
Abstract
Understanding the relationship between Neanderthal groups and their environment and they it is essential to comprehending their ways of life. In this article, we use both zooarchaeology and taphonomy to study the fauna assemblages found in Level 4 of Prado Vargas Cave (Cornejo, Burgos, Spain). The results point to a site in which the main accumulating agent was Neanderthal groups, who transported small- and medium-sized animal carcasses—with deer as the dominant taxon—according to their general utility to systematically and intensely exploit the major muscle bundles, bone marrow, skin, and tendons. According to dental microwear analysis, reiterated and prolonged occupations occurred in diverse moments, suggesting that the site was used most often as a long-term campsite. We detected a minimal amount of carnivore activity, who accessed the cave during periods of human absence and modified some of the remains left by the Neanderthals. Together, this information indicates that Prado Vargas is a key site to understand the dynamics of the Neanderthals in the linking area between the Castillan Plateau and the Cantabrian Range.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The study of fauna plays a fundamental role in the interpretation of human occupations, given what faunal assemblages related to their activities tell us about the subsistence strategies of human groups. Anthropogenic evidence allows us to know the degree of interaction between human groups and the surrounding fauna, both prey and predators (Binford 1981). The subsistence strategies and the method of obtaining animal resources open up a wide range of possibilities that these groups could have used, from scavenging to hunting strategies (Costamagno et al. 2006; Niven et al. 2012; Rendu 2022; Selvaggio 1994; Stiner 1994) (Costamagno et al. 2006; Niven et al. 2012; Rendu 2022; Selvaggio 1994; Stiner 1994). However, both practices are not exclusive. As O’Connell et al. (1988) report, “hominids from all time periods would have welcomed the windfall of scavenging a freshly dead animal when the circumstance arose.”
From an archaeological perspective, it is necessary to establish methods that allow us to assess the patterns that govern occupations in a very precise way regarding function, intensity, and organization (Blasco, 2011; Carbonell and Rosell 2004; Daujeard and Moncel 2010; Rosell 2001; Vaquero 1997). The type of occupation reflects how populations adapt to the environment in which they live and how they take advantage of their resources, being a direct evidence of the behavioral variability expressed through the acquisition and management of biotic resources and territorial mobility (Moncel and Daujeard 2012). Establishing the different types of occupation is as complex, due to the number of influencing factors and the palimpsest effect, as it is interesting as it allows us to learn more about the subsistence strategies and exploitation of the landscape they inhabited. For this reason, several authors have proposed different models. From an ethnographic point of view, Binford (1978a, b, 1980, 1983) proposes two types of groups: (1) foragers, who do not store, have greater residential mobility; and (2) collectors, who do store, have greater logistical mobility and more complex settlement patterns. Both have a base camp, which is the center of their activities. In the case of collector groups, the base camp is the place from which certain members of the group depart to places where resources are obtained (Kelly 1992). From an archaeological perspective, Carbonell and Rosell (2004) propose 4 types of occupation: (1) referential intervention centers correspond to base camps in which we observed a large quantity of remains. With respect to the fauna, herbivores dominate the assemblage, and it is common to observe differential transport of anatomical parts of the hunted animals. The remains show a large number of anthropogenic marks associated with fleshing, disarticulation, and breakage. Non-human carnivores attracted by the abandoned remains would leave their mark in times of human absence; (2) complementary intervention centers are places where specialized activities are carried out in which the accumulations of remains are smaller than in the referential intervention centers, unless the reiteration of their use produces palimpsests. Related to fauna, these sites show a great interpretative complexity, since they are usually related to traps or natural hunting grounds where the carcasses are in anatomical connections or semi-connections and show cut marks related to the first stages of the operative chain. Depending on the body size, the whole animal can be transported to a referral center. Due to the low human presence, the activity of other scavengers is high, finding tooth marks and coporlites; (3) opportunistic and marginal intervention centers are places where accumulations are very low and respond to occupations of short temporal duration. Faunal remains predominate over lithic industry, reflecting the last stages of the operational chain related to the last consumption (mainly breakage); and (4) specialized and singular intervention centers are intentional accumulations that go beyond strictly economic and substantial activities. Several Neanderthal sites have been associated with both foragers (Castel et al. 2017; Daujeard and Moncel 2010; Gaudzinski-Windheuser and Roebroeks 2011; Jauberte et al. 2008; Martínez-Moreno et al. 2004; Mellars 1996; Niven et al. 2012; Rendu et al. 2012; Rendu and Armand 2009) and collectors (Marín et al. 2019; Morin 2012; Patou-Mathis 2000; Rendu 2010; Romandini et al. 2014; Wallace and Shea 2006).
The intensity and length of time that the sites have been occupied also tell us about the function of the site. Daujeard and Moncel (2010) propose three types of sites based on these variables: (1) long-term residential camps, (2) short-term regular hunting camps, and (3) brief stopover camps. However, subsistence strategies may change seasonally, expanding the variability and complexity of occupations (Binford, 1980). In addition, it is generally accepted that nearly all sites are palimpsests, making interpretation of deposits and occupation type difficult (Bailey 2007; Lucas 2005, 2012). Long-term occupations are characterized by a high density of remains (lithic and faunal), a large settlement area, low carnivore activity, and the presence of complete operational chains (Bargalló et al. 2020; Bicho and Cascalheira 2020; Costamagno et al. 2006; Gabucio et al. 2014; Kuntz et al. 2016; Moncel and Rivals 2015; Vaquero et al. 2019). Short-term occupations are characterized by few remains, small areas, significant carnivore activity, and lithic tools configured outside the camp (Costamagno et al. 2011; Niven et al. 2012; Picin and Cascalheira 2020; Vallerdú et al. 2005; Villaverde et al. 2017). At the archaeological level, it has been possible to identify sites used as long-term camps (Bar-Yosef et al., 1992; Blasco et al. 2016; Daujeard, 2008; Morin 2012; Rendu 2010; Romandini et al. 2014), and short-term (Castel et al. 2017; Conard et al. 2012; Delagnes and Rendu 2011; Jauberte et al. 2008; Marín-Arroyo 2013; Rendu and Armand 2009; Rosell et al., 2012b; Valensi et al. 2012) and special purpose sites such as intermediate slaughter or butchering places (Costamagno et al. 2006; Rendu et al. 2012). In the Iberian Peninsula, we find numerous sites in which Neanderthals are the main accumulating agent and which are interpreted as long-term (Gabucio et al. 2014; Luret et al. 2020; Marín et al. 2019; Romero et al. 2019; Rosell et al. 2012a, b; Sañudo et al. 2016; Villaverde et al. 2017) and short-term residential sites, especially hunting camps or hunting stands (Baldeón, 1993; Bargalló et al. 2020; Martínez-Moreno et al. 2004; Moclán et al. 2021; Pérez et al. 2017a, b, 2020; Real Margalef et al. 2019; Rios-Garaizar et al. 2015; Salazar-García et al. 2013; Sánchez-Romero et al. 2020; Sanchis et al. 2019; Utrilla et al. 2010; Uzquiano et al. 2012). However, their functionality is not always known (Ready 2013; Ríos Garaizar et al. 2020; Sánchez-Hernández et al. 2019; Yravedra Sainz de los Terreros and Gómez Castanedo 2011).
Traditionally, zooarchaeological and seasonality studies do not consider the stratigraphic packages that help to distinguish possible events. To confront the problems associated with the palimpsest effect in this study, we are working on a protocol that draws from disciplines such as zooarchaeology—especially taphonomy, dental wear, and bone refits—and microstratigraphy, allowing us to distinguish events and categorize occupations. In this paper, we present the first step of this protocol, a zooarchaeological, taphonomic, and dental wear study which provides data that sheds light on the subsistence strategies of these Neanderthals within the variability shown by the groups of the Iberian Peninsula and, particularly, of the Cantabrian range.
Prado Vargas
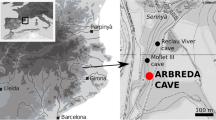
The Prado Vargas cave is located in the Ojo Guareña karst complex (Burgos, Spain) (Fig. 1). Prado Vargas is a 120-m-long subhorizontal gallery interpreted as a karstic paleo-spring located in the sixth level of the complex, which would flow into the Trema river valley (Ortega et al. 2014). The cave opens on the right bank of the valley, currently located about 20 m above the Trema riverbed (Navazo et al. 2021).

Geographic situation of Prado Vargas. a Location of Prado Vargas in the Iberian Peninsula. b Location of the site in Burgos province. c Location of the site at the Ojo Guareña Natural Monument
Prado Vargas is the oldest karst site in Ojo Guareña and was discovered by Trinidad Torres in 1986, who excavated several trenches looking for Ursus spelaeus and ended up finding the Mousterian level (N4). A campaign has been active since 2006, and from 2016 to the present our interdisciplinary team has been in charge of it.
The stratigraphic sequence is composed of 9.3 m of sands, gravels, and clays that are divided into 9 lithostratigraphic units (Fig. 2). To date, layers 4 and 8 are the only ones which contain archaeological material. Unit or level 4 (N4), which is the focus of this paper, has been the only one studied so far (Alonso-García et al. 2020; Cabornero et al. 2016; de la Fuente Juez et al. 2020; Navazo et al. 2005, 2021). N4 has been dated by optically stimulated luminescence (OSL), radiocarbon dating, and AAR dating, yielding dates between 54.7 and 39.8 ka BP (Navazo et al. 2021).

Stratigraphic sequence of Prado Vargas (modified from Navazo et al. 2021)
Pollen analyses reflect a cold climate with a certain degree of humidity, characterized by landscapes of large arboreal spaces and some pine (Pinus), birch (Betula), and juniper coppices (Juniperus). This is confirmed by the macro- and micromammal fauna (Navazo et al. 2021). In previous studies, the presence of different taxa has been documented, highlighting the presence of red deer (Cervus elaphus) and caprines (Capra pyrenaica and Rupicapra pyrenaica), among other species, such as horses (Equus ferus), wild boars (Sus scrofa), Bos/Bison, rabbits (Oryctolagus sp.), cave bear (Ursus spelaeus), wolves (Canis lupus), foxes (Vulpes vulpes), or badgers (Meles meles). These animals were transported to the cave, where the carcasses were intensively processed, from the meat to the marrow (de la Fuente Juez et al. 2020). But their use did not end there, as certain bone fragments became part of the technological sphere and were used as retouchers (Alonso-García et al. 2020).
The lithic industry is mainly made of local flint, although there are other allochthonous raw materials. All structural categories are represented, in other words, the entire reduction sequence, and exploitation is oriented towards intensive use of the material, with hierarchical discoid carving predominating. Flakes are the most representative element, cores represent 5% of the set, and the rest is formed by hammers and fragments. Discoid and Levallois are the best represented exploitation systems. The size of most retouched items is somewhat larger than that of the unretouched flakes, mostly micro- and small-sized. The most representative type is the sidescraper, followed by denticulates, notches and, with much less frequency, points and endscrapers (Navazo et al. 2005, 2021, 2022). Traceology has revealed that some tools were used for skin scraping, and meat and wood processing (Cabornero et al. 2016; Navazo et al. 2022; Santamaría Cabornero et al. 2021).
In addition, Prado Vargas 1360 deserves a special mention. This is the dental crown of a 9–10-year-old Homo neanderthalensis infant individual with evidence of resorption, suggesting that the tooth was lost in life due to tooth replacement (Navazo et al. 2021).
Methodology
The faunal remains from N4 have been taxonomically and anatomically classified using the reference anatomical collection of the Prehistory Laboratory of the University of Burgos and the Centro Nacional de Investigación sobre la Evolución Humana (CENIEH—National Center for Research on Human Evolution) and comparative manuals (e.g., Barone 1976; Pales and Lambert 1971; Schmid 1972). When they could not be identified, they were classified according to the size categories listed by Bunn (1986) based on studies of African mammals and adapted in later work for European sites (Cáceres 2002): very large size (VLS) > 1000 kg; large size (LS) 1000–300 kg; medium size (MS) 300–100 kg; small size (SS) 100–20 kg; very small size (VSS) < 20 kg. Likewise, small–medium size (S-MS) and large–medium size (L-MS) have been used when it has not been possible to discern to which of the two they belong. The Shannon-Wiener index and the inverse of the Simpson index were used to determine species diversity. Because Shannon’s homogeneity index is more sensitive in determining the value of predominant species, Simpson’s dominance index has also been used to look at the degree of specialization of hunting. Age at death was determined by bone fusion (Barone 1976) and tooth eruption and wear (e.g., Analla et al. 2002; Hillson 1990; Pérez-Barbería and Pérez-Fernández 2009; Schmid 1972; Silver 1969). To analyze mortality profiles, the ternary diagram of Stiner (1990), modified by Discamps and Costamagno (2015), was used for a more valid interpretation of the relative abundance of the different age groups in the assemblage.
For the quantification of the record, the NISP (Number of Identified Specimens), MNE (Minimum Number of Elements), and MNI (Minimum Number of Individuals) were taken into account for each taxon and body size (Lyman 1994; Pickering et al. 2003; Rodríguez Hidalgo 2015; Saladié et al. 2011; Stiner 1994). The method described by Morin (2007) was used to calculate the MNE in radius and metapodials. To assess the completeness of the record and its implications, such as biases or transport and exploitation decisions, the %MAU was calculated (Binford 1978, 1984). Various processes can lead to the disappearance of skeletal remains, with density-mediated attrition being particularly influential (R. L. Lyman 1994). In order to study differential preservation as a function of density, Spearman’s correlation coefficient between %MAU and the previously described density values for different body sizes has been calculated (Lam et al. 1999; Lyman 1985; Pavao and Stahl 1999). Since there are no values for VLS, those for LS have been used. Spearman’s correlation coefficient has also been used to analyze the relationship between the selective transport of carcasses and their economic interest, through the %MAU for the different size categories and species identified and different utility indices: [S]FUI, [S]AVGFUI, [S]MAVGMARa, [S]MAVGMARb, UMI (Emerson 1993; Metcalfe and Jones 1988; Morin 2007). To assess the transport strategy, the Shannon unit index (E) for high survival elements (Faith and Gordon 2007) was used and the “Equitability_J” function in the PAST software was employed. In order to evaluate the similarity with long-term (LT) or short-term (ST) models, a discriminant analysis (LDA) has been performed with our %MAU sample and that of other assemblages suggested as long term or short term, as continuous independent variables, with long term, short term, and Prado Vargas (PV) being considered as categorical dependent variables following Marín et al. (2019) and Moclán et al. (2021).
All the remains have been measured in millimeters and breakage has been analyzed distinguishing between green and dry (Villa and Mahieu 1991). Diagnostic elements of intentional breakage have also been documented: percussion notches, percussion pits, pseudo-notches, positive flakes, adhering flakes, flakes, microstriations and striations, and peeling (Capaldo and Blumenschine 1994; Coil et al. 2020; Pickering and Egeland 2006; Vettese et al. 2020). To discern possible breakage patterns, their specific location in the bone has been collected. Furthermore, they have been classified as isolated, opposing, correlative, and consecutive (Rosell 2001).
Bone surface modifications (BSM) were analyzed with a MOTIC SMZ-140 Series binocular loupe. Cut marks (CM) were examined according to their type, location, and morphology. (Binford 1981; Fernández-Jalvo and Andrews 2016; Shipman and Rose 1983). Thermal alterations were classified as follows: 0, no coloration; 1, dispersed brown; 2, widespread brown; 3, completely black; 4, gray and occasionally blue; 5, very pronounced gray or white (Cáceres 2002). To evaluate the relationship between coloration and position, a correspondence analysis was performed between our data and those offered by Pérez et al. (2017a, b) and the biplot representation was carried out with the categories of the variables considered, on the one hand position (2.5 cm, 5 cm, 7 cm, embers, hearth base, and flames) and on the other hand coloration (brown, black, gray, and white). That is to say, from the data provided by Perez et al. (2017a) by means of the correspondence analysis, the biplot has been generated and once the coordinates of the biplot have been obtained, the position of our sample has been represented in this biplot from the data of the same. The action of carnivores has been identified as tooth marks (TM). Classified according to their morphology, location, and purpose, we distinguished scores, pits, punctures, pitting, furrowing, or crenulated edges, in addition to the alterations caused by digestion (Binford 1981; Domínguez-Rodrigo and Piqueras 2003; Selvaggio 1994). Carnivore identification was performed by comparing the collected pits and scores with experimental models (Andrés et al. 2012; Arilla et al. 2014, 2020; Domínguez-Rodrigo and Piqueras 2003; Saladié et al. 2013; Young et al. 2015). One-sample Student’s T-test was used to evaluate if the means of the sample could take values provided by the literature and to be able to assume or discard the action of different agents. The normality of the data was checked using the Kolmogorov-Smirnov test. To evaluate the type of access to the carcasses, the frequency of percussion marks (PM), cut marks, and tooth marks in the different anatomical elements of the identified size categories and species was analyzed.
Other postdepositional processes such as weathering (Behrensmeyer 1978), trampling (Blasco et al. 2008; Courtenay et al. 2020), abrasion (Behrensmeyer 1990; Brett 1990; Cáceres 2002; Lyman 1994), sedimentary pressure (Shipman 1981), humidity (Coard 1999; Courty et al 1989; Fernández-Jalvo and Andrews 2000), and root action (Behrensmeyer 1978; Cáceres 2002; Fernández-Jalvo 1992) were also described.
Meso- and microwear analyses were done on all available dental remains of Equus ferus, Cervus elaphus, Capra pyrenaica, and Rupicapra pyrenaica from level 4. We selected those that were identified at the species level and whose occlusal surfaces presented facets of wear (i.e., belonging to adult individuals). Meso- and microwear will provide evidence of the dietary traits of the ungulates at two different time scales. Mesowear will inform about the annual average diet of the individuals while microwear will indicate the diet at the time of death, informing about the seasonality at the site.
Tooth mesowear is based on the analysis of the shape and relief of the tooth cusps (Fortelius and Solounias 2000). It is directly observed on the buccal or lingual side of the upper and lower molars, respectively, except for horses where only the lower teeth were analyzed. We selected all molars available and discarded those that could belong to the same individual based on size, wear stage, and laterality. Teeth from young individuals (without wear facets) and old adult individuals (with heavy wear) were discarded from the study (Rivals et al. 2007). The method is based on the classification of cusp shape and relief in seven categories, from high relief and sharp cusps (category 0) to blunt cusps with no relief (category 6) (Mihlbachler et al. 2011). The individual mesowear values were then averaged to obtain the mesowear score (MWS).
Tooth microwear was quantitatively studied according to the low-magnification method (Solounias and Semprebon, 2002). The occlusal surfaces were cleaned with acetone and then ethanol 96%. The surfaces were molded using high-resolution dental silicone (Heraeus Kulzer, PROVIL novo, Light C.D. 2 regular set) and casts were created using clear epoxy resin (C.P. Quimica, EP-1060A/1585B). All teeth were screened under a stereomicroscope to check and discard those presenting taphonomic alterations (King et al. 1999; Uzunidis et al., 2021). The quantification of the microwear patterns was done using a Zeiss Stemi 2000C stereomicroscope at × 35 magnification. To avoid inter-observer error, the analysis was performed by a single observer (FR). Microwear features were quantified in two areas using an ocular reticule of 0.16 mm2 on the paracone of the upper molars and the protoconid of the lower molars. We used the classification of features defined by Solounias and Semprebon (2002) and Semprebon et al. (2004) which distinguish pits and scratches. The presence of some other features is recorded qualitatively (presence/absence): cross scratches, hyper-coarse scratches, gouges, large pits. Scratch texture was qualitatively evaluated using the scratch width score (SWS) which is obtained by giving a score of “0” to a surface with predominantly fine scratches, “1” to one with a mixture of fine and coarse scratches, and “2” to one with predominantly coarse. Individual scores for a sample of teeth were then averaged to obtain the SWS.
To estimate the duration of the faunal assemblages’ accumulation at Prado Vargas, we used the standard deviation (SD) and the coefficient of variation (CV) calculated on the numbers of scratches. These two values allow for the quantification of the variability of the microwear signal which is related to seasonal changes in diet and correlates to the duration of the accumulation events, e.g., hunting events (Rivals et al. 2015). The SD and CV values are plotted on a bivariate heat map to classify each sample: event lasting one season or less (zone A); events longer than a season (zone B); and separate events that occurred during non-contiguous seasons (zone C).
PAST, EXCEL, and SPSS software were used for the statistical calculations analysis carried out in this work. Throughout the work, p < 0.05 was considered statistically significant. With the exception of ternary plots, Student’s T-test, diversity indices, and LDA, statistical analyses were performed using SPSS software. Diversity indices and LDA were computed using PAST software, and Student’s T-test and ternary plots using EXCEL. Mesowear and microwear graphs were produced using R. Adobe Photoshop was used to create the artwork.
Results
Integrity of the faunal assemblage
A total of 6343 remains, bones, antlers, and teeth were studied. The remains were found to be highly fractured, with the majority of the remains being less than 5 cm (see Supplementary File 1). A total of 14.98% of the remains have been identified according to their species. Most of these are ungulates (%NISP = 93.37), Bison priscus, Bos primigenius, Bos/Bison (if unable to determine which), Equus ferus, Cervus elaphus, Dama dama, Capreolus capreolus, Capra pyrenaica, Rupicapra pyrenaica, and Sus scrofa (Fig. 3).

Skeletal elements found on Level 4 of Prado Vargas: a mandible of Sus scrofa; b mandible of Cervus elaphus; c antler of Cervus elaphus
Some of the remains were not identified as species and were assigned to Ungulata, Equidae, Cervidae, and Caprinae. Of the total NISP, 5.79% belong to carnivores Ursus spelaeus, Panthera leo, Canis lupus, Vulpes vulpes, Meles meles, and undetermined remains of Carnivora; and 0.84% belong to Oryctolagus sp., Leporidae, and Aves (Table 1). Simpson’s index indicates a medium species diversity (1-D = 0.61), while the Shannon–Wiener index indicates a low-medium diversity (H' = 1.55).
The 88.21% of the NISP were identified according to body size, LS (8.76%), medium size (39.73%), small size (49.53%), very small size (1.14%), and VLS (0.71%). Only a total of 281 remains (NISP = 4.43%) are indeterminate. The remaining percentage are remains identified only at the anatomical level or according to bone type (long or flat).
Among ungulates, the highest MNI values are for Cervus elaphus (20) and Capra pyrenaica (14). The rest of the species obtain values between 1 and 5, except Rupicapra pyrenaica (7). As for carnivores, the highest values correspond to Canis lupus (5) and Ursus spelaeus (5), the rest between 1 and 3 (Table 1).
Mortality profiles indicate that most individuals are adults, except in the case of carnivores and VSS, where subadult and senile individuals are more represented. Prime-dominated mortality profiles are evident in all taxa, although in the case of Equus ferus and Caprinae, whose results are not projected in the prime zone, they are close (Fig. 4).

Profiles of the mortality of the main documented species. a Bovidae LS (Bison priscus, Bos primigenius and Bos/Bison); b Equus ferus (red) and Equidae (blue); c Cervus elaphus (blue) and Cervidae (red); d Capra pyrenaica (blue) and Caprinae (red); e Rupicapra pyrenaica
The NISP and MNE (Supplementary Table A.1) reveal that the main sizes analyzed show a disproportion between elements of the axial skeleton and the appendicular skeleton, being the most represented elements those of the limbs. Only in the case of Cervus elaphus do parts of the axial skeleton, such as the mandibles and maxillae, have a significant presence. This disproportion is less marked for body sizes, so that, although there is a greater representation of limb elements, the number of elements of the axial skeleton is increased. The elements of the axial skeleton only stand out in VSS. The presence of isolated teeth stands out, although there are also complete dental series.
The %MAU confirms the asymmetry in the representation (Fig. 5; Supplementary Table A.2). To see possible biases, it has been studied the correlated correlation with between bone density and utility values by means of Spearman’s correlation coefficient. As for density, Cervus elaphus (ρ = 0.935; p-value = < 0.01) and DM (ρ = 0.915; p-value = < 0.01) show a strong correlation. The correlation coefficient in the case of Capra pyrenaica has a moderate value, and although it is not statistically significant, it is close to the significance limit (ρ = 0.538; p-value = 0.058) and small size (ρ = 0.542; p-value = 0.056). With respect to utility indices, all body sizes show highly significant correlations with (S)MVGMARa and (S)MVGMARb. This happens with all main species identified, except Bos primigenius, Rupicapra pyrenaica, and Sus scrofa. In the case of Equus ferus, there is a moderate relationship which, although not statistically significant, is close to the limit of significance. Similar but less strong correlations exist with UMI, being significant in Bos/Bison, Equus ferus, and LS, and in Cervus elaphus (ρ = 0.643; p-value = 0.119) and medium size (ρ = 0.679; p-value = 0.094), where the correlations are not significant although the moderate value of the correlation coefficient, which is due to the small sample size considered in this analysis. The (S)AVGFUI yields statistically significant correlations in Bos primigenius and Rupicapra pyrenaica (Table 2).

%MAU of the main documented species and sizes. a Bovidae LS (Bison priscus, Bos primigenius, and Bos/Bison; b Equus ferus; c Cervus elaphus; d Capra pyrenaica and Rupicapra pyrenaica; e Sus scrofa; f LS; g MS; h SS
Calculations made with the Equitability_J function indicate a low correlation with unbiased strategies and null correlation with gourmet. Most of the sizes and species identified correspond to Unbiased and Bulk strategies. However, more Unbiased values correspond (E = 0.842) to medium size (E = 0.8776), small size (E = 0.8569), Equus ferus (E = 0.8445), Cervus elaphus (E = 0.8621), and Capra pyrenaica (E = 0.8666). Bison priscus (E = 0.9418) and Sus scrofa (E = 0.961) corresponded to the Bulk strategy (E = 0.980), while only Bos/Bison (E = 1) corresponded to Unconstrained (E = 1). Large size values (E = 0.9135) are between Unbiased and Bulk. In the case of very large size, Bos primigenius and Rupicapra pyrenaica, the values obtained have not been taken into account due to the low number of samples (Fig. 6; Supplementary Table A.3). Regarding the correlation of %MAU with SFUI, only Bos/Bison (ρ = 0.724; p-value = 0.027) shows a statistically significant correlation. In the case of large size, Bison priscus and Equus ferus, there is a strong relationship although there is no statistically significant correlation (Supplementary Table A.4) (Tabla Correlación MAU-SFUI).

Values of E at 95% CI for the main identified species and sizes and the different transport strategies (Faith and Gordon 2007)
Breakage
A total of 18,653 breakage planes were analyzed, of which the majority are curved (55.20%), oblique (80.54%), and smooth (97.72%) (Fig. 7). The presence of several diagnostic elements of intentional breakage has been documented (%NISP = 17.22). Of these, 52.20% are percussion notches (n = 814) and 26.92% are positive flakes (n = 294). Percussion pits (n = 92), cortical flakes (n = 35), flakes (n = 88), and pseudo-notches (n = 88) were also observed. Most of them are found in the diaphysis (n = 950; 93.05%) and are isolated (n = 445; 68.25%), although there are also overlapped notches (n = 77; 11.81%) and opposed (n = 46; 7.06%). The main species identified (Bison priscus, Bos/Bison, Equus ferus, Cervus elaphus, Capra pyrenaica, Rupicapra pyrenaica, and Sus scrofa) and all body sizes except VSS show some of these elements (Table 4). Cervus elaphus is the species that is most present (NISP = 132), while the levels of sizes that are most present are small size (NISP = 373) and medium size (NISP = 310). According to anatomical elements, long bones are the most present, with the humerus (NISP = 82) standing out, followed by the tibia (NISP = 56), the metatarsus (NISP = 54), the radius (NISP = 57), and the femur (NISP = 41). The presence of percussion notches in a 1st phalanx and a 2nd phalanx is noteworthy. The presence of diagnostic elements of intentional breakage has also been documented in elements of the axial skeleton, in skulls (NISP = 2), ribs (NISP = 5), scapula (NISP = 1), and indeterminate flat bones (NISP = 5).

Summary of the characteristics of the different breakage planes according to the criteria of Villa and Mahieu (1991)
The sides on which the diagnostic elements of intentional breakage are located are very similar in the different species and body sizes, with some exceptions. In the case of medium-size metapodials, they are preferentially located on the anterior side, a tendency that is repeated in Cervus elaphus, while in the case of medium-size humeri on the anterior and exterior sides (Table 3).
Bone surface modifications
Bone surface modification (BSM) has been documented in most of the identified remains, with alterations produced by fossildiagenetic and postdepositional processes (e.g., trampling, roots, chemical corrosion) (Fig. 8). A total of 592 remains were identified with cut marks (%NISP = 9.33), especially in ungulates (Bison priscus, Bos/Bison, Equus ferus, Cervus elaphus, Capra pyrenaica, and Rupicapra pyrenaica). The species with the highest number of identified cut marks was Cervus elaphus (n = 94), followed by Capra pyrenaica (NSP = 9). Cut marks have been documented in all body sizes, with more in small size (NSP = 230; 38.85%) and in medium size (NSP = 170; 28,72%).

Taphonomic alterations documented in Level 4 of Prado Vargas. a Cut marks in the first phalanx of Capra pyrenaica; b cut marks in Cervus elaphus maxilla; c cut marks in femur of small size; d cut marks in Cervus elaphus metatarsal; e percussion notch in Cervus elaphus metatarsal; f percussion notch in large-size femur; g diaphyseal cylinder generated in small-size radius
At the anatomical level, cut marks are mainly found in the elements of the appendicular skeleton. They are mostly documented in unidentified long bones (NSP = 319), tibiae (NSP = 46), humeri (NSP = 45), metatarsals (NSP = 42), and radii (NSP = 39). However, they are also documented in unidentified flat bones (NSP = 10), ribs (NSP = 9), mandible (NSP = 8), vertebra (NSP = 4), coxal (NSP = 3), and scapula (NSP = 1). Flat bones with cut marks are from Cervus elaphus, Rupicapra pyrenaica, large size, medium size, and small size (Table 4).
In terms of typology, the majority of cut marks are incisions (%NSP = 93), followed by sawing (%NSP = 2.65), chop (%NSP = 2.65), and scraping marks (%NSP = 1.7). Occasionally, more than one type of markings are present in the same remains.
A total of 239 remains were identified with tooth marks (%NISP = 3.77), especially in ungulates (Cervus elaphus, Capra pyrenaica, Rupicapra pyrenaica, and Caprinae), although there is one carnivore remains, Vulpes vulpes, with tooth marks. The species most in which tooth marks have been most frequently identified are Cervus elaphus (NSP = 23) and Capra pyrenaica (NSP = 5). In the different sizes, tooth marks are also documented in large size (NSP = 21), medium size (NSP = 67), small size (NSP = 101), and VSS (NSP = 1). In addition, they are documented in 14 indeterminate remains.
At the anatomical level, tooth marks are identified especially in unidentified long bones (NSP = 137). We did not observe a significant difference between the bones identified with percussion marks: elements of the appendicular and axial skeleton were similar. Radii (NSP = 20) and tibiae (NSP = 16) stand out. It is worth noting the presence of two horns with percussion marks (Table 4). The measurements of the tooth marks are detailed in Supplementary Table A.5. In addition to pits and scores, remains with digestion (NSP = 34), pitting (NSP = 30), crenulated edges (NSP = 18), furrowing (NSP = 4), and diaphyseal cylinder (NSP = 1) have been documented.
In 9 remains cut marks and tooth marks overlaps have been found, always in long bones, especially medium size (NSP = 6) and small size (NSP = 3). Only two of them could be identified, a tibia of small size and a metopodial of Caprinae. All but one of the superpositions are Human-Carnivore. It should be noted that 4 of them show percussion marks.
A total of 348 (%NISP = 5.49) burned remains were documented. The dimensions of these remains are 74.43% smaller than 25 mm. At the taxonomic level, only 10 remains were identified: Bison priscus (NISP = 1), Capra pyrenaica (NISP = 3), Rupicapra pyrenaica (NISP = 1), Cervus elaphus (NISP = 4), and Ursus spelaeus (NISP = 1). They have been identified in all body sizes, large size (NISP = 14), medium size (NISP = 83), M-LS (NISP = 2), small size (NISP = 191), small–medium size (NISP = 7), very large size (NISP = 2), and very small size (NISP = 1). Most of the burned remains are unidentified long bones (NISP = 297). Burned elements of the axial and appendicular skeleton were identified, most notably the humeri (NISP = 4), metacarpals (NISP = 4), ribs (NISP = 4), and metatarsals (NISP = 3). According to the portion of the bone, 72.13% belonged to the diaphysis, 12.93% to the metaphysis, and 1.72% to the epiphysis. Regarding the degree of cremation, most of them show grade 2 (NISP = 231) and grade 3 (NISP = 274), followed by grade 4 (NISP = 108). It should be noted that 25 remains show percussion marks, 19 cut marks, and 40 tooth marks.
Other postdepositional alterations have been documented, most notably manganese oxide (%NISP = 79.58), fissures (%NISP = 64.40), polishing (%NISP = 61.22%), rounding (%NISP = 58.98), trampling (%NISP = 48.29), and vermiculations (%NISP = 42.39) (Supplementary Table A.6).
Dental mesowear and microwear
A total of 112 teeth were molded and cast. For the mesowear, 47 teeth (i.e., 42%) were discarded due to taphonomic issues (mainly broken cusps). The preservation of the microwear pattern on the ungulate teeth at Prado Vargas is better than that of mesowear. After screening, 23 teeth were discarded (i.e., 20.5%) due to taphonomic damages (weathering, erosion, etc.). The final sample is made of 65 teeth for mesowear and 89 for microwear.
Cervus elaphus, with a mesowear score of 1.52, overlaps with values reported for the extant leaf browsers and mixed feeders (Table 5; Fig. 9). The microwear pattern is characterized by an intermediate number of pits and scratches, a high proportion of individuals with large pits and gouges, a mixture of fine and coarse scratches, and the absence of cross scratches (Table 5). Compared with extant ungulates with known diets, it fits with the range of the leaf browsers (Fig. 10).

Mesowear scores (MWS) for the ungulates from the level 4 of Prado Vargas compared with the dietary categories in extant ungulates (i.e., leaf browsers, mixed feeders, and grazers). Error bars correspond to the standard error of the mean (± 1 SEM) for each sample

Bivariate plot of the average numbers of pits and scratches for the ungulates from the level 4 of Prado Vargas. Error bars correspond to standard error of the mean (± 1 SEM) for each sample. Plain ellipses correspond to the Gaussian confidence ellipses (p = 0.95) on the centroid for the extant leaf browsers (LB) and grazers (G) based on the reference database from Solounias and Semprebon (2002)
Capra pyrenaica has the lowest mesowear score among all the ungulates analyzed (Table 5). The mesowear score (MWS) indicates a diet with low abrasiveness. The mesowear score fits with values of the extant leaf browsers and mixed feeders (Fig. 9). The microwear pattern shows a low number of scratches and the highest number of pits (Table 5). It also records a high percentage of individuals with large pits and gouges and a texture dominated by relatively large scratches. In comparison with extant ungulates, it corresponds to a browsing diet (Fig. 10).
Rupicapra pyrenaica shows a mesowear score very similar to that of the red deer (Table 5; Fig. 9) indicating a diet corresponding to the extant leaf browsers and mixed feeders. Microwear patterns are also similar to that of Cervus elaphus indicating a leaf browsing diet at the time of death (Fig. 10).
Equus ferus is represented by a low sample size; however, some trends can be highlighted. The mesowear score is the highest among the four taxa studied here, but it reflects an intermediate level of abrasion (Table 5). It overlaps with extant grazers and grass-dominated mixed feeders (Fig. 9). The microwear pattern, observed on a slightly higher number of individuals, shows an intermediate number of scratches, a low number of pits, absence of large pits, and low percentage of individuals with large pits (Table 5). On the bivariate plot (Fig. 2), the sample falls in between the confidence ellipses for the extant leaf browsers and grazers, i.e., they would below to the mixed feeders.
When the NMIs (Table 5) are considered, the assemblage comprises 41 individuals belonging to the leaf browsers or browse-dominated leaf browsers (C. elaphus, C. pyrenaica, and R. pyrenaica) and 5 to the grazers (E. ferus). The ungulates analyzed through dental wear are dominated at 89.1% by the leaf browsers or browse-dominated leaf browsers and only 10.9% were grazers.
The CV and SD values computed on the numbers of scratches for each sample show different patterns. Two samples, Equus ferus and Rupicapra pyrenaica, have very low values (Table 1) that fall in zone A of the heat map (Fig. 11). These low values indicate that all the individuals died at a single season. Cervus elaphus and Capra pyrenaica have higher values (Table 1) that would indicate a longer duration for the accumulation of these animals in the cave. Cervus elaphus, however, falls on the boundary between zones B and C where the probability of error is too high. For Capra pyrenaica, the high values indicate the animals died and were brought back to the cave during a period that lasted longer than a single season (Fig. 11).

Boundary lines with an error probability (heat map) based on standard deviations (SD) and coefficient of variation (CV) values of microwear data used for the classification of Prado Vargas samples into short events (region A), long-term events (region B), or two separate short events (region C)
Discussion
In level 4 of Prado Vargas, we observed a certain diversity of represented taxa, documenting remains of all body sizes, which indicates a varied meat diet centered on ungulates. We observed a predilection for Cervus elaphus, as this was the most represented taxon. However, the presence of a greater number of small size remains leads us to believe that taxa such as Capra pyrenaica were also highly consumed. This is consistent with what has been observed in the Cantabrian range in similar chronologies, where the main ungulates exploited are Cervus elaphus (Altuna 1971; Castaños 2005; Fuentes Vidarte 1980; Luret et al. 2020; Pike-Tay et al. 1999) and caprines (Baldeón, 1993; Castaños 2005; Urquijo et al. 2005). The comparison of meso- and micro dental wear of these ungulates at the time of their deaths indicates a diet based on plants with low abrasion (low phytolith concentration). In total, 90% of the individuals hunted and transported into the cave were leaf browsers (Capra pyrenaica, Cervus elaphus, and Rupicapra pyrenaica), i.e., they fed on herbaceous plants and leaves of shrubs or trees. This indicates an exploitation of forest environments by Neanderthal groups. According to pollen and microfauna data (Navazo et al. 2021), the surrounding of the site was covered with sparse forested areas with pine, birch and juniper, and large open meadows. This would suggest a preference by Neanderthals for hunting in the more forested areas rather than in open habitats.
Mortality patterns of the main taxa identified indicate a selection of adult or juvenile-adult individuals. This prime-dominated mortality profile is linked to the hunting practices of hominids, a practice observed in several European sites (e.g., Castel et al. 2017; Niven et al. 2012; Rendu et al. 2012; Romandini et al. 2014; Starkovich 2017) as well as peninsular sites (e.g., Marín et al. 2019; Moclán et al. 2021; Ríos Garaizar et al. 2020; Salazar-García et al. 2013; Sánchez-Romero et al. 2020). According to the ethology of certain species, we can estimate the time of year in which they died through the study of dental eruption. The remains of Ursus spelaeus suggest they died between January and April; Sus scrofa between September and December; Capra pyrenaica between June and October; and Rupicapra pyrenaica between January and June. The bears seem to have encountered death during hibernation, since they show no anthropogenic or carnivore evidence of predation or transport. But the other taxa do, so their predation could have taken place throughout the year, from January to December. However, only in certain months they are available at once, this being the months between June and October. This suggests an occupation of the cave by hominids between spring and autumn.
The uneven presence of anatomical elements may be due to carcass transport decisions or postdepositional processes. The presence of isolated teeth and other taphonomic processes can tell us about the differential destruction of elements of lower skeletal survival. The correlation with density indices indicates that there is differential preservation in Cervus elaphus and medium size, which cannot be ruled out in Capra pyrenaica and small size. Large size, however, does not seem to show it. The significant amounts of polishing and rounding in the cortical and medular sides of remains suggest the presence of water currents. However, there are anatomical elements that should be transported immediately (Behrensmeyer 1975; Voorhies 1969). Regarding food utility indices, we only observed that Bos primigenius and Rupicapra pyrenaica are exploited according to their general utility. However, all body sizes and the main species identified (Bison priscus, Bos/Bison, Cervus elaphus, Equus ferus, and Capra pyrenaica) show a correlation with the medular fat indices. The Equitability_J function indicates that our samples correspond to unbiased and bulk transport strategies, related to the selective transport of the most economically useful and the maximization of all but the least useful elements. Thus, differential conservation seems to be due to the economic interest of the carcasses, with a systematic selection of the parts with high nutritional value.
Although the first stages of the processing sequence are carried out at the place where the prey is obtained, the carcasses are intensively exploited at the site under study. Most of the cut marks are located in the long bones of all the sizes and main documented species, showing evidence of disarticulation and exploitation of the large muscle bundles. The presence of cut marks in jawbones of Cervus elaphus, Rupicapra pyrenaica, large size, medium size, and small size is evidence of the consumption of the muscular bundles of the masticatory apparatus. Skinning is documented in Bos/Bison, Cervus elaphus, Capra pyrenaica, Rupicapra pyrenaica, large size, medium size, medium size, and small size, with cut marks observed in the metapodials and first phalanges. In the case of metapodials, the cut marks identified also indicate the removal of the tendons of the anterior and posterior sides. The presence of scrapings in the long bones is also evidence of the removal of the periosteum to facilitate breakage, as observed by Binford (1981), Blasco et al. (2014), Marean and Assefa (1999), or Vettese et al. (2020). We observed the presence of evisceration and dismemberment in almost all sizes and in Cervus elaphus. This indicates that, although evisceration may be carried out inside or outside the site, on certain occasions the prey are transported in their entirety to be to be processed in the site. Although cut marks are accidents and there are certain activities that lead to their underrepresentation, we did not observe any specialization in any particular activity.
The high fragmentation of our assemblage is a typical feature of anthropogenic exploitation, since it is the result of the exploitation of bone marrow, as noted by Stiner (1994), Vettese et al. (2020), and Villa and Mahieu (1991). The breakage of the bone in its fresh state and the presence of percussion marks show that hominids consumed this resource once the muscle bundles were consumed. All body sizes and the main species identified show this consumption in elements of the axial and appendicular skeleton. Some authors have tried to discern certain patterns in the breakage (e.g., Blasco et al. 2013; Vettese et al. 2017); however, we were not able to identify any pattern.
The identified tooth marks have been measured and compared with reported in the literature (Andrés et al. 2012; Arilla et al. 2014, 2020; Domínguez-Rodrigo and Piqueras 2003; Saladié et al. 2013; Young et al. 2015), so as to discern the responsible agent (see Supplementary File 2). Since our sample coincides with a large number of carnivores and the total data are not available, the means of our values (see Supplementary Table A.7) were compared with those obtained in the literature by means of the Student T-test (Table 6). In this way, we seek to rule out those species for which there is statistically significant evidence that the means are different. We note that we cannot statistically rule out that in cancellous bone affect foxes, bears, dogs, and humans, and in diaphysis, foxes, hyenas, and humans. Thus, they could have altered some remains and made others disappear. The digested bones can be attributed to Canis lupus, since they measure less than 40 mm, although some of greater length could have been the result of the action of hyenas.
The presence of carnivore bones and their marks indicates their access to some remains. The presence of bears may be due to their death in hibernation or to predation, since they are mostly senile or sub-adult individuals. Hyenas and wolves may perform repeated predation on bears hibernating in a cave (Stiner, 1998), although they may also scavenge bear carcasses (Daschek and Mester 2020). Taphonomic analysis suggests that these bears died naturally during hibernation, since no carnivore or hominid action was observed in the recovered remains. The presence of trampling could be the result of bear movements during hibernation, as observed by Diedrich (2011) and Sauqué et al. (2015), although trampling could also be the result of the transit of the Neanderthals themselves. Bears and small carnivores such as Vulpes vulpes and Meles meles have been shown to be potential set modifiers (Arilla et al. 2014, 2020), like wolves and hyenas (Binford 1978, 1981). Of the latter two, remains of mainly adult and subadult individuals of Canis lupus have been found. Their ability to accumulate remains in caves is limited and circumstantial (Sanchís et al. 2019), although the scarce presence of digested bones and remains of subadult individuals may indicate a more intense use of the cave as breeding dens. The existence of manganese and concretion indicates the presence of humid conditions in the cave (Fernández-Jalvo 1992; Fernández-Jalvo and Andrews 2000), which would make it more habitable at certain times of the year for cave carnivores than hominids following Sanchis et al. (2019). However, the low percentage of tooth marks would indicate that Prado Vargas would correspond more to a referential intervention center as defined by Carbonell and Rosell (2004).
However, the presence of cut marks-tooth marks overlaps indicates that carnivores are not the main accumulating agent, but that, in the case where they are responsible, they were able to access the remains left by hominids. Although some authors propose that if tooth marks affect the same species and body sizes, carnivore access to remains is rapid (Rusch et al. 2019), and access does not appear to be immediate, as some remains are incorporated into the technological sphere, being used as retouchers (Alonso-García et al. 2020).
The presence of burned bones is evidence of the use of fire by Neanderthals. The study of the relationship or association between the grades of cremation and their position indicates that some remains seemed to have been in contact with the embers. The absence of epiphyses and metaphyses, or spongy tissues in general, may be related to the hearths, and bone may have been used as fuel. In addition, other remains indicate that the burned remains could have acquired a brown and black coloration (grades 2 and 3) after having been buried between 2.5 cm and 5 cm beneath a hearth (Fig. 12). The reuse of hearths (Vallverdú et al. 2012) and the accumulation of burned remains or different combustion events are typical of long-term occupations. Thus, there seems to be evidence of an intense and recurrent use of the cave by Neanderthals at different times.

Hominids are the main accumulating agent of level 4, although we must not lose sight of the fact that, like most archaeological sites, this is a palimpsest in which occupations (of Neanderthals and Neanderthals-carnivores) are superimposed with remains that may have undergone modifications in respect to their original positions. In order to see to which model of occupation level 4 would correspond, a linear discriminant analysis (LDA) was performed comparing the %MAU of Bos primigenius, Bison priscus, Bos/Bison, Equus ferus, Cervus elaphus, Capra pyrenaica, Rupicapra pyrenaica, large size, medium size, and small size, with levels and deposits catalogued as long term (Bignon et al. 2006; Blasco et al., 2016; Fernádez Laso et al. 2010; Marín et al. 2019; Romandini et al. 2014; Rosell et al. 2012a, b) or short term (Costamagno 2000; Fernádez Laso et al. 2010; Marín et al. 2019; Moclán et al. 2021; Real Margalef et al. 2019; Rosell et al. 2012a, b; Rosell et al. 2017) (Fig. 13). Once we have plotted the data and created the convex hulls, we observed that there is a very significant difference between long term and short term. Our sample does not coincide with either of the two, our sample does not coincide with either of the two, since the convex hulls do not intersect, but if correlated with either, it would do so with long term. However, the ambiguity of the terms makes its classification difficult. Dental microwear provides information on the duration of hunting events. Capra pyrenaica and perhaps Cervus elaphus were hunted during several seasons of the year, indicating either a long-term occupancy of the site or repeated occupations at different seasons during the formation of level 4. Less represented species, such as Equus ferus and Rupicapra pyrenaica, were hunted during short and seasonal events. These results indicate that Neanderthal groups took advantage of specific resources when they were available in the environment.

LDA between the %MAU of our sample (PV) and suggested sets such as LT and ST. (1) Bos primiegnius; (2) Bison priscus; (3) Bos/Bison; (4) Equus ferus; (5) Cervus elaphus; (6) Capra pyrenaica; (7) Rupicapra pyrenaica; (8) Sus scrofa; (9) LS; (10) MS; (11) SS; (12) Equus ferus Pa Abric Romaní; (13) Equus ferus Pb Abric Romaní; (14) Caprinae Abrigo de la Quebrada IV; (15) Cervidae Abrigo de la Quebrada IV; (16) Equidae Abrigo de la Quebrada; (17) IV Saiga tatarica Moulin-Neuf (Gironde); (18) Rangifer tarandus Moulin-Neuf (Gironde); (19) Equus caballus Moulin-Neuf (Gironde); (20) Bovinae Moulin-Neuf (Gironde); (21) Bovinae Navalmaillo; (22) Equus ferus Navalmaillo; (23) Cervidae Navalmaillo; (24) Cervus elaphus Navalmaillo; (25) VLS Navalmaillo; (26) LS Navalmaillo; (27) MS Navalmaillo; (28) SS Navalmaillo; (29) Equidae Ja Abric Romaní; (30) Equidae Jb Abric Romaní; (31) Equidae M Abric Romaní; (32) Equidae Teixoneres III; (33) Cervidae Teixoneres III; (34) Bovidae Teixoneres III; (35) LS Teixoneres III; (36) MS Teixoneres III; (37) SS Teixoneres III; (38) LS Quesem; (39) MS Quesem; (40) SS Quesem; (41) Cervus elaphus Pa Abric Romaní; (42) Cervus elaphus Pb Abric Romaní; (43) Rangifer tarandus Pincevent; (44) Equus Pincevent; (45) Bovidae M Abric Romaní; (46) Cervidae M Abric Romaní; (47) Cervus elaphus Grotta di Fumane A9; (48) Capreolus capreolus Grotta di Fumane A9; (49) Bovidae Ja Abric Romaní; (50) Cervidae Ja Abric Romaní; (51) Bovidae Jb Abric Romaní; (52) Cervidae Jb Abric Romaní
These results would correspond with other diagnostics of long-term occupations. The high density of remains (Gabucio et al. 2014) is a diagnostic feature that we observed in level 4, where in a large area of 80 m2 more than ten thousand archaeological remains have been recovered. The lithic industry is 90% local flint and complete operational chains have been recovered (Navazo et al. 2021), i.e., there is no evidence of the introduction of tools configured outside the site, something typical of short term. In addition, the traceological analysis indicates hide scraping, wood processing, and meat processing, so we see that the activities carried out at the site go beyond those carried out in short-term occupations. The presence of burnt flint, such as that found at Prado Vargas, is also a long-term feature (Rusch et al. 2019). Likewise, the presence of a left Ldm1 of Homo Neanderthalensis (Navazo et al. 2021) indicates the presence of children in the group, something that would not necessarily be expected in short-term or hunting camps, as opposed to long-term or more residential camps.
Subsequent work will allow us to further clarify the functionality and seasonality of the site, by means of spatial analysis and refits, since the spatial relationship of the remains is fundamental to try to individualize events and more accurately identify the type of occupation.
Conclusions
Several analytical methods and techniques were applied to the zooarchaeological assemblage of the level 4 of Prado Vargas: anatomical and taxonomical identification, taphonomic analysis, and tooth wear analysis. The zooarchaeological and taphonomic analysis of level 4 at Prado Vargas suggests that the site was used repeatedly by Neanderthal groups as a camp more akin to a long-term than a short-term camp, in which intensive use of animal resources was made. Also the microwear indicates either a long-term occupancy of the site or repeated occupations at different seasons during the formation of level 4.
They hunted primarily adult animals and transported their carcasses according to their usefulness, although sometimes they transported them whole. There may have been hydraulic transport of some remains, but this does not appear to have affected the whole assemblage, and differential preservation is due to utility interest. Once at the site, they exploited all the food resources, from the muscle packs and marrow to other elements such as skin and tendons, incorporating the resulting remains into the technological sphere by using them as retouchers. The presence of burn remains is evidence of use of fire by Neandertals, who may have been using bones as fuel. The correlation between color and position of burn remains seems to evidence an intense and recurrent use of the cave.
Competition with carnivores was low; few remains are affected by their activity. Although some of them are digested, we cannot rule out that some of the tooth marks were produced by humans. The low percentage of tooth marks, assuming they are all produced by non-human carnivores, and the presence of cut marks-tooth marks overlaps would indicate that these would have affected the assemblage when the hominins were not in the cave, accessing their remains at times when the cave was less habitable.
All the results shown here (zooarchaeological, taphonomic, and dental wear) and the conclusions we draw from them make Prado Vargas Cave a key site for understanding the dynamics of these penultimate Neanderthals, who were found in a place of transition between the Cantabrian region and the interior of the Iberian Peninsula.
Availability of data and materials
All the data used in the preparation of the article can be found in the text or in the supplementary material.
References
Alonso-García P, Navazo Ruiz M, Blasco R (2020) Use and selection of bone fragments in the north of the Iberian Peninsula during the Middle Palaeolithic: bone retouchers from level 4 of Prado Vargas (Burgos, Spain). Archaeol Anthropol Sci 12(9):1–12. https://doi.org/10.1007/S12520-020-01097-Z/FIGURES/9
Altuna J (1971) Los mamíferos del yacimiento prehistórico de Morín. In: González Echegaray J, Freeman LG (eds) Cueva Morín: Excavaciones 1966–1968, pp 367–398. Patronato de las Cuevas Prehistóricas
Analla M, Muñoz-Cobo J, Calvo JA, Carrasco R, Azorit C (2002) Teeth eruption pattern in red deer (Cervus elaphus hispanicus) in southern Spain. Anales de Biología 24:107–114
Andrés M, Gidna AO, Yravedra J, Domínguez-Rodrigo M (2012) A study of dimensional differences of tooth marks (pits and scores) on bones modified by small and large carnivores. Archaeol Anthropol Sci 4(3):209–219. https://doi.org/10.1007/s12520-012-0093-4
Arilla M, Rosell J, Blasco R, Domínguez-Rodrigo M, Pickering TR (2014) The “‘bear’” essentials: actualistic research on Ursus arctos arctos in the Spanish Pyrenees and its implications for paleontology and archaeology. PLoS ONE 9(7):102457. https://doi.org/10.1371/journal.pone.0102457
Arilla M, Rufà A, Rosell J, Blasco R (2020) Small carnivores’ cave-dwelling: neo-taphonomic study of a badger (Meles meles) sett and its archaeological implications. Hist Biol 32(7):951–965. https://doi.org/10.1080/08912963.2018.1558449
Bailey G (2007) Time perspectives, palimpsests and the archaeology of time. J Anthropol Archaeol 26(2):198–223. https://doi.org/10.1016/J.JAA.2006.08.002
Baldeón A (1993) Baldeón 93-El yacimiento de Lezetxiki (Gipuzkoa, País Vasco). Los niveles musterienses. Munibe (Antropologia-Arkeologia) 45:3–97
Bargalló A, Gabucio MJ, Gómez B, Chacón GM, Vaquero M (2020) Rebuilding the daily scenario of Neanderthal settlement. J Archaeol Sci Rep 29:102139. https://doi.org/10.1016/J.JASREP.2019.102139
Bar-Yosef O, Vandermeersch B, Arensburg B, Belfer-Cohen A, Goldberg P, Laville H, Meignen L, Rak Y, Speth J D, Tchernov E, Weiner S, Clark G A, Garrard A, Henry D O, Hole F, Roe D, Rosenberg K R, Schepartz L A, Shea J J, … Wilson L, (1992) The Excavations in Kebara Cave, Mt. Carmel. Curr Anthropol 33(5):497–550. https://doi.org/10.1086/204112
Barone R (1976) Anatomie comparée des mammifères domestiques (Vol. 1). Vigot
Behrensmeyer A (1975) The taphonomy and paleoecology of Plio-Pleistocene vertebrate assemblages east of Lake Rudolf, Kenya. Bulletin of The Museum of Comparative Zoology. https://doi.org/10.5962/BHL.PART.22969
Behrensmeyer AK (1978) Taphonomic and ecologic information from bone weathering. Paleobiology 4(2):150–162. http://www.jstor.org/stable/2400283
Behrensmeyer AK (1990) Transport/hydrodynamics of bones. In: Briggs DEG, Crowther PR (eds) Palaeobiology: a synthesis, pp 232–235. Blackwell Scientific Publications. https://research.si.edu/publication-details/?id=98714
Bicho N, Cascalheira J (2020) Use of lithic assemblages for the definition of short-term occupations in hunter-gatherer prehistory. In: Cascalheira J, Picin A (eds) Short-term occupations in paleolithic archaeology. Interdisciplinary contributions to archaeology, pp 19–38. Springer, Cham. https://doi.org/10.1007/978-3-030-27403-0_2
Bignon O, Enloe JG, Bémilli C (2006) Étude archéozoologique de l’unité T125: originalité de la chasse des rennes et des chevaux. Gallia Préhistoire 48(1):18–35. https://doi.org/10.3406/GALIP.2006.2433
Binford LR (1978a) Nunamiut ethnoarchaeology. Academic Press
Binford LR, (1978b) Dimensional analysis of behavior and site structure: learning from an eskimo. Am Antiq 43(3)
Binford LR (1980) Wilow smoke and dog’s tails: hunter-gatherer settlement systems and archaeological site formation. Am Antiq 45(1):4–20
Binford LR (1981) Bones : ancient men and modern myths. Academic Press
Binford LR (1983) In puirsuit of the past. Decoding archaeological record. Thames and Hudson, London
Binford LR (1984) Faunal remains from Klasies River mouth. Academic Press
Blasco R (2011) La amplitud de la dieta cárnica en el Pleistoceno medio peninsular: una aproximación a partir de la Cova de Bolomor (Tavernes de la Valldigna, Valencia) y del subnivel TD10-1 de Gran Dolina (Sierra de Atapuerca, Burgos). Tesis Doctoral, Universitat Rovira i Virgili, Tarragona
Blasco R, Domínguez-Rodrigo M, Arilla M, Camarós E, Rosell J (2014) Breaking bones to obtain marrow: a comparative study between percussion by batting bone on an anvil and hammerstone percussion. Archaeometry 56(6):1085–1104. https://doi.org/10.1111/ARCM.12084
Blasco R, Rosell J, Domínguez-Rodrigo M, Lozano S, Pastó I, Riba D, Vaquero M, Peris JF, Arsuaga JL, de Castro JMB, Carbonell E (2013) Learning by heart: cultural patterns in the faunal processing sequence during the Middle Pleistocene. PLoS ONE 8(2):e55863. https://doi.org/10.1371/JOURNAL.PONE.0055863
Blasco R, Rosell J, Fernández Peris J, Cáceres I, Vergès JM (2008) A new element of trampling: an experimental application on the Level XII faunal record of Bolomor Cave (Valencia, Spain). J Archaeol Sci 35(6):1605–1618. https://doi.org/10.1016/J.JAS.2007.11.007
Blasco R, Rosell J, Sañudo P, Gopher A, Barkai R (2016) What happens around a fire: faunal processing sequences and spatial distribution at Qesem Cave (300 ka), Israel. Quatern Int 398:190–209. https://doi.org/10.1016/J.QUAINT.2015.04.031
Brett C (1990) Destructive taphonomic processes and skeletal durability. In: Briggs DEG, Crowther PR (eds) Paleobiology, a synthesis. Blackwell Scientific Publications, Oxford, pp 223–226
Bunn HT (1986) Patterns of skeletal representation and hominid subsistence activities at Olduvai Gorge, Tanzania, and Koobi Fora. Kenya Journal of Human Evolution 15(8):673–690. https://doi.org/10.1016/S0047-2484(86)80004-5
Cabornero CS, Cortés SML, Ruiz MN, Benito-Calvo A, Lerma IM (2016) Methodological approach and first results of use-wear traces on flint tools through confocal microscope. DigitAR - Revista Digital de Arqueologia, Arquitectura e Artes 3:15–22. https://doi.org/10.14195/2182-844X_3_2
Cáceres I (2002) Tafonomia de yacimientos antrópicos en karst. Complejo Galeria (Sierra de Atapuerca, Burgos), Vanguard Cave (Gibraltar) y Abric Romaní (Capellades, Barcelona) - Dialnet [Universitat Rovira i Virgili]. https://dialnet.unirioja.es/servlet/tesis?codigo=280069
Capaldo SD, Blumenschine RJ (1994) A quantitative diagnosis of notches made by hammerstone percussion and carnivore gnawing on bovid long bones. Am Antiq 59(4):724–748. https://doi.org/10.2307/282345
Carbonell E, Rosell J (2004) Ocupaciones de homínidos en el Pleistoceno de la Sierra de Atapuerca. In: Baquedano E, Rubio S (eds) Zona Arqueológica. Miscelania en Homenaje a Emiliano Aguirre, vol 4, pp 102–115. Museo Regional
Castaños PM (2005) Revisión actualizada de las faunas de macromamiferos del Würm antiguo en la Región Cantábrica. In: Neandertales cantábricos, estado de la cuestión. Actas de la reunón científica, pp 201–207
Castel JC, Discamps E, Soulier MC, Sandgathe D, Dibble HL, McPherron SJP, Goldberg P, Turq A (2017) Neandertal subsistence strategies during the Quina Mousterian at Roc de Marsal (France). Quatern Int 433:140–156. https://doi.org/10.1016/J.QUAINT.2015.12.033
Coard R (1999) One bone, two bones, wet bones, dry bones: transport potentials under experimental conditions. J Archaeol Sci 26(11):1369–1375. https://doi.org/10.1006/JASC.1999.0438
Coil R, Yezzi-Woodley K, Tappen M (2020) Comparisons of impact flakes derived from hyena and hammerstone long bone breakage. J Archaeol Sci 120:105167. https://doi.org/10.1016/J.JAS.2020.105167
Conard NJ, Bolus M, Münzel SC (2012) Middle Paleolithic land use, spatial organization and settlement intensity in the Swabian Jura, southwestern Germany. Quatern Int 247(1):236–245. https://doi.org/10.1016/J.QUAINT.2011.05.043
Costamagno S (2000) Stratégies d’approvisionnement et traitement des carcasses au Magdalénien : l’exemple de Moulin-Neuf (Gironde) / Magdalenian acquisition strategies and carcass processing : the example of Moulin-Neuf (Gironde). Paléo, Revue d’Archéologie Préhistorique 12(1):77–95. https://doi.org/10.3406/PAL.2000.1597
Costamagno S, Bon F, Valdeyron N (2011) Conclusion. In: Bon F, Costamagno S, Valdeyron N (eds) Hunting camps in prehistory. current archaeological approaches, Proceedings of the International Symposium, May 13–15, vol 3, pp 347–354. University Toulouse II – Le Mirail, P@lethnology. http://www.palethnologie.org
Costamagno S, Liliane M, Cédric B, Bernard V, Bruno M (2006) Les Pradelles (Mc-le-Franc, France): a mousterian reindeer hunting camp? J Anthropol Archaeol 25(4):466–484. https://doi.org/10.1016/j.jaa.2006.03.008
Courtenay LA, Huguet R, Yravedra J (2020) Scratches and grazes: a detailed microscopic analysis of trampling phenomena. J Microsc 277(2):107–117. https://doi.org/10.1111/JMI.12873
Courty MA, Goldberg P, Macphail R (1989) Soils and micromorphology in archaeology. Cambridge University Press
Daschek ÉJ, Mester Z (2020) A site with mixed occupation: Neanderthals and carnivores at Érd (Hungary). J Archaeol Sci Rep 29:102
Daujeard C (2008) Exploitation du milieu animal par les Néanderthaliens dans le Sud-Est de la France [Université Lumière]
Daujeard C, Moncel MH (2010) On Neanderthal subsistence strategies and land use: a regional focus on the Rhone Valley area in southeastern France. J Anthropol Archaeol 29(3):368–391. https://doi.org/10.1016/j.jaa.2010.05.002
de la Fuente Juez H, Domínguez SG, Ruiz MN, López RB (2020) Taphonomy of the fossil record of macromammals of the Upper Pleistocene at level 4 of the Cueva de Prado Vargas (Cornejo, Burgos, Spain). Munibe Antropologia-Arkeologia 71:5–21. https://doi.org/10.21630/maa.2020.71.02
Delagnes A, Rendu W (2011) Shifts in Neandertal mobility, technology and subsistence strategies in western France. In: Journal of Archaeological Science, vol 38, Issue 8, pp 1771–1783. Academic Press. https://doi.org/10.1016/j.jas.2011.04.007
Diedrich CG (2011) An overview of the ichnological and ethological studies in the Cave Bear Den in Urşilor Cave (Western Carpathians, Romania). 18(1), 9–26. https://doi.org/10.1080/10420940.2011.552578
Discamps E, Costamagno S (2015) Improving mortality profile analysis in zooarchaeology: a revised zoning for ternary diagrams. J Archaeol Sci 58:62–76. https://doi.org/10.1016/J.JAS.2015.03.021
Domínguez-Rodrigo M, Piqueras A (2003) The use of tooth pits to identify carnivore taxa in tooth-marked archaeofaunas and their relevance to reconstruct hominid carcass processing behaviours. J Archaeol Sci 30(11):1385–1391. https://doi.org/10.1016/S0305-4403(03)00027-X
Emerson AM (1993) The role of body part utility in small-scale hunting under two strategies of carcass recovery. In: Hudson J (ed) from bones to behavior. ethnoarchaeological and experimental contributions to the interpretation of faunal remains (Vol. 21, pp. 138–168). Center for Archaeological Investigations. Southern Illinois University at Carbondale Occasional Papers. https://www.academia.edu/27030167/_The_role_of_Body_Part_Utility_in_Small_scale_Hunting_under_Two_Strategies_of_Carcass_Recovery_by_Emerson_1993_Spanish_Translation_
Faith JT, Gordon AD (2007) Skeletal element abundances in archaeofaunal assemblages: economic utility, sample size, and assessment of carcass transport strategies. J Archaeol Sci 34(6):872–882. https://doi.org/10.1016/J.JAS.2006.08.007
Fernández-Laso MC (2010) Remontajes de restos faunísticos y relaciones entres áreas domésticas en los niveles K, L y M del Abric Romaní (Capellades, Barcelona, España) tesis doctoral. In: TDX (Tesis Doctorals en Xarxa). Universitat Rovira i Virgili. http://www.tdx.cat/handle/10803/8632
Fernández-Jalvo Y (1992) Tafonomía de microvertebrados del complejo carstico de Atapuerca: (Burgos) - Dialnet [Universidad Complutense de Madrid]. https://dialnet.unirioja.es/servlet/tesis?codigo=201155
Fernández-Jalvo Y, Andrews P (2000) The taphonomy of pleistocene caves, with particular reference to Gibraltar. In: Stringer CB, Barton RNE, Finlayson JC (eds) Neanderthals on the edge. Oxbow Books, pp 171–182
Fernández-Jalvo Y, Andrews P (2016) Atlas of taphonomic identifications. In: Vertebrate Paleobiology and Paleoanthropology (Issue 9789401774307). Springer. https://doi.org/10.1007/978-94-017-7432-1
Fortelius M, Solounias N (2000) Functional characterization of ungulate molars using the abrasion-attrition wear gradient: a new method for reconstructing paleodiets. Am Museum Novit 3301:1–36
Fuentes Vidarte C (1980) Estudio de la fauna del “El Pendo.” In: El Yacimiento de la Cueva de ‘‘El Pendo", vol 17, pp 215–237. Bibliotheca Praehistorica Hispana
Gabucio MJ, Cáceres I, Rosell J, Saladié P, Vallverdú J (2014) From small bone fragments to Neanderthal activity areas: the case of Level O of the Abric Romaní (Capellades, Barcelona, Spain). Quatern Int 330(1):36–51. https://doi.org/10.1016/J.QUAINT.2013.12.015
Gaudzinski-Windheuser S, Roebroeks W (2011) On Neanderthal subsistence in last interglacial forested environments in Northern Europe. In: Vertebrate Paleobiology and Paleoanthropology (Issue 9789400704145, pp 61–71). Springer. https://doi.org/10.1007/978-94-007-0415-2_7
Hillson S (1990) Teeth. Cambridge University Press
Jauberte J, Hublin J-J, Soressi M, McPherron S, Bailey S, Bordes J-G, Claud E, Mallye J-B, Michel A, Niclot M, Rendu W, Richards M, Ritcher D, Roussel M, Steele T, Taylor G, Texier J-P (2008) Paléolithique moyen récent et Paléolithique supérieur ancien a Jonzac (Charente-Maritime): premiers résultats des campagnes 2004–2006 :: MPG.PuRe. In: Jaubert J (ed) Les sociétés du Paléolithique dans un grand Sud-Ouest de la France |nouveaux gisements, nouveaux résultats, nouvelles méthodes: Vol. Mémoire XLVII (pp. 203–243). Société Préhistorique française. https://pure.mpg.de/pubman/faces/ViewItemOverviewPage.jsp?itemId=item_1554689
Kelly RL (1992) Mobility/sedentism: concepts, archaeological measures, and effects. Annu Rev Anthropol 21(1):43–66. https://doi.org/10.1146/ANNUREV.AN.21.100192.000355
King T, Andrews P, Boz B (1999) Effect of taphonomic processes on dental microwear. Am J Phys Anthropol 108:359–373
Kuntz D, Costamagno S, Feyfant L, Martin F (2016) The exploitation of ungulates in the Magdalenian in the Entre-Deux-Mers (Gironde, France). Quatern Int 414:135–158. https://doi.org/10.1016/J.QUAINT.2015.12.079
Lam YM, Chen X, Pearson OM (1999) Intertaxonomic variability in patterns of bone density and the differential representation of bovid, cervid, and equid elements in the archaeological record. Am Antiq 64(2):343–362. https://doi.org/10.2307/2694283
Lucas G (2005) The archaeology of time. Routledge. https://www.routledge.com/The-Archaeology-of-Time/Lucas/p/book/9780415311984
Lucas G (2012) Understanding the archaeological record. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511845772
Luret M, Burke A, Bernaldo de Quiros F, Besse M (2020) El Castillo cave (Cantabria, Spain): archeozoological comparison between the Mousterian occupation level (unit 20) and the “Aurignacien de transition de type El Castillo” (unit 18). J Archaeol Sci Rep 31:102339. https://doi.org/10.1016/J.JASREP.2020.102339
Lyman R (1985) Bone frequencies: differential transport, in situ destruction, and the MGUI. J Archaeol Sci 12(3):221–236. https://doi.org/10.1016/0305-4403(85)90022-6
Lyman RL (1994) Vertebrate taphonomy. Cambridge University Press. https://doi.org/10.1017/CBO9781139878302
Marean CW, Assefa Z (1999) Zooarcheological evidence for the faunal exploitation behavior of Neandertals and early modern humans. Evolutionary Anthropology: Issues, News, and Reviews 8(1):22–37. https://doi.org/10.1002/(SICI)1520-6505(1999)8:1%3c22::AID-EVAN7%3e3.0.CO;2-F
Marín J, Rodríguez-Hidalgo A, Vallverdú J, Gómez de Soler B, Rivals F, Rabuñal JR, Pineda A, Chacón MG, Carbonell E, Saladié P (2019) Neanderthal logistic mobility during MIS3: zooarchaeological perspective of Abric Romaní level P (Spain). Quat Sci Rev 225. https://doi.org/10.1016/j.quascirev.2019.106033
Marín-Arroyo AB (2013) Palaeolithic human subsistence in Mount Carmel (Israel). A taphonomic assessment of Middle and Early Upper Palaeolithic faunal remains from Tabun, Skhul and el-Wad. Int J Osteoarchaeol 23(3):254–273. https://doi.org/10.1002/OA.1241
Martínez-Moreno J, de la Torre I, Mora Torcal R (2004) Methodological approach for understanding Middle Paleolithic settlement dynamics at la Roca dels Bous (Noguera, Catalunya, Northeast Spain) — Universitat Autònoma de Barcelona Research Portal. In: Conard JN (ed) Settlement Dynamics of the Middle Paleolithic and Middle Stone Age (1st ed., vol 2, pp 393–413). Kerns Verlag. https://portalrecerca.uab.cat/en/publications/methodological-approach-for-understanding-middle-paleolithic-sett-2
Mellars P (1996) The Neanderthal legacy : an archaeological perspective from western Europe. Princeton University Press, p 471
Metcalfe D, Jones KT (1988) A reconsideration of animal body-part utility indices. Am Antiq 53(3):486–504. https://doi.org/10.2307/281213
Mihlbachler MC, Rivals F, Solounias N, Semprebon GM (2011) Dietary change and evolution of horses in North America. Science 331:1178–1181
Moclán A, Huguet R, Márquez B, Laplana C, Galindo-Pellicena MÁ, García N, Blain HA, Álvarez-Lao DJ, Arsuaga JL, Pérez-González A, Baquedano E (2021) A Neanderthal hunting camp in the central system of the Iberian Peninsula: a zooarchaeological and taphonomic analysis of the Navalmaíllo Rock Shelter (Pinilla del Valle, Spain). Quatern Sci Rev 269:107142. https://doi.org/10.1016/J.QUASCIREV.2021.107142
Moncel MH, Rivals F (2015) On the question of short-term Neanderthal site occupations: Payre, France (MIS 8–7), and Taubach/Weimar, Germany (MIS 5). J Anthropol Res 67(1):47–75. https://doi.org/10.3998/JAR.0521004.0067.104
Moncel Mh, Daujeard C (2012) The variability of the Middle Palaeolithic on the right bank of the Middle Rhône Valley (southeast France): technical traditions or functional choices? Quat Int 247(1):103–124. https://doi.org/10.1016/J.QUAINT.2010.10.030
Morin E (2007) Fat composition and Nunamiut decision-making: a new look at the marrow and bone grease indices. J Archaeol Sci 34(1):69–82. https://doi.org/10.1016/J.JAS.2006.03.015
Morin E (2012) Reassessing paleolithic subsistence: the Neandertal and modern human foragers of Saint-Césaire. Cambridge University Press
Navazo M, Benito-Calvo A, Alonso-Alcalde R, Alonso P, de la Fuente H, Santamaría M, Santamaría C, Álvarez-Vena A, Arnold LJ, Iriarte-Chiapusso MJ, Demuro M, Lozano M, Ortiz JE, Torres T (2021) Late Neanderthal subsistence strategies and cultural traditions in the northern Iberia Peninsula: insights from Prado Vargas, Burgos Spain. Quat Sci Rev 254:106795. https://doi.org/10.1016/J.QUASCIREV.2021.106795
Navazo M, Díez Fernández-Lomana J, Torres T, Colina A, Ortíz J (2005) La cueva de Prado Vargas. Un yacimiento del Paleolítico Medio en el sur de la cordillera cantábrica | Signed in. In: Neandertales cantábricos, estado de la cuestión: actas de la reunión científica: celebrada en el Museo de Altamira los días 20–22 de octubre de, 2004 (pp. 151–166). Subdirección General de Publicaciones, Información y Documentación. https://www.scopus.com/record/display.uri?eid=2-s2.0-84996855217&origin=inward&txGid=c00b5f54dc7e7007070caad279a6466d&featureToggles=FEATURE_NEW_DOC_DETAILS_EXPORT:1
Navazo M, Santamaría C, Santamaría M (2022) Using cores as tools: use-wear analysis of Neanderthal recycling processes in Level 4 at Prado Vargas (Cornejo, Merindad de Sotoscueva, Burgos, Spain). Lithic Technology 1–19. https://doi.org/10.1080/01977261.2022.2108276
Niven L, Steele TE, Rendu W, Mallye JB, McPherron SP, Soressi M, Jaubert J, Hublin JJ (2012) Neandertal mobility and large-game hunting: the exploitation of reindeer during the Quina Mousterian at Chez-Pinaud Jonzac (Charente-Maritime, France). J Hum Evol 63(4):624–635. https://doi.org/10.1016/j.jhevol.2012.07.002
O’Connell J, Hawkes K, Blurton Jones N (1988) Hazda scavenging: implications for Plio-Pleistocenehominid subsistence. Curr Anthropol 29(2):356–363
Ortega AI, Merino M, Ángel M (2014) Pinilla del Valle Archaeological Sites View project Ojo Guareña View project. https://www.researchgate.net/publication/250916557
Pales L, Lambert C (1971) Atlas Osteologique Herbivores . Atlas osteologique pour servir a l’identification des mammiferes du Quaternaire. Editions du centre national de la recherche scientifique I - II. https://es.scribd.com/document/190047238/Pales-Lambert-1971-Atlas-Osteologique-Herbivores
Patou-Mathis M (2000) Neanderthal subsistence behaviours in Europe. Int J Osteoarchaeol 10:379–395
Pavao B, Stahl PW (1999) Structural density assays of leporid skeletal elements with implications for taphonomic, actualistic and archaeological research. J Archaeol Sci 26(1):53–66. https://doi.org/10.1006/JASC.1998.0299
Pérez L, Machado J, Sanchis A, Hernández CM, Mallol C, Galván B (2020) A high temporal resolution zooarchaeological approach to Neanderthal subsistence strategies on the Southeastern Iberian Peninsula: El Salt Stratigraphic Unit Xa (Alicante, Spain). In: Cascalheira J, Picin A (eds) Short-Term Occupations in Paleolithic Archaeology. Interdisciplinary Contributions to Archaeology, pp 237–289. Springer Nature. https://doi.org/10.1007/978-3-030-27403-0_10/TABLES/13
Pérez L, Sanchis A, Hernández CM, Galván B, Sala R, Mallol C (2017a) Hearths and bones: an experimental study to explore temporality in archaeological contexts based on taphonomical changes in burnt bones. J Archaeol Sci Rep 11:287–309. https://doi.org/10.1016/J.JASREP.2016.11.036
Pérez L, Sanchís Serra A, Hernández Gómez CM, Galván Santos B (2017b) Paleoecología de macromamíferos aplicada a los conjuntos zooarqueológicos de El Salt y el Abric del Pastor: (Alcoy, Alicante) - Dialnet. In: Sanchis Serra A, Pascual Benito L, Sarrión Montañana I (eds) Interaccions entre felins i humans: homenatge a Innocenci Sarrión Montañana, pp 327–353. https://dialnet.unirioja.es/servlet/articulo?codigo=6242918
Pérez-Barbería FJ, Pérez-Fernández E (2009) Identificación, biología y ecología del rebeco cantábrico. In: Pérez-Barbería FJ, Palacios B (eds) El Rebeco Cantábrico (Rupicapra pyrenaica parva). Conservación y Gestión de sus poblaciones, pp 26–69. Ministerio de Medio Ambiente, Medio Rural y Marino. Naturaleza y Parques Nacionales
Picin A, Cascalheira J (2020) Introduction to short-term occupations in palaeolithic archaeology. In: Picin A, Cascalheira J (eds) Short-term occupations in paleolithic archaeology. Interdisciplinary contributions to archaeology, pp 1–15. Springer, Cham. https://doi.org/10.1007/978-3-030-27403-0_1
Pickering TR, Egeland CP (2006) Experimental patterns of hammerstone percussion damage on bones: implications for inferences of carcass processing by humans. J Archaeol Sci 33(4):459–469. https://doi.org/10.1016/J.JAS.2005.09.001
Pickering TR, Marean CW, Domínguez-Rodrigo M (2003) Importance of limb bone shaft fragments in zooarchaeology: a response to “On in situ attrition and vertebrate body part profiles” (2002), by M.C. Stiner. J Archaeol Sci 30(11):1469–1482. https://doi.org/10.1016/S0305-4403(03)00042-6
Pike-Tay A, Cabrera Valdés V, de Quirós FB (1999) Seasonal variations of the Middle-Upper Paleolithic transition at El Castillo, Cueva Morín and El Pendo (Cantabria, Spain). J Hum Evol 36(3):283–317. https://doi.org/10.1006/JHEV.1998.0271
Ready E (2013) Neandertal foraging during the late Mousterian in the Pyrenees: new insights based on faunal remains from Gatzarria Cave. J Archaeol Sci 40(3):1568–1578. https://doi.org/10.1016/J.JAS.2012.10.021
Real Margalef C, Serra AS, Pérez JVMS, Martínez MÁB, Vilanova AE, Zilhão J, Bonilla VV (2019) Estudio de la fauna del nivel IV del Abrigo de la Quebrada y su aportación al conocimiento de la economía y el comportamiento humano en el Paleolítico medio de la vertiente Mediterránea Ibérica. SPAL - Revista de Prehistoria y Arqueología 28(28.2):17–49. https://doi.org/10.12795/SPAL.2019.I28.13
Rendu W (2010) Hunting behavior and Neanderthal adaptability in the Late Pleistocene site of Pech-de-l’Azé I. J Archaeol Sci 37(8):1798–1810. https://doi.org/10.1016/j.jas.2010.01.037
Rendu W (2022) Selection versus opportunism: a view from Neanderthal subsistence strategies. Updating Neanderthals: understanding behavioural complexity in the Late Middle Palaeolithic, 109–122. https://doi.org/10.1016/B978-0-12-821428-2.00013-5
Rendu W, Armand D (2009) Saisonnalité de prédation du Bison du gisement moustérien de la Quina (Gardes-le-Pontaroux, Charente), niveau 6c. Apport à la compréhension des comportements de subsistance. Bulletin de La Société Préhistorique Française 106(4):679–690. https://doi.org/10.3406/BSPF.2009.13890
Rendu W, Costamagno S, Meignen L, Soulier MC (2012) Monospecific faunal spectra in Mousterian contexts: implications for social behavior. Quatern Int 247(1):50–58. https://doi.org/10.1016/j.quaint.2011.01.022
Ríos Garaizar J, San Emeterio Gómez A, Iriarte E, LastNameLópez Bultó O, Arnold LJ, Bourguignon L, Iriarte Chiapusso MJ, Pérez Garrido C, LastNamePiqué R, Campaña Lozano I, Bermejo Albarrán L, Sánchez-Romero L, Demuro M, Marín Arroyo AB, Lahaye C, Aranburu Artano A, Arranz-Otaegui A, Cubas M, Benito-Calvo A, … Ortega Cordellat I (2020) El complejo arqueológico de Aranbaltza (Barrika, Bizkaia). Una ventana a las formas de vida de los neandertales fuera de las cuevas en la región cantábrica. In: Actualidad de la investigación arqueológica en España II (2019–2020). Conferencias impartidas en el Museo Arqueológico Nacional (pp. 47–62). MInisterio de Cultura y Deporte. https://cir.cenieh.es/handle/20.500.12136/2209
Rios-Garaizar J, Maidagan DG, Gómez-Olivencia A, Iriarte E, Arceredillo-Alonso D, Iriarte-Chiapusso MJ, Garcia-Ibaibarriaga N, García-Moreno A, Gutierrez-Zugasti I, Torres T, Aranburu A, Arriolabengoa M, Bailón S, Murelaga X, Ordiales A, Ortiz JE, Rofes J, Pedro ZS (2015) Short-term Neandertal occupations in the late Middle Pleistocene of Arlanpe (Lemoa, northern Iberian Peninsula). CR Palevol 14(3):233–244. https://doi.org/10.1016/J.CRPV.2014.11.006
Rivals F, Mihlbachler MC, Solounias N (2007) Effect of ontogenetic-age distribution in fossil samples on the interpretation of ungulate paleodiets using the mesowear method. J Vertebr Paleontol 27:763–767
Rivals F, Prignano L, Semprebon G, Lozano S (2015) A tool for determiningduration of mortality events in archaeological assemblages using extant ungulate microwear. Sci Rep 5:17330
Rodríguez Hidalgo A (2015) Dinámicas Subsistenciales durante el Pleistoceno Medio en la Sierra de Atapuerca: Los Conjuntos Arqueológicos de TD10.1 y TD10.2. TDX (Tesis Doctorals En Xarxa). http://www.tdx.cat/handle/10803/348869
Romandini M, Nannini N, Tagliacozzo A, Peresani M (2014) The ungulate assemblage from layer A9 at Grotta di Fumane, Italy: a zooarchaeological contribution to the reconstruction of Neanderthal ecology. Quatern Int 337:11–27. https://doi.org/10.1016/J.QUAINT.2014.03.027
Romero AJ, Carlos Díez J, Arceredillo D, García-Solano J, Jordá-Pardo JF (2019) Neanderthal communities in the heart of the Iberian Peninsula: taphonomic and zooarchaeological study of the Mousterian site of Jarama VI (Guadalajara, Spain). Archaeol Anthropol Sci 11(5):1713–1725. https://doi.org/10.1007/S12520-018-0625-7/TABLES/8
Rosell J (2001) Patrons d’aprofitament de les biomasses animals durant el Pleistocè inferior i mig (Sierra de Atapuerca, Burgos) i superior (Abric Romaní, Barcelona) - Dialnet. Universitat Rovira i Virgili
Rosell J, Blasco R, Fernández-Laso MC, Vaquero M, Carbonell E (2012a) Connecting areas: faunal refits as a diagnostic element to identify synchronicity in the Abric Romaní archaeological assemblages. Quatern Int 252:56–67. https://doi.org/10.1016/J.QUAINT.2011.02.019
Rosell J, Blasco R, Huguet R, Cáceres I, Saladié P, Rivals F, Bennàsar M, Bravo P, Campeny G, Esteban-Nadal M, Fernández-Laso C, Gabucio MJ, Ibáñez N, Martín P, Muñoz L, Rodríguez-Hidalgo A (2012b) Occupational patterns and subsistence strategies in level J of Abric Romaní. Vertebr Paleobiol Paleoanthropol 9789400739215:313–372. https://doi.org/10.1007/978-94-007-3922-2_8/COVER
Rosell J, Blasco R, Rivals F, Chacón MG, Arilla M, Camarós E, Rufà A, Sánchez-Hernández C, Picin A, Andrés M, Blain HA, López-García JM, Iriarte E, Cebrià A (2017) A resilient landscape at Teixoneres Cave (MIS 3; Moià, Barcelona, Spain): the Neanderthals as disrupting agent. Quatern Int 435:195–210. https://doi.org/10.1016/J.QUAINT.2015.11.077
Rusch L, Gregoire S, Pois V, Moigne AM (2019) Neanderthal and carnivore occupations in unit II from the Upper Pleistocene site of Ramandils Cave, (Port-la-Nouvelle, Aude, France). J Archaeol Sci Rep 28:102038. https://doi.org/10.1016/J.JASREP.2019.102038
Saladié P, Huguet R, Díez C, Rodríguez-Hidalgo A, Cáceres I, Vallverdú J, Rosell J, Bermúdez De Castro JM, Carbonell E (2011) Carcass transport decisions in Homo antecessor subsistence strategies. J Hum Evol 61(4):425–446. https://doi.org/10.1016/J.JHEVOL.2011.05.012
Saladié P, Rodríguez-Hidalgo A, Díez C, Martín-Rodríguez P, Carbonell E (2013) Range of bone modifications by human chewing. J Archaeol Sci 40(1):380–397. https://doi.org/10.1016/J.JAS.2012.08.002
Salazar-García DC, Power RC, Sanchis Serra A, Villaverde V, Walker MJ, Henry AG (2013) Neanderthal diets in central and southeastern Mediterranean Iberia. Quatern Int 318:3–18. https://doi.org/10.1016/J.QUAINT.2013.06.007
Sánchez-Hernández C, Gourichon L, Pubert E, Rendu W, Montes R, Rivals F (2019) Combined dental wear and cementum analyses in ungulates reveal the seasonality of Neanderthal occupations in Covalejos Cave (Northern Iberia). Sci Rep 9(1): 1–18. https://doi.org/10.1038/s41598-019-50719-7
Sánchez-Romero L, Benito-Calvo A, Marín-Arroyo AB, Agudo-Pérez L, Karampaglidis T, Rios-Garaizar J (2020) New insights for understanding spatial patterning and formation processes of the Neanderthal occupation in the Amalda I cave (Gipuzkoa, Spain). Sci Rep 10(1). https://doi.org/10.1038/s41598-020-65364-8
Sanchis A, Real C, Sauqué V, Núñez-Lahuerta C, Égüez N, Tormo C, Pérez Ripoll M, Carrión Marco Y, Duarte E, de la Rasilla M (2019) Neanderthal and carnivore activities at Llonin Cave, Asturias, northern Iberian Peninsula: faunal study of Mousterian levels (MIS 3). CR Palevol 18(1):113–141. https://doi.org/10.1016/J.CRPV.2018.06.001
Santamaría Cabornero C, Navazo Ruiz M, Benito-Calvo A (2021) Functional analysis of Middle Palaeolithic flint tools through experimental use wear analysis: the case of Prado Vargas (Cornejo) and Fuente Mudarra (Atapuerca), northern Spain. Munibe Antropologia-Arkeologia 72:5–17. https://doi.org/10.21630/maa.2021.72.05
Sañudo P, Blasco R, Fernández Peris J (2016) Site formation dynamics and human occupations at Bolomor Cave (Valencia, Spain): an archaeostratigraphic analysis of levels I to XII (100–200 ka). Quatern Int 417:94–104. https://doi.org/10.1016/J.QUAINT.2015.09.044
Sauqué V, Rabal-Garcés R, García-González R, Gisbert M (2015) Bucardos y osos fósiles del Pirineo: recuperación de restos paleontológicos, catalogación y datación de los yacimientos del Pleistoceno-Holoceno de las cavidades el Pirineo oscense. Lucas Mallada. Revista de Ciencias 17:247–289. https://dialnet.unirioja.es/servlet/articulo?codigo=5968974
Schmid E (1972) Atlas of animal bones. Elsevier Publishing Company. https://books.google.com/books/about/Atlas_of_Animal_Bones.html?hl=es&id=yaMYAAAAYAAJ
Selvaggio MM (1994) Carnivore tooth marks and stone tool butchery marks on scavenged bones: archaeological implications. J Hum Evol 27(1–3):215–228. https://doi.org/10.1006/JHEV.1994.1043
Semprebon GM, Godfrey LR, Solounias N, Sutherland MR, Jungers WL (2004) Can low-magnification stereomicroscopy reveal diet? J Hum Evol 47:115–144
Shipman P (1981) Life history of a fossil : an introduction to taphonomy and paleoecology. Harvard University Press
Shipman P, Rose J (1983) Early hominid hunting, butchering, and carcass-processing behaviors: approaches to the fossil record. J Anthropol Archaeol 2(1):57–98
Silver I (1969) La determinación de la edad en los animales domésticos. In: Brothwell D, Higgins J (eds) Ciencia en arqueología (pp. 229–239).
Solounias N, Semprebon G (2002) Advances in the reconstruction of ungulate ecomorphology with application to early fossil equids. Am Museum Novit 3366:1–49
Starkovich BM (2017) Paleolithic subsistence strategies and changes in site use at Klissoura Cave 1 (Peloponnese, Greece). J Hum Evol 111:63–84. https://doi.org/10.1016/J.JHEVOL.2017.04.005
Stiner MC (1990) The use of mortality patterns in archaeological studies of hominid predatory adaptations. J Anthropol Archaeol 9(4):305–351. https://doi.org/10.1016/0278-4165(90)90010-B
Stiner MC (1994) Honor among thieves : a zooarchaeological study of Neandertal ecology. Princeton University Press
Stiner MC (1998) Mortality analysis of Pleistocene bears and its paleoanthropological relevance. J Hum Evol 34:303–326
Urquijo JG, Estévez JJI, Garaizar JR, Bourguignon L, Ugarte P, Vinagre AT (2005) Excavaciones recientes en Axlor: movilidad y planificación de actividades en grupos de neandertales. Neandertales cantábricos, estado de la cuestión: actas de la reunión científica: celebrada en el Museo de Altamira los días 20–22 de octubre de 2004 / coord. por José Antonio Lasheras Corruchaga. Ramón Montes Barquín 2005:527–539
Utrilla P, Montes L, Blasco F, Torres T, Ortiz J E (2010) La cueva de Gabasa revisada 15 ahos después: Un cubil para las hienas y un cazadero para los Neandertales. Zona Arqueológica 13:376–389. https://www.researchgate.net/publication/264881119_La_cueva_de_Gabasa_revisada_15_ahos_despues_Un_cubil_para_las_hienas_y_un_cazadero_para_los_Neandertales?enrichId=rgreq-db4d1b0426d37cd961c1de934befcdf1-XXX&enrichSource=Y292ZXJQYWdlOzI2NDg4MTExOTtBUzoxMzk1MzI5NjM5NDY0OTZAMTQxMDI3ODY1OTA5MA%3D%3D&el=1_x_2&_esc=publicationCoverPdf
Uzquiano P, Yravedra J, Zapata BR, Gil Garcia MJ, Sesé C, Baena J (2012) Human behaviour and adaptations to MIS 3 environmental trends (>53–30 ka BP) at Esquilleu cave (Cantabria, northern Spain). Quatern Int 252:82–89. https://doi.org/10.1016/J.QUAINT.2011.07.023
Uzunidis A, Pineda A, Jiménez-Manchón S, Xafis A, Ollivier V, Rivals F (2021) The impact of sediment abrasion on tooth microwear analysis: an experimental study. Archaeol Anthropol Sci 13:134. https://doi.org/10.1007/s12520-021-01382-5
Valensi P, Crégut-Bonnoure E, Defleur A (2012) Archaeozoological data from the Mousterian level from Moula-Guercy (Ardèche, France) bearing cannibalised Neanderthal remains. Quatern Int 252:48–55. https://doi.org/10.1016/J.QUAINT.2011.07.028
Vallerdú J, Allué E, Bischoff JL, Cáceres I, Carbonell E, Cebrià A, García-Antón D, Huguet R, Ibáñez N, Martínez K, Pastó I, Rosell J, Saladíe P, Vaquero M (2005) Short human occupations in the Middle Palaeolithic level I of the Abric Romaní rock-shelter (Capellades, Barcelona, Spain). J Hum Evol 48(2):157–174. https://doi.org/10.1016/J.JHEVOL.2004.10.004
Vallverdú J, Alonso S, Bargalló A, Bartrolí R, Campeny G, Carrancho Á, Expósito I, Fontanals M, Gabucio J, Gómez B, Prats JM, Sañudo P, Solé À, Vilalta J, Carbonell E (2012) Combustion structures of archaeological level O and mousterian activity areas with use of fire at the Abric Romaní rockshelter (NE Iberian Peninsula). Quatern Int 247(1):313–324. https://doi.org/10.1016/J.QUAINT.2010.12.012
Vaquero M (1997) Tecnología lítica y comportamiento humano: organización de las actividades y cambio diacrónico en el Paleolitico Medio del Abric Romaní. Universitat Rovira Virgili
Vaquero M, Romagnoli F, Bargalló A, Chacón MG, Gómez de Soler B, Picin A, Carbonell E (2019) Lithic refitting and intrasite artifact transport: a view from the Middle Paleolithic. Archaeol Anthropol Sci 11(9):4491–4513. https://doi.org/10.1007/S12520-019-00832-5/FIGURES/15
Vettese D, Blasco R, Cáceres I, Gaudzinski-Windheuser S, Moncel M-H, Hohenstein UT, Daujeard C (2020) Towards an understanding of hominin marrow extraction strategies: a proposal for a percussion mark terminology. Archaeol Anthropol Sci 12(48). https://doi.org/10.1007/s12520-019-00972-8
Vettese D, Daujeard C, Blasco R, Borel A, Caceres I, Moncel MH (2017) Neandertal long bone breakage process: standardized or random patterns? The example of Abri du Maras (Southeastern France, MIS 3). J Archaeol Sci Rep 13:151–163. https://doi.org/10.1016/J.JASREP.2017.03.029
Villa P, Mahieu E (1991) Breakage patterns of human long bones. J Hum Evol 21(1):27–48. https://doi.org/10.1016/0047-2484(91)90034-S
Villaverde V, Eixea A, Zilhão J, Sanchis A, Real C, Bergadà M (2017) Diachronic variation in the Middle Paleolithic settlement of Abrigo de la Quebrada (Chelva, Spain). Quatern Int 435:164–179. https://doi.org/10.1016/J.QUAINT.2015.09.075
Voorhies MR (1969) Taphonomy and population dynamics of an Early Pliocene vertebrate fauna, Knox County, Nebraska. Taphonomy and population dynamics of an Early Pliocene vertebrate fauna, Knox County, Nebraska. https://doi.org/10.2113/GSROCKY.8.SPECIAL_PAPER_1.1
Wallace IJ, Shea JJ (2006) Mobility patterns and core technologies in the Middle Paleolithic of the Levant. J Archaeol Sci 33(9):1293–1309. https://doi.org/10.1016/J.JAS.2006.01.005
Young A, Stillman R, Smith MJ, Korstjens AH (2015) Scavenger species-typical alteration to bone: using bite mark dimensions to identify scavengers. J Forensic Sci 60(6):1426–1435. https://doi.org/10.1111/1556-4029.12839
Yravedra Sainz de los Terreros J, Gómez Castanedo A (2011) Análisis de los procesos tafonómicos de Cueva Morín. Primeros resultados de un estudio necesario. Zephyrus: Revista de Prehistoria y Arqueología 67:69–90
Acknowledgements
We are grateful for the financial support received from the Junta de Castilla y León, through the Consejería de Cultura y Turismo, Dirección General de Patrimonio Cultural, which funded the project “Intervención arqueológica en la Cueva de Prado Vargas, in Cornejo” (Merindad de Sotoscueva, Burgos) (SUBV-22/011-BU). We also want to thank Beni and the mayors of La Merindad de Sotoscueva and Cornejo for their support and collaboration. The same goes for all of those who participated and participate in excavation campaigns, without whom we would not have been able to do this work. Finally, a special thanks to the Prehistory Laboratory of the UBU partners, Marta Santamaría and Pedro Alonso, for their immeasurable and indispensable support.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Contributions
Conceptualization: HFJ, MN, ABC, FR, MA. Methodology: HFJ, FR, MA. Formal analysis and investigation: HFJ, FR, MA. Writing—original draft preparation: HFJ, FR, MA. Writing—review and editing: HFJ, MN, ABC, FR, MA. Zooarcheological and taphonomical analysis: HFJ. Mesowear and microwear analysis: FR. Statistical analysis: HFJ, MA. Resources: HFJ, MN, ABC, FR, MA. Supervision HFJ.
All the authors reviewed the manuscript and approved the version to be published.
Corresponding author
Ethics declarations
Ethical approval
Not applicable.
Competing interests
The authors declare no competing interests.
Consent to participate
All authors have approved the manuscript.
Consent for publication
All authors agree with the submission to the Archaeological and Anthropological Sciences.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
de la Fuente Juez, H., Navazo, M., Benito-Calvo, A. et al. Too good to go? Neanderthal subsistence strategies at Prado Vargas Cave (Burgos, Spain). Archaeol Anthropol Sci 15, 164 (2023). https://doi.org/10.1007/s12520-023-01857-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-023-01857-7