
Abstract
In this paper, we analyse the fish and small mammal assemblages from the Middle Paleolithic horizons of Hohle Fels Cave to reconstruct the paleoclimatic and paleoenvironmental conditions faced by the Neanderthal groups who occupied the site. The fish assemblage indicates that the freshwater ecosystem around this site was characterized by a pre-mountain river system with the presence of permanent, oxygen-rich, and cold running waters. The results of the Habitat Weighting Method and the Bioclimatic Model applied to the small mammal assemblage, coupled with the new dates obtained for the Archaeological Horizon (AH) IX, allow us to identify two different climatic phases. One phase (AH X-XII) is more temperate possibly corresponding to the end of Marine Isotope Stage (MIS) 5; the other (AH VI-IX) is colder and more arid corresponding to the end of MIS 4 through the beginning of MIS 3. Open environments with a relatively stable forest component dominated the landscape during this part of the Middle Paleolithic. These two climatic phases appear to correspond to different levels of occupational intensity by Neanderthals, with higher occupational intensity during mild climatic periods and lower intensity during cold, arid periods. Our climatic reconstruction and recent absolute dating, together with the recovery of a complete leaf point near the top of AH X, have important implications for the cultural stratigraphy and cultural chronology of the region. Archaeologist have traditionally viewed leaf points as key artefacts of the Blattspitzengruppe, a cultural complex attributed to the end of Middle Paleolithic; however, this stratigraphic, climatic, and chronological context indicates the need to revise this interpretation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Swabian Jura (South-western Germany) is a key region for Paleolithic studies, in particular of the transition between the Middle and Upper Paleolithic and the evolution of the Upper Paleolithic industries (Aurignacian, Gravettian, Magdalenian). The caves in this region (Fig. 1) have been the subject of intensive research for the last 150 years and produced an astonishing archaeological record that helped develop and improve our understanding of past lifeways, subsistence strategies, and technologies of both Neanderthal and modern human groups. The Ach and the Lone Valleys are located along two tributaries of the Danube in the Swabian Jura. The caves and shelters that can be found along these valleys, such as in Hohle Fels, Geißenklösterle, Vogelherd, and Hohlenstein-Stadel (Conard et al. 2009; Conard and Kind 2017), are renowned for the musical instruments and the ivory figurines that have been recovered there. The study of subsistence strategies during the Middle and Upper Paleolithic provides useful insights into the behaviour of human groups (Barth et al. 2009; Conard et al. 2013; Kitagawa 2014; Wong et al. 2020b; Bertacchi et al. 2021, among others) and recently the potential of small vertebrate studies for the reconstruction of the climatic and environmental conditions faced by those groups was highlighted by the publications of the small mammal assemblage at Hohle Fels (Rhodes et al. 2019), Geißenklösterle (Rhodes et al. 2018), and Langmahdhalde (Wong et al. 2020a).

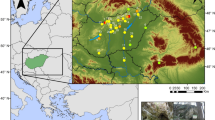
Location of Hohle Fels Cave and other sites mentioned in the discussion. 1, Gironde; 2, Charente-Maritimes and Charente; 3, Scladina and Marie-Jeanne Caves; 4, Sesselfelsgrotte; 5, Biśnik and Koziarnia Caves; 6, Obłazowa Cave. Detailed map of the Swabian Jura modified after Sommer 2019
In this paper, we propose a new climatic and environmental reconstruction of the conditions faced by Neanderthal groups during their occupations of Hohle Fels Cave and try to understand how these conditions might have influenced their strategies and occupation patterns.
The site
Hohle Fels Cave (48° 22′ 45″ N; E9° 45′14″ E) is located in the Ach River valley (Fig. 1), at 564 m a.s.l. and ca. 7 m above the current floor of the valley. The first scientific investigation of the cave led by Fraas and Hartmann started in 1870–1871 and yielded faunal remains (mostly cave bears), lithic tools, and bone and antler artifacts (Hahn 1978). After that, Schmidt (1912) published the results of his research conducted in 1906 in the book Die Diluviale Vorzeit Deutschland. Between 1958 and 1960, further excavations were carried out by Matschak and Riek, and between 1977 and 1979 by Hahn (Hahn 1978, 1979). This last effort provided proof of a well-preserved stratigraphy an abundance of artifacts (Hahn 1979). Systematic investigation of Hohle Fels started in 1988 under the direction of Hahn (until his death in 1997) and has continued every year from 1997 until the present under the direction of Conard. The excavations exposed a ca 5.5 m-thick sequence, spanning from the Middle Paleolithic to the Magdalenian (Fig. 2). In the past 25 years of researches, the site yielded abundant lithic and faunal remains, but it is especially renowned for the symbolic artifacts including ivory animal and human figurines, personal ornaments, and musical instruments recovered in the Aurignacian levels (Conard 2009; Conard et al. 2009; Wolf and Conard. 2015; Velliky et al. 2021, among others). The material analysed in this study is derived from the horizons assigned to Middle Paleolithic, i.e., Archeological Horizons (AH) VI to XII, which correspond to Geological Horizons (GH) 9–15, excavated between 2014 and 2021 (Fig. 2). The description of the GHs follows Miller (2015) and the field notes from the excavation campaigns.
-
GH 9: ca. 30 cm thick. The sediment is brown (7.5 YR 5/6) with yellowish-brown layers alternating with reddish-brown layers. It is composed of silt, with little clay and some lime sand. Limestone blocks make up 20–30% of the units, are sub-rounded, and range in size between 8 and 25 cm. The transition to GH 10 appears gradually over 4 cm.
-
GH 10: ca. 30 cm thick. The sediment is brown (7.5 YR 6/4–5/4), has a relatively high proportion of calcite sand and is composed mostly of silt. Limestone blocks are less present than in GH 9 and are angular to slightly rounded. The passage to GH 11 is clear in the east part, over 2 cm, and more gradual in the west part, over 4 cm.
-
GH 11: ca. 10–20 cm. The sediment is yellowish-brown (7.5 YR 6/4–5/6) and composed of silty clay. Limestone blocks are smaller, 2–15 cm in size. Gradual transition to GH 12.
-
GH 12: the sediment is greenish-brown (10 YR 5/4), composed of clayey silt with a high proportion of coarse limestone sand. Limestone blocks are small, 2–8 cm in size.
-
GH 13: the sediment is yellow–brown, darker, and almost reddish in the lower part (10 YR 5/8 upper, 10 YR 4/6 lower). It is composed of silty clay with a relatively high proportion of coarse and fine lime sand. Irregular limestone rubbles up to 20 cm. Transition to GH 14 is gradual but visible.
-
GH 14: The sediment is yellow–brown, almost reddish (10 YR 4/6). It is composed of silty clay with a lot of coarse and fine limestone sand. In its lower portion, the sediment is yellowish-brown (10YR 5/6) and composed of clayey silt. Limestone blocks up to 30 cm. GH 14.1 is a feature characterized by grey-brown sediment (10YR 5/4) and unusually high find density. Transition to GH 15 is very clear, sometimes over a few millimetres.
-
GH 15: the sediment is yellowish-brown (10 YR 5/4), lighter colour in the northern Sect. (10 YR 5/6). It is composed of clayey silt with a high proportion of limestone sand. High amount of limestone clasts, 50–80%. GH 15.2 is a feature at the contact with GH 14.1 that presents lighter colour (10 YR 5/6) and a high density of bones and small mammal bones.

Profile 2 of Hohle Fels cave, modified after Conard and Janas 2021. Graphic by A. Janas
GH 9 to 11 are dated between 35 and 40 ka years BP, corresponding to ~ 39–44 ka cal BP (OxCal 4.4, IntCal 20, Bronk Ramsey 2009) and clearly represent minimum ages (Conard and Bolus 2008; Conard 2009; Higham et al. 2012), while new ESR dates for GH 12 put it at 62 ± 4 ka (Conard et al. 2021).
One previous study explored the presence of fish remains in Hohle Fels from the Middle Paleolithic layers and indicated the presence of burbot (Lota lota), grayling (Thymallus thymallus), and unidentifiable cyprinids, all of which correspond to species of fish commonly found in the Danube and its tributaries such as the Ach River (Conard et al. 2013). In this study, the authors reported the common presence of fish bones and scales in the Aurignacian and Gravettian deposits but suggested this presence to be only sporadic in the Middle Paleolithic layers. In the present paper, the sample was enlarged to more than 3400 remains in contrast to the original study that included only 19 specimens from the upper portion of the Middle Paleolithic deposits. Additionally, we present a detailed paleoenvironmental study based on the fish species recovered from the site.
Recent studies (Rhodes et al. 2019; Rhodes and Conard 2021) analysed the small mammal assemblages from the Middle Paleolithic GHs available at the time (GH 9–12) and compared them to the early Upper Paleolithic (i.e. Aurignacian) in order to explore the climatic and environmental changes during the Middle to Upper Paleolithic transition. The paleoclimatic reconstruction based on the rodent assemblage describes GH 12 as the less cold and arid layer in the Middle Paleolithic sequence and indicates a decrease in temperature and precipitation in GH 9–11 (Rhodes and Conard 2021). The landscape surrounding the site was a mosaic of arctic tundra and boreal and deciduous forests, with a steppe component always present but fluctuating through time (Rhodes 2019; Rhodes et al. 2019). In this study, we increased the samples for the Middle Paleolithic GHs already published and add samples from the new Middle Paleolithic GHs 13, 14, and 15 excavated in 2020 and 2021.
Material and methods
Small vertebrate material was recovered through water screening using 0.5-mm sieves. The material analysed comes from one square meter excavation units 11, 25, 26, 27, and 28. These are the squares that currently present the whole Middle Paleolithic sequence, going from GH 9 to 15 (Fig. 2). For each GH, we present the number of identified specimens (NISP) and the minimum number of individuals (MNI).
Fishes
We analysed this assemblage using a Zeiss Stemi 305 stereomicroscope and photographed the material using a Dino-Lite Edge Digital Microscope. Anatomical and taxonomical classifications were done by comparison method of external characteristics of those remains using the reference collection hosted in Senckenberg Centre for Human Evolution and Palaeoenvironment (University of Tübingen) and several osteological atlases (Lepiksaar 1994; Watt et al. 1997; Conroy et al. 2005). The taxonomic nomenclature used has been extracted from Cannon (1987) and Wheeler and Jones (2009).
Small mammals
The identification of the small mammal was based on the best diagnostic elements available: for lagomorphs and rodents, mandibles, maxilla, and isolated teeth; for shrews, mandibles, and maxilla; for moles and bats, mandibles, maxilla, isolated teeth, and postcranial bones were analysed. The criteria for the identification of the material, the nomenclature for morphological descriptions, and the measurements taken follow Niethammer and Krapp (1978; 1982), Nadachowski (1982), and Borodin and Markova (2015) for Rodentia; Laplana et al. (2015) for Lagomorpha; and Niethammer and Krapp (1990) for Eulipothyphla; Sevilla García (1988) for Chiroptera. Comparison of the fossil specimens to the osteological collection of the University of Tübingen was also crucial for the identification. Taxonomic nomenclature follows Wilson and Reeder (2005), except for Clethrionomys glareolus (see Tesakov et al., 2010; Kryštufek et al., 2019), Alexandromys oeconomus (Lissovsky et al. 2018, among others), and Lasiopodomys anglicus (see Petrova et al., 2015; Baca et al., 2019). Photos of the material have been taken using a Zeiss Stereo Discovery V8.
Morphometric analysis has been conducted on complete first lower molars of adult specimens of Arvicola amphibius. The Schmelzband-Differenzierung-Quotient or SDQ index (Heinrich 1978) quantifies the difference in enamel thickness between the anterior and the posterior wall of each triangle of the tooth (Fig. 3).

Scheme for the measurement of the Schmelzband-Differenzierung-Quotient (SDQ) index in first lower molar of Arvicola amphibius
To calculate the SDQ value of a population, it is necessary to obtain the mean SDQ for each m1 (comprehending all the triangles (T1 to T5) and the two sides of PL)
and then the mean of the SDQ of all (n) m1 taken into account
Photos of the specimens have been taken using a Leica EZ4HD microscope. Measurements have been taken on the photos using ImageJ software. Data have been analysed using PAST software (Hammer et al. 2001).
Paleoenvironmental reconstruction
The habitat weighting method (Evans et al. 1981; Andrews 2006) based on small mammals has been applied in order to obtain a reconstruction of the landscape and environment surrounding the site. Each mammal species has been assigned to the habitat(s) where it can be currently found. We decided to use two different sets of values, one based on ecological preferences (Suppl. Material 1), and one based on habitat suitability (Suppl. Material 2).
For ecological preferences, six categories have been identified (following López-García et al. 2014, 2017a, 2017b): open dry grassland and steppe (OD), open humid evergreen meadows (OH), open woodland with moderate ground cover, woodland margins, and forest patches (OW), woodland, or mature forest (Wo), rocky environment (R), and water-related environments (Wa).
For habitat suitability, we follow the attributions of the IUNC Red List (IUCN 2021): “forest” including forested and wooded areas, and forest margins; “shrubland” intended as shrubs, bushland and thickets; “grassland,” characterized by grasses and broadleaf herbaceous plants with sparsely distributed or absent woody plants; “wetland”; and “desert.” Since most of the species are no longer present in Southern Germany, we use the attribution(s) given in the global distribution for each species.
We use Spermophilus citellus as current reference for S. superciliosus and Cricetulus migratorius as current reference for Allocricetus vel Cricetulus sp.
Paleoclimatic reconstruction
Paleoclimatic data have been calculated for each GH based on the insectivore and rodent assemblage, using the Bioclimatic model described in Hernández Fernández (2001, 2005) and recently updated by Royer et al. (2020). The insectivores and rodents’ assemblages have been analysed using the Climatic Restriction Index (CRIi = 1/n, where “n” is the number of climatic zones where the species are represented and “i” is the climatic zone where the species appears). Starting from the CRI, the Bioclimatic Component has been calculated (BC; representation by the level of each of the available climatic zones): BCi = (ΣCRIi) X 100/S (where S is the number of species per GH). From the BC, it was possible to estimate climatic parameters by means of multiple linear regression method (Hernández Fernández and Peláez-Campomanes 2005; Royer et al. 2020), using the R script file published by Royer et al. (2020). For this study, the mean annual temperature (MAT expressed in °C) and mean annual precipitations (P expressed in mm) are calculated. Current data for the town of Schelklingen (Fig. 1) show a MAT of 8.3ºC and P of 1059 mm (climate-data.org). Talpa sp. has been considered as T. europaea, being the only species for this genus present at Hohle Fels. Sorex sp., Neomys sp., and Sicista sp. have not been included in the calculation. As for the environmental reconstructions, we use Spermophilus citellus as current reference for S. superciliosus and Cricetulus migratorius as current reference for Allocricetus vel Cricetulus sp.
Results and discussion
Taphonomic analysis: preliminary results
Fishes
Anthropogenic marks such as burned bones or cut marks and non-anthropogenic damage indicators on the bone surface of the fish that remain such as digestion marks, compression, uniaxial mechanical deformation, or bite marks were analysed. Previous studies have distinguished several categories of digestion marks, depending on the intensity of digestion (Andrews 1990; Guillaud et al. 2018; Frontini et al. 2021; Blanco-Lapaz et al. 2021). The presence of bite marks or crushed bones can give us insight into the gnawing activities carried out by terrestrial carnivores such as foxes or wolves (Nicholson 1993) while digestive marks are indicative of other carnivorous birds (Andrews 1990).
Taphonomic analysis of the fish assemblage from the Middle Paleolithic horizons of Hohle Fels indicates no damages such as mechanical deformation, rounding, or polishing due to compression during the digestion process (Frontini et al. 2021; Blanco-Lapaz et al. 2021). Bite marks, crush marks, or digestive marks were absent from all the fish remains, which means that carnivores and birds can be ruled out as accumulating agents (Nicholson 1993). No direct anthropogenic marks that indicate burning or butchery activities were documented.
The lack of carnivore modifications on fish bones likely indicates that carnivores, which visited the cave, including red/arctic foxes, wolf, cave/brown bears, hyenas, and birds of prey (Conard et al. 2013; Rhodes et al. 2019; Baumann et al. 2020; Beattie et al. 2021), were not the primary agents of accumulation for the fish remains, leaving humans as most probable accumulation agents. On the other hand, we cannot yet definitively prove that the fish remains were accumulated by humans since there is no direct evidence of anthropogenic modifications such as cut marks or burned bones on the assemblage. Further, more detailed taphonomic studies (ongoing) might provide more data and help identify the main agent of the fish accumulation during the Middle Paleolithic at Hohle Fels.
Small mammals
A comprehensive taphonomic analysis of small mammals from GH 9 to 12 has been published by Rhodes et al. 2019) and suggests that the small mammal material was most likely accumulated by the snowy owl Bubo scandiacus, the Eurasian eagle-owl Bubo bubo, and/or the great grey owl Strix nebulosa, with a limited contribution by foxes. As these predators are mostly generalist hunters, the composition of the assemblages should accurately reflect the diversity of the small mammal community on the landscape at the time of the accumulation.
In this study, a total of 2093 first lower molars of Arvicolids from GH 13 to 15 have been analysed according to the criteria described in Fernández-Jalvo et al. (2016), observing the degree and frequency of digestion signs, in order to determine the principal agents of the small mammal accumulation (Table 1). Due to the high percentage of teeth with absent to moderate modifications, this preliminary study indicates that predators of category 1 or 2 are most likely responsible for the accumulation. Nocturnal birds of prey, such as Tyto alba, Asio flammeus, and Asio otus (cat. 1) or Bubo scandiacus and Strix nebulosa (cat.2) are listed as possible accumulators (Fernández-Jalvo et al. 2016). Nevertheless, the low percentage of teeth with heavy or extreme digestion signs might indicate that small carnivores (e.g., Mustela spp. or Vulpes spp.) or diurnal birds of prey (such as Falco spp.) also played a role in the formation of the small mammal assemblages. An ongoing study regarding the bird assemblage of Hohle Fels already identified four species of birds of prey from GH 13: Falco tinnunculus, Corvus corax, Asio flammeus, and Bubo bubo (Beattie et al. 2021). These preliminary results confirm that opportunistic hunters (Andrews 1990) are the principal responsible for the small mammal accumulation that should therefore accurately reflect the composition of the small mammal community on the landscape surrounding the site.
Paleoclimatic and paleoenvironmental reconstructions for Neanderthal occupations at Hohle Fels Cave
Fishes
A total of 3482 fish remains, belonging to 8 species, were analysed (Table 2, Fig. 4). The Middle Paleolithic fish assemblage from Hohle Fels comprises five families: Salmonidae, Lotidae, Cyprinidae, Cottidae, and Esocidae. The predominant family recovered is Salmonidae, represented by three species, the European grayling (Thymallus thymallus, NISP = 1381, MNI = 15), the brown trout (Salmo trutta, NISP = 3, MNI = 2), and the Danubian trout (Hucho hucho, NISP = 2, MNI = 1). Lotidae, represented by the burbot (Lota lota, NISP = 1112, MNI = 11), is the second most common family in the assemblage. The third most represented family is Cottidae, with one species, the bullhead (Cottus gobio, NISP = 73, MNI = 10). Two species belonging to the carp family, Cyprinidae, have been identified: the common chub (Leuciscus cephalus, NISP = 6, NMI = 5) and the common nase (Chondrostoma nasus, NISP = 2, NMI = 1). Esocidae is the least represented family classified in the assemblage with one specimen identified as the northern pike (Esox lucius, NISP = 1, MNI = 1).

A Precaudal vertebra. Pike (Esox lucius), HF21 25 3727 15 XII; B Left praeoeperculum fragment. Bullhead (Cottus gobio), HF21 25 3786.1 15 XII; C Left praemaxilar. Burbot (Lota lota), HF21 25 3786.2 15 XII; D Caudal vertebra. Burbot (Lota lota), HF21 25 3786.3 15 XII; E Precaudal vertebra. Danuvian trout (Hucho hucho), HF21 25 3551 14 XI; F. Precaudal vertebra. Brown trout (Salmo trutta), HF17 26 1633 12 IX; G Precaudal vertebra. Grayling (Thymallus thymallus), HF21 25 3785.1 15 XII; H Scale. Grayling (Thymallus thymallus), HF21 25 3785.2 15 XII; I1 Left pharyngeal arch fragment. Common nase (Chondrostoma nasus), HF21 25 3316.1 14 XI; I2 pharyngeal tooth fragment. Common nase (Chondrostoma nasus), HF21 25 3316.2 14 XI; J Left pharyngeal arch fragment. Common chub (Leuciscus cephalus), HF21 11 2411 14 XI. Scale 2 mm
As mentioned above, the Salmonidae family is represented by three different species. The European grayling (T. thymallus), which is present throughout the sequence, is native to the northern parts of the Palearctic and Nearctic ecozones, ranging from the UK and northern Europe across Eurasia to Siberia. These fishes require cool, well-oxygenated water, preferably with a swift current; they are found in large, sandy- or gravel-bottomed rivers and lakes, but T. thymallus may occasionally be found in brackish conditions. Generally omnivorous, they feed primarily on crustaceans, insects, and zooplankton. As they are highly sensitive to changes in water quality, Thymallus fishes may be considered indicator species. This species can reach a maximum weight of 1.5 kg and a maximum size of 50 cm (Füllner et al. 2016). The brown trout (S. trutta), which has only been found in layers GH 9 and 12, requires oxygen-rich water and a specific temperature for hatching and growing. Nowadays, S. trutta is usually found in cold running water with temperatures ranging from 0 to 20ºC (Jonsson and Jonsson 2009; Elliott and Elliott 2010). The Danubian trout (H. hucho), only present in GH 10 and 14, is endemic to the Danube basin in Europe and reaches about 1.5 m in length and more than 50 kg in weight, although the average length is between 60 to 120 cm. This permanent cold-water salmonid spawns in April, when water reaches a temperature of 6 to 9 °C (Füllner et al. 2016).
The burbot (Lota lota), the only gadiform (cod-like) freshwater species, is present in the Hohle Fels assemblage throughout the sequence. This species has a circumpolar distribution above 40°N. Populations are continuous from the British Isles across Europe and Asia to the Bering Strait. Burbots live in large, cold rivers, lakes, and reservoirs, primarily preferring freshwater habitats, but can thrive in brackish environments for spawning. During the summer, they are typically found in the colder water below the thermocline. As benthic fish, they tolerate an array of substrate types, including mud, sand, rubble, boulder, silt, and gravel for feeding. Burbot populations are fluvial during the winter months, and they migrate to near-shore reefs and shoals to spawn on the grounds of sand or gravel. As adults, they are primarily piscivores, preying on lamprey, whitefish, young northern pike, suckers, bullhead, and stickleback. This species can reach a maximum weight of 4 kg and a maximum size of 80 cm (Füllner et al., 2016).
The European bullhead (C. gobio) is found in Hohle Fels from GH 12 to 15 and is missing in the rest of the layers (GH 9–11). The European bullhead is a small demersal fish and prefers cold, clear, fast-flowing small streams and middle-sized rivers. It occurs on gravelly shores of cold lakes as well. The diet of this species is typically based on insects, crustaceans, and other invertebrates and it breeds in the spring. Some individuals can measure up to 15 cm although they are usually about 6 cm in length. This species is also a common prey of the burbot and other carnivorous fishes (Füllner et al., 2016).
The two species of cyprinids recovered from Hohle Fels are more typical of temperate waters and live in waters with temperatures between 10 and 25 °C (Doadrio 2002; Tissot and Souchon 2010). In Hohle Fels, the common chub (L. cephalus) is present in GH 12–14. This species is distributed throughout most of northern Eurasia, can reach lengths up to 60 cm, can weigh between 2 and 4 kg, and reach ages of 15–16 years (Füllner et al., 2016). The common nase (C. nasus) is only present in GH 14 and the individuals of this species are naturally found in drainages of the Black Sea (e.g. Danube), the southern Baltic Sea, and the southern North Sea. They range from 25 to 40 cm in length and weigh about 1 kg (Füllner et al., 2016). Accordingly, cyprinids such as the genus Leuciscus or Chondrostoma can be found in salmonid zones, as they also thrive in running water. However, cyprinids in general are present in multiple environments and show enormous diversity in their diet, which includes arthropods and other fishes (Doadrio, 2002).
The least represented species classified at Hohle Fels corresponds to the northern pike (E. lucius), represented in the assemblage by one caudal vertebra in GH 15. This species is carnivorous and typical of the fresh waters of the Northern Hemisphere. The northern pike has an average length of 50 cm but it can measure up to 150 cm and weigh 25 kg (Füllner et al., 2016).
No relevant environment changes are observed during the Hohle Fels sequence based on the fish remains since the majority of the identified species, such as the European grayling, the brown trout, the Danubian trout, the burbot, the bullhead, and the pike are common cold-water species (Füllner et al., 2016). The presence of these species characterizes the fish assemblages as belonging to a pre-mountain river system with the presence of permanent, oxygen-rich, and cold running waters. Only the common chub and nase are associated with habitat with temperate water but can also live in cold waters (Doadrio, 2002).
Small mammals
A total of 6538 small mammal remains, corresponding to a MNI of 2310, were identified. Six insectivores, one bat (only identifiable at genus level as Myotis sp.), one small lagomorph (Ochotona pusilla), and 14 rodents are represented at Hohle Fels (Table 3, Fig. 5). A few taxonomical remarks: the red-toothed shrews, with intermediate size between Sorex ex gr. araneus and S. minutus, a slightly posterior position of the mandibular foramen and a somewhat slender condylar articulation (Fig. 5B) has been assigned to S. tundrensis, following van Kolfschoten (2014); Neomys fodiens and N. anomalus (Fig. 5D and 5E) have been separated following the metric criteria in Popov and Zidarova (2008); Sicista cf. subtilis and S. cf. betulina (Fig. 5T and 5U) have been separated following the morphological and metric criteria of Niethammer and Krapp (1982); the small size hamsters (Fig. 5S) have been assigned to Allocricetus vel Cricetulus sp. due to the low number of specimens which does not allow the morphological and morphometric population study necessary to discriminate between the two genera (Hir 1993; Cuenca Bescós 2003).

A Sorex ex gr. araneus, condylar process and right mandible, HF_25-GH12-154; B Sorex tundrensis, condylar process and right mandible, HF_25-GH12-155; C: Sorex minutus, left mandible and condylar process, HF_28d_GH13-432; D Neomys fodiens, condylar process and right mandible, HF_28c-GH15-n4001; E Neomys anomalus, condylar process and right mandible, HF_27a-GH15-n3434; F Talpa europaea, left humerus, HF_27b-GH15-n3411. Scale 2 mm. G Microtus arvalis, right m1, HF_27b-GH15-672; H: Chionomys nivalis, right m1, HF_27b-GH9-345; I Alexandromys oeconomus, right m1, HF_26a-GH15-115; J Lasiopodomys anglicus, right m1, HF_27b-GH9-355; K Lasiopodomys anglicus, right m1, HF_27b-GH9-356; L Lasiopodomys anglicus, left m1, HF_27b-GH9-357; M Ochotona pusilla, left mandible, lateral and occlusal view, HF_26a-GH14-n1887; N Dicrostonyx torquatus, right m1, HF_26a-GH15-117; O Lemmus lemmus, right m1, HF_25-GH12-77; P Arvicola amphibius, left m1, HF_27a-GH15-25; Q Clethrionomys glareolus, left M2, occlusal and lateral view, HF_27b-GH9-316; R Apodemus cf sylvaticus, right m1, HF_27a-GH14-505; S Allocricetus vel Cricetulus sp., left m1, m2, m3, HF_25d-GH15-329; T Sicista cf. subtilis, left m1, HF_27a-GH14-506; U Sicista cf. betulina, left m1, HF_28-GH13-446; V Spermophilus cf. superciliosus, right M3, occlusal and lateral view, HF_25d-GH15-42; W Allactaga cf. major, right m1, HF_25c-GH 14.1–187. Scale 1 ms
To reconstruct the environment and landscape in the surrounding of the site, we decided to apply the Habitat Weighting Method with two different sets of values, one indicating the ecological preferences of each species, to obtain a general view of the changes in the environment while the other is based on habitat suitability, which allows us to recognize oscillation in the sequence by including different categories and dividing them further into sub-habitats.
We also decided to give particular weight to the changes in the relative frequency of the two lemming species present at Hohle Fels, Lemmus lemmus and Dicrostonyx torquatus. These two species are in fact strongly linked to subarctic and tundra environments (Wilson et al. 2017; IUCN 2021). They are easily identifiable so that they cannot be confused with other arvicolines in the sequence (Nadachowski 1982) and are therefore good indicators of cooling periods. A summary of the results of the environmental analysis is given in Fig. 6.

Paleoenvironmental reconstruction for the Middle Paleolithic Horizons of Hohle Fels. GH: geological horizons; OD: open dry; OH: open humid; Wa: water-related environment. % of Lemmus + Dicrostonyx indicates subarctic and tundra environments
Based on the percentage of lemmings (Table 3, Fig. 6), GH 14 seems to represent a more temperate period than GH 15. Conditions got slightly cooler and then colder respectively in GH 13 and 12. During GH 11 and 10 it is possible to observe an amelioration but at GH 9 there seems to be a recrudescence of the colder climate. The results of Habitat Weighting based on ecological preferences describe four different phases (Table 4). GH 15 to 13 represent a humid phase, with dominating open humid meadows (OH) and water-related environments (Wa). GH 12 and 11 represent a more arid phase, with an increase in open dry grassland (OD) and open woodland (OW) and a decrease in Wa and OH. Elements of mature forest (Wo) slightly decrease in this phase. In these two GHs, the relative percentage of Microtus arvalis reached its maximum, alongside a decrease in the proportion of Lasiopodomys anglicus, which however remains dominant (Table 3). This indicates a moment in the Late Pleistocene that can be considered relatively less cold than the following GHs, but still colder than the preceding phase when subarctic and tundra elements are less represented. GH 10 represents another relatively humid phase, while GH 9 presents again as cold and relatively dry (Fig. 6).
The Habitat Weighting based on habitat suitability offers a different perspective and allows us to understand better the changes in the landscape and environmental conditions (Table 5). It is interesting to notice that the forest component seems to be relatively stable thought the sequence, except for GH 15 and GH 9 that present the lowest percentages. This stability might be linked to the constant presence of water in the Ach River system, as testified by the fish assemblage, that might have been able to support a riparian forest. The relative percentage of wetlands also does not vary much, although it is possible to divide the sequence into two phases: GH 15 to 13 with wetlands at > 9%, and GH 12 to 9 with < 9%. An interesting result is an increase in shrublands with a general decrease of all the other habitats, i.e. grassland, forest, wetlands, rocky, and desert, in GH 12 and partially in GH 11. This might indicate a process of degradation of the grasslands, possibly brought by the loss of ground-water resources, as indicated by the % of Wa and Wetland. It might also point to a stronger seasonality in precipitations that could generate generally drier conditions and facilitate woody plant encroachment (Archer et al. 2017).
To summarize, the landscape surrounding Hohle Fels during the deposition of the Middle Paleolithic Horizons was dominated by open environments, with forest components always present, due to the influence of the Ach River. GH 15 presents as relatively temperate and humid; GH 14 as more temperate and humid; GH 13 less temperate and humid; GH 12 and 11 cold and arid but relatively more temperate than the following periods; GH 10 is still cold but less arid; GH 9 clearly colder and arid.
The data from the bioclimatic model are consistent with our environmental reconstruction (Table 6). The MATs inferred for the sequence are always lower than modern temperatures, between 4.2 (GH 14) and 9.48 °C (GH 9) less than the present. Ps are also significantly lower, between 638 (GH13) and 672.73 mm (GH9) less than present mean annual precipitation. After a relatively temperate and humid period (GH 13–15), climatic conditions became increasingly colder and more arid (GH 9–12). These results confirm the trend already observed by Rhodes and Conard (2021) for GH 9–12. Furthermore, our reconstruction also coincides with the paleoclimatic data obtained from micromorphology analysis of GH 9–12 (Miller 2015), which showed a gradual decrease in temperate and humid conditions throughout the Middle Paleolithic layers excavated at the time.
Based on the limited radiocarbon dates available at the time for the lower GHs, Rhodes (2019) suggested that the shift in climatic conditions between GH 12 and 11 might take place around the time of the Greenland Interstadial (GI) 12, at ca. 47 ka cal BP. Using the new ESR dates available for GH 12 and the radiocarbon dates for the upper Middle Paleolithic Horizons, we can tentatively try to relate the different GHs to the oxygen isotope curve for the Late Pleistocene (Fig. 7). GH 12, dated at 62.5 ± 4 ka (Conard et al. 2021), corresponds to the end of Marine Isotope Stage (MIS) 4—beginning of MIS 3. Given its relatively temperate signal, it might correspond to the GI 18 (ca. 64 ka BP) or to the beginning of MIS 3 (ca. 59–57 ka BP) (after Rasmussen et al. 2014). Consequently, GH 15 to 13 probably correspond to the end of MIS 5 (ca. 87–71 ka BP, after Rasmussen et al. 2014), and we can cautiously assign GH 14, with the highest MAT, to GI 21, 20, or 19, while GH 15 and 13 to the cooling periods preceding and following it (as described in Wohlfarth 2013). GH 11 to 9 show a trend toward colder and more arid conditions, typical of MIS 3, but it is not possible at the time to correlate these GHs to any particular point of the curve.

Bioclimatic reconstruction for the Middle Paleolithic Horizons of Hohle Fels with tentative correlation to the Isotope Curve (after Lisiecki and Raymo 2005). MAT: mean annual temperature; P: mean annual precipitations
Small mammals: biochronological inferences
Furthermore, the stage of evolution of the water vole Arvicola amphibius as quantified by the Schmelzband-Differenzierung-Quotient (SDQ) can help with the chronological attribution of the lower Middle Paleolithic Horizons. A general decrease in the value of SDQ index is observed in water vole populations all across Europe during the Middle and Late Pleistocene (Heinrich 1990; van Kolfschoten 1992; Koenigswald von 1994; van Kolfschoten and Turner 1996; Desclaux et al. 2000; Maul et al. 2000; Paunescu et al. 2004; Kalthoff et al. 2007; Cuenca-Bescós et al. 2010). However, this trend is not perfectly smooth through time or synchronous in all regions (Maul and Markova 2007; Maul et al. 2007, 2014; Masini et al. 2020). Hence, criticisms have been made about the use of the SDQ index as a tool for specific determination and/or stratigraphic correlations (Escudé et al. 2008; Martin 2014). The SDQ pattern is in fact influenced by factors including climate, geography, altitude, latitude, and migration (Kratochvil 1981; Röttger 1987; van Kolfschoten 1992; Ruddy 2011) but, even considering its limits, the SDQ index remains a very powerful descriptive tool and, given the amount of study and analysis presented in literature, a solid base for comparison. Furthermore, once the context is taken into account (i.e., the small mammal assemblage as a whole, the geographic position, the possibility of contact between different populations, and the morphology of the molars), it is possible to use the SDQ index to obtain a reliable biochronological indication for the relative age of a site.
Only GH 15, 14, 13, and 12 at Hohle Fels present complete first lower molars of A. amphibius which could be measured (Table 7). Although the number of specimens analysed is limited, it is possible to see that the values for the index in GH 15, 14, and 13 are > 90, while the value of the sample from GH 12 is only 84.2. Compared with SDQ values of populations from Central Europe (Heinrich 1990; Kalthoff et al. 2007; Maul et al. 2014), our data suggest that GH 15 to 13 might be related to the Early Weichselian (MIS 5d-5a) while GH 12 belongs to a later period, in accordance with the ESR dates obtained for GH 12 and with our paleo-environmental and -climatic reconstruction.
Were Neanderthal occupations at Hohle Fels influenced by climatic and environmental changes?
Neanderthal subsistence practices in freshwater ecosystems
Several authors presented evidence of aquatic resources exploitation during the Lower Paleolithic in Africa (Brooks et al. 1995; Yellen et al. 1995) and Neanderthal marine coastal adaptation, including fishing activities, in the Iberian Peninsula and the Atlantic coast (e.g., Morales-Muñiz and Roselló-Izquierdo 2005; Stringer et al. 2008; Brown et al. 2011; Zilhão et al. 2020). Nevertheless, the possibility of the exploitation of continental freshwater ecosystems in Western-Central Europe by Neanderthal groups has not often been the subject of systematic research. This is partially due to the lack of well-preserved, abundant assemblages. However, the study of inland fishing activities, occurring close to rivers or lakes, during the Middle Paleolithic has been shown to have great potential by authors who have already analysed this topic with interesting results (e.g., Blanco-Lapaz 2019; Guillaud et al. 2021).
Our analysis shows that the freshwater ecosystem around this site was characterized by a pre-mountain river system with the presence of permanent, oxygen-rich, and cold running waters inhabited by preys suitable to humans, due to their relatively larger size and weight (Morales-Muñiz and Roselló-Izquierdo 2005). Other Middle Pleistocene sites in Western Europe show smaller fish accumulations, indicating probably that this abundance of fish results from birds of prey or small-medium carnivores (Guillaud et al. 2021). Based on our preliminary results on the Hohle Fels fish remains, we cannot discard fishing as part of the subsistence practices of Paleolithic hunter-gatherers in Central Europe. However, future studies on fish remains would be necessary to better understand the local freshwater ecosystems, the taphonomic history of fish accumulation, and how fish constitute part of the human diet. In an effort to address some of these issues, additional studies from other Swabian Palaeolithic sites are underway. Exploring more fish assemblages from Middle Paleolithic sites could also help to investigate, and possibly refute, the existing paradigm that Neanderthals were unable to use fishes as a resource in continental waters (Morales-Muñiz and Roselló-Izquierdo 2005; Conard et al. 2013; Guillaud et al. 2021; Blanco-Lapaz et al. 2021).
Neanderthal occupations at Hohle Fels Cave
The upper units attributed to the Middle Paleolithic, corresponding to AH VI-IX / GH 9–12, are relatively poor in anthropogenic finds (i.e., lithic industries, burned bones, modified bones) (Conard et al. 2012). Going from bottom to top, the density of the finds decreases and the layer underlying the first Aurignacian proved to be essentially sterile (Conard and Bolus 2008). This led to the hypothesis that the last Neanderthal groups and the first modern human inhabited the cave with little or no overlaps (Conard et al. 2006). The low density of finds in AH VI-IX might indicate frequent moves and few long-term occupations by large groups of Neanderthals and, although well-adapted to the steppe environment, population density also appears to be low (Conard et al. 2012). On the other hand, the new lower Middle Paleolithic units, corresponding to AH X-XI / GH 13–15, present a relatively high finds density, not comparable with the density of the Aurignacian layers, but remarkably higher than those of AH VI-IX (Conard et al. 2021). This increase in anthropogenic findings coincides with the more temperate climatic phase detected by the analysis of the small mammal assemblage. This suggests that Neanderthals used the cave more intensely during periods with less harsh conditions. The increased levels of all classes of anthropogenic materials reflect the presence of larger groups, occupations for longer periods, or both. The archaeological signal might also point to higher population density at the end MIS 5 than during MIS 3. Changes in sedimentation rates and taphonomic factors including biological, geological, and/or anthropogenic influences might also contribute to the differences in anthropogenic signals between AH VI-IX and AH X-XII. Thus, more studies are ongoing to ascertain the characteristics of the newly excavated AHs. Here, it is important to note that the deepest part of the excavation currently corresponds to only 6m2, and a far greater sample is needed to confirm or refute these trends.
Among the findings from AH X, the most remarkable is the presence complete leaf point, the first one recovered in situ in the Swabian Jura after the 1936 excavation at Haldenstein (Lone Valley) conducted by Riek (Riek 1938). The leaf point is 7.6 cm long, 4.1 cm wide, 0.9 cm thick, made from locally available, grey Jurassic chert from the Swabian region. It was hafted at the less pointy end, probably used as a projectile during hunting, and discarded after a failed attempt at resharpening (Rots et al. 2021). In Germany, leaf points are referred to as “Blattspitzen” and until now, the Blattspitzengruppe, the cultural unit to which the leaf points belong, has been assigned to the final Middle Paleolithic (Bosinski 1967; Conard and Fischer 2000; Bolus 2004, 2011). The new dates for GH 12, combined with the biostratigraphic signal of A. amphibius and the climatic reconstruction here presented, would place the leaf point of Hohle Fels at a much earlier time than expected and thereby questioning the traditional cultural chronology of Swabian Jura. Here, it is important to reiterate that our biostratigraphic arguments are consistent with new radiometric dates (Conard et al. 2021). A reappraisal of this cultural unit and its chronological attribution is needed to document Middle Paleolithic technology and hunting strategies, as well as to refine our understanding of the circumstances that led to the extinction of Neanderthals. Every indication suggests that the ongoing excavations at Hohle Fels will continue to contribute to these research questions.
Neanderthal occupations at Hohle Fels Cave in the context of Central-Western Europe (Germany, Belgium, Poland, and France)
The small mammal assemblages related to the Middle Paleolithic GHs at Hohle Fels provide a fundamental regional reference for this chronology, for its sample size, and for the reliable dates available for the sequence. The assemblage of Geißenklosterle (GHs 23–18, Ach Valley, Rhodes et al. 2018) is dated to the end of MIS 5 up to MIS 3 (Richard et al. 2019b). It is smaller in size, but consistent in faunal composition with the Hohle Fels one. The assemblage of Stadel-Höhle at Hohlenstein (Lone Valley, Ziegler 2019) is dated to the MIS 3 (Richard et al. 2019a) and it differs from the Hohle Fels one for the presence, although sporadic, of Clethrionomys glareolus, Glis glis, Sciurus vulgaris, Apodemus sp., indicators of woodland and forest patch, and of Ochotona pusilla and large- and small-size hamsters, indicators of open and arid environments. These differences might indicate that micro-environmental and micro-climatic conditions were not the same in the two valleys during the last Neandertal occupations.
A recent publication by Rhodes and Conard (2021) suggests that climatic conditions in the Ach Valley were colder and more arid than in other Central European sites with the same chronology as GHs 12–7 of Hohle Fels. The present study confirms these results also for the lower GHs 15–13. The small mammal assemblage from Sesselfelsgrotte (Germany, layers L-G, MIS 4–3; van Kolfschoten 2014) indicates generally more temperate conditions than those registered at Hohle Fels, due to the relative dominance of M. arvalis/agrestis over L. anglicus (= Microtus gregalis) and the presence of C. glareolus. Data from small mammal assemblages directly related to Middle Paleolithic occupations at Scladina Cave (MIS 5–3, López-García et al. 2017a, 2017b) and Marie-Jeanne Cave (MIS 3, López-García et al. 2017a), both located in Belgium, indicate more temperate conditions than at Hohle Fels, testified by the presence of C. glareolus, Microtus (Terricola) subterraneus, Glis glis, and Apodemus gr. sylvaticus/flavicollis. During MIS 5, the landscape at Scladina and Marie-Jeanne Caves was always dominated by woodland, while that at Hohle Fels open environment is always dominant. During both MIS 5 and MIS 3, the mean annual precipitation indicates more humid conditions than at Hohle Fels. Similar comparisons can be drawn between the Hohle Fels assemblage and the assemblage from Biśnik Cave (MIS 5–3, layers 13 to 9; Socha 2014) in Poland. Both faunal composition and bioclimatic reconstructions indicate more temperate and humid conditions than at Hohle Fels. The small mammals from Obłazowa Cave (Poland, early MIS 3; Valde-Nowak and Nadachowski 2014) also indicate more humid conditions, due to the relatively high percentage of A. oeconomus. Another recent study concerning small mammal assemblages in Middle Paleolithic contest at Koziarnia Cave (Poland, Berto et al. 2021) indicates a general dominance of the open environment, but the sample size does not allow for further comparisons. According to the environmental reconstructions of Discamps and Royer (2017), the conditions at Hohle Fels seem to be close to those of the sites of the Atlantic region (i.e. Charente-Maritime, Charente, and Gironde, see Fig. 1), with L. anglicus dominant over M. arvalis/agrestis and generally dry and open landscape, while in the rest of southern France conditions seem more humid and temperate, with a higher component of shrubland and woodland.
In this context, the Middle Paleolithic occupations registered at Hohle Fels both during MIS 5 (GHs 15- 13) and MIS4-3 (GHs 12–9) took place in a more arid and colder environmental setting than those registered in Sesselfelsgrotte, Poland, and Belgium. Further studies are needed to clarify if this is a local (Ach Valley) or regional (Swabian Jura) peculiarity.
Conclusions
The joint study of fish and small mammal assemblages from the Middle Paleolithic of Hohle Fels provided insights into the climatic and environmental conditions faced sby Neanderthals in the Ach Valley. The freshwater ecosystem around this site was characterized by a pre-mountain river system with the presence of permanent, oxygen-rich, and cold running waters. The landscape was dominated by an open environment, and thanks to the presence of the Ach river, the forest component was always relatively stable throughout the sequence. Our research has identified two major climatic oscillations: one more temperate corresponding to Neanderthal occupations of relatively high intensity, the other colder and more arid, presenting a trend toward increasingly harsher conditions, characterized by low-intensity Neanderthal occupations. Future studies, including additional material from Aurignacian, Gravettian, and Magdalenian units of Hohle Fels (already ongoing) will provide further knowledge about the changes in climate and environment during the human occupations at Hohle Fels. This ongoing research will document how human groups interacted with and adapted to the different climatic settings and will further illuminate their use of freshwater resources.
Data availability
Raw data are available upon request. Corresponding author contact: elisa.luzi@ifu.uni-tuebingen.de.
References
Andrews P (1990) Owl, Caves and Fossils. British Museum (Natural History), London
Andrews P (2006) Taphonomic effects of faunal impoverishment and faunal mixing. Palaeogeogr Palaeoclimatol Palaeoecol 241:572–589. https://doi.org/10.1016/j.palaeo.2006.04.012
Archer SR, Andersen EM, Predick KI, et al (2017) Woody Plant Encroachment: causes and Consequences. In: Briske DD (ed) Rangeland Systems. Processes, Management and Challenges, Springer S. pp 25–84
Baca M, Popović D, Lemanik A et al (2019) Highly divergent lineage of narrow-headed vole from the Late Pleistocene Europe. Sci Rep 9:1–10. https://doi.org/10.1038/s41598-019-53937-1
Barth MM, Conard NJ, Münzel SC (2009) Palaeolithic subsistence and organic technology in the Swabian Jura. Search Total Anim Exploit Case Stud from up Palaeolithic Mesolith Proc XVth UISPP Congr Sess C61(42):5–20
Baumann C, Wong GL, Starkovich BM et al (2020) The role of foxes in the Palaeolithic economies of the Swabian Jura (Germany). Archaeol Anthropol Sci 12:208. https://doi.org/10.1007/s12520-020-01173-4
Beattie S, Starkovich BM, Conard NJ (2021) Middle Palaeolithic Avian fauna from Hohle Fels, Germany and Neanderthal lifeways. In: Back to the Gravettian. 62nd conference of the Hugo Obermaier Society, April 6th-8th 2021, Brno. pp 21–22
Bertacchi A, Starkovich BM, Conard NJ (2021) The Zooarchaeology of Sirgenstein Cave: a Middle and Upper Paleolithic site in the Swabian Jura, SW Germany. J Paleolit Archaeol 4:1–44
Berto C, Krajcarz MT, Hoyo MM del, et al (2021) Environment changes during Middle to Upper Palaeolithic transition in southern Poland (Central Europe). A multiproxy approach for the MIS 3 sequence of Koziarnia Cave (Kraków-Częstochowa Upland). J Archaeol Sci Reports 35: https://doi.org/10.1016/j.jasrep.2020.102723
Blanco-Lapaz A, Martínez-Monzón A, Blain H-A, Cuenca-Bescós G (2021) Early-Middle Pleistocene freshwater ecosystems in the Sierra de Atapuerca (northern Iberia) based on the Gran Dolina fish record. Palaeogeogr Palaeoclimatol Palaeoecol 574:110444. https://doi.org/10.1016/j.palaeo.2021.110444
Blanco-Lapaz Á (2019) Fische. In: Kind C-J (ed) Löwenmensch und mehr. Die Grabungen 2009 bis 2013 in der Stadel-Höhle. pp 175–178
Bolus M (2004) Settlement analysis of sites of the Blattspitzen complex in central Europe. Settl Dyn Middle Paleolit Middle Stone Age 2(2):201–226
Bolus M (2011) The late Middle Paleolithic and the Aurignacian of the Swabian Jura, Southwestern Germany. In: Proceedings of the International Symposium “Characteristic Features of the Middle to Upper Paleolithic Transition in Eurasia: Development of Culture and Evolution of Homo Genus” (July 4–10, 2011, Denisova Cave, Altai). pp 3–10
Borodin AV, Markova EA (2015) Keys to identify modern and Pleistocene arvicolines (Arvicolinae, Rodentia) from the Urals and Western Siberia based on odontological characteristics. Biol Bull 42:652–663
Bosinski G (1967) Die mittelpaläolithischen Funde im westlichen Mitteleuropa. Böhlau Verlag, Köln and Graz, Fundamenta
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51:337–360
Brooks AS, Helgren DM, Cramer JS et al (1995) Dating and context of three middle stone age sites with bone points in the Upper Semliki Valley, Zaire. Science (80-) 268:548–553. https://doi.org/10.1126/science.7725099
Brown K, Fa DA, Finlayson G, Finlayson C (2011) Small game and marine resource exploitation by Neanderthals: the evidence from Gibraltar. Trekking the Shore. Springer, New York, pp 247–272
Cannon DY (1987) Marine fish osteology: a manual for archaeologists. Archaeology Press, Simon Fraser University, Burnaby, 133 p. Archaeology Press, Simon Fraser University, Burnaby
Conard NJ (2009) A female figurine from the basal Aurignacian of Hohle Fels Cave in southwestern Germany. Nature 459:248–252. https://doi.org/10.1038/nature07995
Conard NJ, Bolus M (2008) Radiocarbon dating the late middle paleolithic and the aurignacian of the Swabian Jura. J Hum Evol 55:886–897. https://doi.org/10.1016/j.jhevol.2008.08.006
Conard NJ, Fischer B (2000) Are there recognizable cultural entities in the German Middle Palaeolithic? In: Ronen A, Weinstein-Evron M (eds) Toward Modern Humans: The Yabrudian and Micoquian 400–50 kyears ago, British Ar. Archaeopress, Oxford, pp 7–24
Conard NJ, Janas A (2021) Ausgrabungen im Hohle Fels: Fundschichten aus dem Mittelpaläolithikum und Neues zur Jagdtechnik der Neandertaler. Archäologische Ausgrabungen Bad Württemb 2020:60–65
Conard NJ, Malina M, Münzel SC (2009) New flutes document the earliest musical tradition in southwestern Germany. Nature 460:737–740. https://doi.org/10.1038/nature08169
Conard NJ, Bolus M, Münzel SC (2012) Middle Paleolithic land use, spatial organization and settlement intensity in the Swabian Jura, southwestern Germany. Quat Int 247:236–245. https://doi.org/10.1016/j.quaint.2011.05.043
Conard. NJ, Kind CJ (ed) (2017) Als der Mensch die Kunst erfand: Eiszeithöhlen der Schwäbischen Alb. Theiss Verlag
Conard NJ, Bolus M, Goldberg P, Münzel SC (2006) The last Neanderthals and first modern humans in the Swabian Jura. In: When Neanderthals and modern humans met. pp 305–341
Conard NJ, Kitagawa K, Krönneck P, et al (2013) The importance of fish, fowl and small mammals in the Paleolithic diet of the Swabian Jura, Southwestern Germany. In: Zooarchaeology and Modern Human Origins: Human Hunting Behavior during the Later Pleistocene, Vertebrate Paleobiology and Paleoanthropology. pp 173–190
Conard NJ, Janas A, Richard M et al (2021) The cultural and chronostratigraphic context of a new leaf point from Hohle Fels Cave in the Ach Valley of southwestern Germany. Mitteilungen der Gesellschaft für Urgeschichte 30
Conroy JWH, Watt J, Webb JB, Jones A (2005) A guide to the identification of prey remains in otter spraint, 3rd edn. The Mammal Society, London
Cuenca Bescós G (2003) Análisis filogenético de Allocricetus del Pleistoceno (Cricetidae, Rodentia, Mammalia ) from the Pleistocene. A Phylogenetical Approach Coloquios Paleontol 1:95–113
Cuenca-Bescós G, Agustí J, Lira J et al (2010) A New species of water vole from the Early Pleistocene of Southern Europe. Acta Palaeontol Pol 55:565–580. https://doi.org/10.4202/app.2009.0027
Desclaux E, Abbassi M, Marquet J-C et al (2000) Distribution of Arvicola (Mammalia, Rodentia) in France and Liguria (Italy) during the Middle and Upper Pleistocene. Acta Zool Cracoviensia 43:107–125
Discamps E, Royer A (2017) Reconstructing palaeoenvironmental conditions faced by Mousterian hunters during MIS 5 to 3 in southwestern France: a multi-scale approach using data from large and small mammal communities. Quat Int 433:64–87. https://doi.org/10.1016/j.quaint.2016.02.067
Doadrio Villarejo JI (2001). Atlas y libro rojo de los peces continentales de España. Museo Nacional de Ciencias Naturales, Dirección General de Conservación de la Naturaleza, Madrid
Elliott JM, Elliott JA (2010) Temperature requirements of Atlantic salmon Salmo salar, brown trout Salmo trutta and Arctic charr Salvelinus alpinus: predicting the effects of climate change. J Fish Biol 77:1793–1817. https://doi.org/10.1111/j.1095-8649.2010.02762.x
Escudé E, Montuire S, Desclaux E et al (2008) Reappraisal of ‘chronospecies’ and the use of Arvicola (Rodentia, Mammalia) for biochronology. J Archaeol Sci 35:1867–1879. https://doi.org/10.1016/j.jas.2007.11.018
Evans EM, Van Couvering JAH, Andrews P (1981) Palaeoecology of Miocene sites in Western Kenya. J Hum Evol 10:99–116. https://doi.org/10.1016/S0047-2484(81)80027-9
Fernández-Jalvo Y, Andrews P, Denys C et al (2016) Taphonomy for taxonomists: implications of predation in small mammal studies. Quat Sci Rev 139:138–157
Frontini R, Roselló-Izquierdo E, Morales-Muñiz A et al (2021) Compression and digestion as agents of vertebral deformation in Sciaenidae. Merlucidae and Gadidae Remains: an Experimental Study to Interpret Archaeological Assemblages J Archaeol Method Theory. https://doi.org/10.1007/s10816-021-09527-5
Füllner G, Pfeifer M, Völker F, Yarske A (2016) Atlas der Fische Sachsens. Landesamt für Umwelt, Landwirtschaft und Geologie, Senckenber
Guillaud E, Lebreton L, Béarez P (2018) Taphonomic signature of Eurasian eagle owl (Bubo bubo) on fish remains. Folia Zool. 67:143. https://doi.org/10.25225/fozo.v67.i3-4.a1.2018
Guillaud E, Béarez P, Daujeard C et al (2021) Neanderthal foraging in freshwater ecosystems: a reappraisal of the Middle Paleolithic archaeological fish record from continental Western Europe. Quat Sci Rev 252:106731. https://doi.org/10.1016/j.quascirev.2020.106731
Hahn J (1979) Ausgrabungen in urgeschichlichen Fundstellen des Achtales zwischen Blaubeuren und Schelklingen, Alb-Donau-Kreis. Archäologische Augrabungen Baden-Württemb 1978:11–15
Hahn J (1978) Die altsteinzeitliche Schichtenfolge des „Geißenklösterle“ bei Blaubeuren nach der. Grabung 1977 Archäeologische Ausgrabungen Baden-württemb 1977 7–10
Hammer Ø, Harper DA, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4(1):9
Heinrich W-D (1978) Zur biometrischen Erfassung eines Evolutionstrends bei Arvicola (Rodentia, Mammalia) aus dem Pleistozän Thüringens. Säugetierkundliche Informationen 2:3–21
Heinrich W-D (1990) Some aspects of the evlution and biostratigraphy of Arvicola (Mammalia, Rodentia) in the Central European Pleistocene. In: Fejfar O, Heinrich W-D (eds) International Symposium Evolution, Phylogeny, and Bostratigraphy of Arvicolids (Rodentia, Mammalia). Pfeil-Verlag, Munchen, pp 165–182
Hernández Fernández M (2001) Bioclimatic discriminant capacity of terrestrial mammal faunas. Glob Ecol Biogeogr 10:189–204. https://doi.org/10.1046/j.1466-822x.2001.00218.x
Hernández Fernández M, Peláez-Campomanes P (2005) Quantitative palaeoclimatic inference based on terrestrial mammal faunas. Glob Ecol Biogeogr 14:39–56. https://doi.org/10.1111/j.1466-822X.2004.00125.x
Hernández Fernández M (2005) Análisis paleoclimático y paleoecológico de las sucesiones de mamíferos del Plio-Pleistoceno de la Península Ibérica. PhD dissertation: Universidad Complutense de Madrid
Higham T, Basell L, Jacobi R et al (2012) Τesting models for the beginnings of the Aurignacian and the advent of figurative art and music: the radiocarbon chronology of Geißenklösterle. J Hum Evol 62:664–676. https://doi.org/10.1016/j.jhevol.2012.03.003
Hir J (1993) Cricetulus migratorius (PALLAS 1773)(Rodentia, Mammalia) population from the Toros Mountains (Turkey)(With a special reference to the relation of Cricetulus and Allocricetus genera). Folia Hist Nat Musei Matraensis 18:17–34
IUCN (2021) The IUCN red list of threatened species. Version 2021. https://www.iucnredlist.org/
Jonsson B, Jonsson N (2009) A review of the likely effects of climate change on anadromous Atlantic salmon Salmo salar and brown trout Salmo trutta, with particular reference to water temperature and flow. J Fish Biol 75:2381–2447. https://doi.org/10.1111/j.1095-8649.2009.02380.x
Kalthoff DC, Mörs T, Tesakov A (2007) Late Pleistocene small mammals from the Wannenköpfe volcanoes (Neuwied Basin, western Germany) with remarks on the stratigraphic range of Arvicola terrestris. Geobios 40:609–623
Kitagawa K (2014) Exploring hominins and animals in the Swabian Jura: study of the Paleolithic fauna from Hohlenstein-Stadel. PhD dissertation, Universitätsbibliothek Tübingen
Kratochvil J (1981) Arvicola cantiana vit-elle encore? Folia Zool 29:209–224
Kryštufek B, Tesakov AS, Lebedev VS et al (2019) Back to the future: the proper name for red-backed voles is Clethrionomys Tilesius and not Myodes Pallas. Mammalia 84:214–217
Laplana C, Sevilla P, Arsuaga JL et al (2015) How far into europe did pikas (Lagomorpha: Ochotonidae) go during the Pleistocene? New Evidence from Central Iberia. PLoS ONE 10:e0140513. https://doi.org/10.1371/journal.pone.0140513
Lepiksaar J (1994) Introduction to Osteology of Fishes for Paleozoologists. Göteborg University Press, Göteborg
Lisiecki LE, Raymo ME (2005) A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records. Paleoceanography 20:1–17. https://doi.org/10.1029/2004PA001071
Lissovsky AA, Petrova TV, Yatsentyuk SP et al (2018) Multilocus phylogeny and taxonomy of East Asian voles Alexandromys (Rodentia, Arvicolinae). Zool Scr 47:9–20. https://doi.org/10.1111/zsc.12261
López-García JM, Berto C, Colamussi V et al (2014) Palaeoenvironmental and palaeoclimatic reconstruction of the latest Pleistocene-Holocene sequence from Grotta del Romito (Calabria, southern Italy) using the small-mammal assemblages. Palaeogeogr Palaeoclimatol Palaeoecol 409:169–179. https://doi.org/10.1016/j.palaeo.2014.05.017
López-García JM, Blain H-A, Lozano-Fernández I et al (2017a) Environmental and climatic reconstruction of MIS 3 in northwestern Europe using the small-mammal assemblage from Caverne Marie-Jeanne (Hastière-Lavaux, Belgium). Palaeogeogr Palaeoclimatol Palaeoecol 485:622–631. https://doi.org/10.1016/j.palaeo.2017.07.017
López-García JM, Blain HA, Cordy JM et al (2017b) Palaeoenvironmental and palaeoclimatic reconstruction of the Middle to Late Pleistocene sequence of Scladina Cave (Namur, Belgium) using the small-mammal assemblages. Hist Biol 29:1125–1142. https://doi.org/10.1080/08912963.2017.1288229
Martin RA (2014) A critique of vole clocks. Quat Sci Rev 94:1–6. https://doi.org/10.1016/j.quascirev.2014.03.004
Masini F, Maul LC, Abbazzi L et al (2020) Independent water vole (Mimomys savini, Arvicola: Rodentia, Mammalia) lineages in Italy and Central Europe. Foss Impr 76:59–83. https://doi.org/10.37520/fi.2020.005
Maul LC, Markova AK (2007) Similarity and regional differences in Quaternary arvicolid evolution in Central and Eastern Europe. Quat Int 160:81–99. https://doi.org/10.1016/j.quaint.2006.09.010
Maul LC, Rekovets L, Heinrich W-D et al (2000) Arvicola mosbachensis (Schmidtgen 1911) of Mosbach 2: a basic sample for the early evolution of the genus and a reference for further biostratigraphical studies. Senckenb Lethaea 80:129–147
Maul LC, Heinrich WD, Parfitt SA, Paunescu AC (2007) Comment on the correlation between magnetostratigraphy and the evolution of Microtus (Arvicolidae, Rodentia, Mammalia) during the Early and early Middle Pleistocene. CFS Courier Forschungsinstitut Senckenberg. CFS Cour Forschungsinstitut Senckenb 259:243–263
Maul LC, Masini F, Parfitt SA et al (2014) Evolutionary trends in arvicolids and the endemic murid Mikrotia - New data and a critical overview. Quat Sci Rev 96:240–258. https://doi.org/10.1016/j.quascirev.2013.09.017
Miller CE (2015) A Tale of Two Swabian Caves. Geoachaeological investigations ah Hohle Fels and Geißenklösterle. P. Kerns Verlag, Tübingen
Morales-Muñiz A, Roselló-Izquierdo E (2005) Ictiofaunas musterienses de la Península Ibérica:¿ evidencias de la pesca Neanderthal? Munibe Antropol 57:183–195
Nadachowski A (1982) Late quaternary rodents of Poland with special reference to morphotype dentition analysis of voles. P.W.N., Warszava - Krakow
Nicholson RA (1993) An investigation into the effects on fish bone on passage through the human gut: some experiments and comparisons with archaeological material. Circaea 10:38–51
Niethammer J, Krapp F (1978) Handbuch der Säugetiere Europas. Band 1/Rodentia I (Sciuridae, Castoridae, Gliridae, Muridae). Akademische Verlagsgesellschaft
Niethammer J, Krapp F (1982) Handbuch der Säugetiere Europas. Band 2/I Rodentia II (Cricetidae, Arvicolidae, Zapodidae, Spalacidae, Hystricidae, Capromyidae). Akademische Verlagsgesellschaft
Niethammer J, Krapp F (1990) Handbuch der Säugetiere Europas. Band 3/I Insectivora, Primates (Erinaceidae, Talpidae, Spricidae, Cercopithecidae). AULA-Verlag
Paunescu A, Maul LC, Masini F (2004) Comparison of evolutionary patterns of Arvicola in Germany , France and Italy. In: Maul LC, Kahlke R-D (eds) Late Neogene and Quaternary biodiversity and evolution: regional developments and interregional correlations. Conference Volume. 18 th International Senckenberg Conference. VI International Palaeontological Colloquium in Weimar . Weimar (Germany), 25th -, Terra Nost. Berlin, pp 203–204
Petrova TV, Zakharov ES, Samiya R, Abramson NI (2015) Phylogeography of the narrow-headed vole Lasiopodomys (Stenocranius) gregalis (Cricetidae, Rodentia) inferred from mitochondrial cytochrome b sequences: an echo of Pleistocene prosperity. J Zool Syst Evol Res 53:97–108. https://doi.org/10.1111/jzs.12082
Popov VV, Zidarova SA (2008) Patterns of craniometric variability of Neomys fodiens and Neomys anomalus (Mammalia, Insectivora) in Bulgaria - role of abiotic and biotic factors. Acta Zool Bulg 60:171–185
Rasmussen SO, Bigler M, Blockley SP et al (2014) A stratigraphic framework for abrupt climatic changes during the Last Glacial period based on three synchronized Greenland ice-core records: Refining and extending the INTIMATE event stratigraphy. Quat Sci Rev 106:14–28. https://doi.org/10.1016/j.quascirev.2014.09.007
Rhodes SE (2019) A synthesis of the paleoecology of the Ach Valley during the Middle to Upper Paleolithic Transition with an expanded discussion of the small mammal record from Hohle Fels and Geißenklösterle Caves. Mitteilungen Der Gesellschaft Für Urgeschichte 28:51–104
Rhodes SE, Conard NJ (2021) A quantitative paleoclimatic reconstruction of the non-analogue environment of oxygen isotope stage 3: new data from small mammal records of southwestern Germany. Archaeol Anthropol Sci 13: https://doi.org/10.1007/s12520-021-01363-8
Rhodes SE, Ziegler R, Starkovich BM, Conard NJ (2018) Small mammal taxonomy, taphonomy, and the paleoenvironmental record during the Middle and Upper Paleolithic at Geißenklösterle Cave (Ach Valley, southwestern Germany). Quat Sci Rev 185:199–221. https://doi.org/10.1016/j.quascirev.2017.12.008
Rhodes SE, Starkovich BM, Conard NJ (2019) Did climate determine Late Pleistocene settlement dynamics in the Ach Valley, SW Germany? Plos one, 14(5), e0215172.
Richard M, Falguères C, Valladas H et al (2019b) New electron spin resonance (ESR) ages from Geißenklösterle Cave: a chronological study of the Middle and early Upper Paleolithic layers. J Hum Evol 133:133–145. https://doi.org/10.1016/j.jhevol.2019.05.014
Richard M, Falguères C, Pons-Branchu E, Richter D (2019a) Dating the stratigraphic sequence at Hohlenstein- Stadel Cave using ESR and U-Th dating methods. In: Kind C-J (ed) Löwenmensch und mehr. Die Ausgrabungen 2008–2013 in den altsteinzeitlichen Schichten der Stadel-Höhle im Hohlenstein (Lonetal), Gemeinde Asselfingen, Alb-Donau-Kreis. Dr. Ludwig Reichert Verlag, Wiesbaden, pp 95–101
Riek G (1938) Ein Beitrag zur kenntnis des süddeutschen Solutréen. Ger Anzeiger Der Römisch-Germanischen Kommission Des Dtsch Archäologischen Instituts 22:147–150
Rots V, Coppe J, Conard N (2021) A leaf point documents hunting with spears in the Middle Paleolithic at Hohle Fels, Germany. Mitteilungen der Gesellschaft für Urgeschichte 2021:1–28. https://doi.org/10.51315/mgfu.2021.30004
Röttger U (1987) Shmelzbandbreiten an Molaren von Schermausen (Arvicola, Lacépède, 1799). Bonn Zool Beitr 38:95–105
Royer A, Yelo BAG, Laffont R, Fernández MH (2020) New bioclimatic models for the Quaternary Palaearctic based on insectivore and rodent communities. Palaeogeogr Palaeoclimatol Palaeoecol 560:110040
Ruddy M (2011) The Western Palaearctic evolution of the water vole Arvicola. PhD dissertation, University of London
Schmidt RR (1912) Die Diluviale Vorzeit Deutschland. E. Schweizerbartsche Verlasbuchhandlung, Stuttgart
Sevilla García P (1988) Estudio paleontológico de los Quirópteros del Cuaternario español. Paleontol i Evol 22:113–233
Socha P (2014) Rodent palaeofaunas from Biśnik cave (Kraków-czestochowa upland, Poland): Palaeoecological, palaeoclimatic and biostratigraphic reconstruction. Quat Int 326–327:64–81. https://doi.org/10.1016/j.quaint.2013.12.027
Sommer C (2019) A map collection of the Paleolithic of the Swabian Jura. Zenodo. https://doi.org/10.5281/zenodo.3460301
Stringer CB, Finlayson JC, Barton RNE et al (2008) Neanderthal exploitation of marine mammals in Gibraltar. Proc Natl Acad Sci 105:14319–14324. https://doi.org/10.1073/pnas.0805474105
Tesakov AS, Lebedev VS, Bannikova AA, Abramson NI (2010) Clethrionomys Tilesius, 1850 is the valid generic name for red-backed voles and Myodes Pallas, 1811 is a junior synonym of Lemmus Link, 1795. Russ J Theriol 9:83–86
Tissot L, Souchon Y (2010) Synthèse des tolérances thermiques des principales espèces de Poissons des rivières et fleuves de plaine de l’Ouest européen. Hydroécol Appliquée Hydroécol Appliquée 17:17–76
Valde-Nowak P, Nadachowski A (2014) Micoquian assemblage and environmental conditions for the Neanderthals in Obłazowa Cave, Western Carpathians, Poland. Quat Int 326–327:146–156. https://doi.org/10.1016/j.quaint.2013.08.057
van Kolfschoten T (1992) Aspects of the migration of mammals to northwestern Europe during the Pleistocene, in particular the reimmigration of Arvicola terrestris. Cour Forsch-Inst Senckenb 153:213–220
van Kolfschoten T, Turner E (1996) Early Middle Pleistocene mammalian faunas from Kärlich and Miesenheim I and their biostratigraphical implications. In: Turner C (ed) The early Middle Pleistocene in Europe. Balkema, Rotterdam, pp 227–253
van Kolfschoten T (2014) The smaller mammals from the Late Pleistocene sequence of the Sesselfelsgrotte (Neuessing, Lower Bavaria). In: Freund G, Reisch L (eds) Sesselfelsgrotte VI. Naturwissenschaftliche Untersuchungen: Wirbeltierfauna 1. Das Paläolithikum und Mesolithikum des Unteren Altmühltals II. Franz Steiner Verlag, Stuttgart, pp 27–117
Velliky EC, Schmidt P, Bellot-Gurlet L et al (2021) Early anthropogenic use of hematite on Aurignacian ivory personal ornaments from Hohle Fels and Vogelherd caves. Germany J Hum Evol 150:102900. https://doi.org/10.1016/j.jhevol.2020.102900
von Koenigswald W (1994) Functional symmetries in the schmelzmuster and morphology of rootless rodent molars. Zool J Linn Soc 110:141–179
Watt J, Pierce GJ, Boyle PR (1997) Guide to the Identification of North Sea Fish Using Premaxillae and Vertebrae. ICES Cooperative research report, 220. International Council for the Exploration of the Sea. Palaegade 2–4, Copenhagen
Wheeler A, Jones AKG (2009) Fishes. Cambridge University Press, Cambridge, Manuals in
Wilson DE, Reeder DM (eds) (2005) Mammal species of the world. A taxonomic and geographic reference, 3rd edn. Johns Hopkins University Press, Baltimora
Wilson DE, Mittermeier RA, Lacher TE (2017) Handbook of the Mammals of the World – Volume 7 Rodents II. Lynx Edicions, Barcelona
Wohlfarth B (2013) A review of Early Weichselian climate ( MIS 5d-a ) in Europe. Technical report/Svensk kärnbränslehantering AB 40 (50)
Wolf S, Conard NJ (2015) Personal Ornaments of the Swabian Aurignacian. Palethnologie 7: https://doi.org/10.4000/palethnologie.897
Wong GL, Drucker DG, Starkovich BM, Conard NJ (2020a) Latest Pleistocene paleoenvironmental reconstructions from the Swabian Jura, southwestern Germany: evidence from stable isotope analysis and micromammal remains. Palaeogeogr Palaeoclimatol Palaeoecol 540:109527. https://doi.org/10.1016/j.palaeo.2019.109527
Wong GL, Starkovich BM, Drucker DG, Conard NJ (2020b) New perspectives on human subsistence during the Magdalenian in the Swabian Jura, Germany. Archaeol Anthropol Sci 12: https://doi.org/10.1007/s12520-020-01119-w
Yellen JE, Brooks AS, Cornelissen E et al (1995) A Middle Stone Age Worked Bone Industry from Katanda, Upper Semliki Valley Zaire. Science (80-) 268:553–556. https://doi.org/10.1126/science.7725100
Ziegler R (2019) Kleinesäuger. In: Kind C-J (ed) Löwenmensch und mehr. Die Ausgrabungen 2008–2013 in den altsteinzeitlichen Schichten der Stadel-Höhle im Hohlenstein (Lonetal), Gemeinde Asselfingen, Alb-Donau-Kreis. Dr. Ludwig Reichert Verlag, Wiesbaden, pp 167–171
Zilhão J, Angelucci DE, Igreja MA, et al (2020) Last Interglacial Iberian Neandertals as fisher-hunter-gatherers. Science (80- ) 367: https://doi.org/10.1126/science.aaz7943
Acknowledgements
The authors would like to thank Alexander Janas and Maria Malina for their many contributions to the excavations at Hohle Fels, for their help in navigating the Hohle Fels database and for the useful discussions. We thank Jacob Schlechtendahl for the photos of the material, and Julia Zastrow for her help in the lab. The authors would like to thank the two reviewers that help improve the quality of the manuscript with their comments and suggestions.
Funding
Open Access funding enabled and organized by Projekt DEAL. E.L. was supported by the Alexander von Humboldt Foundation with a Humboldt Research Fellowship for postdoctoral researchers (ESP1209403HFST-P). S.E.R was supported by a German Academic Exchange Service (DAAD) long-term research grant and a DAAD P.R.I.M.E. postdoctoral grant during this project. The excavations at Hohle Fels have been funded for decades from many sources including the University of Tübingen, the Deutsche Forschungsgemeinschaft, the Heidelberger Akademie der Wissenschaften, the Ministry of Science, Research and Art of Baden-Württemberg, HeidelbergCement, as well as the cities of Schelklingen and Blaubeuren,
Author information
Authors and Affiliations
Contributions
E. Luzi and S. Rhodes analysed the small mammal assemblage. Á. Blanco-Lapaz analysed the fish assemblage. N. Conard directs the excavation at Hohle Fels provided archaeological and geological context and funding. The manuscript was written by E. Luzi. All authors contributed to sections of the manuscript. All authors reviewed and approved the manuscript before submission.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Microvertebrate Studies in Archaeological Contexts: Middle Paleolithic to early Holocene past environments
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Luzi, E., Blanco-Lapaz, À., Rhodes, S.E. et al. Paleoclimatic and paleoenvironmental reconstructions based on the small vertebrates from the Middle Paleolithic of Hohle Fels Cave, SW Germany. Archaeol Anthropol Sci 14, 107 (2022). https://doi.org/10.1007/s12520-022-01568-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-022-01568-5