell traits and inferred modes of life are discussed below.
Stratigraphic ranges of selected species of the lamarcki group
The lamarcki group comprises a rather heterogeneous complex of about 50 species and subspecies (many of which probably synonymous) with poorly understood phylogentic relationships that appeared in the Middle Turonian and ranged into the basal Upper Coniacian (Walaszczyk and Cobban 2000: p. 50). Taxa of the group occur all over the northern hemisphere and show considerable morphological plasticity. Herein, we focus on large to huge Upper Turonian to Lower Coniacian representatives from selected sections in northern Germany.
There are three sections in Lower Saxony where both Inoceramus stuemckei and I. annulatus were found (Fig. 12): the former quarry at Lüneburg, study area of Rudolf Heinz, which is inaccessible since a long time (site 4 in Fig. 2), the Staffhorst shaft section, from which the rich assemblage of inoceramid bivalves is housed in the BGR collection (site 2 in Fig. 2), and the famous quarry section in Salzgitter-Salder, GSSP for the base of the Coniacian (Walaszczyk et al. 2021, site 5 in Fig. 2).
The range of Inoceramus stuemckei in northern Germany is fairly well constrained, comprising the basal Upper Turonian up to the first appearance of M. scupini in the upper Upper Turonian. An overlap of small- to medium-sized Inoceramus lamarcki (< 150 mm in height) and large to very large Inoceramus stuemckei (commonly > 250 mm in height) in the Middle–Upper Turonian boundary interval is proposed by Walaszczyk (1988, 1992, pers. commun. May 2021), but not seen in our material. In the Staffhorst shaft section, the lowermost Upper Turonian I. perplexus/I. stuemckei/I. inaequivalvis Zone is differentiated in a lower part without, and an upper part with Inoceramus perplexus (Walaszczyk and Wood in Niebuhr et al. 1999; see Wiese et al. 2020 for an alternative Upper Turonian inoceramid biostratigraphy based on interval zones only); however, very large Inoceramus stuemckei, such as specimen BGR: Ma 14106 (Fig. 6), are a characteristic feature of the lower Upper Turonian I. perplexus/I. stuemckei/I. inaequivalvis Zone (Ernst et al. 1983; Wood et al. 1984; Walaszczyk and Wood in Niebuhr et al. 1999; Kaplan 2011). Also in Saxony, thin-shelled I. perplexus, limited to the offshore marl and Pläner facies, are associated with large I. stuemckei. Thus, the appearance of common large I. stuemckei in Sandstone a3 of the Postelwitz Formation in the nearshore zone (Saxonian Switzerland; cf. Wilmsen and Niebuhr 2009, 2014a) is considered as simultaneous, i.e., marking the base of the Upper Turonian in the shallow-water sandstone facies where the thin-shelled index inoceramid I. perplexus is missing. This is also supported by the fact that below this level, also in the sandstone facies of the Saxonian Switzerland, the lamarcki group is only represented by small- to medium-sized Inoceramus lamarcki ssp. which usually do not exceed a height of 150 mm, being a feature of the upper Middle Turonian of central Europe (Walaszczyk and Wood in Niebuhr et al. 1999). The upper limit of the stratigraphic range of I. stuemckei is demonstrated in the Staffhorst shaft section where the first appearance of M. scupini is associated with the last occurrence of large Inoceramus ex gr. lamarcki and I. ex gr. inaequivalvis (Walaszczyk and Wood in Niebuhr et al. 1999: p. 188). The same stratigraphic level is indicated in the marl facies of Saxony where the youngest I. stuemckei occur in the Strehlen Limestone of the Strehlen Formation (upper Mytiloides striatoconcentricus/M. labiatoidiformis Zone; Inoceramid Assemblage Zone 19 of Tröger 1998), immediately followed by the M. scupini Zone (Inoceramid Assemblage Zone 19a of Tröger 1998).
In the Lüneburg section, the range of Inoceramus annulatus Goldfuss, 1836 according to Heinz [(1928a), i.e., Goldfuss’ specimen 7a (1836: pl. 110, fig. 7a)], is identical with the range of Mytiloides scupini (= Inoceramus stillei of Heinz 1928a: pl. 3; Inoceramus stillei var. scupini n. var. of Heinz 1930). This interval is in turn characterized by small- to medium-sized representatives of the lamarcki group, e.g., Inoceramus lusatiae, in central European sections (Walaszczyk and Wood 1998). Very large and huge inoceramids > 250 mm in height are missing in this interval. This suggests that Heinz (1926, 1928a) by no means saw in the small Goldfuss specimen 7a (< 85 mm in height) a representative of a huge inoceramid species with “shells exceeded three feet [914 mm] in length and twenty inches [508 mm] in width” (Mantell 1822: p. 213; see Fig. 13), often noted from the “schloenbachi beds” (Lower Coniacian) and the Lower–Middle Coniacian transitional beds (Walaszczyk 1992), e.g., Inoceramus annulatus in the sense of Walaszczyk and Wood (1998).

Inoceramus annulatus sensu Walaszczyk and Wood (1998). BGR: Ma 14107; double-valved specimen in butterfly position with most of the shell preserved, more than 450 mm in height, with nicely preserved “Anwachsringreifen” on the composite mould, but not on the outer surface of the shell; Erwitte Formation, (2) 443.50–447 m depth in the Staffhorst shaft, Lower Saxony; Cremnoceramus crassus crassus Zone, Lower Coniacian
Likewise, the large suite of ca. 200 inoceramid bivalves of the Andert collection (housed in the SNSD) from the Sonnenberg quarry of the Zittau Sandstone Mountains (Turonian/Coniacian boundary interval, upper M. scupini to lower C. deformis erectus zones, Andert 1911; Niebuhr et al. 2020), lacks any Inoceramus specimens > 150 mm. There, at its type locality, Mytiloides scupini disappears probably in the C. waltersdorfensis waltersdorfensis Zone (Niebuhr et al. 2020: fig. 10). From exactly the same ca. 20 m thick interval Heinz (1930: p. 26) identified Inoceramus annulatus Goldfuss, 1836 in the large suite of ca. 100 Inoceramus lusatiae [deformed specimen MMG SaK 13304, ca. 80 mm in height; original of Inoceramus lusatiae of Andert (1911: pl. 8, fig. 5)]. However, this identification was not confirmed by Andert (1934: p. 128), Tröger (1967: p. 73) and Walaszczyk (1996: p. 369). Moreover, Walaszczyk and Wood (1998) are of the opinion that Inoceramus lusatiae and I. annulatus, both belonging to the lamarcki group, are extremely difficult to distinguish and do not include the original of Andert (1911: pl. 8, fig. 5) in their taxonomic concept of the small- to medium-sized species I. lusatiae. However, I. stuemckei, I. lusatiae, the Goldfuss specimen 7a (1836: pl. 110, fig. 7a), and the Early Coniacian I. annulatus exhibit very similar lamarcki-like ornaments with nearly concentric rugae, covered by regularly spaced, raised growth lines (Anwachsringreifen, cf. Heinz 1928b: p. 19, 23; 1928c: p. 75). Furthermore, the fact that I. annulatus of Heinz (1926, 1928a) and I. lusatiae have an overlap in the topmost Turonian Mytiloides scupini Zone suggests that Goldfuss’ specimen 7a (1836: pl. 110, fig. 7a) may rather correspond to Andert’s species concept of I. lusatiae. This, in turn, would indicate that I. annulatus of Heinz (1926, 1928a) is a younger synonym of Inoceramus lusatiae Andert, 1911 and the name Inoceramus annulatus Goldfuss, 1836 is a nomen nudum which does not fulfil the criteria of the ICZN (Ride et al. 1999).
Inoceramus annulatus in the sense of Walaszczyk and Wood (1998) first appears some way-up in the Lower Coniacian (Fig. 12), maybe as a phylogenetic successor of Inoceramus lusatiae, its most closely resembling species (Walaszczyk and Wood 1998). I. lusatiae disappears nearly simultaneously with the first appearance of I. annulatus, however, at different stratigraphic levels at certain central European sections, e.g., the C. c. crassus Zone at Staffhorst (Walaszczyk and Wood 2018; site 2 in Figs. 2, 12; see also Fig. 13) and the base of the C. w. hannovrensis Zone of Słupia Nadbrzeżna, Poland (Walaszczyk and Wood 1998). Furthermore, a transitional form of I. lusatiae/annulatus was found at the latter section (Walaszczyk and Wood 1998: pl. 10, fig. 5) which equals the small Goldfuss specimen (1836: pl. 110, fig. 7a) very well. Also, in the Salzgitter-Salder section, I. lusatiae has its highest occurrence at the base of the C. w. hannovrensis Zone (163 m depth according to Wood and Ernst 1998: figs. 55, 56) while I. annulatus first appears 12 m above, respectively (175 m depth; Walaszczyk et al. 2021). This strongly suggests that I. annulatus should be regarded as a successor of I. lusatiae, and that I. lusatiae from the C. w. hannovrensis and C. c. inconstans zones at Staffhorst equals to small- to medium-sized specimens of I. annulatus of Salzgitter-Salder and Słupia Nadbrzeżna, Poland. I. annulatus ranges to the top of the Lower Coniacian, possibly also into the basal Middle Coniacian (Walaszczyk and Wood 1998: p. 424).
In a nutshell, I. annulatus Goldfuss, 1836 sensu Heinz (1926, 1928a) and I. annulatus sensu Walaszczyk and Wood (1998) are almost certainly different species with different stratigraphic ranges—so far with an identical lectotype of latest Turonian age (see Fig. 1). The current paper does not provide a final solution for the taxonomic issues arising from this conclusion but we suggest that the name I. annulatus sensu Walaszczyk and Wood (1998) should be retained for the very large to huge Early Coniacian forms in order to reflect the taxonomic usage of the last decades. Medium-sized I. annulatus sensu Heinz (1926, 1928a) from the latest Turonian may be conspecific to contemporaneous I. lusatiae Andert, 1911 but further systematic studies are needed in this respect in order to satisfactorily solve the complex nomenclatorial dilemma revolving around I. annulatus according to the ICZN (Ride et al. 1999).
Palaeoecological implications
When evaluating the morphological trends in the above considered late Middle Turonian to Early Coniacian representatives of the lamarcki group, an overall maximum size increase becomes evident, accompanied by a thickening of the shell and buttressing the hinge areas (Fig. 14). The relatively abrupt increase in shell size from medium-sized Inoceramus lamarcki to very large Inoceramus stuemckei at the transition from the Middle to the Late Turonian may have been fostered by a global eustatic sea-level fall at this time (sequence boundary SB Tu 3 of Janetschke et al. 2015), resulting in widespread shallowing of the shelf seas and triggering speciation in high-stress situations (cf. Kauffman 1978a). This speciation event seems to follow the evolutionary mode of punctuated equilibria sensu Eldridge and Gould (1972) and Gould and Eldredge (1977) and have been documented for other Upper Cretaceous inoceramids (e.g., Kauffman 1977). Corresponding evolutionary events often occurred during high-stress situations affecting whole communities during short time intervals such as rapid marine regression (Kauffman 1978a) when habitat segregation favored allopatric speciation. In the henceforth shoaling early Late Turonian seas, the newly evolved large-sized and thickened shells were probably beneficial in two ways: on the one hand, a thick and strongly ribbed shell may provide increased mechanical protection and sea-floor adhesion in agitated shallow waters. However, this interpretation alone is certainly not the main reason because the same inoceramid morphs also occur in contemporaneous deeper water facies, i.e., under low-energy conditions. On the other hand, size increase is, together with sculptural features such as ornamentation and thick shells, a common passive defense strategy among shelled organisms in order to reach a size refugium in which the prey becomes unassailable for a predator (e.g., Klompmaker et al. 2019). This is especially important for immobile epifaunal organisms such as inoceramid bivalves that simply cannot escape from attacks. Most likely the development of the massive hinges in I. stuemckei also aimed at improved breakup-protection against predator attempts to open the valves, converting the live shells into a “security container” (Coen 1985 pointed to the important role of the bivalve hinge in withstanding shear forces). Thus, large, thick and strongly ribbed shells with buttressed hinges in the first place provided improved shelter against shell-breaking marine predators such as ptychodontoid sharks and marine reptiles (e.g., mosasaurid lizards) that are much more common in shallow waters (cf. Ward 1996; Kelley and Hansen 2001) and increased in importance during the Mesozoic marine revolution, especially in the (Late) Cretaceous (Vermeij 1977). We thus propose that predation pressure increased during the Late Turonian shallowing phase, triggering the size increase and the development of high-strength shells in some representatives of the lamarcki group (Fig. 14), providing additional evidence for the escalation hypothesis of Vermeij (1987, 1994). In support of this view, Ekrt et al. (2001) and Wiese et al. (2004) provided firm evidence for the abundance of marine reptiles and sharks (i.e., potential predators) from the lower Teplice Formation (mid-Upper Turonian) of the Bohemian Cretaceous Basin; also, the contemporaneous Strehlen Limestone of Saxony yielded numerous corresponding vertebrate remains (Licht et al. 2016; Sachs et al. 2016, 2017), especially massive shark vertebrae and teeth, including specimens that can be attributed to the durophagous taxon Ptychodus (Fig. 15a). Moreover, a relatively abrupt increase in predation intensity on inoceramids by vertebrates was proposed for the Late Turonian of the US Western Interior Seaway by Harries and Ozanne (1998); they quoted an increasing abundance of Ptychodus (shark) teeth and inferred predation marks on up to 40% of the inoceramids in certain levels of the Upper Turonian. Ptychodontid sharks attained considerable sizes during the Late Cretaceous (Jambura and Kriwet 2020) and are known to have preyed upon inoceramids (Kauffman 1978b). Furthermore, durophagous adaptions such as molariform teeth on the fingers of the chelae, heterochely (e.g., enlarged crusher claws and/or specialized cutting claws) and calcified mandibles had appeared among multiple clades of the Decapoda by the Late Cretaceous (Schweitzer and Feldmann 2010). Especially nephropid and erymid lobsters were widely distributed during the Cretaceous and had claws that could be used for crushing or capturing prey (Feldmann and Schweitzer 2014); large erymids are not uncommon in the Saxo-Bohemian Cretaceous (see Geinitz 1875; Fritsch and Kafka 1887; Gründel 1974), e.g., Enoploclytia leachii (Mantell) which has also been found in the Strehlen Limestone (Fig. 15c), along with abundant shark remains including ptychodontoid teeth and vertebrae (Fig. 15a, b). Harries and Ozanne (1998: p. 382) also brought two further decapod groups into play who attained durophagy during the Late Cretaceous, corresponding to the interval where evidence for inoceramid predation increases: stomatopod and brachyuran crustaceans. Experiments on crushing crab predation showed that the shells of the Blue Mussel Mytilus edulis, a well-suited model organism for the typical shell bauplan of many Cenomanian–Middle Turonian inoceramids, works well with its thin shell when staying small, but an increase in size is structurally disadvantageous (Nagel-Myers et al. 2019). This observation may be the reason that the size increase of Late Turonian inoceramids is not seen in myti-liform taxa and only certain groups started the arms race with the diversifying durophagous predators, also applying shell-thickening and sculpture formation. Consequently, the numerous isolated massive hinges and thick shell fragments found in Upper Turonian strata, originally giving rise for the erection of I. hercules, might also be seen as evidence of (successful) predation. However, such fragments may be indistinguishable from shell material resulting from post-mortem shell breakdown by physical processes (Schweitzer and Feldmann 2010). After a gap in the fossil record during the late Late Turonian and earliest Coniacian when large- to huge-sized inoceramids largely disappeared (interval with I. lusatiae in Figs. 12, 14), I. annulatus readopted the trend of maximum size increase in the lamarcki group during the Early Coniacian (Fig. 14). Even if I. annulatus may be seen as an phylogenetic successor of I. lusatiae (to which it has the closest morphological relationship; cf. Walaszczyk and Wood 1998), the exact evolutionary relationships in this interval remain elusive; further detailed stratigraphic and systematic studies are needed.

Stratigraphic ranges, details on regional occurrences and possible evolutionary patterns of selected inoceramid species of the lamarcki group supplemented with thumbnail illustrations of inferred durophagous predators (see text for further explanation). Inoceramid illustrations based on Tröger (1967, I. lamarcki, I. stuemckei), Tröger and Niebuhr (2014, I. lusatiae) and Walaszczyk and Cobban (2000, I. annulatus), predator thumbnail images redrawn/modified after https://commons.wikimedia.org/wiki/File:Mosasaurus_beaugei1DB.jpg (mosasaur, Creative Commons Attribution 3.0 Unported license), Fritsch and Kafka (1887: E. leachi, public domain) and http://www.deviantart.com/art/Ptychodus-mortoni-533593712 (Ptychodus, Creative Commons Attribution-Share Alike 4.0 International license). A1 = lower I. stuemckei acme, A2 = upper I. stuemckei acme; note that both acmes follow global sea-level falls

Fossil remains of potential inoceramid predators from the mid-Upper Turonian Strehlen Limestone of the lower Strehlen Formation (Elbtal Group, Saxony). a MMG: SaK 10330a; cracker tooth of the durophagous shark Ptychodus mammillaris (Mantell). b MMG: SaK 10537; large vertebra assigned to Ptychodus (cf. Schlesiger 2021). c MMG: SaK 10196; right claw of the durophagous crustacean Enoploclytia leachii (Mantell)
Mode of life
Inoceramid bivalves were epibenthic, predominantly sessile organisms that often dominated Late Cretaceous offshore level-bottom communities (e.g., Kauffman et al. 2007; Wilmsen 2008; Wilmsen et al. 2021), but they are also found in nearshore sands, suggesting that they had comparatively wide tolerances for physico-chemical environmental factors at genus and species level (e.g., Harries et al. 1996). From the perspective of functional morphology, the flat, disc-like shells of I. stuemckei and I. annulatus provide large resting surfaces that are, in conjunction with the thick shell, most likely an adaption to a recumbent lifestyle (see Kauffman et al. 2007 for Platyceramus and Wilmsen et al. 2021 for disc-like Gnesioceramus crippsi). An upright, semi-infaunal mode of life with the dorsal part buried and the ventral part directed into the water column is less likely given the fact that in the sandy nearshore facies of Saxony I. stuemckei has always been found in horizontal position, whereas contemporaneous pinnid bivalves often occurred in their vertical semi-infaunal life position (e.g., Wilmsen and Niebuhr 2014b: figs. 13c, 13d).
Where preserved, the regular growth rings of both I. stuemckei and I. annulatus show a rather equal width between 2–5 mm across the major part of the shell (or external imprint) of individual specimens (see for example Figs. 6 and 11b). The equally broad increments suggest constantly recurring growth cycles with no or little ontogenetic variation. Environmental variables in fact significantly influence modern bivalve shell formation with temperature and food availability exerting the main controls on growth rate (Schöne et al. 2005; Walliser et al. 2018). Regular growth rings of similar dimension (2 mm) in Late Cretaceous inoceramids from Demerara Rise have been assigned to short-term cycles of food availability in deep-oceanic bottom waters, driven by seasonal/annual productivity changes in surface waters (Jiménez Berrocoso et al. 2008). Furthermore, Walliser et al. (2020) suggest that growth bands in Late Turonian Inoceramus hercules (= I. stuemckei) were produced annually and shell growth was nearly continuous throughout the year. Accepting this, a half- to one-meter-high inoceramid bivalve would have attained a maximum age of 250–500 years. However, even if some modern bivalves may reach ages of > 200 year under more temperate conditions and with slower growth rates (e.g., Arctica islandica; Thompson et al. 1980), such an old age (and consequently slow growth) for inoceramids seems very unlikely, especially when considering that ontogenetic size increase as fast as possible was of utmost importance in the predator-littered Late Cretaceous marine environment in order to reach the size refugium, following the leitmotif: grow fast or die young. Furthermore, intra-annual growth increments have been observed by Schöne et al. (2005) and we speculate that the incremental growth is reflecting in fact sub-annual growth cycles; monthly neap–spring tidal cycles, known to potentially be recorded in bivalve shells (e.g., Pannella and MacClintock 1968; Richardson et al. 1980), may be an option. Their effects are most pronounced in inter- to shallow subtidal environments but spring–neap modulation of coastal water circulation may result in subtidal flows that are known to be capable of rapidly flushing buoyant material out of the coastal zone and into the sea at neap tides (Ribeiro et al. 2004), exerting a strong control on food availability for and, thus, incremental shell growth of shelf biota. In this scenario, the possible life span of the large inoceramids would be reduced to 20–40 years, but a final statement on the origin of the inoceramid growth rings is currently not possible.
Conclusions
Based on the systematic and stratigraphic revision of selected large inoceramids of the lamarcki group from Upper Turonian and Lower Coniacian strata of Germany, the following taxonomic, biostratigraphic, evolutionary and palaeoecological conclusions can be drawn:
-
In his Petrefacta Germaniae, Goldfuss (1836) illustrated two inoceramid specimens as Inoceramus annulatus which were differentiated by Heinz (1926, 1928a, c) into “Inoceramus latus Mant., var. stümckei n. var.” (i.e., the specimen on pl. 110, fig. 7b in Goldfuss 1836) and “Inoceramus annulatus Goldf., part. Heinz” (i.e., the specimen on pl. 110, fig. 7a in Goldfuss 1836). Both taxa became entrenched into the literature later on, as Late Turonian I. lamarcki stuemckei Heinz, 1928a and (Early) Coniacian I. annulatus Goldfuss, 1836. However, both specimens of Goldfuss (1836) are apparently of Late Turonian age.
-
Inoceramus stuemckei Heinz, 1928a, characterized by a box-shaped to pentagonal outline and irregularly spaced, distant concentric rugae, is well represented by its lectotype (Goldfuss 1836, pl. 110, fig. 7b). This large to very large species is very common and characterizes the lower to middle Upper Turonian, including two conspicuous acme occurrences.
-
Inoceramus stuemckei Heinz, 1928a has a facies-independent occurrence from sandy shallow-water into fine-grained marly-calcareous offshore facies. After its possible first appearance in the late Middle Turonian, it ranges from the lower Upper Turonian I. perplexus/I. stuemckei/I. inaequivalvis assemblage zone to the top of the mid-Upper Turonian Mytiloides striatoconcentricus/M. labiatoidiformis concurrent range zone. As thin-shelled I. perplexus, the widely accepted index inoceramid for the base of the Upper Turonian, are largely restricted to the offshore facies, the first acme occurrence of I. stuemckei is an excellent inoceramid marker for the base of the substage in shallow-water facies.
-
Large and massive calcitic hinges, found isolated in contemporaneous Upper Turonian strata and newly introduced by Heinz (1932a) as Heroceramus hercules, are merely parts of the time-equivalent large I. stuemckei Heinz, 1928a as emended herein; the name is thus no longer valid.
-
Inoceramus annulatus Goldfuss, 1836 remains taxonomically problematic. Despite being based on an only medium-sized lectotype from the latest Turonian, the name was later applied to very large to huge forms characterized by a triangular-rounded outline, more widely and regularly spaced concentric rugae, an upright form and a much larger maximum size, first appearing in the mid-Early Coniacian. Thus, I. annulatus in the sense of Walaszczyk and Wood (1998) is poorly represented by its lectotype that we regard being very close the latest Turonian–earliest Coniacian species I. lusatiae Andert, 1911, a medium-sized representative of the lamarcki group. The complex nomenclatorial issues revolving around I. annulatus cannot be solved herein and further systematic studies are needed.
-
I. annulatus sensu Walaszczyk and Wood (1998) is comparably rare and appears in the mid-Lower Coniacian Cremnoceramus waltersdorfensis hannovrensis Zone, ranging into the basal Middle Coniacian.
-
Valves of both I. stuemckei Heinz, 1928a and I. annulatus sensu Walaszczyk and Wood (1998) are ornamented with regularly spaced, round-topped rugae, covered by fine, regularly spaced and slightly raised growth lines (“Anwachsringreifen” of Heinz 1928b). The presence/absence of this feature in imprints and moulds is related to different preservation forms and cannot be used as a taxonomic criterion.
-
Inoceramus stuemckei Heinz, 1928a and I. annulatus sensu Walaszczyk and Wood (1998) are characterized by a stratigraphically abrupt enlargement of their shells and buttressing of the hinges across the Middle/Upper Turonian boundary interval and within the Early Coniacian, respectively. Up to 500 mm were reached in Late Turonian I. stuemckei, and Early Coniacian I. annulatus attain shell heights up to one meter.
-
The evolutionarily fairly abrupt shift to large/very large inoceramid shells in the Late Turonian is accompanied by widespread evidence for increased predation by durophagous organism such as sharks, mosasaurs and crustaceans. Size increase is a common passive defense strategy among (shelled) organisms in order to reach a size refugium in which the prey becomes unassailable for a predator. We thus speculate that the considerable size increase in Late Turonian to Early Coniacian inoceramid bivalves is part of an escalating arms race between prey and predators, ultimately triggering Late Cretaceous inoceramid gigantism.
-
We infer a recumbent lifestyle for the very large to huge, flat and disc-like I. stuemckei Heinz, 1928a and I. annulatus sensu Walaszczyk and Wood (1998) and rapid ontogenetic shell growth, reflected by the regularly spaced growth lines that most likely document sub-annual growth cycles.
References
Andert, H. 1911. Die Inoceramen des Kreibitz-Zittauer Sandsteingebirges. Festschrift des Humboldtvereins zur Feier seines 50-jährigen Bestehens: 1–63.
Andert, H. 1934. Die Kreideablagerungen zwischen Elbe und Jeschken. Teil III: Die Fauna der obersten Kreide zwischen Sachsen, Böhmen und Schlesien. Abhandlungen der Preußischen Geologischen Landesanstalt, Neue Fassung 159: 1–477.
Bodylevsky, V.I. 1958. Upper Cretaceous fauna from the lower reaches of the river Yenisei. In Jurassic and Cretaceous faunas of the lower course of Yenisei, eds. V.I. Bodylevsky and N.I. Shulgina. Transactions of the Arctic Institute, Geology 93: 69–86.
Bräutigam, F. 1962. Zur Stratigraphie und Paläontologie des Cenomans und Turons im nordwestlichen Harzvorland. PhD thesis Technische Hochschule Carola-Wilhelmina zu Braunschweig: 1–261 [unpubl.].
Carter, J.G., C.R. Altaba, L.C. Anderson, R. Araujo, A.S. Biakov, A.E. Bogan, D.C. Campbell, M. Campbell, J.C.W. Chen Jin-hua, G. Cope, H.H. Dijkstra. Delvene, R.N. Fang Zong-jie, V.A. Gardner, I. Gavrilova, P.J. Goncharova, J.H. Harries, M. Hartman, W.R. Hautmann, J. Hylleberg. Hoeh, P. Jiang Bao-yu, L. Johnston, K. Kirkendale, J. Kleemann, J. Koppka, D. Kříž, N. Machado, A. Malchus, J.-P. Márquez-Aliaga, C.A. Masse, P.U. McRoberts, S. Middelfart, L.A. Mitchell, S. Nevesskaja, J. Özer Jr., I.V. Pojeta, J.M. Polubotko, S. Pons, T. Popov, A.F. Sánchez, R.W. Sartori, I.I. Scott, J.H. Sey, V.V. Signorelli, P.W. Silantiev, T. Skelton, J.B. Steuber, G.L. Wingard. Waterhouse, and T. Yancey. 2011. A synoptical classification of the Bivalvia (Mollusca). Paleontological Contributions 4: 1–47.
Cleevely, R.J., and N.J. Morris. 1987. Bivalven. In Kreide-Fossilien: Bestimmungsatlas der Fossilien des Chalk, eds. E. Owen and A.B. Smith. 49–74.
Cleevely, R.J., and N.J. Morris. 2002. Introduction to molluscs and bivalves. In Fossils of the Chalk, eds. A.B. Smith and D.J. Batten. The Palaeontological Association, Field Guide to Fossils 2: 99–160.
Coen, L.D. 1985. Shear resistance in two bivalve molluscs: Role of hinges and interdigitating margins. Journal of Zoology 205: 479–487.
Dacqué, E. 1939. Die Fauna der Regensburg-Kelheimer Oberkreide (mit Ausschluß der Spongien und Bryozoen). Abhandlungen der Bayerischen Akademie der Wissenschaften, Neue Fassung 45: 1–218.
Deecke, W. 1908. Eine Tiefbohrung in Heringsdorf (Usedom). Mittheilungen aus dem naturwissenschaftlichen Vereine von Neu-Vorpommern und Rügen 40: 56–60.
Delvaque, C., F. Robaszynski, and B. Matrion. 2014. Les autres faunes. In Le Turonien des Massifs d’Uchaux et de la Cèze (S.E. France)—Migration globale d’ammonites et conséquences sur la zonation internationale, rudistes et corrélations entre les massifs, eds. F. Robaszynski, F. Amédro, C. Delvaque and B. Matrion. Mémoires de la Classe des Sciences 4/4: 175–177.
Ekrt, B., M. Košťák, M. Mazuch, J. Valíček, S. Voigt, and F. Wiese. 2001. Short note on new records of late Turonian (Upper Cretaceous) marine reptiles remains from the Úpohlavy quarry (NW Bohemia, Czech Republic). Bulletin of Geosciences 76: 101–106.
Eldridge, N., and S.J. Gould. 1972. Punctuated equilibria: an alternative to phyletic gradualism. In Models in Paleobiology, ed. T.J.M Schopf. 82–115.
Ernst, G., F. Schmid, and E. Seibertz. 1983. Event-Stratigraphie im Cenoman und Turon von NW-Deutschland. Zitteliana 10: 531–554.
Feldmann, R.M., and C.E. Schweitzer. 2014. Lobster (Decapoda) diversity and evolutionary patterns through time. Journal of Crustacean Biology 34: 820–847.
Fritsch, A., and J. Kafka. 1887. Die Crustaceen der böhmischen Kreideformation. 54 pp., 10 pls.; Praha (Františak Rivnáce).
Geinitz, H.B. 1839. Charakteristik der Schichten und Petrefacten des sächsischen Kreidegebirges, Erstes Heft. Der Tunnel von Oberau in geognostischer Hinsicht, und die dieser Bildung verwandten Ablagerungen zwischen Oberau, Meissen und dem Plauen‘schen Grunde bei Dresden. 1–30.
Geinitz, H.B. 1849. Das Quadersandsteingebirge oder Kreidegebirge in Deutschland. 1–292.
Geinitz, H.B. 1873. Brachiopoden und Pelecypoden. Das Elbthalgebirge in Sachsen. Zweiter Theil. Der mittlere und obere Quader. Palaeontographica 20 (II): II.21–II.52.
Geinitz, H.B. 1875. Crustacea, Krebse. Das Elbthalgebirge in Sachsen. Zweiter Theil. Der mittlere und obere Quader. Palaeontographica 20 (II): II.202–II.205.
Giebel, C.G. 1852. Deutschlands Petrefakten. Ein systematisches Verzeichnis aller in Deutschland und den angrenzenden Gebieten vorkommenden Petrefakten nebst Angabe der Synonyme und Fundorte. 329–441.
Goldfuss, A. 1834–1840. Petrefacta Germaniae 2 (1): 1–68, pls 72–96 [1834], (2): 69–140, pls 97–121 [1836], (3): 141–224, pls 122–146 [1837], (4): I–III + 225–312, pls 147–165 [1840].
Gould, S.J., and N. Eldredge. 1977. Punctuated Equilibria: The tempo and mode of evolution reconsidered. Paleobiology 3: 115–151.
Gründel, J. 1974. Die Decapoda (Crustacea) der sächsischen Kreide. Abhandlungen des Staatlichen Museums für Mineralogie und Geologie zu Dresden 21: 79–100.
Hagenow, F. von 1842a. Greifswald, 31. März 1842. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde: p. 317.
Hagenow, F. von 1842b. Monographie der Rügen´schen Kreide-Versteinerungen, III. Abtheilung: Mollusken. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde: 528–575.
Harries, P.I., and C.R. Ozanne. 1998. General trends in predation and parasitism upon inoceramids. Acta Geologica Polonica 48: 377–386.
Harries, P.J., E.G. Kauffman, and J.S. Crampton. 1996. Lower Turonian Euramerica Inoceramidae; a morphologic, taxonomic, and biostratigraphic overview: a report from the First workshop on Early Turonian inoceramids (Oct. 5–8, 1992) in Hamburg, Germany. Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 77: 641–671.
Heinz, R. 1926. Beitrag zur Kenntnis der Stratigraphie und Tektonik der oberen Kreide Lüneburgs. PhD thesis, Mathematisch-Naturwissenschaftliche Fakultät der Hamburgischen Universität. 1–109.
Heinz, R. 1928a. Das Inoceramen-Profil der Oberen Kreide Lüneburgs. Mit Anführung der neuen Formen und deren Kennzeichnung. Beiträge zur Kenntnis der oberkretazischen Inoceramen I. Jahresbericht des Niedersächsischen geologischen Vereins 21: 64–81.
Heinz, R. 1928b. Über die bisher wenig beachtete Skulptur der Inoceramen-Schale und ihrer stratigraphischen Bedeutung. Beiträge zur Kenntnis der Inoceramen IV. Mitteilungen aus dem Mineralogisch-Geologischen Staatsinstitut in Hamburg 10: 3–39.
Heinz, R. 1928c. Über die Oberkreide-Inoceramen Süd-Amerikas und ihre Beziehungen zu denen Europas und anderer Gebiete. Beiträge zur Kenntnis der Inoceramen V. Mitteilungen aus dem Mineralogisch-Geologischen Staatsinstitut in Hamburg 10: 41–97.
Heinz, R. 1930. Zur stratigraphischen Stellung der Sonnenbergschichten bei Waltersdorf i. Sa. (westsüdwestlich von Zittau). Beiträge zur Kenntnis der Inoceramen IX. Jahresbericht des Niedersächsischen geologischen Vereins 23: 25–30.
Heinz, R. 1932a. Zur Gliederung der sächsisch-schlesisch-böhmischen Kreide unter Zugrundelegung der norddeutschen Stratigraphie. Beiträge zur Kenntnis der Inoceramen X. Jahresbericht des Niedersächsischen geologischen Vereins 24: 22–53.
Heinz, R. 1932b. Aus der neuen Systematik der Inoceramen. Beiträge zur Kenntnis der Inoceramen XIV. Mitteilungen aus dem Mineralogisch-Geologischen Staatsinstitut in Hamburg 13: 1–26.
Hiss, M., B. Niebuhr, and U. Teipel. 2018. Die Kreide in der Stratigraphischen Tabelle von Deutschland 2016 – The Cretaceous System in the Stratigraphic Table of Germany 2016. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften 169: 247–266.
Jambura, P.L., and J. Kriwet. 2020. Articulated remains of the extinct shark Ptychodus (Elasmobranchii, Ptychodontidae) from the Upper Cretaceous of Spain provide insights into gigantism, growth rate and life history of ptychodontid sharks. PLoS ONE 15: e0231544.
Janetschke, N., B. Niebuhr, and M. Wilmsen. 2015. Inter-regional sequence stratigraphical synthesis of the Plänerkalk, Elbtal and Danubian Cretaceous groups (Germany): Cenomanian-Turonian correlations around the Mid-European Island. Cretaceous Research 56: 530–549.
Jiménez Berrocoso, Á., K.G. MacLeod, S.E. Calvert, and J. Elorza. 2008. Bottom water anoxia, inoceramid colonization, and benthopelagic coupling during black shale deposition on Demerara Rise (Late Cretaceous western tropical North Atlantic). Paleoceanography 23: PA3212 (20 pp.). https://doi.org/10.1029/2007PA001545.
Kaplan, U. 2011. Turonium und Unterconiacium (Oberkreide) im Steinbruch DIMAC bei Halle (Westfalen), Teutoburger Wald (Östliches Münsterländer Kreidebecken). Geologie und Paläontologie in Westfalen 81: 75–105.
Kauffman, E.G. 1977. Evolutionary rates and biostratigraphy. In Concepts and methods of biostratigraphy, eds. E.G. Kauffman and J.E. Hazel, 109–141.
Kauffman, E.G. 1978a. Evolutionary rates and patterns among Cretaceous Bivalvia. Philosophical Transactions of the Royal Society of London B 284: 277–304.
Kauffman, E.G. 1978b. Ptychodus predation upon a Cretaceous Inoceramus. Paleontology 15: 439–444.
Kauffman, E.G., P.J. Harries, C. Meyer, T. Villamil, C. Arango, and G. Jaecks. 2007. Paleoecology of giant Inoceramidae (Platyceramus) on a Santonian (Cretaceous) seafloor in Colorado. Journal of Paleontology 81: 64–81.
Keller, S. 1982. Die Oberkreide der Sack-Mulde bei Alfeld (Cenoman–Unter-Coniac) Lithologie, Biostratigraphie und Inoceramen. Geologisches Jahrbuch A 64: 3–171.
Kelley, P.H., and T.A. Hansen. 2001. Mesozoic marine revolution. In Palaeobiology II, eds. D.E.G. Briggs and P.R. Crowther. 94–97.
Kelly, S.R.A., and C.J. Wood. 1996. A revision of the inoceramids in Henry Woods´ Monograph of the Cretaceous Lamellibranchia of England (1899–1913). Progress Report. Abstract Volume, 5th International Cretaceous Symposium and Inoceramid workshop, Freiberg, Germany, 16–24 September 1996: 1–13.
Klompmaker, A.A., P.H. Kelley, D. Chattopadhyay, J.C. Clements, J.W. Huntley, and M. Kowalewski. 2019. Predation in the marine fossil record: Studies, data, recognition, environmental factors, and behavior. Earth-Science Reviews 194: 472–520.
Leonhard, R. 1897. Die Fauna der Kreideformation in Oberschlesien. Palaeontographica 44: 11–70.
Licht, M., I. Kogan, J. Fischer, and S. Reiss. 2016. Knochenfische (Osteichthyes). In Kreide-Fossilien in Sachsen, Teil 2, eds. B. Niebuhr and M. Wilmsen. Geologica Saxonica 62: 143–168.
Mantell, G. 1822. The fossils of the South Downs; or illustrations of the geology of Sussex. 1–320. https://www.biodiversitylibrary.org/item/97604#page/11/mode/1up
Nagel-Myers, J., I. Mastorakos, P. Yuya, and G. Reeder. 2019. Modelling crushing crab predation on bivalve prey using finite element analysis. Historical Biology. https://doi.org/10.1080/08912963.2019.1699555.
Niebuhr, B. 2006. Multistratigraphische Gliederung der norddeutschen Schreibkreide (Coniac bis Maastricht), Korrelation von Aufschlüssen und Bohrungen. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften 157: 245–262.
Niebuhr, B., R. Baldschuhn, G. Ernst, I. Walaszczyk, W. Weiss, and C.J. Wood. 1999. The Upper Cretaceous succession (Cenomanian–Santonian) of the Staffhorst Shaft, Lower Saxony, northern Germany: Integrated biostratigraphic, lithostratigraphic and downhole geophysical log data. Acta Geologica Polonica 49: 175–213.
Niebuhr, B., M. Hiss, U. Kaplan, K.-A. Tröger, S. Voigt, T. Voigt, F. Wiese, and M. Wilmsen. 2007. Lithostratigraphie der norddeutschen Oberkreide. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 55: 1–136.
Niebuhr, B., M. Wilmsen, and T. Voigt. 2020. Die Oberkreide (Cenomanium–Mittelconiacium) im Zittauer Sandsteingebirge (Deutschland, Tschechien). Zeitschrift der Deutschen Gesellschaft für Geowissenschaften 171: 163–197.
Pannella, G., and C. MacClintock. 1968. Biological and environmental rhythms reflected in molluscan shell growth. The Paleontological Society Memoir 2: 64–80.
Ribeiro, C.H.A., J.J. Waniek, and J. Sharples. 2004. Observations of the spring–neap modulation of the gravitational circulation in a partially mixed estuary. Ocean Dynamics 54: 299–306.
Richardson, C.A., D.J. Crisp, and N.W. Runham. 1980. Factors influencing shell growth in Cerastoderma edule. Proceedings of the Royal Society (b) 210: 513–531.
Ride, W.D.L., H.G. Cogger, C. Dupuis, O. Fraus, A. Minelli, F.C. Thompson, and P.K. Tubbs 1999 (Eds.). International Code of Zoological Nomenclation (pp. 1–306). London: The Natural History Museum.
Roemer, F. 1870. Geologie von Oberschlesien. Atlas von 50 Tafeln mit Abbildungen von Versteinerungen.
Sachs, S., J.J. Hornung, J. Knüppe, M. Wilmsen, and B.P. Kear. 2016. Reptilien. In Kreide-Fossilien in Sachsen, Teil 2, eds. B. Niebuhr and M. Wilmsen. Geologica Saxonica 62: 169–180.
Sachs, S., M. Wilmsen, J. Knüppe, J.J. Hornung, and B.P. Kear. 2017. Cenomanian-Turonian marine amniote remains from the Saxonian Cretaceous Basin of Germany. Geological Magazine 154: 237–246.
Schlesiger, E. 2021. Elasmobranchier-Wirbel aus der sächsischen Kreide: Verbreitung, Systematik und Paläobiologie. BSc thesis, Fakultät für Geowissenschaften, Geotechnik und Bergbau der TU Bergakademie Freiberg. 65 pp.
Schlüter, C. 1877. Kreide-Bivalven. Zur Gattung Inoceramus. Palaeontographica 24: 249–288.
Schöne, B.R., S.D. Houk, A.D. Freyre Castro, J. Fiebig, and W. Oschmann. 2005. Daily growth rates in shells of Arctica islandica: Assessing subseasonal environmental controls on a long-lived bivalve mollusk. Palaios 20: 78–92.
Schweitzer, C.E., and R.M. Feldmann. 2010. The Decapoda (Crustacea) as predators on Mollusca through geologic time. Palaios 25: 167–182.
Seitz, O. 1921. Die stratigraphisch wichtigen Inoceramen des norddeutschen Turons. Zeitschrift der Deutschen Geologischen Gesellschaft, Monatsberichte 73: 99–107.
Sowerby, J. 1812–1822. The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the Earth. 1–803; 1, pls 1–9 [1812], pls 10–44 [1813], pls 45–78 [1814], pls 79–102 [1815]; 2, pls 103–114 [1815], pls 115–150 [1816], pls 151–186 [1817], pls 187–203 [1818]; 3, pls 204–221 [1818], pls 222–253 [1819], pls 254–271 [1820], pls 272–306 [1821]; 4, pls 307–318 [1821], pls 319–383 [1822].
Sowerby, J. de C. 1823–1845. The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the Earth. 1–558; 5, pls 384–443 [1823], pls 444–485, [1824], pls 486–503 [1824]; 6, pls 504–544 [1826], pls 545–580 [1827], pls 581–597 [1828], pls 598–609 [1829]; 7, pls 610–618 [1840], pls 619–623 [1841], pls 624–628 [1843], pls 629–643 [1844], pls 644–648 [1845].
Tarkowski, R. 1991. Stratigraphy, macrofossils and palaeogeography of the Upper Cretaceous from the Opole Trough. Zeszytów Naukowych Akademii Górniczo - Hutniczej, Geologia 51: 1–156.
Tarkowski, R. 1996. Inoceramid biostratigraphy in the Turonian of the Opole Trough (southwestern Poland). Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 77: 489–501.
Thompson, I., S.D. Jones, and D. Dreibelbis. 1980. Annual internal growth banding and life history of the ocean quahog Arctica islandica (Mollusca: Bivalvia). Marine Biology 57: 25–34.
Tröger, K.-A. 1967. Zur Paläontologie, Biostratigraphie und faziellen Ausbildung der unteren Oberkreide (Cenoman bis Turon). Teil 1: Paläontologie und Biostratigraphie der Inoceramen des Ceno-mans und Turons Mitteleuropas. Abhandlungen des Staatlichen Museums für Mineralogie und Geologie zu Dresden 12: 13–207.
Tröger, K.-A. 1981. Zu Problemen der Biostratigraphie der Inoceramen und der Untergliederung des Cenomans und Turons in Mittel- und Osteuropa. Newsletters on Stratigraphy 9: 139–156.
Tröger, K.-A. 1984. Über zwei außergewöhnlich große Inoceramen-Arten aus dem Ober-Turon Europas. Freiberger Forschungshefte C 395: 47–53.
Tröger, K.-A. 1987. Der Strehlener Kalkstein – ein Beitrag zur Paläontologie und Biostratigraphie des höheren Oberturons. Zeitschrift der Geologischen Wissenschaften 15: 205–212.
Tröger, K.-A. 1998. Remarks concerning morphometric parameters, biostratigraphy and palaeobiogeography of Turonian inoceramids (Bivalvia) in Europe. Zentralblatt Geologie und Paläontologie 1: 1489–1499.
Tröger, K.-A. 2009. Katalog oberkretazischer Inoceramen. Geologica Saxonica 55: 1–188.
Tröger, K.-A., and W.K. Christensen. 1991. Upper Cretaceous (Cenomanian–Santonian) inoceramid bivalve faunas from the island of Bornholm, Denmark. Danmarks Geologiske Undersøgelse A 28: 4–47.
Tröger, K.-A., and B. Niebuhr. 2014. Inoceramide Muscheln. In Kreide-Fossilien in Sachsen, Teil 1, eds. B. Niebuhr and M. Wilmsen. Geologica Saxonica 60: 169–199.
Tröger, K.-A., and L. Wolf. 1960. Zur Stratigraphie und Petrographie der Strehlener Schichten. Geologie 9: 288–298.
Tröger, K.-A., B. Niebuhr, and M. Wilmsen. 2009. Inoceramen aus dem Cenomanium bis Coniacium der Danubischen Kreide-Gruppe (Bayern, Süd-Deutschland). In Litho- und Biostratigraphie der außeralpinen Kreide von Bayern, ed. B. Niebuhr. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 65: 59–110.
Vejbæk, O.V., C. Andersen, M. Dusa, W. Herngreen, H. Krabbe, K. Leszczynski, G.K. Lott, J. Mutterlose, and A.S. van der Molen. 2010. Cretaceous. In Petroleum Geological Atlas of the Southern Permian Basin Area, eds. H. Doornenbal and A. Stevenson. 195–209.
Vermeij, G.J. 1977. The Mesozoic marine revolution: Evidence from snails, predators and grazers. Paleobiology 3: 245–258.
Vermeij, G.J. 1987. Evolution and escalation: An ecological history of life. Princeton: Princeton University Press. https://doi.org/10.1046/j.1420-9101.1988.1040369.x
Vermeij, G.J. 1994. The evolutionary interaction among species—selection, escalation, and coevolution. Annual Review of Ecology and Systematics 25: 219–236.
Walaszczyk, I. 1988. Inoceramid stratigraphy of the Turonian and Coniacian strata in the environs of Opole (Southern Poland). Acta Geologica Polonica 38: 51–61.
Walaszczyk, I. 1992. Turonian through Santonian deposits of the Central Polish Uplands; their facies development, inoceramid paleontology and stratigraphy. Acta Geologica Polonica 42: 1–122.
Walaszczyk, I. 1996. Inoceramids from Kreibitz-Zittauer area (Saxony and northern Bohemia): revision of ANDERT's (1911) description. Palaeontologische Zeitschrift 70: 367–392.
Walaszczyk, I., and W.A. Cobban. 2000. Inoceramid faunas and biostratigraphy of the Upper Turonian-Lower Coniacian of the Western Interior of the United States. Special Papers in Palaeontology 64: 1–118.
Walaszczyk, I., and C.J. Wood. 1998. Inoceramids and biostratigraphy at the Turonian/Coniacian boundary; based on the Salzgitter-Salder Quarry, and the Słupia Nadbrzeżna section, Central Poland. Acta Geologica Polonica 48: 395–434.
Walaszczyk, I., and C.J. Wood. 2018. Inoceramid bivalves from the Coniacian (Upper Cretaceous) Staffhorst shaft (Lower Saxony, Germany)—Stratigraphical significance of a unique succession. Cretaceous Research 87: 226–240.
Walaszczyk, I., L.F. Kopaevich, and V.N. Beniamovski. 2013. Inoceramid and foraminiferal record and biozonation of the Turonian and Coniacian (Upper Cretaceous) of the Mangyshlak Mts., western Kazakhstan. Acta Geologica Polonica 63: 469–487.
Walaszczyk, I., A.G. Plint, and N.H. Landman. 2017. Inoceramid bivalves from the Coniacian and basal Santonian (Upper Cretaceous) of the Western Canada Foreland Basin. In Allostratigraphy and biostratigraphy of the Upper Cretaceous (Coniacian–Santonian) Western Canada Foreland Basin, eds. N.H. Landman, A.G. Plint and I. Walaszczyk. Bulletin of the American Museum of Natural History 414. 53–103.
Walaszczyk, I., S. Čech, J.S. Crampton, Z. Dubicka, C. Ifrim, I. Jarvis, W.J. Kennedy, J.A. Lees, D. Lodowski, M. Pearce, D. Peryt, B.B. Sageman, P. Schiøler, J. Todes, D. Uličný, S. Voigt, and F. Wiese; With contributions by C. Linnert, T. Püttmann and S. Toshimitsu. 2021. The Global Boundary Stratotype Section and Point (GSSP) for the base of the Coniacian Stage (Salzgitter-Salder, Germany) and its auxiliary sections (Słupia Nadbrzeżna, central Poland; Střeleč, Czech Republic; and El Rosario, NE Mexico). Episodes 44: 129–150. https://doi.org/10.18814/epiiugs/2021/021022
Walliser, E.O., R. Mertz-Krause, and B.R. Schöne. 2018. The giant inoceramid Platyceramus platinus as a high-resolution paleoclimate archive for the Late Cretaceous of the Western Interior Seaway. Cretaceous Research 86: 73–90. https://doi.org/10.1016/j.cretres.2018.01.010.
Walliser, E.O., R. Vodrážka, N. Höche, S. Voigt, and B.R. Schöne. 2020. Late Turonian climate variability in the Bohemian Cretaceous Basin – A sclerochronological study of Inoceramus hercules shells from the Úpohlavy quarry (Czech Republic). Palaeogeography, Palaeoclimatology, Palaeoecology 560: 109996 (13pp.). https://doi.org/10.1016/j.palaeo.2020.10996
Ward, P.D. 1996. Ammonoid extinction. In Ammonoid paleobiology, eds. N.H., Landman, K. Tanabe and R.A. Davies. Topics in Geobiology 13: 815–824.
Wiese, F., S. Čech, B. Ekrt, M. Košt’ák, M. Mazuch, and S. Voigt. 2004. The Upper Turonian of the Bohemian Cretaceous Basin (Czech Republic) exemplified by the Úpohlavy working quarry: integrated stratigraphy and palaeoceanography of a gateway to the Tethys. Cretaceous Research 25: 329–352.
Wiese, F., S. Čech, I. Walaszczyk, and M. Košt’ák. 2020. An upper Turonian (Upper Cretaceous) inoceramid zonation and a round-the-world trip with Mytiloides incertus (Jimbo, 1894). Zeitschrift der Deutschen Gesellschaft für Geowissenschaften 171: 211–226.
Wilmsen, M. 2008. An Early Cenomanian (Late Cretaceous) maximum flooding bioevent in NW Europe: Correlation, sedimentology and biofacies. Palaeogeography, Palaeoclimatology, Palaeoecology 258: 317–333.
Wilmsen, M., and E. Nagm. 2014. Ammoniten. In Kreide-Fossilien in Sachsen, Teil 1, eds. B. Niebuhr and M. Wilmsen. Geologica Saxonica 60: 201–240.
Wilmsen, M., and B. Niebuhr. 2009. Exkursion 9–Die Kreide der Elbtalzone. Exkursionsführer der Deutschen Gesellschaft für Geowissenschaften 241: 199–218.
Wilmsen, M., and B. Niebuhr. 2014a. Die Kreide in Sachsen. In Kreide-Fossilien in Sachsen, Teil 1, eds. B. Niebuhr and M. Wilmsen. Geologica Saxonica 60: 3–12.
Wilmsen, M., and B. Niebuhr. 2014b. Stratigraphie und Ablagerungsbedingungen der Kreide in Sachsen (Elbtal-Gruppe, Cenomanium–Unterconiacium). Geologica Saxonica 60: 347–369.
Wilmsen, M., B. Niebuhr, and M. Hiss. 2005. The Cenomanian of northern Germany: Facies analysis of a transgressive biosedimentary system. Facies 51: 242–263.
Wilmsen, M., B. Niebuhr, M. Fengler, T. Püttmann, and M. Berensmeier. 2019a. The Late Cretaceous transgression in the Saxonian Cretaceous Basin (Germany): Old story, new data and novel findings. Bulletin of Geosciences 94: 71–100.
Wilmsen, M., M. Dölling, M. Hiss, and B. Niebuhr. 2019b. The lower Upper Cretaceous of the south-eastern Münsterland Cretaceous Basin, Germany: facies, integrated stratigraphy and inter-basinal correlation. Facies 65: 13/1-13/29.
Wilmsen, M., D. Schumacher, and B. Niebuhr. 2021. The Early Cenomanian crippsi Event at Lüneburg (Germany): Palaeontological and stratigraphical significance of a widespread Late Cretaceous bioevent. Palaeobiodiversity and Palaeoenvironments 101: 927–946. https://doi.org/10.1007/s12549-020-00459-8.
Wolansky, D. 1932. Die Cephalopoden und Lamellibranchiaten der Ober-Kreide Pommerns. Abhandlungen des Geologisch-Paläontologischen Institutes Greifswald 9: 1–72.
Wood, C.J., G. Ernst, and G. Rasemann. 1984. The Turonian-Coniacian stage boundary in Lower Saxony (Germany) and adjacent areas: The Salzgitter-Salder Quarry as a proposed international standard section. Bulletin of the Geological Society of Denmark 33: 225–234.
Wood, C.J., and G. Ernst. 1998. Turonian–Coniacian of Salzgitter-Salder. In Key locations of the Northwest European Cretaceous, eds. J. Mutterlose, A Bornemann, S. Rauer, C. Spaeth and C.J. Wood. Bochumer Geologische und Geotechnische Arbeiten 48: 94–102.
Woods, H. 1912. A monograph of the Cretaceous Lamellibranchiata of England. Inoceramus (continued). Monographs of the Palaeontological Society 2 (8): 285–340. https://archive.org/details/monographofcreta02wood
Acknowledgements
Constructive reviews by J. Todes (Chicago) and I. Walaszczyk (Warsaw) are gratefully acknowledged. Furthermore, I. Walaszczyk is thanked for further discussion in the aftermath of the review process. We also thank R. Winkler and M. Röthel (Senckenberg Naturhistorische Sammlungen Dresden) for digital photography of the MMG specimens and collection work, respectively. D. Erler (Senckenberg Naturhistorische Sammlungen Dresden) helped to obtain important literature. B. Gaitzsch and I. Kogan helped with the photographic documentation of the specimens housed in the geological collection of the TU Bergakademie Freiberg. C. Heunisch and A. Bornemann (Geozentrum Hannover) supported the collection work in the BGR in various ways. Our final thanks go to M. Amler (Köln) und M. Reich (Munich) for the editorial handling.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Michael Amler.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
Cite this article
Niebuhr, B., Wilmsen, M. Large-sized Late Turonian–Early Coniacian (Late Cretaceous) inoceramid bivalves from Germany: taxonomic issues, temporal framework and palaeoecological implications. PalZ 97, 217–243 (2023). https://doi.org/10.1007/s12542-022-00615-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-022-00615-9