
Abstract
Cattle and, to a lesser extent, sheep/goat and pigs, were crucial components of Norway’s socio-cultural and economic dynamics in the Middle Ages. Zooarchaeological analyses of faunal samples from different urban sites reveal that changes in cattle husbandry practices occurred in Norway over the medieval period, and especially in its early phases. This is clearly demonstrated in the case of medieval Oslo, where an in-depth zooarchaeological study provides the first comprehensive review of cattle husbandry strategies. In the first centuries of the medieval period (11th -mid 12th c. AD), cattle were mainly raised for milk and meat production, while fewer young and more numerous older individuals, most of which probably oxen, were present from mid 12th -13th c. AD onwards. This hypothesis is supported by biometrical analyses of cattle postcranial bones, indicating changes in the sexual composition of cattle herds over time. Therefore, from ca. mid 12th -13th c. AD onward, cattle seem to have been more used for ploughing and, only once old, to have been culled for their meat. In turn, this evidence suggests that more arable lands had to be ploughed, especially in eastern and central Norway. Here, demographic growth and settlement expansion would have led to an increase in food demand; at the same time, more favourable climatic conditions might have contributed to an increase in the availability of arable fields. This article suggests that medieval Norwegian farming was not static as historically thought, but rather connected to wider-scale agricultural innovations characterising other parts of medieval Europe.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cattle, sheep/goat, and pig remains have rarely been used to investigate dietary habits as well as socio-cultural, economic, and environmental dynamics in medieval Norway. With few exceptions (e.g. Lie 1979; 1988; Lie and Lie 1990; Hufthammer 2000; 2003; Lie and Lie 2013; Walker et al. 2019; 2020; 2021), most zooarchaeological studies from medieval sites in southern-central Norway are presented as concise technical reports, offering essential yet significant information. This is particularly valid for the zooarchaeological studies of medieval Bergen, Oslo, and Trondheim; here, archaeological excavations conducted in the 1970–80s recovered large faunal samples currently stored at the Department of Natural History-University Museum of Bergen (Norway). The related zooarchaeological reports, however, often lack a broader contextualisation within the historical and archaeological framework, as well as comparisons with additional samples. The FOODIMPACT Project, funded by the Research Council of Norway and of which this study is an integral part, represented an excellent opportunity to fill such a gap (https://www.khm.uio.no/english/research/projects/foodimpact). As the first study with a special focus on medieval Norwegian urban food culture, the project aimed to provide an in-depth analysis of food consumption patterns at urban sites through a multidisciplinary approach, including the study of animal remains, plants, pottery, organic residues (Garnier and Vedeler 2021; Vedeler and Garnier 2024; Vedeler et al. 2023; Martínez-García et al. in prep. a, b; Fuglevik in prep; Vedeler and Bergstøl in prep), and written sources (Vedeler 2023). The project placed special emphasis on medieval Oslo. The archaeological excavations conducted almost continuously in the ancient part of the city over the last 30 years, coupled with its numerous attestations in written sources (e.g. sagas, annals etc.), make Oslo an ideal laboratory for the study of the Norwegian Middle Ages (Christie 1966). Aligned with the primary objectives of the FOODIMPACT Project, Oslo is the central focus of this zooarchaeological study, with a specific emphasis on the faunal material from the urban site of Oslogate 6 - one of the largest samples ever recovered from the medieval city (Archive Department of Natural History-The University Museum of Bergen). Faunal reports from contemporary urban sites in Norway have also been analysed to facilitate comparisons among site-types, regions, and chronological periods. Although large-scale comparisons are presented, this paper does not aim to provide an exhaustive review of medieval Norwegian animal husbandry and dietary practices. Rather, the focus is on presenting a targeted zooarchaeological investigation, emphasising the pivotal role of the main domesticates (cattle, sheep/goat, and pig) in food production, with particular emphasis on cattle. The study offers insights into how cattle, among other factors, significantly influenced the socio-cultural, economic, and environmental dynamics of urban societies in medieval Norway.
Archaeological and historical background
The growth of larger towns such as Bergen, Oslo, Tønsberg, and Trondheim as well as smaller ones like Stavanger in the late 10th -early 11th c. AD is intricately connected with the establishment and consolidation of the national kingdom, the development of trade, and the gradual adoption of Christianity by the Norwegian population (Hansen 2017). Notably, the construction of buildings specifically designated for the use of the monarchy and the Church played a pivotal role in defining the core of these urban settlements, shaping them with an entirely new socio-economic identity (Hill 2010). Archaeological excavations conducted from the 1970s onward, suggest that the first major urban development in Oslo took place during the first half of the 11th c. AD (Christie 1966; Schia 1989; Nordeide and Gulliksen 2007). In the early years of its development, Oslo possibly served as a local trading hub or a designated landing site. During this period, the town was repeatedly destroyed by fire and rebuilt several times. By the mid-12th c. AD, the Church’s independence and subsequent growth played a crucial role in fortifying the town’s political and economic position in the region. Indeed, it was in this period that the city became the seat of the east-Norwegian episcopacy. Despite its status as a bishop’s seat and royal residence, the development of the town of Oslo proceeded rather slowly during its first 200 years (Hill 2010).
However, in the 13th -early 14th c. AD, the Oslofjord region, an inlet stretching from Oslo to the Skagerrak strait on the North Sea, assumed a position of paramount importance within Norway, owing to its favourable proximity to Denmark and southern Sweden; similar considerations apply to Tønsberg, located 100 km south of Oslo. During the reign of Håkon V Magnusson (AD 1299–1319), the functions of the capital were divided between Oslo and Bergen (Blackmore 1994). In AD 1349, Oslo was hit by a plague outbreak that probably killed more than 50% of the city’s inhabitants, thus having a devastating impact on the city’s economy and social structure (Nedkvitne and Norseng 2000). The city was also affected by wars and political turmoil. At the end of the 14th c. AD, Oslo lost its political and administrative function when Norway entered the Kalmar Union with Denmark and Sweden (AD 1397–1523), and the official capital was relocated to Copenhagen. Following the dissolution of the Kalmar Union in AD 1523, Norway and Denmark established a personal union.
After a devastating fire in AD 1624, the medieval city of Oslo was abandoned, and a new town named Christiania was established near the old settlement (Christie 1966; Blackmore 1994).
Several archaeological excavations were conducted in the medieval town during the 1970–80s, revealing some of the old structures of the city. Among these areas, archaeologists concentrated on the site of Oslogate 6, known for its historical and archaeological significance. The site, situated on a slope, occupied a position overlooking the bay of Bjørvika (Fig. 1) and was bordered by the Hovinbekken river to the north. Here, archaeological excavations unveiled a stratigraphic sequence encompassing two pre-urban and seven urban phases, all culminating in a fire event (Table 1). The relative chronology of each phase is based on a sequence of fire events, while the absolute chronology primarily relies on the dating of pottery, combs, and shoes. 14C-datings were performed and were especially valuable for dating the oldest urban phases (Phases 1–4; Molaug 2015; Molaug et al. 2018). The settlement area of Oslogate 6 consisted of five tenements; Tenements 2, 3, and 4 underwent extensive excavations, while Tenements 1 and 5 were only partially investigated. All tenements were likely connected by a street, possibly Nordre strete, which is mentioned in late medieval historical documents (Smedstad 1991). The earliest occupation of the site dates to the 11th c. AD; in this period human settlement appeared to have been rather sparse and most of the area was uninhabited. Archaeological features, interpreted as the remains of fences and/or ditches, indicated that the tenement areas were defined by wooden perimeters, which did not undergo severe modifications throughout the medieval period. Historical sources (Frimanslund 1961) suggest that such fences were used to separate fields and/or for enclosing animals; similar structures were also found at other contemporary sites in Oslo (e.g. Arups gate site; Schia 1987; Martens 2010). In the first decades since its foundation, metalworking activities characterised the Oslogate 6 site. Later, in the late 12th c. - mid 13th c. AD, the area was transformed into a shoemaker centre as suggested by the large number of discarded leather fragments recovered. In the 13th -14th c. AD, the site’s architectural changes followed a similar pattern as was detected in other parts of medieval Oslo. During this period, the city underwent significant urban growth, which encompassed both population growth and the expansion of physical infrastructure. New residential areas, workshops, and commercial spaces were established. Streets and marketplaces were developed or expanded to accommodate the growing trade and to facilitate the movement of goods and people. The city’s expansion also necessitated the construction of new buildings, including warehouses, harbours, and administrative structures. Overall, the urban growth of Oslo in the 13th - early 14th c. AD was driven by the city’s economic potential, attracting settlers from various backgrounds. The increasing population provided a labour force for the expanding industries and contributed to the growth of local markets. In contrast to the extensive information available for the 13th - early 14th c. AD, there is a relatively limited amount of archaeological and historical evidence available regarding Oslogate 6 in the later medieval phases (AD 1350–1550; Molaug 2015; Molaug et al. 2018). Such a paucity of information might be related to the gradual decline of medieval Oslo but also to less favourable conservation conditions of the archaeological context (Vedeler pers. comm.).

Materials and methods
The faunal material from Oslogate 6 is currently stored at the Department of Natural History-The University Museum, University of Bergen. The sample was previously studied in the 1980s, but a more selective and in-depth analysis was carried out due to the limited availability of crucial zooarchaeological information (e.g. the recording of mandibular tooth wear stages, biometrical and butchery data), which were key to the FOODIMPACT Project research questions. This second investigation specifically focuses on the study of the main domesticates (cattle, sheep/goat, and pig). Animal remains were anatomically and taxonomically identified using the zooarchaeological reference collection held at the Department of Natural History-The University Museum of Bergen (Norway). During this step, animal bone atlases were also consulted (Schmid 1972; Barone 1976). The recording was conducted following the Diagnostic Zone Method and a 50% integrity threshold was applied (Albarella and Davis 1994), thereby minimizing interdependence problems (Grayson 1984; Lyman 2008).
In this article, sheep (Ovis aries) and goat (Capra hircus) remains have been analysed together; a forthcoming article will reconstruct in detail their exploitation practices in Oslo during the Middle Ages (Aniceti et al. in prep.).
Both archaeological and historical sources indicate that the wild boar (Sus scrofa) had disappeared from Norway before the Middle Ages as a consequence of agricultural expansion; this led to the fragmentation of suitable habitats and an intensified hunting pressure due to the expanding human population (Rosvold et al. 2010). Therefore, the suid remains from Oslogate 6, as well as those recovered at other medieval sites included in this study, most likely refer to domestic pigs (Sus scrofa domesticus). This is further corroborated by the consistent predominance of immature individuals in all sites, a well-known pattern of domestic pig culling suggesting that most pigs were domestic animals subjected to a planned culling strategy.
The Number of Identified Specimens (NISP) was used to quantify the relative proportion of each taxon; the NISP represents the raw count of specimens representing the selected diagnostic zones and considered ‘countable’ (Aniceti and Albarella 2022).
As mentioned before, the faunal sample from Oslogate 6 was examined in conjunction with other medieval faunal samples from different urban sites in Norway that had previously been recorded, in order to allow inter-site, regional and chronological comparisons. In addition, an in-depth review of the relevant zooarchaeological literature was carried out (Fig. 2; Table 2). Such a comparative study focuses on the taxonomic frequency of the main domesticates as well as on cattle ageing profiles. Different protocols were employed to record animal bones and teeth from these additional assemblages. To ensure reliable comparisons between sites and time periods, precautionary measures were taken. For example, only samples providing data on recorded anatomical parts and indicating the preservation level of each element were chosen. Among these, only parts corresponding to the diagnostic zones selected for the study of the Oslogate 6 material and exhibiting a preservation level exceeding 50% of the original bone size, were included in the quantitative analyses. Moreover, only faunal samples with > 100 remains were selected.

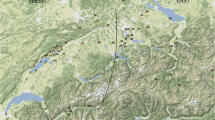
Map of Norway indicating the medieval urban centres from which the faunal samples included in this study have been recovered as well as the regions of Østlandet and Trøndelag (adapted from d-maps.com; for further details see Table 2)
In most cases, cattle age-at-death was estimated by analysing the epiphyseal fusion stage of postcranial bones, while mandibular wear stages were only available for Oslogate 6. To reconstruct mortality profiles, post-cranial bone fusion data were divided into three fusion events following Silver (1969). Mandibular tooth wear was recorded using Grant (1982), and mandibular wear stages were then determined using O’Connor (1988); these were estimated only for jaws with at least two recordable teeth. When the furthest tooth in the jaw could not be recorded or was not present, the complete tooth wear stage sequence was reconstructed using Grant’s correlation Table (1982). Kill-off patterns were reconstructed when the number of available mandibular wear stages was ≥ 20.
At Oslogate 6, pathological evidence on cattle was systematically recorded; such evidence is used in this paper to complement the results from other zooarchaeological analyses.
A biometric analysis of cattle postcranial bones from Oslogate 6 was undertaken to examine potential variations in size and shape, sexual composition, and the potential presence of multiple cattle groups, as well as to provide further ageing information. Cattle size in medieval Norway has been previously analysed at some sites using withers heights (Lie 1988, 1989; Undheim 1985). However, the overall scarcity of raw data from comparative sites compromised inter-site and regional comparisons. Furthermore, while withers heights provide useful information on overall body size, they do not allow the reconstruction of body shape. In this study, measurement values from different axes were used to investigate both body size and shape, thus providing a more complete picture of morphometric characters. Measurements were taken following von den Driesch (1976) and Davis (1996) on fully fused bones, while fusing or unfused specimens were excluded from analyses. Measurements from different elements were plotted on the same scale using a size index scaling technique (the log ratio or LSI), which allowed to maximise the informative potentials of biometrical datasets (Simpson et al. 1960; Albarella 2002). This method involves calculating the logarithm of the ratio between archaeological and standard values (Meadow 1999). The standard values used in this study refer to the means of measurements of cattle bones from Early Anglo-Saxon West Stow (Suffolk, United Kingdom) as reported in Rizzetto and Albarella (2022); the list of measurements used in the histograms is provided in Table 3. Though useful, size index scaling techniques come with limitations. Measurements from various anatomical elements can be influenced differently by factors like age, environmental conditions, and sex. As a result, combining all measurements on a single scale can obscure factors influencing metric variation. To address this, measurements should be chosen based on the research question (Payne and Bull 1988) and analysed separately if they lie on different axes. Due to the scarcity of width and depth measurements, they were combined in this study, while the rarer length measurements were excluded. In addition, it was decided to not include the smallest diaphysis widths of long bones (SD), as well as the smallest length of collum scapulae (SLC), as they are especially affected by ageing.
In this study, the decision was made to focus on width/depth measurements, opting to combine them to avoid an unwarranted reduction in the sample size, while excluding the potential biases derived from the inclusion of length measurements. Only one value per specimen was selected. Biometric analyses are supported by statistical ones. The statistical approach utilised in this study encompasses the use of the two-tailed Student’s t-test. Such a test is known for its robustness, indicating that it is minimally affected by variance (Simpson et al. 1960). In data analysis, statistical tests have traditionally relied on fixed thresholds, used to assess differences between groups of data (e.g. p > or < 0.05); these determine whether differences are statistically significant or not. This study, however, avoids the arbitrary use of such thresholds by directly commenting on the lower or higher values of p. The main aim behind the adoption of this approach is to consider statistical analyses as complementary instruments that can be scrutinised and critically interpreted alongside the graphic representations of biometrical analyses (Stern 2016; Imbens 2021). Although statistical significance can serve as a useful benchmark for decision-making or inference, it should not serve as the sole criterion for interpreting research findings especially when, as in this study, multiple lines of evidence (ageing, biometry and, although to a lesser extent, pathology) contribute to the assessment of change or continuity in animal use.
Due to the absence of detailed chronological information at certain comparative sites, only a limited number of faunal samples could be examined across different phases. At some sites, chronological phases had to be merged for the sake of facilitating comparisons (e.g. Phases 2 and 3). To facilitate comparisons, all analysed sites/faunal samples were organised into these chronological phases:
-
Phase 1: AD 1000/1100–1150 (Early Middle Ages, EM)
-
Phase 2: AD 1125/1150–1250 (Early Middle Ages, EM)
-
Phase 3: AD 1250–1350 (Early Middle Ages, EM)
-
Phase 4: AD 1350–1550 (Late Middle Ages, LM)
Results
Taxonomic frequency
This section presents the taxonomic frequencies of the main domesticates at Oslogate 6 and compares them with those from other urban sites. Additional faunal samples from monastic sites, and from other urban sites broadly attributed to the medieval period (Table 2), are analysed in a ternary plot. This approach allowed for a further investigation of potential variations in species frequency across different site types, providing complementary information on animal husbandry dynamics in medieval Norway.
In all Oslo early medieval sites, cattle are the predominant species (Table 4a-c and Fig. 3). In Phase 1, sheep/goat and pigs are also well-represented (Table 4a; Fig. 3). In Phases 2–3, the incidence of sheep/goat increases primarily at the expense of pig and, to a lesser extent, of cattle (Table 4b-c and Fig. 3). Early medieval Tønsberg (Table 5; Fig. 4) and Trondheim (Table 6; Fig. 5) show an analogous pattern to Oslo’s. A similar consideration is valid for early medieval Bergen, although in this case sheep/goat appear to be more abundant compared to other sites; such a higher frequency also characterises the late medieval period (Table 7; Fig. 6). With the exception of pig frequencies, which show a continuous decline over time, no significant changes between the early and late medieval phases are observed (Table 4d, 6; Fig. 7). Overall, taxonomic frequency data reveal a relatively consistent pattern, wherein the main domestic animals are similarly represented across all the analysed sites and phases. This observation is reinforced by the ternary plot, which highlights a predominantly uniform pattern, revealing no discernible variations among urban and monastic sites (Fig. 8). The combination of various medieval phases into a single period in the ternary plot may have impacted the resolution of the results. Nevertheless, the in-depth chronological analyses conducted for multiple sites in Oslo, Tønsberg, Trondheim, and Bergen largely substantiate this overall homogeneous trend.






Ternary plot - NISP frequencies of cattle, sheep/goat, and pig at all the analysed sites, including monastic sites and those generally dated to the medieval period (raw data from: Brendalsmo 1986; Eriksson 1975; Hufthammer 1999; Lie 1979; 1988; 1989; 1991; Lindh 1984; Marthinussen 1992; Nordman et al. 1986; Undheim 1985; Wienberg 1983; Wiig 1981; Aniceti et al. this article; Archive Natural History – UMB).
Ageing
Early medieval period; Phase 1 (AD 1000–1150)
Cattle culling profiles at urban Oslo and Trondheim show similar patterns (Fig. 9a). At Oslogate 6, early culling events, which refer to animals culled in their 1st -2nd year of life (calves), account for almost 40%; Televerkstomten and Erkebispegården (Trondheim) show a similar incidence of early culls. At Oslogate 6, foetal individuals were also recovered, thus suggesting on-site breeding. At Oslogate 6, ca. 20% of individuals are slaughtered by their 2nd -3rd year of age (middle fusing stage), and ca. 15% during their 3rd -4th year of life (late fusing stage); similar considerations apply to Televerkstomten and Erkepispegården. At all the three sites, ca. 20% of the individuals survives into late adulthood (beyond the late fusing stage; ≥3–4 years of age). The kill-off pattern reconstructed from the mandibular wear stages largely corroborates the results of the epiphyseal fusion analysis (Fig. 10). Overall, during the early centuries of the medieval period in Oslo and Trondheim, cattle use appears to have focused on cows for milk, cheese and butter production.


Frequency of cattle mandibular wear stages (MWS; O’Connor 1988) at Oslogate 6. Phase 1 (n:20), Phase 2 (n:59), Phase 3 (n:22), and Phase 4 (n:15)
Early medieval period; Phases 2,3 (AD 1200–1350)
At Oslogate 6, a higher number of cattle are raised to full adulthood in comparison to the previous phase (ca. 60%), and fewer early culling events are present (ca. 20%; Fig. 9b, c). Data on mandibular wear stages support this observation (Fig. 10). An even higher incidence of adults is observed at Oslogate 7 and Mindets Tomt/Søndre Felt, where ca. 70% and ca. 65% of cattle are culled after their 3rd -4th year of age. Similar patterns are evident in Tønsberg, except for Baglergaten 3; here, early culls are similarly scarce but over ca. 80% of cattle were culled before reaching adulthood, suggesting a potential emphasis on meat production. A similar trend to that of Baglergaten 3 is also observed at contemporary Dreggen (Bergen). At Trondheim (Televerkstomten and Erkebispegården) there is a similar scarcity of early culls (< 10%), although the same amount of individuals survive into adulthood as in Phase 1 (ca. 45%). During this period, cattle husbandry practices in Oslo and Trondheim appear to be no longer primarily on dairy products. Instead, there is a specialisation in the use of cattle for its traction force. This trend is also visible in Tønsberg, with the exception of Baglergaten 3, and to a lesser extent in Bergen, where a distinct interest in beef and dairy production is evident.
Late medieval period; Phase 4 (AD 1350–1537)
In Oslo and Trondheim, cattle husbandry practices appear similar to those of the previous phase. There is a slight increase in the number of individuals kept until late adulthood (ca. 70%), and early culling events have become even less frequent (< 5%; Fig. 9d). Similar patterns are reflected in the mandibular wear stage data (Fig. 10). In Bergen, however, a larger proportion of individuals are slaughtered before reaching their 3rd -4th year of age compared to the previous phase (ca. 80 and 95% at Rosenkrantzgt 1–2).
During the Late Medieval period, therefore, at both Oslo and Trondheim cattle continued being raised for secondary products beyond as well as dairy production, i.e. for their traction force. These animals were probably mainly used for ploughing the fields and for breeding purposes, reflecting a specialised economy that had already developed during the Phases 2–3 of the Early medieval period (although more abruptly in Oslo, and more gradually at Trondheim). In contrast, Bergen’s cattle economy appears to have maintained its emphasis on meat production, a trend observed since the Early medieval phases.
Pathology
Pathological conditions on cattle postcranial bones are mostly present in Phases 2–3, accounting for ca. 2% of the total cattle specimens in both phases. In Phase 4, fewer pathological evidence is observed, and none are found in Phase 1. Pathological evidence has been exclusively recorded on cattle metapodials and on phalanges (Table 8). The intensive and specialised butchery practices on metapodials and on other long bones, resulting from marrow extraction, might have limited the visibility of pathological evidence, possibly suggesting a quantitative underrepresentation of the features described in Table 8.
Biometry
Overall, the log ratio histograms of cattle post-cranial bone width and depth measurements at Oslogate 6 display a unimodal distribution of data in all phases (Fig. 11). Cattle increase in size from Phase 1 to Phase 2. A slight further increase in size is observed in Phase 3, while in Phase 4 individuals show a similar size to those from the previous period. The p-values obtained from the Student’s t-test are generally low (p < 0.05), indicating significant differences between datasets from consecutive phases. This suggests that changes in cattle from one phase to the next reflect actual differences in the population. The exceptions are Phase 2 vs. Phase 3, which show marginally significant differences, and Phase 3 vs. Phase 4, where no statistically significant differences were observed (SI, Fig. 1, Table 2). The extensive dataset from Oslogate 6 allowed to separately analyse widths/depths from the most sex dependent bones (i.e. humeri, metacarpals, and metatarsals). This analysis evaluates the influence of potential changes in the sex ratio in the assessment of overall size changes among phases.

Distribution of log ratio values for cattle post-cranial bone measurements (humerus, metacarpal, tibia, astragalus, calcaneum, metatarsal widths and depths) from Oslogate 6; Period 1 (n:65), Period 2 (n:200), Period 3 (n:145), and from Period 4 (n:127). The stars indicate the logarithmic means; the standard values used are the means of measurements from a sample of cattle bones from Early Anglo-Saxon West Stow (Suffolk, UK; Rizzetto and Albarella 2022)
As illustrated in Fig. 12, during Phase 2 and, to a lesser extent, Phases 3 and 4, the data exhibit a bimodal distribution, with distinct groups of larger values – likely castrated males (oxen), plus a few breeding bulls. A similar group of larger animals is almost absent in Phase 1. This is reflected by the logarithmic means: the mean does not change in Phase 1 when all values and only those from the most sex-dependent measurements are considered (-0.03), while it is higher in the samples of sex-dependent measurement values in Phases 2–4. This is the result of the higher incidence of (castrated) males in the latter phases, as evidenced by the groups of larger values in Fig. 12. Therefore, it is likely that differences in the proportion of cows and oxen/bulls contribute to the overall variations in size observed in Fig. 11. At the same time, the presence, in Phases 2–4, of multiple cattle group(s) of different size, thus influencing the distribution of data in conjunction with a different sex composition, cannot be entirely ruled out.

‘Distribution of log ratio values for cattle post-cranial bone measurements (humerus, metacarpal, metatarsal widths and depths) from Oslogate 6’; Period 1 (n:24), Period 2 (n:65), Period 3 (n:52), and from Period 4 (n:48). The stars indicate the logarithmic means; the standard values used are the means of measurements from a sample of cattle bones from Early Anglo-Saxon West Stow (Suffolk, UK; Rizzetto and Albarella 2022)
Discussion
Cattle played a central role in medieval Norway’s food production practices and socio-economic dynamics; in both the Early and Late medieval periods, these were indeed the most abundant domesticates, followed by sheep/goat and pigs at Oslo, Tønsberg, Trondheim, and Bergen. Through time, sheep/goat increased at the expenses of pig. Similarly, a decline in the incidence of pigs is observed at other contemporary northern European sites in Sweden and England (Vretemark 1997; Hamilton and Thomas 2012; Grau-Sologestoa and Albarella 2019). After the 14th c. AD, pigs also became very rare or absent in Iceland and Greenland as well (McGovern et al. 2014). In both Sweden and England, this development seems to find its roots in key changes in the landscapes, which impacted on free-range pig husbandry practices. The massive deforestation resulting from the expansion of urban settlements and cultivated areas severely diminished the availability of forests. Such landscape changes might have favoured the transition from free-range pig husbandry to keeping these animals primarily indoor; while this development was facilitated by the increased availability of food waste in urban areas, which could be used to feed pigs, it also facilitated keeping a smaller number of animals. A similar hypothesis might also apply to medieval Norway, or at least to some areas. Pigs in 12th -13th c. AD Bergen exhibited notably high δ13C and δ15N values, suggesting that their main source of sustenance was marine fish leftovers, namely food waste, rather than plant-based food sources which characterise free-range husbandry practices (Halley and Rosvold 2014). Socio-cultural and religious factors might also explain pig decline in medieval Norway. In the period spanning from AD 930 to 1030, Norway gradually started its conversion to Christianity. During this process, food taboos were introduced into Norway, consisting of prohibitions on the consumption of meat from specific animals. Written sources, such as the medieval Islandic Grágás (e.g. Kristinna laga þáttr) and ecclesiastic regulations (kristinn réttr; e.g. the Gulathing, Frostathing, Borgarthing, and Eidsivathing laws), discuss various types of tabooed species. Although pig was not generally forbidden, its consumption was prohibited under certain circumstances, such as when pigs consumed horse or human flesh (Helle 2001; Sanmark 2002, 2005; Bourns 2018; Maraschi 2020). However, these conditions do not seem to be so common as to explain a substantial decline in pork consumption.
While the incidence of cattle, sheep/goat, and pig changes slightly but consistently across phases and various types of medieval sites, different patterns in the utilisation of cattle have been observed. At Early medieval Oslo and Trondheim (Phase 1; AD 1000–1150), cattle husbandry practices seem rather generalised, with beef and secondary products playing equally significant roles. However, the high number of foetal - suggesting onsite breeding - and young individuals at Oslogate 6 and Trondheim, alongside the presence of adults/elderly animals, may suggest a strong emphasis on dairy production (i.e. calves being culled to maximise milk exploitation from adult cows). In this context, it is likely that butter production played a central role in the site’s economy. Indeed, butter has always been an important commodity in Norway: it served multiple purposes, being used as a source of food, an export commodity, and even as a form of currency. In Norway, Sweden, and Denmark, as well as in other northern European countries such as England, butter made from cow’s milk was considered a high-status food and, unlike meat, it was not subject to clear religious restrictions (Woolgar 2016; Vedeler 2023). Its popularity stemmed from its ease of preservation and high energy content. This made it an ideal food source, particularly in a country, as Norway, where short growing seasons and crop failures were common (Challinor 2004). Proper storage methods were employed to reduce microbial activity, allowing butter to be preserved for extended periods. Consequently, converting milk into butter, especially in preparation for the winter months, represented a wise strategy for effective food resource management (Critch et al. 2018).
Compared to the preceding period, cattle husbandry practices in 12th -13th c. AD Oslo exhibit significant changes, with a similar, albeit milder, shift also observed in Trondheim. These changes consist of an increased emphasis on keeping animals until they reached their late adulthood, while reducing the culling of young and subadult individuals. A similar situation was also observed in Tønsberg, with the sole exception of Baglergaten 3, where a noticeable focus on meat production is evident. As the Middle Ages progressed (14th -early 16th c. AD), a greater number of animals were kept until they reached old age, resulting in a diminished presence of young and subadult animals. This shift, which started in Oslo and Trondheim in the 12th -13th c. AD, may suggest a potential change in the role of cattle from food providers (milk, meat) to the utilization of their traction force in agricultural activities. Bergen, showing a more specialised interest in meat production, is the only exception to this trend.
It can be argued that such a general decrease of neonatal and young individuals is the result of taphonomic processes affecting the more fragile bones of immature animals. While taphonomic processes certainly led to an underrepresentation of younger animals in all sites and phases, their relative decrease after Phase 1, occurring nearly simultaneously across different Norwegian sites, strongly suggests otherwise. The following paragraphs investigate how the use of draught animals in agricultural works can be identified, and how and why such use could have intensified in medieval Norway.
Identifying the use, spread and the intensification of cattle traction technology is a rather debated topic in archaeology. In zooarchaeology, multiple lines of evidence can be used, in combination, to validate the identification of traction animals. The evidence from taxonomic frequencies and ageing data has already been discussed; biometrical analyses can also contribute to such studies. The scarcity of published biometric data from medieval sites in Norway prevents us from interpreting the results from Oslogate 6 in a broader regional context. Despite this, the large biometric dataset presented in this paper is a good starting point. Biometrical analyses suggest that cattle increased in size over the medieval period, and especially from the 13th to mid-14th c. AD. The analysis of the most sex-dependent measurements suggests that such an increase is likely the result of changes in livestock sex ratios, namely of a higher incidence of oxen. While archaeological and ethnographic studies have shown that cows, especially on smaller farms, were also used as draft animals (Johannsen 2011), historical records indicate that draught cattle primarily consisted of castrated males (Fowler 2002). Such a preference for oxen was due to their greater size and strength, which allow them to generate more power than cows, while remaining easier to handle than bulls (Holmes 2022). The increase in cattle size at Oslogate 6, however, does not necessarily rule out the possibility that cattle was also improved locally, for example by improving animal management and/or through selective breeding, or through inter-breeding local cattle with ‘larger’ types imported from Sweden or other nearby countries. Recently, Walker and Meijer (2020) presented zooarchaeological data indicating the existence of various domestic fowl types in medieval Norway; similarly, other domestic animals may have been represented by different types, especially in the case of the multi-purpose cattle.
In addition to ageing and biometry, the identification of draught cattle in zooarchaeology has traditionally relied also on the interpretation of specific pathological conditions (Groot 2005; Thomas 2008; Holmes et al. 2021). Indeed, cattle have not evolved for the specific purpose of pulling ploughs or carts, and it is widely acknowledged that the additional mechanical stress imposed by such activities can result in discernible effects on skeletal tissues (Lanyon and Rubin 1985). While some pathological evidence on cattle metapodials and phalanges at Oslogate 6 may be indicative of what are commonly referred to as ‘draught cattle signatures’ (Holmes 2022), these occurrences are relatively scarce in the whole assemblage, thus making it difficult to draw definitive conclusions. Moreover, other factors such as age, sex, castration, the timing of castration, soil conditions, nutrition, the nature and intensity of traction use, plough technology, and the presence and nature of stalling facilities can also play a role in the development of pathological conditions, thereby adding complexity to their interpretation (Thomas 2008). Conversely to ageing and biometric data, pathological conditions do not provide substantial evidence indicating a transition in cattle husbandry practices during the Early Medieval period - specifically, the shift from a focus on dairy production to a combined use of cattle as draught animals in arable farming. However, their exclusive occurrence in Phases 2–4 provides interesting complementary data in this direction.
The contextualisation of zooarchaeological data within the wider socio-economic and political framework of Norway and northern Europe may also offer important insights to further strengthen this hypothesis.
From the 9th to the late 13th c. AD, the medieval European economy experienced a remarkable increase in productivity. This shift, attributed to new agricultural technologies and land utilisation practices, has led many scholars to consider this period ‘revolutionary’ (White 1962; Langdon et al. 1997). Geographically located at the northernmost tip of Europe and characterized by an overall harsh physical environment and climate, medieval Norwegian agriculture and animal farming may seem marginal from a European perspective. In comparison to its Scandinavian neighbours, such as Sweden and Denmark, Norway only has ca. 3% of arable land and presents a much more varied and complex topography, with small margin for agricultural improvement and growth (Øye 2004). While the south-eastern and central parts of the country (Østlandet and Trøndelag, Fig. 2) are characterized by large areas and flat lands of marine and moraine deposits, the western regions offer a more uneven distribution of post-glacial deposits separated by fjords and small valleys (Øye 2009). In the past, the long winter season and abundant rainfall caused frequent crop failures in Norway. However, climatic conditions were more varied than they might appear, exhibiting significant variations between the north and south as well as between coastal and inland regions. Between the 9th and 14th c. AD, the climate was warmer and drier than it is today, reaching an optimum around the mid-10th -13th c. AD (the so called ‘Medieval Warm Period’); later, more humid, and colder climatic conditions developed. During the Middle Ages, the hilly terrain, and vast forests of south-eastern and central parts of Norway (Østlandet and Trøndelag, Fig. 2) offered more favourable agricultural conditions in comparison to other parts of the country, resembling those of northern England. In the Early medieval period, the emergence and development of urban centres and a warmer climate promoted demographic growth. Since the 13th c. AD, the Norwegian population doubled and, in certain areas even tripled, with the eastern regions including the most densely populated areas (Øye 2013).
Given the growing population and the higher food demands, the necessity to transform less fertile areas into cultivatable land became important, thereby promoting clearances and advances in agricultural technology. Compared to the previous period, palaeobotanical studies have attested more diversified cereal productions in 12th -13th c. AD Norway (Øye 2013). Such diversification also extends to vegetables and fruits. In the regions of eastern and central Norway, known for their larger arable areas, medieval legal codes mandated that one-fourth of arable land remain fallow, a measure aimed at preventing soil depletion. The cultivation of different species with varying characteristics emerged as a novel strategy. Although productivity levels may not have been exceptionally high, people appear to have maximised the utilisation of cultivatable lands while minimising the risks (Øye 2004). The impact of the introduction of the heavy plough in northern European land use practices remains a debated topic among scholars (White 1962). This plough was more robust and capable of turning over heavy and wet soils, therefore being particularly suitable for the clayey soils prevalent in northern Europe. It replaced the earlier ard (a lighter plough) and enabled more extensive cultivation of the land. The use of the heavy plough in Scandinavia is scarcely documented in archaeology, and it is only vaguely mentioned in 13th c. AD written sources. Based on the scarce archaeological evidence, it appears that Norway and other Scandinavian nations embraced heavier ploughs several centuries after their implementation in southern Europe, roughly between the 11th -13th c. AD, although lighter ploughs remained in use. The presence of solid horseshoes in Early Medieval contexts served as archaeological indicators to suggest the utilisation of horses as the main draught animals (Øye 2009). However, this has never been proved zooarchaeologically. Although it is possible that some horses were used as working animals, their higher maintenance costs, coupled with the cultural reluctance to consume them at the end of their working life as opposed to cattle, made them less economically viable to be reared in large numbers. Towards the end of the 13th c. AD, Norway’s arable farming had achieved a technological state close to that of the period predating the ‘agricultural revolution’ in the 18th and 19th c. AD, with cattle being a key part of such achievements. The obtained zooarchaeological data suggest that, indeed, cattle played a major role as draught animals in the Early medieval period. Their increased use in agricultural works, observed between the 12th and 14th c. AD, coincided with a series of events (warmer period, population growth, establishment and expansion of urban centres, surplus production, trade) for which this animal had become essential for maximising agricultural outputs. Within this context, the fact that the shift towards a use of cattle as draught animals to be used in the field occurs in Oslo and Trondheim, where most of the arable land was available, while western sites such as Bergen - which at that time was a major town - focused on meat production, further supports such interpretation. The shift in cattle exploitation strategies - from being primarily used as a source of food to playing a more important role in agriculture - was therefore instrumental in Norway’s agrarian economic development.
Conclusions
Cattle played a pivotal role in medieval Norway, evolving from an important source of meat, dairy products to indispensable working animals, contributing to agricultural activities before being culled for their meat. In the early centuries, they mainly provided milk, which could be transformed into precious butter, and, to a lesser extent, meat, meeting the nutritional and economic needs of local communities. As agricultural production expanded, cattle took on a dual role, being more intensively used in the fields. The combination of zooarchaeological evidence presented in this paper, encompassing taxonomic frequencies, ageing data, biometrical analyses, and pathological conditions, reveal and describe in detail such process.
The unique landscape of eastern and central Norway, characterized by a hilly terrain and fertile areas, played a crucial role in this transition. Here, cattle became essential draught animals, ploughing fields and fulfilling various agricultural tasks. Favourable climatic conditions further enhanced agricultural production and, hence, the use of cattle in the fields, allowing communities to refine and develop farming technologies.
This study focused on eastern Norway, examining site-specific data, and establishing a foundation for more comprehensive research into medieval Norwegian animal husbandry. Future investigations will need to delve deeper into the intertwined relationships between animal exploitation, landscape utilisation, climatic conditions, and evolving societal needs. A thorough exploration of these interconnected variables is essential for a comprehensive understanding of the intricate dynamics shaping the multifaceted role of cattle in medieval Norway and northern Europe.
Data availability
No datasets were generated or analysed during the current study.
Data code
Not applicable.
References
Albarella U (2002) Size matters’: how and why biometry is still important in zooarchaeology. In: Dobney K, O’Connor TP (eds) Bones and the man. Studies in honour of Don Brothwell. Oxbow, Oxford, pp 51–62
Albarella U, Davis SJM (1994) The Saxon and medieval animal bones excavated 1985-1989 from West Cotton, Northamptonshire. Research Department Reports Series 1717/94
Aniceti V, Albarella U (2022) Who’s eating pork? Investigating pig breeding and consumption in byzantine, islamic and Norman/Aragonese Sicily (7th -14th c. AD). J Archaeol Science: Rep 41:103299
Barone R (1976) Anatomie comparée des animaux domestiques: ostéologie 1. Vigot frères, Paris
Blackmore L (1994) The Bryggen pottery in the wider context. Oslo. In: Blackmore L, Vince A (eds) The Bryggen papers 5. Scandinavian University, Oslo, pp 110–111
Bourns T (2018) Meat and taboo in medieval scandinavian law and literature. Viking Medieval Scandinavia 14:61–80
Brendalsmo J (1986) Innberetning: Over de arkeologiske utgravningene 1, Baglergaten 2–4, Tønsberg, 1981–1982, Del 1 - Tekst
Challinor C (2004) Butter as an Economic Resource in the Northern Isles. In: Housley RA, Coles G (eds) Atlantic connections and adaptations: economies, environments and subsistence in Lands bordering the North Atlantic. Oxbow Books, Oxford, pp 163–174
Christie H (1966) Old Oslo. Medieval Archaeol 10:45–48. https://doi.org/10.1080/00766097.1966.11735281
Critch A, Harland J, Barrett J (2018) Tracing the late viking age and medieval butter economy: the view from Quoygrew, Orkney. In: Kershaw J, Williams G (eds) Silver, butter, Cloth: Monetary and Social Economies in the viking age. Medieval history and Archaeology. Oxford University Press, Oxford, pp 278–297
Davis SJ (1996) Measurements of a group of adult females Shetland sheep skeletons from a single flock: a baseline for zooarchaeologists. J Archaeol Sci 23:593–612
Eriksson JEG (1975) Hvistendahl – Tomten, Storgaten 35, Tønsberg. Preliminær rapport. De arkeologiske undersøkelser Tønsberg, Riksantikvaren
Fowler P (2002) Farming in the first millennium AD. British agriculture between Julius Caesar and William the Conqueror. Farming in the First Millennium AD. Cambridge University Press, Cambridge
Frimanslund R (1961) Hegn (Norge). In: Hödnebö F (ed) Kulturhistorisk leksikon for nordisk middelalder VI. Gyldendal, Oslo, pp 287–290
Fuglevik LM (in prep) An Archaeology of consuming passions. Household pottery and urban commensal strategies in Early and High Medieval Oslo, PhD Thesis. Oslo, University of Oslo
Garnier N, Vedeler M (2021) Revealing medieval culinary practices in Norway: a first metabolomic-based approach. J Archaeol Science: 40. https://doi.org/10.1016/j.jasrep.2021.103206
Grant A (1982) The use of tooth wear as a guide to the age of domestic ungulates. In: Wilson B, Grigson C, Payne S (eds) Ageing and sexing animal bones from archaeological sites. Archaeo, Oxford, pp 91–108
Grau-Sologestoa I, Albarella U (2019) The ‘long’ sixteenth century: a key period of animal husbandry change in England. Archaeol Anthropol Sci 11:2781–2803
Grayson DK (1984) Quantitative zooarchaeology. Academic, Orlando
Groot M (2005) Paleopathological evidence for draught cattle on a roman site in the Netherlands. In: Davies J, Fabiš M, Mainland I, Richards M, Thomas R (eds) Diet and health in past animal populations: current research and future directions. Oxbow, Oxford, pp 52–57
Halley DJ, Rosvold J (2014) Stable isotope analysis and variation in medieval domestic pig husbandry practices in northwest Europe: absence of evidence for a purely herbivorous diet. J Archaeol Sci 49:1–5. https://doi.org/10.1016/j.jas.2014.04.006
Hamilton J, Thomas R (2012) Pannage, pulses and pigs: isotopic and zooarchaeological evidence for changing pig management practices in later medieval England. Medieval Archaeol 56(1):234–259. https://doi.org/10.1179/0076609712Z.0000000008
Hansen G (2017) Domestic and exotic materials in early medieval Norwegian towns. An archaeological perspective on production, procurement and consumption. In: Glørstad ZT, Loftsgarden K (eds) Viking-age transformations. Trade, Craft and resources in Western Scandinavia. Routledge, London, pp 59–94
Helle K (2001) Gulatinget Og Gulatingslova. Leikanger, Skald
Hill D (2010) Medieval towns and the rural economy in eastern Norway: central place theory, settlement and taxation AD 1000 - AD 1350. VDM Verlag Dr. Müller, Saarbrücken
Holmes M (2022) Innovation, technology and social change: the adoption of the mouldboard plough and its impact on human–animal relationships. In: McKerracher M, Hamerow H (eds) New perspectives on the medieval ‘Agricultural revolution’: crop, stock and furrow. Liverpool University, Liverpool, pp 87–108
Holmes M, Thomas R, Hamerow H (2021) Identifying draught cattle in the past: lessons from large-scale analysis of archaeological datasets. Int J Paleopathol 33:258–269. https://doi.org/10.1016/j.ijpp.2021.05.004
Hufthammer AK (1999) Utgravningene i Erkebispegården i Trondheim: Kosthold Og Erverv i Erkebispegården, En osteologisk analyse. Trondheim: NIKU Temahefte 17:1–47
Hufthammer AK (2000) Kosthold hos overklassen Og Hos Vanlige husholdninger i middelalderen. En sammenligning mellom animalosteologisk materiale fra Trondheim Og Oslo. In: Dybdahl A (ed) Osteologisk materiale som historisk kilde. Centre for Medieval Studies Norwegian University of Science and Technology, Trondheim, pp 163–188
Hufthammer AK (2003) Med kjøtt Og Fisk på Menyen. In: Skevik O (ed) Middelalder-gården i Trøndelag, foredrag fra to seminar. Stiklestad Nasjonale Kultursenter, Verdal, pp 182–196
Imbens GW (2021) Statistical significance, 푝-Values, and the reporting of uncertainty. J Economic Perspect 35:3
Johannsen N (2011) Past and present strategies for draught exploitation of cattle. In: Albarella U, Trentacoste A (eds) Ethnozooarchaeology: the present and past of human-animal relationships. Oxbow Books, Oxford, pp 13–19
Langdon J, Astill G, Myrdal J (1997) Introduction. Medieval farming and technology. Brill, Leiden
Lanyon LE, Rubin CT (1985) Regulation of bone mass by mechanical strain magnitude. Calcif Tissue Int 37:411–417. https://doi.org/10.1007/BF02553711
Lie RW (1979) Osteologisk Materiale Fra Oslogate 7. In: Schia E (ed) De Arkeologiske Utgravninger i Gamlebyen, Oslo 2. Universitetsforlaget, Oslo, pp 108–123
Lie RW (1988) Animal bones. In: Schia E (ed) De Arkeologiske Utgravninger i Gamlebyen, Oslo. Mindets Tomt – Søndre Felt; animal bones, moss-, plant-, insect – and parasite remains 5. Alvheim and Eide, Bergen, pp 153–195
Lie RW (1989) Dyr i byen – en osteologisk analyse. Fortiden i Trondheim bygrunn: Folkebibliotekstomten. Meddelelser 18. Riksantikvaren, Utgravningskontoret for Trondheim, Trondheim
Lie RW (1991) Dyrebein Fra Oslogt. 4 Og Kanslergt. 10. I De Arkeologiske Utgravninger i Gamlebyen, Oslo bind 10. Øvre Ervik, Bergen, pp 75–83
Lie RT, Lie RW (1990) Changes in survival of cattle (Bos taurus) in Trondheim during the medieval period. Fauna Norvegica 11:43–49
Lie SA, Lie RW (2013) Changes in survival of cattle (Bos taurus) during medieval times in two Norwegian cities. Environ Archaeol 18:2178–2183. https://doi.org/10.1179/1461410313Z.00000000028
Lindh J (1984) Innberetning: Over de arkeologiske utgravningene 1, Storgaten 24/26, Tønsberg, 1979. Riksantikvaren: Utgravningskontoret for Tønsberg
Lyman RL (2008) Quantitative palaeozoology. Cambridge University Press, Cambridge
Maraschi A (2020) The impact of christianization on identity-marking foods in the medieval North: between pagan survivals, new dietary restrictions, and magic practice. Food History 17:2153–2181
Martens VV (2010) Arups gate, Gamlebyen, Oslo. Arkeologiske undersøkelser Og overvåking i forbindelse med utskifting av vannledning. NIKU Oppdragsrapport nr.111/2010. Norwegian Institute for Cultural Heritage Research, Oslo
Marthinussen KL (1992) Et osteologisk materiale fra Televerkstomten, Undersøkelse av et utgravd beinmateriale fra middelalderens Trondheim (Nordre gt. 1). Unpublished thesis. Bergen, University of Bergen
Martínez-García L, Cuevas A, Ferrari G, Hufthammer AK, Vedeler M, Hirons A, Star B (in prep. a) Tracing 600 years of long-distance Atlantic Cod trade in medieval and post-medieval Oslo using stable isotopes and ancient DNA
Martínez- García L, Cuevas A, Ferrari G, Jakobsen KS, Jentoft S, Blevis R, Harland J, Küchelmann HC, Gotfredsen AB, Hufthammer AK, Vedeler M, Barrett JH Star B (in prep. b) Long-term population dynamics of Atlantic Cod revealed by ancient DNA and genomics
McGovern TH, Harrison R, Smiarowski K (2014) Sorting sheep and goats in medieval Iceland and Greenland: local subsistence, climate change, or world system impacts? In: Harrison R, Maher RA (eds) Human ecodynamics in the North Atlantic: a collaborative model of humans and nature through space and time. Lexington Books, Lanham
Meadow RH (1999) The use of size index scaling techniques for research on archaeozoological collections from the Middle East. In: Becker C, Manhart H, Peters J, Schibler J (eds) Historia animalium ex ossibus. Beiträge Zur Paläoanatomie, Archäologie, Ägyptologie, Ethnologie Und Geschichte Der Tiermedizin. Festschrift für Angela Von Den Driesch. Verlag Marie Leidorf GmbH, Rahden, pp 285–300
Molaug P (2015) From the farm of Oslo to the townyard of Miklagard. In: Baug I, Larsen J, Mygland SS (eds) Nordic Middle ages - artefacts, landscapes and society. Essays in honour of Ingvild Øye on her 70th birthday. University of Bergen Archaeological Series, Bergen, pp 213–226
Molaug P, Skre D, Flodin L (2018) Oslogate 6 Arkeologiske utgravninger 1987–1989. NIKU Oppdragsrapport Nr. 4/2018. Norwegian Institute for Cultural Heritage Research, Oslo
Nedkvitne A, Norseng PG (2000) Middelalderbyen ved Bjørvika. Oslo 1000–1536. Cappelen Damm, Oslo
Nordeide SW, Gulliksen S (2007) First generation christians, second generation radiocarbondates: the Cemetry at St. Clement’s. Oslo nor Archaeol Rev 40:1–25. https://doi.org/10.1080/00293650601054362
Nordman AM, Pettersson C, Tjeldvoll GA (1986) Innberetning: over de arkeologiske utgravningene 1, Storgaten 33, Tjømegaten 1, Tønsberg, 1986. Utgravningskontoret for Tønsberg, Riksantikvaren
O’Connor T (1988) Bones from the general accident side, Tanner Row. Council for British Archaeology, London
Øye I (2004) Agricultural conditions and rural societies c. 800–1350. In: Almas R (ed) Norwegian Agricultural History, Trondheim, pp 79–140
Øye I (2009) Settlement patterns and field systems in medieval Norway. Landsc History 30(2):37–54. https://doi.org/10.1080/01433768.2009.10594607
Øye I (2013) Technology, land use and transformations in scandinavian landscapes, c. 800–1300 AD. In: Kerig T, Zimmermann A (eds) Economic archaeology: from structure to performance in European archaeology. Habelt, Bonn, pp 295–309
Payne S, Bull G (1988) Components of variation in measurements of pig bones and teeth, and use of measurements to distinguish wild from domestic remains. Archaeozoologia 2:27–66
Rizzetto M, Albarella U (2022) Livestock size and the roman-early anglo-saxon transition: Britain in North-West Europe. Archaeol Anthropol Sci 14:65. https://doi.org/10.1007/s12520-021-01494-y
Rosvold J, Halley DJ, Hufthammer AK, Minagawa M, Andersen R (2010) The rise and fall of wild boar in a northern environment: evidence from stable isotopes and subfossil finds. Holocene 20(7):1113–1121. https://doi.org/10.1177/0959683610369505
Sanmark A (2002) The Christianization of Scandinavia. A comparative study. PhD Thesis, University College of London
Sanmark A (2005) Dietary regulations in early christian Norway. Viking Medieval Scandinavia 1:203–224
Schia E (1987) Ch.C Bebyggelsesrester Og datering, Ch.D Topografi Og bebyggelsesutvikling, Ch. E Bygården. In: Schia E (ed) Søndre felt Stratigrafi, bebyggelsesrester og daterende funngrupper. De Arkeologiske Utgravninger i Gamlebyen, Oslo, vol 3. Alvheim og Eide, Bergen, pp 41–228
Schia E (1989) De Arkeologiske Utgravningene i Gamlebyen, Oslo, bind 6: hus Og Gjerde. Alvheim & Eide, Bergen
Schia E (1991) Oslo Innerst i Viken. Aschehoug, Oslo
Schmid E (1972) Atlas of animal bones. For prehistorians, archaeologists and quaternary geologists. Elsevier Publishing Company, London
Silver IA (1969) The ageing of domestic animals. In: Brothwell DR, Higgs E (eds) Science in archaeology. Thames and Hudson, London
Simpson GG, Roe A, Lewontin RC (1960) Quantitative zoology. Harcourt Brace, New York
Smedstad I (1991) Oslogate 2–8, 1987–1988. In: Wiberg T, Schia E (eds) Grøftegravinger. De Arkeologiske Utgravninger i Gamlebyen, Oslo 10. Alvheim og Eide, Bergen, pp 7–62
Stern HS (2016) A test by any other name: p values, bayes factors, and statistical inference. Multivar Behav Res 51(1):23–29. https://doi.org/10.1080/00273171.2015.1099032
Thomas R (2008) Diachronic trends in lower limb pathologies in later medieval and post-medieval cattle from Britain. In: Grupe G, McGlynn G, Peters J (eds) Limping Together Through the Ages: Joint Afflictions and Bone Infections. Documenta Archaeobiologiae 6:187–201
Undheim P (1985) Osteologisk Materiale Fra Dreggen, En økologisk studie fra middelalderens Bergen. Universitetet i Bergen
Vedeler M (2023) Middelalderens kjøkken, oppskrifter og historiene bak dem. Spartacus Forlag, Oslo
Vedeler M Bergstøl J (in prep) Maten i middelalderen: Fra naturens spiskammer til bur og bord
Vedeler M, Garnier N (2024) Fish, Wine and Fruit Juice in Medieval Soapstone Cooking. Food & History 22(1):33–66. https://doi.org/10.1484/j.food.5.137476
Vedeler M, Samdal M, Christiansen L (2023) Kartlagte matplanter fra middelalderens Oslo. Oslogate 7. Research Report. Kulturhistorisk museum. Universitetet i Oslo Arkeologisk seksjon, Oslo
von den Driesch A (1976) A guide to the measurement of animal bones from archaeological sites. Peabody Museum, Harvard
Vretemark M (1997) Från Ben till boskap: kosthåll och djurhållning med utgångspunkt i medeltida benmaterial från Skara 1. Skaraborgs länsmuseum, Skara
Walker SJ (2021) Archaeological bird remains from Norway as a means to identify as a means to identify long-term patterns in a Northern European avifauna. PhD Thesis. Bergen, University Museum of Bergen
Walker SJ, Meijer HJM (2020) More than food: evidence for different breeds and cockfighting in Gallus gallus bones from medieval and Post-medieval Norway. Quatern Int 543:125–134. https://doi.org/10.1016/j.quaint.2020.03.008
Walker SJ, Hufthammer AK, Meijer HJM (2019) Birds in Medieval Norway. Open Quaternary 5(1):1–33
Website: http://d-maps.com. Last access Nov 21 2023
White L (1962) Medieval technology & social change. Oxford University Press, Oxford
Wienberg J (1983) Rapport: over den arkeologiske udgravning 1, Kammegaten 10, Tønsberg, 1981. Utgravningskontoret for Tønsberg, Riksantikvaren
Wiig O (1981) Faunal remains from Mediaeval Bergen. Fauna Norvegica A:2:36–40
Woolgar CM (2016) The culture of food in England. Yale University Press, New Haven, pp 1200–1500
Acknowledgements
We would like to express our gratitude to everyone who contributed to the completion of this research article. We extend our sincere appreciation to the Research Council of Norway for funding the FOODIMPACT Project, of which this study is an integral part. Special thanks are due to Olaug Flatnes Bratbak for her invaluable assistance in data collection. We also wish to acknowledge Hanneke Farstad Meijer, Johnny Magnussen, Liselotte Takken-Beijersbergen, and all colleagues at the Department of Natural History-The University Museum, University of Bergen, for their valuable suggestions and support. We are grateful to Sam Walker and Mauro Rizzetto for their constructive feedback and comments. We acknowledge with gratitude the resources and facilities provided by the Department of Natural History, particularly the osteological reference collection and archive, which played an essential role in facilitating this research project. Finally, we thank the reviewers for their invaluable contributions and insightful suggestions that have significantly improved our article.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. The content of this article is a product of a Postdoctoral grant of the FOODIMPACT Project, funded by the Research Council of Norway, and awarded to Veronica Aniceti. The sponsor was not involved in the study design, data collection, analysis and interpretation, article writing, or the decision to submit it for publication.
Author information
Authors and Affiliations
Contributions
Veronica Aniceti: Conceptualisation, Methodology, Formal analysis, Data Curation, Writing - Original Draft, Review & Editing, Visualisation. Marianne Vedeler: Review & Editing, Funding acquisition. Anne Karin Hufthammer: Data curation, Review & Editing, Supervision.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 21.9 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Aniceti, V., Vedeler, M. & Hufthammer, A.K. Time for a change? Investigating shifts in agricultural economies and food in southern-central Norway (11th -16th c. AD). Archaeol Anthropol Sci 16, 144 (2024). https://doi.org/10.1007/s12520-024-02039-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-024-02039-9