
Abstract
Although many historians have extensively discussed the agricultural history of England between the Late Middle Ages and the Modern Era, this period of crucial changes has received less attention by archaeologists. In this paper, zooarchaeological evidence dated between the Late Middle Ages and the Early Modern period is analysed to investigate changes in animal husbandry during the ‘long’ sixteenth century. The size and shape of the main domestic animals (cattle, sheep, pig and chicken) is explored through biometrical data and discussed in line with evidence of taxonomic frequencies, ageing and sex ratios. Data from 12 sites with relevant chronologies and located in different areas of the country are considered. The results show that, although a remarkable size increase of animals occurred in England throughout the post-medieval period, much of this improvement occurred as early as the sixteenth century. The nature and causes of such improvement are discussed, with the aim of understanding the development of Early Modern farming and the foundations of the so-called Agricultural Revolution.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
During the sixteenth century, Europe shifted from being ‘medieval’ to being ‘modern’ because some crucial changes occurred in all aspects of life (e.g. Rice and Grafton 1994; Johnson 1996). Abundant documentary evidence has allowed historians to investigate some of these changes, such as the exploration of other continents (e.g. Penrose 1952; Cipolla 1970; Parry 1981; Arnold 2002; Love 2006), the opening of oceanic trade routes and expansion of commerce (e.g. Braudel 1992; Parker 2010), the remarkable development of politics (e.g. Allen 1928; Skinner 2007), science (e.g. Henry 2008; Burns 2015; Wooton 2015), philosophy (e.g. Allen 1928; Kraye 1996; Nauert 2006), international law (e.g. Orakhelashvili 2006; Jeffery 2006), art (e.g. Bohn and Saslow 2013) and the Protestant Reformation (Allen 1928; Bainton 1952; Appold 2011) and, in England, the dissolution of monasteries (e.g. Moorhouse 2012; Guinn-Chipman 2013).
Changes also occurred in the rural world which, arguably, affected the character of human societies even more profoundly than the political and religious upheavals. In fact, the countryside of England underwent some major transformations during the post-medieval period, with remarkable consequences in farming. These changes included the following:
-
The introduction of new crops (Crosby 2003; Fernández and González 1990; Nunn and Qian 2010; Ratcliffe 1984)
-
The introduction of new animal species such as the turkey (e.g. Yalden 1999; Reed 2008; Thomas 2010; Fothergill 2014)
-
New farming technologies and techniques such as wooden harnesses, light iron ploughs, drainage systems and four-course systems of crop rotation (e.g. Thirsk 1967; Rusell 1986; Langdon 1986; Astill and Langdon 1997; Williamson 2002)
-
The process of enclosure (e.g. Havinden 1961; Kerridge 1967)
-
A notable demographic increase associated with the growth of urban demand (e.g. Beckett 1990; Jack 1996; Dyer 2002)
For many, these are the causes of the so-called British Agricultural Revolution (e.g. Kerridge 1967; Beckett 1990; Williamson 2002), which ultimately led to the birth of modern farming. Lord Ernle’s influential work (Prothero 1912) identified such important agricultural changes with the second half of the eighteenth century, but his ideas were subsequently challenged by historians, such as Fussell (1961), Kerridge (1967) and Thirsk (1987), who suggested a much earlier onset of those changes. Regardless of its starting date and its ‘revolutionary’ nature, the British Agricultural Revolution is a concept that is widely used by historians of Modern Britain (e.g. Jones 1965; Kerridge 1967 and 1969; Beckett 1990; Overton 1984, 1996a, b; Turner et al. 1996; Allen 1999; Clark 1999), and the expression will therefore be referred to in this paper.
When it comes to investigating agricultural change, the sixteenth century has, however, been somewhat disregarded, perhaps because, traditionally, it has been overshadowed by two major events (Allen 1991) that unquestionably transformed the European countryside in both its appearance and practices: the Black Death (mid-fourteenth century) (e.g. Postan 1939; Dyer 1981; Verhulst 1990; Miller 1991; Hopcraft 1994; Dodds and Britnell 2008) and the Agricultural Revolution as originally interpreted (eighteenth century) (e.g. Prothero 1912). Most of the period in between these two events will be referred to in this work as the ‘long’ sixteenth century (spanning, broadly, from the mid-fifteenth century to the mid-seventeenth century) and will be the main focus of this paper, although some phenomena will also be considered in a longer time perspective.
Historians have paid a substantial amount of attention to the investigation of late- and post-medieval changes in agriculture and animal husbandry (e.g. Prothero 1912; Trow-Smith 1957, 1959; Fussell 1961; Thirsk 1987; Campbell and Overton 1993). Archaeologists, however, have been slower in recognising the importance of the topic, with the exception of some early work conducted by Philip Armitage (1978, 1980, 1982, 1990). In later years, the main topics that have been investigated by zooarchaeologists are represented by late medieval innovations in animal husbandry (Albarella 1997a), evidence for the onset of the British Agricultural Revolution (Davis 1997; Davis and Beckett 1999; Thomas 2005a, 2009) and urban provisioning during the post-medieval period (e.g. Gordon 2016). Some of the trends that have been identified through zooarchaeological research include the substitution of cattle with horse as the main draught animal, the increased meat (mainly veal) and milk production and the enhanced productivity related to a notable size increase of livestock (Langdon 1986; Albarella 1997a, b; Davis 1997; Davis and Beckett 1999; Thomas 2005a, 2009; Sykes 2006; Vann and Grimm 2010; Thomas et al. 2013). Zooarchaeological research concerning late- and post-medieval agricultural changes and ‘improvement’ has mainly focused on biometrical analyses. The greatest attention has been paid to cattle and sheep, the animals whose remains are the most frequent in English sites dated to this period. Other livestock species that were central to agriculture and everyday life, such as pig, chicken or horse, on the contrary, have been somewhat neglected (for turkey, however, see Reed 2008 and Fothergill 2014).
Most of the discussion has focused on when these changes occurred (e.g. O’Connor 1995; Albarella and Davis 1996; Davis 1997; Davis and Beckett 1999; Thomas 2005a, b; Thomas et al. 2013), with no clear solution to the problem, partly due to most researchers focusing only on datasets limited to specific case-studies and partly due to the fact that post-medieval bone assemblages have sparked the interest of scholars only rather recently. While all agree that changes in animal husbandry were gradual rather than revolutionary, it is still unclear if changes in England as a whole occurred as early as the Late Middle Ages or later in time. For instance, a remarkable size increase of livestock is visible at Dudley Castle during the late medieval period (Thomas 2005b), while in York, changes occurred only after the eighteenth century (O’Connor 1995). Regional diversity in post-medieval English agriculture has already been pointed out (Kerridge 1967; Thirsk 1987), with zooarchaeological evidence suggesting that changes occurred earlier and more notably in the south-eastern and central areas of the country than in the more peripheral areas (Davis 1997; Davis and Beckett 1999; Thomas et al. 2013; Thomas 2005b). The main research questions of this work concern whether improvements in livestock management can be regarded as having already been underway at the beginning of the Modern period, during the ‘long’ sixteenth century and whether they should be regarded as part of a countrywide trend or were rather punctuated and localised.
It is argued here that such innovations were concerned with increasing production and enhancing quality and therefore related to ‘improvement’ in animal husbandry. This research will mainly consider biometrical analysis, to look at size and shape changes of the main food domesticates (cattle, sheep, pig and chicken), during the period considered. Other evidence, such as taxonomic proportions, kill-off patterns and sex ratios will be utilised comparatively, to discuss the patterns observed in the biometrical analysis.
A concept that needs to be clarified in this paper is that of improvement, which has received wide attention by archaeologists and historians of the later Modern Era, particularly in relation to the Enlightenment period and the idea of progress. According to Tarlow (2007), “Improvement (in the Modern Era) was a cross-cutting ethic, directed not only at the improvement of agricultural production (…), but also at the moral, intellectual and physical improvement of the self, of the labouring people, of society, of production and of the human environment” (p. 16) and “encompasses and contextualises traditional ‘industrial archaeology’ and joins material culture with ideology” (p. 18). Therefore, investigating ‘improvement’ is key for understanding of “all aspects of material life in Britain in the period: agriculture and landscape change, houses and architecture, town morphology, artefacts and technology, social practices (etc.)” (p. 18). With these considerations in mind, it is important to clarify that ‘improvement’, although largely regarded by farmers as a way to maximise yield, indicates a different rather than necessarily better way to manage animals or the landscape. For example, in the contemporary world, industrialised intensive farming can increase meat output and profit, but is not necessarily desirable for the society, the environment, let alone the welfare of the animals.
This work provides clear evidence, based on biometrical data, for a series of ‘improvements’ in animal husbandry already during the ‘long’ sixteenth century, long before the eighteenth century. We suggest that, although many of these innovations occurred already in the Late Middle Ages, their impact was limited to some particular geographic areas; it was not until the sixteenth century that they really became widespread, making this century a turning point for English agricultural history.
Materials and methods
The sites
The faunal remains from the following archaeological sites have been re-analysed for the specific purpose of this paper: Little Pickle (Bletchingley, Surrey), Flaxengate (Lincoln, Lincolnshire) and The Shires (including Little Lane and St. Peter’s Lane) (Leicester, Leicestershire). The stratigraphic integrity of these assemblages and the chronological attribution of the archaeological contexts were assessed by the excavators and the zooarchaeologists who first analysed these materials (see details below), who regarded them as valid and representative. Only some contexts, stratigraphically sound and securely dated, as well as relevant for this paper (roughly, from the fourteenth to the seventeenth centuries), were selected for the current research.
Faunal remains from Little Pickle (Bletchingley, Surrey, 51.2406° N, 0.0992° W) are dated between 1490 and 1555 (Poulton 1998), when the site functioned as the manor country house of Hextalls. The property was bought in the early sixteenth century by the Duke of Buckingham and after his execution for treason in 1521 it was taken into royal control. The site was excavated in 1988 and 1989, and the animal bones were first studied by Bourdillon (1998). The faunal remains were loaned to us by the Guildford Museum.
Two of the sites analysed by us (Little Lane and St. Peter’s Lane) are both located in The Shires, an area in the city centre of Leicester (52.6369° N, 1.1398° W), known nowadays to the locals as the Highcross. Both sites were excavated in 1988 and 1989 (Lucas and Buckley 2007). The sites provided faunal remains dated to a wide chronology, spanning from Roman to contemporary times, but only the late medieval and early post-medieval contexts (fourteenth–seventeenth centuries), first studied by Gidney (1991a, b, c, 1992, 1993), were re-analysed for this work. The faunal remains were made available to us by the Leicester Museum.
The last set of faunal remains that has been re-examined specifically for this paper comes from Flaxengate, in the city centre of Lincoln (53.2307° N, 0.5406° W). The site provided evidence spanning from Roman to contemporary times (Jones 1980), but only the late medieval and early post-medieval faunal remains (late fourteenth–mid sixteenth centuries) were considered for this paper. The animal bones were first analysed by O’Connor (1982). The faunal remains were loaned to our lab by The Collection Museum in Lincoln.
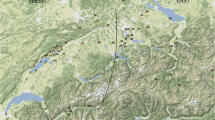
In addition to these primary data (raw data are available in the supplementary online resources, file ‘Online Resource 2’), further evidence was collected for other English assemblages (Fig. 1). These include Dudley Castle (Thomas 2005b), Exeter (Maltby 1979), Launceston Castle (Albarella and Davis 1996), various sites from London (Thomas et al. 2013), Norton Priory (Wright et al. 2016), Norwich Castle (Albarella et al. 2009), Okehampton Castle (Maltby 1982), Saint Giles by Brompton Bridge (Stallibrass 1993) and West Cotton (Albarella and Davis 1994, 2010). This large body of data has allowed us to undertake a full comparative analysis aimed at identifying the timing and nature of animal husbandry changes. These sites cover most of England (Fig. 1, Table 1), from north to south, and from east to west, although the analysis is inevitably punctuated, with large sections of West Midlands and the South West not represented. Some of the sites are urban (e.g. London, Exeter and The Shires) or castles (e.g. Dudley and Launceston), but faunal assemblages from rural and monastic sites dated to the Early Modern period are rare, making a full comparison between different types of sites unfeasible on the basis of the current state of knowledge.

Map showing the location of the sites
The different ‘period-sites’ (i.e. assemblages from the same site but different periods) have been combined into three main periods (Late Middle Ages, c. sixteenth century and seventeenth–nineteenth centuries). The period-sites considered for each general chronological period are shown in Table 2.
The log-ratio technique and standards used
The log-ratio technique (Simpson 1941) has been used for the present biometrical analysis. This technique increases the sample size and allows direct comparison between measurements (Meadow 1999; Albarella 2002). As a size index scaling technique, the log-ratio relates our measurements to a standard individual or the mean of a given population (Payne and Bull 1988), calculating the decimal logarithm of the ratio between the measurement and its standard. For the comparative analysis between different period-sites, only those that provided a sample size equal to or larger than 30 measurements were used, thus discarding period-sites with very small samples, following a common practice in statistical literature (Hogg et al. 2015).
The focus of the study is on cattle, sheep, pig and chicken. It would have been valuable to include horse too, but the relevant datasets tended to be too small. Goat was excluded from the analysis when it was identified; however, some measurements from specimens identified as sheep/goat have been included due to the difficult distinction of the two species in certain elements. Nonetheless, goat is rare in British medieval and post-medieval sites (Albarella 1997a, 2003), and therefore, the inclusion of a few goat measurements to a sample largely predominated by sheep does not affect the observed patterns in any significant way.
Only a selection of all possible measurements was considered for the analysis based on the log-ratio technique. The parts of the skeleton that were chosen are especially common in archaeological assemblages as they tend to preserve well. Also, the chosen measurements can be easily defined and taken and are well suited for comparison across different researchers. Finally, the measurements were chosen because of their relatively low sexual dimorphism and because they are not amongst the most age dependent (Popkin et al. 2012). In this work, the priority was to look at morphometric differences between livestock types, rather than the identification of age and sex groups, though these must be taken into account in the interpretation, as changes in sex ratios can generate changes in size (Hadjikoumis 2010).
There is better correlation between measurements taken along the same axis than between those on different axes (Davis 1996) and, ideally, only measurements taken in the same plane should be combined, making sure that only one measurement per bone is considered. Lengths, widths and depths have therefore been plotted separately. Fused and fusing, but not unfused, bones have been included in the analysis. The inclusion of fusing bones may result in a potential bias in the size patterns, as some fusing bones may still be in the process of growth. However, their number was small and therefore they are unlikely to have produced any substantial effect on the observed patterns. The list of anatomical elements and measurements used for this analysis, as well as some further information on them, is provided as part of the supplementary online resources (‘Online Resource 1’).
The set of standard measurements used for cattle, sheep and chicken have been calculated from our own data from The Shires, following well-known statistical methods (see ‘Online Resource 1’ for details). These standards have been calculated from the combination of the contexts dated between the fifteenth and mid-sixteenth centuries (both Little Lane and St. Peter’s Lane). As part of the supplementary online resources (‘Online Resource 1’), the standards calculated for The Shires are provided (Tables S2 to S4), and the measurements themselves can be found in the supplementary online resources (‘Online Resource 2’). For the biometrical analysis of pig postcranial bone and tooth measurements, the standards are those derived from Late Neolithic Durrington Walls (Albarella and Payne 2005).
In total, almost 17,600 measurements were collected. Nearly 5200 of those are from cattle, 7400 from sheep, 1500 from pig and 3500 from chicken. Of these, approximately 6400 are dated to the Late Middle Ages, 7300 to the c. sixteenth century and 3900 to the seventeenth–nineteenth centuries.
Results
In the following sections, the results of the biometrical analyses for the four main livestock species used as food (cattle, sheep, pig and chicken) are shown. In each case, box plots comparing the log-ratios of all period-sites are provided.
Cattle
In Figs. 2, 3 and 4, the log-ratios for cattle postcranial bones (widths, lengths and depths, respectively) are shown for each period-site, with the earliest ones plotted at the bottom of the graphs and the latest at the top. A progressive increase in size is noticeable for all three axes. Depth measurements (Fig. 4) seem to show greater variability within each of the three main chronological groups, as exemplified by the notable size difference between cattle from London 17th-19th c., Launceston 17th-19th c. and Exeter 18th c.

Box-plot showing the log-ratios of cattle postcranial widths, for each period-site

Box-plot showing the log-ratios of cattle postcranial lengths, for each period-site

Box-plot showing the log-ratios of cattle postcranial depths, for each period-site
A histogram of the axis with most measurements (widths) is shown in Fig. 5, taking into consideration only those period-sites (with sample size larger than 30) that provided measurements only dated to the sixteenth century, or to the centuries just before or after, with no chronological overlap. The graph shows a remarkable size increase in the Early Modern period at The Shires and the very large size of the sixteenth century Little Pickle cattle.

Histogram with cattle width measurements
When all period-sites are combined into three general chronological periods, the sixteenth century size increase of cattle is already very clear, though further increase occurs later (Fig. 6). Although measurements along the three axes all increased in size through time, the change is more marked in depths and, particularly, widths. Post-medieval cattle were, on average, more robust than late medieval ones.

Mean values of the three postcranial axes for cattle
In the online resources (‘Online Resource 1’), statistical analyses of the biometrical data of cattle postcranial widths (S5), lengths (S6) and depths (S7) are shown. The results of the pairwise comparison (using a Mann-Whitney U test), summarised in Table 3, show very highly statistically significant differences between the three periods, suggesting a size increase of cattle in the three skeletal axes through time, and already in the period around the sixteenth century.
Sheep
The log-ratios of sheep postcranial bones comparing different period-sites are shown in Figs. 7, 8 and 9 (widths, lengths and depths, respectively). The three graphs show an overall increase in size through time, for the three axes. An interesting pattern appears when looking at the range of the width values: it seems that width values progressively become more variable, perhaps suggesting that, in later times, sheep breeds of very different robustness occurred in England.

Box-plot showing the log-ratios of sheep postcranial widths, for each period-site

Box-plot showing the log-ratios of sheep postcranial lengths, for each period-site

Box-plot showing the log-ratios of sheep postcranial depths, for each period-site
In Fig. 10, a histogram of the axis with most measurements (widths) is shown, taking into consideration only those period-sites (with sample size larger than 30) that provided measurements solely dated to the sixteenth century, or to the centuries just before or after, with no chronological overlap. Similarly to cattle, the graph shows a substantial size increase at The Shires and that sixteenth century Little Pickle and Exeter had very large sheep.

Histogram with sheep width measurements
When grouped into three chronological periods, the pattern seems clear: the evidence suggests that sheep increased in size remarkably in post-medieval times and already in the sixteenth century. Figure 11 summarises the evolution of the means for the three axes through time. This graph suggests a very notable size increase of sheep in all axes, although lengths did not change as much as widths and depths in the post-sixteenth century. As for cattle, post-medieval sheep were, on average, more robust than late medieval ones.

Mean values of the three postcranial axes for sheep
In the supplementary resources (‘Online Resource 1’), the statistics analysing the biometrical data of sheep postcranial widths (S8), lengths (S9) and depths (S10) are shown. The results of the pairwise comparison (using a Mann-Whitney U test), summarised in Table 4, show very highly statistically significant differences between the three periods, suggesting a size increase of sheep in the three skeletal axes through time, and already in the c. sixteenth century.
Pig
Many pig bones recovered from late and post-medieval English sites are not fused, and thus, their measurements cannot be used for this analysis. For this reason, pig postcranial measurements represent the smallest sample size of the four taxa analysed in this paper, and therefore, the results must be taken cautiously. However, unlike for the other species, a relatively large number of pig tooth measurements could be taken, which provides the opportunity to investigate whether observed size changes are related to genetic or phenotypic factors. Teeth are less plastic than bones and therefore less affected by environmental factors (such as nutrition and breeding conditions), as well as age and sex variation (Degerbøl 1963; Payne and Bull 1988; Albarella 2002); thus, change in tooth size is more likely to indicate genotypic variation, which could be caused by the introduction of new animals or the evolution of local ones (Albarella et al. 2007).
The log-ratios of pig postcranial bones by period-sites are shown in Figs 12 and 13 (widths and lengths, respectively). The sample size for many of the period-sites is limited, especially for width measurements, so a clear pattern is not visible. The graph showing the log-ratios of pig postcranial lengths, however, shows a clear progressive size increase through time. Depth measurements have not been taken into account because, aside from their small number, no depth postcranial standard was available from the Durrington Walls dataset (Albarella and Payne 2005).

Box-plot showing the log-ratios of pig postcranial widths, for each period-site

Box-plot showing the log-ratios of pig postcranial lengths, for each period-site
The pattern for pig teeth is different from that suggested by postcranial bones. Figs. 14 and 15 show the log-ratios for pig teeth (lengths and widths, respectively), for each period-site. Both figures suggest that those period-sites that are shown in the middle of the graph, in general dated to the sixteenth century, show larger values than the late medieval and later post-medieval pig teeth. Pig teeth increased in size in the sixteenth century, but decreased in later centuries.

Box-plot showing the log-ratios of pig tooth lengths, for each period-site

Box-plot showing the log-ratios of pig tooth widths, for each period-site
These observations are supported by further analysis grouping all period-sites into three broad chronological periods. Figure 16 summarises the evolution of the means for the four types of measurements (postcranial widths and lengths, and teeth widths and lengths). Between the Late Middle Ages and the sixteenth century, all four types of measurements increased in size, suggesting that pig improvement involved some genetic change, perhaps in combination with environmental factors. The most marked increase in this period is nonetheless visible on postcranial lengths, making pigs taller in the sixteenth century than they were in late medieval times. On the other hand, when comparing the values of the sixteenth century with those of the post-sixteenth century, an interesting pattern emerges: the postcranial values did increase, while tooth values decreased in size. The increase in bone size could be explained with better feeding and more efficient managing practices, but the decrease in tooth size is likely to be genetic. It reflects a trend observed in modern pig breeds, namely the shortening of the snout, a characteristic that can be used to assess primitiveness in pigs (Albarella et al. 2006). Large, short-snouted pigs (relatively similar in appearance, for instance, to the modern Berkshire pig, the Oxford Sandy & Black Pig or the British Saddleback pig) were likely to have been imported into Britain after the sixteenth century.

Mean values of the two postcranial axes and the two tooth axes for pig
In the supplementary resources (‘Online Resource 1’), the statistics analysing biometrical data of pig postcranial widths (S11) and lengths (S12), as well as tooth lengths (S13) and widths (S14) are shown. The results of the statistical analyses are summarised in Table 5. As explained in the supplementary online resources, for the comparison between the late medieval and the sixteenth century samples, a Mann-Whitney U test was used, due to the non-normal distribution of the data. On the other hand, for comparing the postcranial data for the pair sixteenth c.–post-sixteenth c., a parametric test (a Student’s independent-samples t test) was used, as the data show a normal distribution. In contrast, for teeth of this chronology a non-parametric test (Mann-Whitney U test) was carried out, as the data are not normally distributed. The results show that statistically there are very highly or highly significant differences between the first two periods in all axes, confirming a pig size increase already in the c. sixteenth century. Between the c. sixteenth century and the post-sixteenth century, there is no statistical significance in the size of postcranial bones, but this may be due to the small sample size in the later period. Conversely, the reduction in tooth size is validated by statistical analysis.
Chicken
The log-ratios of chicken postcranial bones, arranged by period-sites, are shown in Figs. 17, 18 and 19 (widths, lengths and depths, respectively). The three graphs show a progressive increase in the size of chicken through time, and already in the sixteenth century. Size change in later post-medieval width and depth measurements, however, appears to be less clear, perhaps in relation to the introduction of a diversity of chicken breeds (Aldrovandi 1598), which may have confused the pattern. The lack of a distinct trend may, however, also be due to the smaller sample size of the more modern period-sites, as shown at the top of the graphs.

Box-plot showing the log-ratios of chicken postcranial widths, for each period-site

Box-plot showing the log-ratios of chicken postcranial lengths, for each period-site

Box-plot showing the log-ratios of chicken postcranial depths, for each period-site
When comparing the evolution of the mean of the three axes for the chicken postcranial values divided into three broad chronological periods (Fig. 20), all of them indicate increase through time but, more remarkably, between the Late Middle Ages and the sixteenth century. For the post-sixteenth centuries, the most marked increase is in length values. This evidence suggests that in the sixteenth century, domestic fowl became taller and more robust, while for the later period, increase was largely limited to height.

Mean values of the three postcranial axes for chicken
In the supplementary online resources (‘Online Resource 1’), the statistics analysing the biometrical data of chicken postcranial widths (S15), lengths (S16) and depths (S17) are shown. The results of the pairwise comparison (with a Mann-Whitney U test), summarised in Table 6, support the pattern mentioned above. There are very highly statistically significant differences for the three postcranial axes between the Late Middle Ages and the sixteenth century and for length measurements in the period after the sixteenth century. On the other hand, no statistically significant differences can be demonstrated for widths and lengths between the sixteenth century and the following centuries.
Discussion
The results of the analysis carried out in this paper show that a clear size increase occurred in all four livestock species considered (cattle, sheep, pig and chicken) between the Late Middle Ages and the post-medieval period in England. More importantly, this work suggests that a very significant part of this size increase happened already during the ‘long’ sixteenth century, at the very beginning of the Modern Era. Traditionally, research on agricultural change in this period has been overshadowed by the remarkable developments in farming that happened in later post-medieval times. Moreover, many scholars of the Early Modern period have paid greater attention to other social, economic and political developments (e.g. international trade and colonialism), rather than investigating changes in the rural world. Plainly, not all regions experienced the same degree of change, but there seems to be little doubt that the sixteenth century was a dynamic period of experimentation in husbandry practices.
Much research so far has focused on the timing of livestock size increase, but the evidence has mainly been based on individual case-studies. Research has mainly dealt with cattle and sheep, suggesting that size increase happened earlier in the south-eastern and central areas of England. This is corroborated by our results, showing, for example, the large size of late medieval cattle from centrally located Dudley Castle. Earlier research also pointed out a later improvement for sheep than cattle. Relying on historical evidence, Overton (1996b) has proposed that cattle size increased between the Middle Ages and the sixteenth century, rather than later. Davis and Beckett (1999), combining documentary and archaeological evidence, have also suggested that size increase in cattle and sheep was under way by the sixteenth century, although O’Connor (1995) suggested that the size increase happened much later, in the late eighteenth–nineteenth centuries. This paper provides—for the first time—a large-scale, regional analysis of this question, which confirms and emphasises the suggestion that a substantial size increase occurred in the sixteenth century. This affected cattle and sheep, but also pig and chicken, and continued in later centuries. It is also worth noticing that all domesticates in the sixteenth century manor house of Little Pickle are much larger than at other contemporary sites. This may be a consequence of the more intensive farming practices that operated at large rural properties.
Our research has shown a clear size increase through time in bones aligned along all three axes of the body. Why did this size increase occur? The size (and shape) of animals is determined by both genotypic and phenotypic factors. A phenotypic change is due to environmental factors, as well as age and sex variation, and can be reflected in the size and shape of postcranial bones. However, we suggest that the size increase that we have documented is unlikely to be explained with a change in the age and sex composition of the faunal assemblages included in our analysis, as discussed below.
Sex variation can affect size: if one compares two samples and if one sample has more males and the other more females, then the average size of the former is likely to be larger than the latter as, in most mammals and many birds, males are larger than females. All animals discussed in this paper are sexually dimorphic, with males on average larger than females, and therefore, size needs to be discussed as a potential reason for the observed size change. Sexing evidence was considered for the largest of the assemblages that we re-analysed: data from The Shires did not show any substantial change in sex ratios through time for pigs (‘Online Resource 1’, S18) and chicken (S19 and S20). Sample sizes for sex data were, however, small and the evidence must be considered cautiously.
For pigs and chicken, if the size increase were due to sex, an increase of the proportions of males should be expected, but this is unlikely from a management point of view, as for both species a higher proportion of females (to ensure enhanced reproduction rates) is more profitable. Furthermore, this is supported by the ageing evidence (see below), which suggests a decrease in the age-at-death of both species, due to increased emphasis on meat production. In this context, the slaughtering of males in young age, which therefore would not feature in the biometric analysis, is expected.
For cattle and sheep, if the size increase were due to sex, we should expect an increase in the proportions of males or castrates. The first is highly improbable from a management point of view; both species are more profitable as castrates or females if they are raised to an advanced age. Concerning cattle, a sex-related size increase is made unlikely by the age pattern. Since oxen are used for traction, such change should be accompanied by an increase in the overall age. However, as we will see below, the opposite is the case. For sheep, on the other hand, the size change could perhaps be related to an increase in the proportions of wethers, linked to the increasing importance of wool production, as castration is known to improve the quality of the fleece (Kiley 1976). However, if we consider the fact that castrates tend to have longer bones than females and entire males (Silberberg and Silberberg 1971; Davis 2000), a hypothetical increase in ox and whether numbers should result in a size increase that is more pronounced in lengths than widths and depths. As we have seen before, the evidence, however, shows otherwise. Therefore, there is overwhelming indication that the size increase of cattle and sheep is not due to changes in the sex ratios.
Our biometrical analysis carefully selected those measurements that are less age dependent. Although unfused bones were excluded from the analysis, age can still be an influence on size as some bones can be subjected to post-fusion increase, particularly in width and depth (Payne and Bull 1988). Chicken measurements were all taken from fully formed individuals, with dense and not spongy bones; age, therefore, should not represent a substantial factor in their size variation. If age was the determinant factor for the size increase in mammals, we should expect to see an increase of the proportion of animals slaughtered at older stages. Ageing evidence for cattle, sheep and pig from The Shires is shown in the supplementary online resources (‘Online Resource 1’, S21 to S23). This evidence suggests that, at this particular site, the age-at-death of both cattle and pig decreased through time, and only sheep became slightly older during the sixteenth century and younger in the post-sixteenth century. At other case-studies considered in our regional analysis, such as Launceston Castle (Albarella and Davis 1996), age was ruled out as a potential factor to explain size change. Indeed, several regional reviews have suggested that the age of most livestock decreased in post-medieval times (this is the case for cattle, pig and chicken in central England—Albarella et al. forthcoming—cattle in the south—Holmes forthcoming—and sheep in the north—Huntley and Stallibrass 1995), with the only exception of sheep in southern and central England, where they became slightly older (Holmes forthcoming and Albarella et al. forthcoming, respectively). The overall pattern, therefore, indicates that age represents a very unlikely factor to explain livestock size increase.
Environmental factors may have played a role in the observed pattern. The introduction of new or different foodstuff, such as forage crops (Davis and Beckett 1999: 3), carrots, introduced to England at the end of the sixteenth century (Kerridge 1967), and turnips, also in the sixteenth century (Beckett 1990), as well as the use of stalling or sty farming, which allowed closer control over feeding and breeding (Thomas 2005a: 83–84), could have all contributed to improve the diet of the livestock and, consequently, its size.
A genotypic change would happen through a genetic alteration, either by introducing foreign animals or through selection of the local ones. In zooarchaeology, genetic modifications can be identified by looking at changes in the size and shape of teeth, since these are less plastic and less affected by environmental factors than postcranial bones (Payne and Bull 1988; Albarella 2002). A change in the size and shape of teeth will mean that a new type of livestock was introduced or that selective breeding of local types occurred. Too few tooth measurements were unfortunately available for cattle and sheep, but the analysis of pig teeth has demonstrated that genetic factors contributed to the size increase of pig in the sixteenth century. Further genetic modifications were introduced in later centuries. These caused the shortening of the snout (as proven by the reduction in tooth lengths), a characteristic that can be found in modern (‘non-primitive’) breeds. Conversely, body size increased further and affected all bone dimensions.
The abundant documentary evidence available for this period allows us to compare our results with such complementary sources. As communication and trade improved (Parker 2010), livestock trade increased, both within Britain and with foreign countries. This “led to cross-breeding and hence improvement through ‘hybrid vigour’” (Davis 1997: 416), which must have been accompanied by greater breed diversification. There is documentary evidence suggesting, for instance, that large ‘long-legged’ Dutch cattle were introduced to England in the sixteenth century (Trow-Smith 1957: 203) and that new large pig breeds were imported in the eighteenth century (Epstein and Bichard 1984). It has been suggested that cattle from post-medieval Launceston were perhaps the result of such introduction from the Netherlands (Albarella 1997b: 45–46).
As a consequence of a substantial demographic increase in the sixteenth century (Wrigley and Schofield 1981), and the subsequent increase of the mainly urban-driven demand for meat (Albarella 1997a: 28) and other animal products, such as wool (Albarella 1999), there was a shift towards a more intensive type of farming and animal husbandry. By the sixteenth century, wool was of great importance to England’s economy, with cloth representing four-fifths of the country’s exports (Hoskins 1955). Labour productivity and agricultural efficiency rose in Early Modern England (Allen 2003), as a result of new farming practices, such as grass-arable rotation and the increased use of manure, soap ashes, marl and lime for improving the soil (Kerridge 1967), together with the growth of stock density (Davis and Beckett 1999). The size increase of domestic animals therefore constitutes only one element of a larger system of support to the improvement, intensification and increased sophistication of animal husbandry.
In which sense is animal size increase related to improvement? Zooarchaeological research dealing with biometrical data generally assumes that the size increase of animals mostly responds to the will of farmers to enhance agricultural production and productivity and, therefore, represents an agricultural innovation (as defined by Van der Veen 2010: 2). When dealing with the late- and post-medieval periods, zooarchaeologists label size increase in livestock as an improvement. The understanding of ‘improvement’ in agriculture has been “a central element of historical studies of the feudal/capitalist transition” (Thomas 2005a: 71). It can, however, be argued that interpreting livestock size increase as an improvement is just a product of a contemporary point of view, which sees enhanced productivity exclusively in positive terms. An animal’s larger body size will ensure a greater meat output when slaughtered and, while still alive, will provide greater traction power. Also, improved animal breeds tend to grow faster than more primitive breeds (Albarella 1997a). This means that they can be slaughtered earlier, thus generating the desired meat output in a shorter time. There are, however, many situations in which smaller animals could be preferable—for instance, in transhumant or otherwise highly mobile communities, or when adaptation to local, even harsh, environmental conditions is more important than the maximisation of yield.
However, the important point to make is not whether animal size increase in Early Modern England was an improvement or not; more importantly, it was perceived as an improvement by Early Modern English farmers. In contrast to medieval subsistence farming, modern farmers sought profit from their livestock and were therefore particularly motivated to enhance both quantity and quality. All early examples of British writers who gave advice on animal husbandry (e.g. Fitzherbert 1523a/34 and 1523b/39; Weston 1645; Blith 1649, 1653; Worlidge 1668) advocated the selection of larger animals. It could be argued that written sources tend to emphasise theory rather than actual practice and to idealise the role of noble men in achieving agricultural innovations, as exemplified by R. Bakewell, seen as one of the great English stockbreeders of the eighteenth century. However, farmers’ knowledge based on observation and experience should not be underestimated (Grau-Sologestoa 2015: 128–129). Before genetics were scientifically understood, “crossing of animals, in an attempt to improve the quality of output, had taken place for centuries, as farmers sought -pragmatically- to improve the weight and quality of the beasts they took to the market” (Davis and Beckett 1999: 2).
Although the evidence for size increase presented in this work is meaningful by itself, other zooarchaeological evidence, such as taxonomic proportions and kill-off patterns, confirm that important changes occurred in animal husbandry between the Late Middle Ages and the post-medieval period in England. Two forthcoming studies provide an overview of the zooarchaeological evidence of the transition between these two periods in central and southern England (Albarella et al. forthcoming; Holmes forthcoming, respectively). A separate review was previously carried out for the northern part of the country (Huntley and Stallibrass 1995). The zooarchaeological evidence shows that post-medieval changes in animal husbandry and consumption patterns affected the various English regions in different ways. This is consistent with the information we gather from documentary sources (e.g. Kerridge 1967; Thirsk 1987).
Holmes (forthcoming: 142–143) suggests that there was an increase of arable land in southern England, between the Late Middle Ages and the Modern period. Sheep frequencies increased due to the enhanced value of manure and wool, whereas cattle became less common, as large herds of this species became unsustainable. Horse remains continued to be uncommon in the post-medieval period, but Holmes records a noteworthy increase of horse proportions in high-status sites (p. 158). Chicken was the most frequent bird in both periods, but its frequency decreases substantially in post-medieval times. Based on the ageing evidence, she also inferred (p. 185) an enhancement of cattle meat production in the post-medieval period, while for sheep there was evidence suggesting an increasing importance of secondary products.
The patterns observed in central England appear to be rather different (Albarella et al. forthcoming). No substantial differences in livestock frequencies were found between the two periods. Regional differences were, however, observed, as cattle were more common than sheep in western and eastern central England, in continuity with the medieval period. The frequencies of horse and chicken remains increased in the post-medieval period. The slaughtering of calves (for veal and dairy production) became more common in the latest medieval and the post-medieval period, while sheep got older, for wool production. There is an overall decrease of the slaughtering age for pigs, due to faster growing breeds (Albarella 1997a; Albarella et al. 2006), and chicken, probably connected to a greater emphasis on rapid meat returns.
There is less available evidence for Northern England (Huntley and Stallibrass 1995), but the authors suggested two general patterns: first, sheep was the most frequent domesticate in both periods, and secondly, a higher proportion of young sheep, raised for meat, is attested for the post-medieval period.
Regional variability aside, the emerging picture is that of a country that, in general, was progressively more and more dominated by sheep husbandry. The written sources suggest that horse substituted cattle as the main traction animal (Langdon 1986; Overton and Campbell 1992), though, this did not happen in all regions (Trow-Smith 1959: 173–177). The zooarchaeological evidence shows that horse is uncommon in all available sites; however, the relative frequency of this species needs to be considered carefully (discussed in Albarella 1997a). With horse not being a food animal, it can be suggested that disposal practices for this animal were different from those used for other livestock species considered here. Perhaps, what seems more interesting, is that the reduction in cattle frequencies, accompanied by some decrease of the age-at-death of cattle, may be related to a gradual switch of traction duties from oxen to horses, helped by the introduction of lighter iron ploughs (Trow-Smith 1959) and wooden harnesses (Lefebvre des Noëttes 1931). Horse carcasses could have been disposed of at special dumping sites, rather than in the middens, pits and other contexts where most faunal remains are found. Also, it is possible that most elderly horses would arrive at rural sites to the end of their lives, where they were used for agricultural purposes. Further excavations and zooarchaeological analyses are needed at rural sites to elucidate this, as post-medieval excavations have been carried out mainly at urban settlements.
Another important trend is that, in all regions, the kill-off patterns show—for the late fifteenth to the sixteenth centuries—an increase in the proportions of domesticates being slaughtered young. This suggests an increase in meat production, mainly represented by veal, pork and chicken, and mutton also in northern England. The reduction in cattle frequencies and age-at-death may also reflect the growing importance of dairy products and specialisation in the post-medieval period (Thirsk 2007) resulting in higher amounts of bones from veal which may have lower bone preservation/survival rates. The important change in consumption patterns, mainly in towns, reflected by the increasing proportion of domesticates raised for producing meat, could have been one of the main reasons why farmers aimed to breed larger animals. Finally, if animals were getting larger but at the same time they were culled at an earlier age, they probably belonged to fast growing breeds (Albarella 1997a; Albarella et al. 2006). By reaching full weight at a younger age, they would have been more profitable for farmers. This trend has continued in the contemporary world, in response to the rapid explosion of global demand for (cheap) meat (Meat Atlas 2014: 48–49).
Conclusions
Although zooarchaeological evidence for livestock improvements in post-medieval England has been investigated before, this has mainly relied on individual case studies and the subject—in view of its importance for our understanding of the shaping of the modern world—has been in dire need of a re-analysis. In this paper, for the first time, the evidence from all main food animals (cattle, sheep, pig and chicken) has been considered at a countrywide scale and using a large number of case-studies, as well as relying on first-hand re-analysis of a substantial amount of evidence (for London see Thomas et al. 2013).
Our work incontrovertibly demonstrates that, in England, livestock improvements were well on their way in the sixteenth century, well before the Agricultural Revolution as historically defined. This is indicated by clear body size increases in all analysed animals as well as changes in mortality patterns. Obviously changes did not affect all regions equally or at the same pace, but enough areas were involved to consider this to have been a large-scale phenomenon.
Having identified the sixteenth century as an important period of change, we should not underestimate the innovations introduced in animal husbandry in the Late Middle Ages, which were many and important. It has been suggested that some mechanisms of livestock improvement were already in place during the late medieval period (see Thomas 2005b, mainly relying on the important example of Dudley Castle). Although there is certainly some truth in this, the medieval evidence for improvement is fairly limited. Some of the innovations that we have attested for the sixteenth century may have had their roots in the Middle Ages, but it seems clear that the rate and scale of change substantially increased.
Our research suggests that the sixteenth century was a key turning point for innovation and improvement in animal husbandry, which led to the birth of modern farming. The idea that this was a period of relative stagnation in agriculture, sandwiched between the Black Death and the Agricultural Revolution, can be discounted. The ‘long’ sixteenth century was the time when those agricultural innovations that had begun to emerge in England during the Late Middle Ages became widespread, causing substantial and long-lasting transformations in the rural world. This ultimately led to the ‘modernisation’ of English agriculture.
References
Albarella, U (1997a) Size, power, wool and veal: zooarchaeological evidence for late medieval innovations. In: De Bow G, Verhaeghe F (eds) Environment and subsistence in medieval Europe—papers of the ‘Medieval Europe Brugge 1997’ Conference, Volume 9. Instituut voor het Archeologisch Patrimonium, p 19–30
Albarella U (1997b) Shape variation of cattle metapodials: age, sex or breed? Some examples from mediaeval and postmediaeval sites. Anthropozoologica 25/26:37–47
Albarella U (1999) ‘The mystery of husbandry’: medieval animals and the problem of integrating historical and archaeological evidence. Antiquity 73:867–975
Albarella U (2002) Size matters: how and why biometry is still important in zooarchaeology. In: Dobney K, O’Connor T (eds) Bones and the man. Studies in honour of Don Brothwell. Oxbow Books, Oxford, pp 51–62
Albarella U (2003) Tanners, tawyers, horn working and the mystery of the missing goat. In: Murphy P, Wiltshire P (eds) The environmental archaeology of industry. Oxbow Books, Oxford, pp 71–86
Albarella U, Beech M, Curl J, Locker A, Moreno-García M, Mulville J (2009) Norwich Castle: excavations and historical surveys 1987–98. Part III: a zooarchaeological study. East Anglian Archaeology Occasional Papers 22, Norwich
Albarella U, Davis SJM (1994) The Saxon and medieval bones excavated 1985-1989 from West Cotton, Northamptonshire. English Heritage, London AML Report 17/94
Albarella U, Davis SJM (1996) The mammals and birds from Launceston Castle, Cornwall: decline in status and the rise of agriculture. Circaea 12(1):1–156
Albarella U, Davis SJM (2010) The animal bones. In: Chapman A (ed) West Cotton, Raunds: a study of medieval settlement dynamics AD 450-1450. Oxbow Books, Oxford, pp 516–537
Albarella U, Dobney K, Rowley-Conwy P (2006) The domestication of the pig (Sus scrofa): new challenges and approaches. In: Zeder MA, Bradley DG, Emshwiller E, Smith DD (eds) Documenting domestication. New genetic and archaeological paradigms. University of California Press, Berkeley, pp 209–227
Albarella U, Johnstone C, Vickers K (2007) The development of animal husbandry from the Late Iron Age to the end of the Roman period: a case study from South-East Britain. J Archaeol Sci 20:1–21
Albarella U, Payne S (2005) Neolithic pigs from Durrington Walls, Wiltshire, England: a biometrical database. J Archaeol Sci 32:589–599
Albarella U, Pirnie T, Viner S (forthcoming) Animals of our past: zooarchaeological evidence from Central England. Historic England
Aldrovandi, U (translated by LR Lind) (1598) Aldrovandi on chickens: the ornithology of Ulisse Aldrovandi (1600). Volume II. Book XIV. University of Oklahoma Press, Norman
Allen JW (1928) A history of political thought in the sixteenth century. Routledge, London
Allen RC (1991) The two English agricultural revolutions, 1450-1850. In: Campbell B, Overton M (eds) Land, labour and livestock: historical studies in European agricultural productivity. Manchester University Press, Manchester, pp 236–254
Allen RC (1999) Tracking the Agricultural Revolution in England. Econ Hist Rev 52:209–235
Allen RC (2003) Progress and poverty in Early Modern Europe. Econ Hist Rev LVI(3):403–443
Appold KG (2011) The reformation: a brief history. Wiley-Blackwell, Malden
Armitage P (1978) Hertfordshire cattle and London meat markets in the 17th and 18th centuries. London Archaeologist 3(8):217–223
Armitage P (1980) A preliminary description of British cattle from the late twelfth to the early sixteenth century. Ark 7(12):405–413
Armitage P (1982) A system for ageing and sexing the horn cores of cattle from British post-medieval sites (17th to early 18th century) with special reference to unimproved British longhorn cattle. In: Wilson B, Grigson C, Payne S (eds) Ageing and sexing animal bones from archaeological sites. BAR, British Series, Oxford, pp 37–54
Armitage P (1990) Post-medieval cattle horn cores from the Greyfriars site, Chichester, West Sussex, England. Circaea 7(2):81–90
Arnold D (2002) The age of discovery, 1400–1600. Routledge, London
Astill G, Langdon J (1997) Medieval technology and farming: the impact of agricultural change in Northwest Europe. Brill, Leiden
Bainton R (1952) The reformation of the sixteenth century. The Beacon Press, Boston
Beckett JV (1990) The Agricultural Revolution. Blackwell, Oxford
Blith W (1649) The English improver, or, a new survey of husbandry
Blith W (1653) The English improver improved, or, the survey of husbandry surveyed
Bohn B, Saslow JM (2013) A companion to Renaissance and Baroque art. Wiley-Blackwell, Malden
Bourdillon, J (1998) The mammal and bird bones. In: Poulton R (ed) The lost manor of Hextalls. Little Pickle, Bletchingley. Surrey County Archaeological Unit, p 139–164
Braudel F (1992) Civilization and capitalism, 15th–18th century. Vol. II. The wheels of commerce. University of California Press, Berkeley
Burns WE (2015) The scientific revolution in global perspective. Oxford University Press, Oxford
Campbell MS, Overton M (1993) A new perspective on medieval and Early Modern agriculture: six centuries of Norfolk farming c.1250-c.1850. Past & Present 141:38–105
Cipolla C (1970) European culture and overseas expansion. Penguin Books, London
Clark G (1999) Too much revolution: agriculture and the industrial revolution 1700-1860. In: Mokyr J (ed) The British Industrial Revolution: an economic assessment. Westview Press, Boulder, pp 206–240
Crosby AW (2003) The Columbian exchange: biological and cultural consequences of 1492. Greenwood Publishing Group, Westport
Davis SJM (1996) Measurements of a group of adult female Shetland sheep skeletons from a single flock: a baseline for zooarchaeologists. J Archaeol Sci 23:593–612
Davis SJM (1997) The Agricultural Revolution in England: some zoo-archaeological evidence. Anthropozoologica 25(26):413–428
Davis SJM (2000) The effect of castration and age on the development of the Shetland sheep skeleton and a metric comparison between bones of males, females and castrates. J Archaeol Sci 27:373–390. https://doi.org/10.1006/jasc.1999.0452
Davis SJM, Beckett JH (1999) Animal husbandry and agricultural improvement: the archaeological evidence from animal bones and teeth. Rural Hist 10(1):1–17
Degerbøl M (1963) Prehistoric cattle in Denmark and adjacent areas. In: Mourant AE, Zeuner FE (eds) Man and cattle. Royal Anthropological Institute, London, pp 69–79
Dodds B, Britnell R (2008) Agriculture and rural society after the Black Death: common themes and regional variations. University of Hertfordshire Press, Hatfield
Dyer C (1981) Warwickshire farming 1349-c.1520: preparations for the Agricultural Revolution. Dugdale Society Occasional papers 27. Oxford
Dyer A (2002) Small towns in England. 1600-1800. In: Borsay P, Proudfoot L (eds) Provincial towns in Early Modern England and Ireland. Change, convergence and divergence. Proceedings of the British Academy 108. Oxford University Press, Oxford, pp 53–67
Epstein H, Bichard M (1984) Pig. In: Mason I (ed) Evolution of domesticated animals. Longman, London, pp 145–161
Fernández J, González I (1990) La agricultura viajera: cultivos y manufacturas de plantas industriales y alimentarias en España y en la América virreinal. Real Jardín Botánico (CSIC), Madrid
Fitzherbert A (1523/34) Boke of husbandry
Fitzherbert A (1523/39) The boke of surveying and improvements
Fothergill BT (2014) The husbandry, perception and “improvement” of the Turkey in Britain, 1500-1900. Post-Mediev Archaeol 48(1):210–231
Fussell GE (1961) Introduction part one: before 1815. In: Prothero RE, English farming, past and present, 6th edn. Heineman, London, pp 21–75
Gidney LJ (1991a) Leicester, The Shires 1988 excavations. The animal bones from the post-medieval deposits at St Peter’s Lane. English Heritage ancient monuments laboratory report, London 131/91
Gidney LJ (1991b) Leicester, The Shires 1988 excavations: the animal bones from the medieval deposits at Little Lane. English Heritage Ancient Monuments Laboratory Report, London 57/91
Gidney LJ (1991c) Leicester, The Shires 1988 excavations: the animal bones from the medieval deposits at St Peter’s Lane. English Heritage Ancient Monuments Laboratory Report, London 116/91
Gidney LJ (1992) Leicester, The Shires 1988 excavations. The animal bones from the post-medieval deposits at Little Lane. English Heritage Ancient Monuments Laboratory Report, London 24/92
Gidney LJ (1993) Leicester, The Shires 1988 excavations: further identifications of small mammal and bird bones. English Heritage Ancient Monuments Laboratory Report London 92/93
Gordon RL (2016) Feeding the city: zooarchaeological perspectives on urban provisioning and consumption behaviours in post-medieval England (AD1500-AD1900). PhD thesis, University of Leicester. Available from: https://lra.le.ac.uk/handle/2381/37809
Grau-Sologestoa I (2015) Livestock management in Spain from Roman to post-medieval times: a biometrical analysis of cattle, sheep/goat and pig. J Archaeol Sci 54:123–134. https://doi.org/10.1016/j.jas.2014.11.038
Guinn-Chipman S (2013) Religious space in reformation England: contesting the past. Religious cultures in the Early Modern world. Routledge, London
Hadjikoumis A (2010) The origins and evolution of pig domestication in prehistoric Spain. PhD dissertation, The University of Sheffield
Havinden M (1961) Agricultural progress in open field Oxfordshire. Agric Hist Rev 9:73–83
Henry H (2008) The scientific revolution and the origins of modern science. Palgrave Macmillan, Basingstoke
Hogg RV, Tanis E, Zimmerman D (2015) Probability and statistical inference. 9th ed. Pearson Education
Holmes M (forthcoming) Southern England: a review of animals from Saxon, Medieval and Post Medieval Archaeological Sites. Historic England Research Report, Portsmouth
Hopcraft RL (1994) The social origins of agrarian change in late medieval England. Am J Sociol 99(6):1559–1595
Hoskins WG (1955) Sheep farming in Saxon and medieval England. Department of Education of the International Wool Secretariat, London
Huntley J, Stallibrass S (1995) Plant and vertebrate remains from archaeological sites in Northern England: data reviews and future directions. Research Report N. 4. Architectural and Archaeological Society of Durham and Northumberland
Jack SM (1996) Towns in Tudor and Stuart Britain. Macmillan Press, London
Jeffery R (2006) Hugo Grotius in International Thought. Palgrave MacMillan History of International Thought. Palgrave Macmillan, Basingstoke
Johnson M (1996) An archaeology of capitalism. Blackwell, Oxford
Jones EL (1965) Agricultural and economic growth in England 1650-1750: agricultural change. J Econ Hist 25:1–18
Jones RH (1980) Medieval houses at Flaxengate, Lincoln. In: The archaeology of Lincoln, volume XI: medieval Lincoln: the Lower City, p 1–45
Kerridge E (1967) The Agricultural Revolution. Allen & Unwin, London
Kerridge E (1969) The Agricultural Revolution reconsidered. Agric Hist 43:463–476
Kiley M (1976) A review of the advantages and disadvantages of castrating farm livestock with particular reference to behavioural effects. Br Vet J 32:323–331
Kraye K (1996) The Cambridge companion to Renaissance humanism. Cambridge University Press, Cambridge
Langdon J (1986) Horses, oxen and technological innovation: the use of draught animals in English farming from 1066 to 1500. Cambridge University Press, Cambridge
Lefebvre des Noëttes, RJEC (1931) L'Attelage. Le cheval de selle à travers les ages: Contribution à l'histoire de l'ésclavage. Paris, Picard
Love RS (2006) Maritime exploration in the age of discovery, 1415–1800. Greenwood Publishing Group, Westport
Lucas, J, Buckley, R (2007) Excavations at Little Lane and St Peter’s Lane (Shires), Leicester 1988–9, unpublished University of Leicester Archaeological Services report
Maltby M (1979) The animal bones from Exeter 1971-1975. Sheffield University Press, Sheffield
Maltby M (1982) The animal and bird bones. In: Higham R, Allen J, Blaglock S (eds) Excavations at Okehampton Castle, Devon part 2—the Bailey. Proceedings of the Devon archaeological society. 40:114–138
Meadow R (1999) The use of size index scaling techniques for research on archaeozoological collections from the Middle East. In: Becker C, Manhart H, Peters J, Schibler J (eds) Historia animalium ex ossibus. Festschrift für Angela von den Driesch. Verlagmarie Leidorf GmbH, Rahden/Westf, pp 285–300
Meat Atlas (2014) Facts and figures about the animals we eat. Heinrich Böll Foundation, Berlin, Germany, and Friends of the Earth Europe, Brussels, Belgium
Miller E (1991) The agrarian history of England and Wales, vol III. Cambridge University Press, Cambridge, pp 1348–1500
Moorhouse G (2012) The last divine office: Henry VIII and the dissolution of the monasteries. Bluebridge, New York
Nauert CG (2006) Humanism and the culture of Renaissance Europe (new approaches to European history). Cambridge University Press, Cambridge
Nunn N, Qian N (2010) The Columbian exchange: a history of disease, food, and ideas. J Econ Perspect 24(2):163–188
O’Connor TP (1982) Animal bones from Flaxengate, Lincoln, c.870-1500. In: Jones MJ (ed) The archaeology of Lincoln. Volume XVIII: environmental studies, p 1–51
O’Connor TP (1995) Size increase in post-medieval English sheep: the osteological evidence. Archaeofauna 4:81–91
Orakhelashvili A (2006) The idea of European international law. Eur J Int Law 17(2):315–347
Overton M (1984) Agricultural Revolution? Development of the agrarian economy in Early Modern England. In: Baker ARH, Gregory D (eds) Explorations in historical geography: interpretative essays. Cambridge University Press, Cambridge, pp 118–235
Overton M (1996a) Re-establishing the English Agricultural Revolution. Agric Hist Rev 44:1–34
Overton M (1996b) Agricultural Revolution in England: the transformation of the agrarian economy 1500-1850. Cambridge University Press, Cambridge
Overton M, Campbell B (1992) Norfolk livestock farming 1250-1740: a comparative study of manorial accounts and probate inventories. J Hist Geogr 18(4):377–396
Parker CH (2010) Global interactions in the Early Modern Age, 1400–1800. Cambridge University Press, Cambridge
Parry JH (1981) The age of reconnaissance. University of California Press, Berkeley
Payne S, Bull G (1988) Components of variation in measurements of pig bones and teeth, and the use of measurements to distinguish wild from domestic pig remains. Archaeozoologia II/1(2):27–66
Penrose B (1952) Travel and discovery in the Renaissance, 1420-1620. Harvard University Press, Harvard
Popkin P, Baker P, Worley F, Payne S, Hammon A (2012) The sheep project (1): determining skeletal growth, timing of epiphyseal fusion and morphometric variation in unimproved Shetland sheep of known age, sex, castration status and nutrition. J Archaeol Sci 39:1775–1792
Postan M (1939) Revisions in economic history, IX—the fifteenth century. Econ Hist Rev 9:160–167
Poulton R (1998) The lost manor of Hextalls. Little Pickle, Bletchingley. Surrey County Archaeological Unit
Prothero RE (1912) English farming, past and present, 6th edn. Heineman, London
Ratcliffe DA (1984) Post-medieval and recent changes in British vegetation; the culmination of human influence. New Phytol 98:73–100
Reed KM (2008) Turkey. In: Cockett NE, Kole C (eds) Genome mapping and genomics in domestic animals. Springer, Heidelberg, pp 143–164
Rice EF, Grafton A (1994) The foundations of Early Modern Europe, 1460-1559. W.W. Norton, New York
Russell N (1986) Like Engend’ring like: heredity and animal breeding in Early Modern England. Cambridge University Press, Cambridge
Silberberg M, Silberberg R (1971) Steroid hormones and bone. In: Bourne GH (ed) The biochemistry and physiology of bone, vol III. Academic Press, London, pp 401–484
Simpson GG (1941) The large Pleistocene felines of North America. Am Mus Novit 1136:1–27
Skinner Q (2007) Renaissance virtues: visions of politics: volume II. Cambridge University Press, Cambridge
Stallibrass S (1993) Animal bones from excavations at a medieval hospital and a post-medieval farmstead at St Giles by Brompton Bridge, North Yorkshire, 1989-1990. English Heritage, London, AML Report 95/93
Sykes N (2006) From Cu and Sceap to Beffe and Motton: the management, distribution and consumption of cattle and sheep in medieval England. In: Woolgar C, Serjeantson D, Waldron T (eds) Food in medieval England: diet and nutrition. Oxford University Press, Oxford, pp 56–71
Tarlow S (2007) The archaeology of improvement in Britain, 1750–1850. Cambridge University Press, Cambridge
Thirsk J (1967) The agrarian history of England and Wales, vol IV. Cambridge University Press, Cambridge, pp 1500–1640
Thirsk J (1987) Agricultural regions and agrarian history in England, 1500-1750. Macmillan, London
Thirsk J (2007) Food in Early Modern England: phase, fad, fashions 1500-1760. Hambledon Continuum, London
Thomas R (2005a) Zooarchaeology, improvement and the British Agricultural Revolution. Int J Hist Archaeol 2:71–87
Thomas R (2005b) Animals, economy and status: the integration of zooarchaeological and historical evidence in the study of Dudley Castle, West Midlands (c.1100-1750). BAR British Series 392. Archaeopress, Oxford
Thomas R (2009) Bones of contention: why later post-medieval assemblages of animal bones matter. In: Horning A, Palmer M (eds) Crossing paths or sharing tracks: future directions in the archaeological study of post-1550 Britain and Ireland. Boydell and Brewer Ltd, Woodbridge, pp 133–148
Thomas R (2010) Translocated testudinidae: the earliest archaeological evidence for tortoises in Britain. Post-Mediev Archaeol 44:165–171
Thomas R, Holmes M, Morris J (2013) “So bigge as bigge may be”: tracking size and shape change in domestic livestock in London (AD 1220–1900). J Archaeol Sci 40(8):3309–3325. https://doi.org/10.1016/j.jas.2013.02.032
Trow-Smith R (1957) A history of British livestock husbandry to 1700. Routledge and Kegan Paul, London
Trow-Smith R (1959) A history of British livestock husbandry 1700-900. Routledge and Kegan Paul, London
Turner ME, Beckett JV, Afton B (1996) Taking stock: farmers, farm records and agricultural output in England, 1700-1850. Agric Hist Rev 44:21–34
Van der Veen M (2010) Agricultural innovation: invention and adoption or change and adaptation? World Archaeol 2(1):1–12
Vann S, Grimm J (2010) Post-medieval sheep (Ovis aries) metapodia from southern Britain. J Archaeol Sci 37:1532–1542. https://doi.org/10.1016/j.jas.2010.01.014
Verhulst A (1990) The Agricultural Revolution of the Middle Ages reconsidered. In: Bachrach BS, Nicholas D (eds) Law, custom and the social fabric of medieval Europe: essays in honor of Bryce Lyon, Medieval Institute Publications. Western Michigan University, Michigan, pp 17–28
Weston R (1645) A discourse of husbandry of Brabant and Flanders
Williamson T (2002) The transformation of rural England: farming and the landscape 1700-1870. University of Exeter Press, Exeter
Wooton D (2015) The invention of science: a new history of the scientific revolution. Penguin Books, London
Worlidge J (1668) Systema Agriculturae, Being the mystery of husbandry discovered and laid open
Wright, E, Corbino, C, Albarella U (2016) The animal bones from Norton Priory, Runcorn, Cheshire. Unpublished report
Wrigley E, Schofield R (1981) The population history of England, 1541-1871: a reconstruction. E. Arnold, London
Yalden D (1999) The history of the British mammals. T. & A.D. Poyser, London
Acknowledgements
The authors would like to thank those colleagues who generously provided raw data to support our analysis: the Museum of London Archaeology, Richard Thomas and Jim Morris (London evidence); Mark Maltby (Exeter); Lizzie Wright and Chiara Corbino (Norton Priory); Sue Stallibrass (Saint Giles by Brompton Bridge). We would also like to thank Matty Holmes for letting us consult her forthcoming review of the zooarchaeology of southern England.
We wish to express our gratitude to the Museums of Guildford, Lincoln and Leicester for providing us access to the faunal remains from Little Pickle, Flaxengate and The Shires, respectively.
Many thanks also to Tamsyn Fraser, Simon Davis and Konstantinos Sechidis for their valuable comments.
Funding
This research was carried out thanks to a Post-Doctoral Fellowship funded by the Basque Government (Gobierno Vasco – Eusko Jaurlaritza) awarded to Idoia Grau-Sologestoa. This research was supported by the project “Peasant agency and social complexity in north-western Iberia in the medieval period” (Spanish Ministry of Economy, Industry and Competitiveness AEI/FEDER UE HAR2016-76094-C4-2R), the Research Group in Heritage and Cultural Landscapes (Government of the Basque Country, IT931-16) and the Group of Rural Studies (Unidad Asociada UPV/EHU-CSIC).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Online Resource 1
(PDF 792 kb)
Online Resource 2
(XLSX 162 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Grau-Sologestoa, I., Albarella, U. The ‘long’ sixteenth century: a key period of animal husbandry change in England. Archaeol Anthropol Sci 11, 2781–2803 (2019). https://doi.org/10.1007/s12520-018-0723-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-018-0723-6