
Abstract
Located at the crossroads between Africa, Europe and Asia, the Southern Caucasus is a prime location to study occupations by H. heidelbergensis, H. neanderthalensis and anatomically modern humans. Azokh Cave is an important site for the understanding of human evolution in its archaeological, palaeontological, environmental and ecological context. The main objective of this work is to use rodents to infer the climatic and environmental conditions that prevailed during the formation of the site. The small-mammal remains come from the archaeological excavation campaigns carried out in Azokh 1 in 2003, 2005, 2014, 2015 and 2018; they are from Unit V, Units III–IV and Unit II. The small-mammal assemblage is composed of at least 13 taxa: seven arvicoline, two cricetine, two gerbilline, one dipodid and one murine species. Units III–IV do not yield enough material to draw palaeoclimatic inferences. The palaeoclimatic conditions for Units V and II, ascertained by means of the bioclimatic model, suggest temperatures and precipitation similar to nowadays; the climate seems to be relatively warm-temperate in both units. The palaeoenvironmental reconstruction by means of habitat weighting points to an environment mainly composed of desert and steppe habitats, as well as portions of grassland and forest. This interpretation differs from that inferred from the large-mammal and archaeobotanical data, which indicate a woodland environment. These differences could be explained by the origin of the accumulation. There was no evidence of a major palaeoenvironmental or palaeoclimatic change between the Middle and Late Pleistocene layers, indicating favourable conditions throughout the study period.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The area of Western Asia plays an important role in efforts to understand the biological history of human lineages and their techno-cultural complexes during the Pleistocene, as well as large- and small-mammal migrations (Agustí and Lordkipanidze 2019; Bermúdez de Castro and Martinón-Torres 2013; Belmaker et al. 2016). Western Asia was a natural corridor and refuge between Africa and Eurasia during the Pleistocene. The work of Abbate and Sagri (2012) indicates that hominin dispersals occurred during favourable climatic periods, and this area was very attractive for humans. This is evidenced by the high number of sites located in this region, such as Dmanisi (Lordkipanidze et al. 2013; Coil et al. 2020), Qesem Cave (Maul et al. 2016), Misliya Cave (Hershkovitz et al. 2018; Weissbrod and Weinstein-Evron 2020), Nesher Ramla (Hershkovitz et al. 2021), Agithu-3 (Kandel et al. 2017) and Dzudzuana Cave (Belmaker et al. 2016).
Studies of small vertebrates from archaeological sites in this region have been on the increase in the last decade (Belmaker and Hovers 2011; Demirel et al. 2011; Weissbrod and Zaidner 2014; Maul et al. 2015, 2016, 2020; Smith et al. 2015; Belmaker et al. 2016; Parfitt 2016; Kandel et al. 2017; Weissbrod and Weinstein-Evron 2020; Rey-Rodríguez et al. 2020; Tilby et al. 2022). Here we analyse the Azokh 1 site, which is one of the most complete and, to date, one of the oldest archaeological sites found in the Southern Caucasus.
The main objective of our work is to use rodents to infer the climatic and environmental conditions that prevailed during the formation of the site. We compare our results with previous studies carried out in Azokh 1 Cave, as well as other sites in the region where rodents have been analysed, in order to obtain a framework in which to discuss the palaeoecological conditions where humans pursued their activities in the Southern Caucasus.
Azokh Cave system
Azokh Cave takes its name from the village situated nearby. The site is also known as Azykh Cave. It is located in the Ishkhanaget river valley, Southern Caucasus (39° 37′ 15″ N; 46° 59′ 32″ E) (Asryan et al. 2020). It is an important site for the understanding of human evolution in its archaeological, palaeontological, environmental and ecological context (Fernández-Jalvo et al. 2016b).
The cave is developed in thickly bedded Mesozoic carbonates. This system comprises a succession of sub-rounded chambers oriented NNW to SSE and interconnected for almost 130 m. Several entrance passageways connect the inner parts of the cave to the exterior, but geo-archaeological sediments have only been found in Azokh 1, Azokh 2 and Azokh 5 (Fig. 1). Our research focused on Azokh 1, because this is the only entrance found to date filled with Pleistocene-Holocene sediments (Murray et al. 2016).

Azokh Cave system: a view of exterior of cave system, showing three main entrance passages (Azokh 1, 2 and 5); b plan of Azokh Cave showing the main entrances and internal galleries (Murray et al. 2010); c location of Azokh Cave
Azokh 1
Azokh 1 is a broadly linear chamber 40 m long and 11.5 m high, with a WSW-ENE alignment. The cave has provided evidence of occupation by hominins from the Middle Pleistocene to the Holocene, and is the only well-stratified and dated sequence from this period in the region (Asryan 2015).
It was discovered by M. Huseinov (also named Guseinov by other authors) at the end of the 1950s, and the first excavation started in the 1960s. Excavations of Azokh 1 were carried out from the 1960s to the 1980s, yielding abundant archaeological remains (Guseinov 2010). During this period, the volume of the sediment excavated was about 70% of the cave. Unfortunately, the information and descriptions of the excavation procedures and finds before 1975 were too schematic, making it difficult to interpret the area (Asryan 2015; Fernández-Jalvo et al. 2016b). The current excavations began in 2002, after an initial survey of the site in 1999 and 2001. The excavation project at Azokh Cave was restarted through the collaboration of an interdisciplinary research team. The work focused primarily on the undisturbed, complete sequence of deposits in the upper levels (I–V) at the back of the cave. The systematic recovery and the detailed recording of the material, with new methodologies applied, provided high-quality information on the formation of the site and on human behaviour and evolution (Asryan 2015).
In Azokh 1, nine stratigraphic units are identified, currently separated into two sediment sequences, which can no longer be readily correlated due to the removal of all the intervening stratigraphy (Fig. 2) (Murray et al. 2010, 2016):
-
1.
Sediment Sequence 1: contains Units IX to VI. The test-trench excavation carried out has so far shown that this sequence does not contain archaeological material. The exact age of Sediment Sequence 1 remains unclear, although previous palaeomagnetic results suggest that it is probably older than the upper units and may be of Early Pleistocene age.
-
2.
Sediment Sequence 2: contains Units V to I. The new excavations focus on these units; all of them present an archaeological record, with an age range from the Middle (MIS 9–8) to the Late Pleistocene (MIS 5) (Units V to II), with some Holocene sediments at the top (Unit I) (Fernández-Jalvo et al. 2010, 2016b; Murray et al. 2010; Asryan 2015; Asryan et al. 2017, 2020).

modified from Murray et al. (2016))
Stratigraphy of Azokh 1: a cross section through the entrance passage (facing NW) showing the extent of the cave sediments remaining in the chamber. These are physically separated and are labelled Sediment Sequences 1 and 2. The undated Sequence 1 includes Units VI to IX. The archaeological Sequence 2 (inside the rectangle) includes Units I to V; b cross section (orthogonal to the section shown in a) of Azokh 1 showing the keyhole shape of the passage; c stratigraphy of Sediment Sequence 1; d stratigraphy of Sediment Sequence 2 with dating results of archaeological units (photos
Unit V
Unit V is located at the base of the sequence and is the largest unit, being approximately 4.5 m thick. It is divided into two subunits: Vb (located at the base) and Va (above Vb). Uranium-series dating indicated an age of ca. 200 ka; racemization analysis (D/LAsp) indicated an age closer to ca. 300 ka; and ESR dating suggested an age of 293 ± 23 ka (Murray et al. 2016). For the present study, we decided to refer to it simply as Unit V without considering the subdivisions, notably because the latter may be subject to further revision (Murray et al. 2016).
According to Asryan et al. (2020), the lithic artifacts from this unit include a relatively high presence of retouched flakes and flake fragments, as well as a few cores. There are no unknapped cobbles/pebbles or large tools (bifaces, choppers, chopping tools). The characteristics of the Unit V lithic assemblage indicate that most artifacts made from all the raw materials were introduced as ready-made tools; however, the presence of a refit set may point to some isolated in situ knapping. The techno-typological characteristics and chronology (~ 300 ka) of Unit V share similarities with the Acheulo-Yabrudian techno-culture of the Caucasus. From a broader perspective, this assemblage is late Acheulean or pre-Mousterian, without large cutting tools.
In 1968, a fragment of hominin mandible was discovered in Unit V by M. Huseinov (King et al. 2016). It is a right mandible consisting of the posterior portion of the body and the inferior part of the ramus. The study carried out by Kasimova (2001) suggests that the mandible belonged to a female aged 20–25 years. This specimen has been assigned to Homo heidelbergensis, based on the primitive features (relief of the mylohyoid line) that it displays (King et al. 2016).
According to Van der Made et al. (2016), Unit V is mainly composed of the following large mammals: carnivores (Canis aureus, Crocuta crocuta, Lynx sp., Felis chaus, Panthera pardus, Ursus spelaeus), artiodactyls (Cervus elaphus, Capra aegagrus) and perissodactyls (Stephanorhinus hemitoechus, Stephanorhinus kirchbergensis, Equus hydruntinus, Equus ferus). The faunal spectrum is mainly composed of “interglacial” species; however, during glacial times, the altitude of Azokh Cave (926 m above sea level) would have resulted in a harsh environment around the cave (Van der Made et al. 2016).
According to the herpetofauna of Azokh 1 published by Blain (2016), Unit V is composed of amphibians (Pseudepidalea viridis sensu lato, Ranidae/Hylidae indet. and Pelobates cf. syriacus) and squamates (Pseudopus apodus, Lacerta sp., Eryx jaculus, cf. Coronella austriaca, cf. Elaphe sp., cf. “Coluber” sp. and “Colubrinae” indet.). The herpetofauna seems to be more consistent with a meadow-steppe environment, indicated by species such as Pelobates syriacus.
The study of bats was carried out by Sevilla (2016) and shows an assemblage in Unit V characteristic of an open-ground landscape with steppe vegetation in Unit V, mainly by the presence of Myotis blythii and the genus Rhinolophus. The rodent remains were studied by Parfitt (2016), and these results will be compared with the new study in “Results and discussion” (see “Palaeoecological reconstruction of Azokh 1 Cave Sequence 2” for more details).
Allué (2016) identified a low number of charcoal remains in Unit V, attributed to three taxa only: Prunus, Maloideae and deciduous Quercus sp. These taxa reflect mild, humid environmental conditions.
Unit IV
The contact between the top of Unit V and the overlying Unit IV is 100–130 cm thick but diffuse and irregular. No systematic excavation was carried out in this unit, although bone and charcoal were recovered from the test-trench. ESR dating indicates an age of 205 ± 16 ka for the contact between Units IV and V (Fernández-Jalvo et al. 2016b).
Unit III
The transition between Unit IV and III is marked by a change in the colour of the matrix. This unit is approximately 60 cm thick and contains charcoal, fossil bones and a few stone tools. No date is available for Unit III.
The faunal list identified by Van der Made et al. (2016) for Unit III is composed of carnivores (Panthera pardus, Ursus spelaeus), artiodactyls (Cervus elaphus, Capra aegagrus and Sus scrofa) and perissodactyls (Stephanorhinus hemitoechus and Equus hydruntinus). Unit III can be considered practically sterile in fossil bat remains (Sevilla 2016). Blain (2016) recognized several squamates: Lacerta sp., Eryx jaculus, “Coluber” sp. and Vipera sp. The rodent remains were studied by Parfitt (2016), and his results will be commented on in the “Results and discussion” (see “Palaeoecological reconstruction of Azokh 1 Cave Sequence 2”).
The material from this unit is not abundant enough to draw climatic inferences.
Unit II
Unit II is 100–200 cm thick and includes fossil fauna and stone tools. Sediment diagenesis most likely caused by bat guano has strongly affected the preservation of fossil bones and some stone artifacts in this unit. ESR dating indicates an age of 184 ± 13 ka for the bottom of Unit II and 100 ± 7 ka for the contact between Units I and II (Fernández-Jalvo et al. 2016b).
Lithic artifacts were studied by Asryan et al. (2015, 2020), showing that the operative chain of different raw materials is based primarily on knapping products, including flakes, flake fragments and broken flakes. Levallois technology clearly dominates the core assemblage and knapping products. The small size of most cores and the further retouch of some indicate maximum exploitation and use of good-quality, non-local raw materials. These characteristics imply that almost all artifacts entered this area of Unit II as ready-made tools, with most presenting a clearly predetermined character. There may have been some in situ knapping and retouching activities, as suggested by the presence of knapping waste.
During the excavations in August 2010, an isolated hominin tooth was discovered in this unit and studied by King et al. (2016). It was identified as an upper left first permanent molar that showed typical features of Homo neanderthalensis, on the basis of its morphology (swollen hypocone and skewed shape) and taurodontism, as well as the crown dimensions and root robusticity.
The list of large mammals (Van der Made et al. 2016) is composed of carnivores (Panthera pardus, Ursus spelaeus, Ursus sp., Vulpes vulpes and Canis lupus), artiodactyls (Cervus elaphus, Capra aegagrus, Saiga tatarica, Dama sp. and Sus scrofa) and perissodactyls (Stephanorhinus kirchbergensis). The faunal spectrum suggests that this unit was characterized by a typical “interglacial” temperate environment.
Unit II yielded slightly more bat remains than Unit III (Sevilla 2016). The bat representation in Unit II shows a low diversity and hints at a change towards colder conditions compared to Unit V, indicated notably by the presence of Myotis mystacinus and the absence of Rhinolophus mehelyi (Sevilla 2016).
According to Blain (2016), the fossil herpetofauna from Unit II is composed of amphibians (Pseudepidalea viridis sensu lato, Pelobates cf. syriacus) and squamates (Pseudopus apodus, Lacerta sp., Ophisops elegans, Eryx jaculus, cf. Coronella austriaca, cf. Elaphe sp., cf. “Coluber” sp., the Vipera berus complex and Vipera sp.). This assemblage represents a drier period, with the presence of a representative of the V. berus complex (probably V. ursinii) and the small colubrine Coronella austriaca. A rodent study was carried out by Parfitt (2016); see “Results and discussion” (“Palaeoecological reconstruction of Azokh 1 Cave Sequence 2”).
According to the study carried out by Allué (2016), the charcoal remains from Unit II show a wide diversity of taxa, with Prunus, Acer, Quercus sp., Maloideae, Lonicera sp., Paliurus/Ziziphus sp., Celtis/Zelkova sp., Euonymus sp. and Ulmaceae. This assemblage composition indicates mild, humid environmental conditions.
Unit I
Unit I is 80–135 cm thick. This unit presents considerable disturbance and reworking of the sediments by recent mammal burrowing activity. Palaeolithic faunal and lithic remains were recovered from these burrows together with modern artifacts. Unit I evidences the presence of domestic animals (Sus scrofa, Equus asinus, Equus caballus, Capra hircus) and is considered to be of Holocene age (Van der Made et al. 2016). The amphibians (Pseudepidalea viridis sensu lato) and squamates (Agamidae indet., Pseudopus apodus, Lacerta sp., Ophisops elegans, Lacertidae indet., Eryx jaculus, cf. Coronella austriaca, cf. Elaphe sp., cf. “Coluber” sp., Vipera sp.) present in this unit indicate an environment corresponding to an arid mountain steppe (Blain 2016).
The large-scale bioturbation has greatly interfered with the internal stratigraphic details. Charcoal from the fumier in Unit I provided a radiocarbon age of 157 ± 26 years BP (OxAC1419424). A Soviet coin from around the mid-1960s was also found in 2006, arguing in favour of high bioturbation in this level (Fernández-Jalvo et al. 2016b). Consequently, palaeoenvironmental interpretations must be treated with caution.
In summary, Azokh Cave is an important site for several reasons: (1) it is located on the migration route through the Caucasus used by hominins and fauna at the crossroads between Africa, Europe and Asia; (2) the site was occupied by three hominin species (documented by human remains and lithic artifacts): H. heidelbergensis (Azokh 1 Unit V) (Fernández-Jalvo et al. 2010), H. neanderthalensis (Azokh 1 Unit II) and H. sapiens (Azokh 2 and 5) (Fernández-Jalvo et al. 2016b); (3) it is a site that has been well studied using a multidisciplinary approach (Fernández-Jalvo et al. 2016c) including the study of lithic assemblages (Asryan 2015; Asryan et al. 2017, 2020) as well as of geology, geomorphology (Murray et al. 2016), taphonomy (Andrews et al. 2016; Marin-Monfort et al. 2016), zooarchaeology, palaeontology (Blain 2016; Parfitt 2016; Van der Made et al. 2016) and anthracology (Allué 2016).
The aim of this study is to investigate the palaeoenvironmental and palaeoclimatic conditions that prevailed during the formation of Azokh 1 Cave using these new data and to compare our results with other sites in the study area and with previous works carried out in Azokh 1 Cave. Before undertaking our palaeoecological study, we had to identify the origin of the accumulation and the potential influence of this on the palaeoecological results.
Material and methods
The rodent remains used in this study come from the archaeological excavation campaigns carried out in Azokh 1 in 2003, 2005, 2014, 2015 and 2018. They are from Sediment Sequence 2: Unit V, Units III–IV and Unit II.
The samples comprise disarticulated bones and isolated teeth that were collected in the field by water screening using superimposed 5 and 0.5 mm mesh screens. Most of the samples were sieved in the field by the excavation team, and the resulting residues were air-dried and sorted in the site laboratory. In subsequent years, the small-vertebrate bones were picked out by hand and under a microscope. The material was identified with a binocular microscope, and photos were taken with a Dino-Lite (model AM7915MZTL). In total, 855 samples were analysed: 251 from Unit V, 10 from Units III–IV and 594 from Unit II. The material is housed in the village of Azokh.
Taxonomy
We focused on rodents because these can be considered one of the most useful tools for palaeoenvironmental and palaeoclimatic reconstructions in archaeological sites (e.g. López-García et al. 2010; Maul et al. 2015). The taxonomic identification of the rodent remains is based mainly on molar morphology and measurements (Rey-Rodríguez et al. 2020). We focused especially on the lower m1 (the most discriminant) for Arvicolinae; mandibles and maxillae were used for the taxonomic identification of Cricetidae, Gerbillinae and Murinae.
The remains were identified to species level whenever possible. We used specimens from the modern reference collections of the Natural History Museum of London, the Field Museum of Chicago and the American Museum of Natural History of New York. We also used comparative morphological and biometric data from the literature, notably for Microtus (Coşkun 2016; Kryštufek & Shenbrot 2016; Kryštufek and Vohralík 2009; López-García 2008; Rusin 2017; Shenbrot, Kryštufek & Molur 2016; Tesakov 2016), Clethrionomys (Kryštufek et al. 2009, 2020), Chionomys (Krystufek 2017; Kryštufek and Vohralík 2009; López-García 2011), Cricetulus (Bogicevic et al. 2011; Kryštufek et al. 2017; Sándor 2018), Mesocricetus (Kryštufek and Vohralík 2009), Meriones (Coşkun 1999; Kryštufek and Vohralík 2009; Darvish 2011; Dianat et al. 2017; Stoetzel et al. 2017), Allactaga (Karami et al. 2008; Shenbrot 2009) and Apodemus (López-García 2008; Kryštufek and Vohralík 2009; Bogicevic et al. 2011; Amori et al. 2016; Knitlová and Horáček 2017).
For the genus Ellobius, we based our identifications on data from the literature (Moradi Gharkheloo 2003; Kryštufek and Vohralík 2009; Tesakov 2016), and we also applied geometric morphometric methods (GMM) in order to detect morphological differences and variations in shape and size, following Rey-Rodríguez et al. (2021). The Ellobius first lower molars were all photographed in occlusal view under constant conditions with a digital camera. To investigate the first lower molar size and shape, we combined 2D landmarks (LM) and semi-landmarks (SLM) on the photographs using the TPSdig2 v.2.32 software package (Rohlf 2016) for two-dimensional geometric morphometric analyses. All the analyses were performed with R (R Core Team 2020) using the geomorph (Adams et al. 2020) and Morpho (Schlager 2017) packages.
For the genus Arvicola, we used the molar morphology (Maul et al. 2020) as well as the enamel differentiation index developed by Heinrich (1987), also named SDQ (Schmelzband-Differenzierungs-Quotient). The enamel differentiation index was calculated in accordance with the formula:
SDQ = [Σ(teet × 100/leet)]/N.
teet: posterior part of the triangle.
leet: distal part of the triangle.
N: number of triangles.
where N refers to the number of dentine fields of the studied tooth, teet (trailing edge enamel thickness) refers to the maximum thickness of the posterior enamel band and leet (leading edge enamel thickness) refers to the maximum thickness of the anterior enamel band of each dentine field (Heinrich 1987; Lozano-Fernández et al. 2013).
We also used the standard measurements of the total length (L) and total width (W) of the molars in order to identify some species, particularly among the genera Cricetulus and Mesocricetus.
The quantification of taxonomic frequencies was based on standard indices used in zooarchaeological analyses, including the number of identified specimens (NISP) and minimum number of individuals (MNI) (Weissbrod and Zaidner 2014). The latter was estimated using the most abundant skeletal element present in the assemblage (molars in our case).
Taphonomy
A preliminary study was performed on the rodent remains from Azokh 1 Cave. This was based on the systematic descriptive method that examines the modifications of prey bones induced by predation, focusing on the degree of digestion observed on teeth during identification (Andrews 1990; Fernández-Jalvo et al. 2016a).
Predation is closely related to ecology due to two main factors: the density dependence of prey according to population fluctuations and the trophic preferences of the predator. Accordingly, identifying the predator(s) makes it easier to identify the nature and origin of the assemblage. It is necessary to take these factors into account in undertaking palaeoecological interpretations based on small-mammal accumulations (Fernandez-Jalvo and Andrews 2016).
We performed a preliminary study focusing on a subsample composed of 100 incisors and 90 molars per unit (for Unit V and Unit II), looking at the digestion intensity and broadly defining the category of modification produced by the predator. A taphonomic study was previously performed by Andrews in Azokh 1 Cave (Andrews et al. 2016); in this work, we include an analysis of the new material.
Palaeoenvironmental and palaeoclimatic reconstructions
Habitat weighting method
Palaeoecological interpretations derived from faunal data are based on analyses of community composition (Belmaker and Hovers 2011). The method used for the palaeoenvironmental reconstruction is the habitat weighting method (Evans et al. 1981; Andrews 2006; modified by Blain et al. 2008; López-García et al. 2011), which is based on the current distribution of each taxon in the habitat(s) where it can be found nowadays, according to the values proposed by the IUCN red list. We adapt the method to our studied area (Rey-Rodríguez et al. 2020) in assuming that the Azokh 1 Cave species had equivalent ecological requirements to their present-day relatives. We consider the following types of habitats: forest (Fo), a large area covered with trees; shrubland (Sh), vegetation dominated by shrubs; grassland (Gr), an open area covered with grass; desert (De), an area with little precipitation and no vegetation cover; wetland (We), an area where water covers the soil; steppe (St), a dry grassy plain; and rocky (Ro), a rocky or stony substrate. Each species has a score of 1.00, which is divided between the habitats where the species can be found at present (Table 1).
Bioclimatic model
In order to reconstruct the past climate at Azokh 1 Cave, we applied the bioclimatic model (BM), which was developed by Hernández-Fernández (2001) and updated by Royer et al. (2020). This method is based on the faunal spectrum, assuming that small- and large-mammal species can in general be ascribed to ten different climates (Hernández-Fernández 2001; Hernández-Fernández et al. 2007). It was first necessary to calculate the climatic restriction index (CRIi = 1/n, where i is the climatic zone where the species appear and n is the number of zones where the species is present) and the bioclimatic component (BCi = (Σ CRIi) 100/S, where i is the climatic zone and S is the number of species). All statistical analyses were performed using the R software package (R Core Team 2020) using the script PalBER developed by Royer et al. (2020) in order to calculate the bioclimatic spectra and infer the climatic zone.
The different climatic groups defined by Hernández-Fernández (2001) and Hernández-Fernández et al. (2007) present in this work are as follows: II/III, transition tropical semi-arid; III, subtropical arid; IV, subtropical with winter rains and summer droughts; VI, typical temperate with winters that are cold but not very long, but summers that are cool; VII, arid-temperate with large temperature contrasts between winter and summer; VIII, cold-temperate with cool summers and long cold winters (boreal) and IX, arctic (Table 2).
By means of the BM, we were able to estimate various climatic parameters, such as the mean annual temperature (MAT), the mean temperature of the coldest month (MTC), the mean temperature of the warmest month (MTW) and the mean annual precipitation (MAP). This method enabled us to infer the palaeoclimatic conditions that prevailed during the Middle and Late Pleistocene at Azokh 1 Cave.
Results and discussion
Systematics, current distribution and ecology
A total of 434 molars were identified following the identification keys, representing a minimum number of 237 individuals (MNI) (Table 3). This sample comprises the new remains from the 2003, 2005, 2014, 2015 and 2018 excavation campaigns, the previous material having been analysed by Parfitt (2016), as later discussed in “Palaeoecological reconstruction of Azokh 1 Cave Sequence 2”.
Order Rodentia Bowdich, 1821
Family Cricetidae Fischer, 1817
Genus Cricetulus Milne-Edwards, 1867
Cricetulus migratorius Pallas, 1773
Material: 10 isolated teeth. Unit II: eight isolated teeth, one left lower m1, two right lower m1, one right upper M1, one left upper M1, one right upper M2 and two left upper M2. Unit V: two isolated teeth, one left lower m1 and one right maxilla with M1 and M2.
Discussion: the first molars (m1 and M1) are brachyodont and cuspidate, with two longitudinal series of cusps. Each series of cusps consists of three pairs. The m1 and M1 are the largest, and the m3/M3 the smallest. The lower m3 only has two pairs of cusps (Kryštufek and Vohralík 2009). We identified Cricetulus migratorius (Fig. 3.1) in Unit II and Unit V of Azokh 1 Cave in accordance with the measurements (Table 4) and the identification keys for molars based on the morphology and arrangement of the tubercles and cusps provided by Kryštufek and Vohralík (2009). We also drew comparisons with the reference collection from Iran, Afghanistan and Azerbaijan housed in the Natural History Museum of London regarding the morphology of the teeth. The grey hamster, or migratory hamster, is the smallest hamster species (Bogicevic et al. 2011; Sándor 2018). We measured the teeth, and the results indicate that our specimens are the same size as those from the reference collection, excluding the small-sized hamster Allocricetus (Parfitt 2016).

Some rodents identified at Azokh 1 Cave. 3.1 Cricetulus migratorius, Azokh 1 Cave, 2003, Unit V, D45, Z: 224, right M1 and M2, number 221. 3.2 Mesocricetus brandti, Azokh 1 Cave, 2014, Unit II, E49, Z: 320–330, left m1, number 74. 3.3 Clethrionomys glareolus, Azokh 1 Cave, 2018, Unit II, G56, Z: 46.48, left m1 occlusal and lingual view, number 703. 3.4 Arvicola ex. gr. persicus, Azokh 1 Cave, 2014, Unit II, G51, Z: 330–340, left m1, number 536. 3.5 Chionomys gud, Azokh 1 Cave, 2003, Unit V, D46, Z: 111, left m1, number 349. 3.6 Chionomys nivalis, Azokh 1 Cave, 2014, Unit II, F48, Z: 330–340, left m1, number 529. 3.7 Microtus gr. arvalis-socialis, Azokh 1 Cave, 2014, Unit II, G53, Z: 330–340, right m1, number 5. 3.8 Microtus (Terricola) sp., Azokh 1 Cave, 2005, Unit V, E40, Z: 136, left m1, number 43. 3.9 Microtus (Terricola) sp., Azokh 1 Cave, 2005, Unit V, D45, right m1, number 170. 3.10 Ellobius cf. lutescens, Azokh 1 Cave, 2015, Unit II, F51, Z: 320–330, left m1 occlusal and lingual view, number 226. 3.11 Meriones gr. persicus- tristrami, Azokh 1 Cave, 2003, Unit V, D45, right m1 occlusal and lingual view, number 169. 3.12 Meriones gr. dahli-libycus, Azokh 1 Cave, 2003, Unit V, D45, Z: 224, left M1, number 222. 3.13 Apodemus (Sylvaemus) sp., Azokh 1 Cave, 2014, F52, Z: 330–340, left m1, number 80. 3.14 Allactaga cf. williamsi, Azokh 1 Cave, 2018, H49, G50, right m1, number 635. Scale 1 mm
Habitat and distribution: Cricetulus migratorius extends from eastern Europe through Russia and Central Asia to Mongolia and western China (Kryštufek et al. 2017; Kryštufek and Vohralík 2009). The habitats of this species are mostly dry grasslands, steppes and semi-deserts. Arid areas with relatively sparse vegetation are preferred (Kryštufek et al. 2017; Maul et al. 2015).
Cricetulus sp.
Material: 10 isolated teeth. Unit II: six isolated teeth, two left lower m1, two right lower m2 and two left lower m2. Unit V: four isolated teeth, two left lower m1, one right lower m1 and one left lower m2.
In our record, we recognized as Cricetulus sp. broken teeth that could not be measured and identified to the species level.
Genus Mesocricetus Nehring, 1898
Mesocricetus cf. brandti Nehring,
Material: 11 isolated teeth. Unit II: five isolated teeth, one left lower mandible with m1 and m2, one left lower m1, three left upper M2 and one left upper M3. Unit V: six isolated teeth, one right lower m1, one left upper M1, three left upper M2 and one right upper maxilla with M1 and M2.
Discussion: the specimens from Azokh 1 Cave are attributed to Mesocricetus brandti (Fig. 3.2) on the basis of the morphology of the teeth, the disposition of the cusps and the size. The molars are significantly larger than Cricetulus migratorius, but smaller than representatives of the Cricetus genus. The first molars have six tubercles; the second and third molars only four. The largest molars are m1 and M1, whereas m2/M2 and m3/M3 are reduced (Kryštufek and Vohralík 2009). In the region of Azokh, Mesocricetus raddei is also found, presenting a similar morphology of the teeth to Mesocricetus brandti, but compared with the NHM reference collection of the latter, Mesocricetus raddei is bigger (Table 5).
Habitat and distribution: Mesocricetus brandti has the largest distributional area of the species belonging to the genus Mesocricetus, ranging from Anatolia, Transcaucasia and southeast Dagestan to northwest Iran (Qazvin in the east, Lorestan in the south; Lay 1967). This species is found at altitudes ranging from sea level up to 2600 m. However, the primary range is from 1000 to 2200 m. Mesocricetus brandti is found in arid and semi-arid steppe habitats in lowlands and in mountainous areas (Kryštufek, Yigit & Amori 2015; Kryštufek and Vohralík 2009; Neumann et al. 2017).
Subfamily Arvicolinae Gray, 1821
Genus Clethrionomys Tilesius, 1850
Clethrionomys glareolus Schreber, 1780
Material: four isolated teeth. Unit II: one right lower m1 and three left lower m1.
Description and discussion: In our sample, we identified Clethrionomys glareolus (Fig. 3.3) on the basis of the presence of roots and the morphology of the first lower molar (m1) (Kryštufek et al. 2020). In our study region, there are only two genera of Arvicolinae with rooted molars: Clethrionomys and Ellobius, and these can be easily differentiated by their molar morphology. In Clethrionomys, the m1 displays two roots, and has five triangles with a highly variable anteroconid complex; however, T4–T5 do not vary. T5 is integrated into the anterior cap or more rarely entirely isolated from it; the anterior cap is rarely simple and oval but mostly broadly confluent with T5. There are three or four re-entrant angles on the inner side and three on the outer side (Kryštufek & Vohralík 2005).
Habitat and distribution: The bank vole (Clethrionomys glareolus) is widely distributed in the Palaearctic area, which stretches from the British Isles through continental Europe and Russia to Lake Baikal. It was present at Azokh 1 Cave in the past; however, this species no longer exists in the study region today.
Regarding its habitat, it is present in all kinds of woodland, preferring densely vegetated clearings, woodland edges and river and stream banks in forests. It can also be found in scrub, parkland and hedges (Bergl et al. 2017).
Arvicola ex. gr. persicus de Filippi, 1865
Material: one isolated tooth. Unit II: one left lower m1.
Description and discussion: recent works have shown that the Iranian phylogroup of water voles can be considered a valid species, named Arvicola persicus (Fig. 3.4) (Chevret et al. 2020; Mahmoudi et al. 2020). We chose to follow Maul et al. (2020) in using the name Arvicola ex. gr. persicus. In our material, only one m1 could be attributed to the genus Arvicola based on its size and morphology. The m1 is rootless, with clear Mimomys-type enamel differentiation and crown cementum filling the re-entrant angle. The m1 consists of an anterior cap (AC), five triangles (T) and a posterior lobe (PL). Enamel thickness (SDQ) is a parameter applied to lower molars for the identification of Arvicola species. In our material, we could only use SDQ5 on the triangles, because the AC and the PL were digested. We obtained a value of 124, which assigns our sample to Arvicola ex. gr. persicus according to Maul et al. (2020). The shape of the anteroconid complex also corresponds to this species, with a BRA3 that is not as deep as in A. nahalensis. The Mimomys-type enamel differentiation excludes the possibility that it could be A. amphibius, the other current representative of the genus in the region (Mahmoudi et al. 2020). The SDQ values show that there is a close relation between A. persicus from Iran and Turkey, A. italicus from Italy and A. sapidus from Spain, because all of these display a SDQ value > 100 (Maul et al. 2020). In our sample, Arvicola ex. gr. persicus is the only possible assignation, considering the geographic distribution of the other taxa.
Habitat and distribution: the precise ecology and distribution of Arvicola persicus remain to be clarified (Mahmoudi et al. 2020). However, the group of water voles, within which our species is included, is always rather scarce in dry areas, most probably because these animals require suitable aquatic habitats, i.e. the nearby presence of permanent water bodies such as rivers, streams and marshes. We assume that A. persicus has a semi-aquatic lifestyle, indicating the presence of water habitats (Harrison and Bates 1991; Maul et al. 2020; Mahmoudi et al. 2020). Nowadays, there is a river called the Ishkhanaget near the site at the bottom of the mountain, which may very well have existed in the past and favoured the presence of Arvicola around the site.
Genus Chionomys Miller, 1908
In our sample, eight isolated teeth show the typical traits of the genus Chionomys. The enamel is of the Microtus type with cement in the re-entrant angles (Kryštufek 2017; Kryštufek and Vohralík 2005; López-García 2011). These specimens differ from Microtus in that they only have five triangles, and an AC with a typical rounded morphology.
Chionomys gud Satunin, 1909
Material: three isolated teeth. Unit V: one isolated right lower m1 and two isolated left lower m1.
Description and discussion: in Unit V we observed three m1, which mainly show four closed triangles, with the wear facets of triangles T5 and T6 confluent with the anterior cap; they are typical of the Caucasian snow vole, Chionomys gud (Fig. 3.5) (Kryštufek and Vohralík 2005; Sözen et al. 2009). These specimens were also compared to the Chionomys roberti specimens from the NHM of London and were found to be quite different in appearance, notably in the configuration of the triangles and the morphology of the AC.
Habitat and distribution: Chionomys gud is closely associated with rocky habitats. It occurs in alpine meadows, in sparse fir and spruce forests and in the valleys of brooks and small rivers. It prefers more mesic habitats than Chionomys nivalis (Kryštufek and Vohralík 2005).
Chionomys gud is endemic to the Caucasus and the easternmost part of the Pontic Mountains of Turkey, and is not represented today in Armenia (Kryštufek 1999).
Chionomys nivalis Martins, 1842
Material: four isolated teeth. Unit II: one right lower m1 and three left lower m1.
Description and discussion: in Unit II, four first lower molars display five closed triangles and an anteroconid complex (AC) with a morphology characteristic of the nivalis morphotype (Fig. 3.6), where triangles T6 and T7 are reduced and tightened at their base, and the anterior cap is of an arrowhead or oval shape, inclined towards the labial part (Nadachowski 1991; Kryštufek and Vohralík 2005).
Habitat and distribution: Chionomys nivalis has a global distribution extending from southwestern Europe through southeastern Europe to the Caucasus, Turkey, Israel, Lebanon, Syria and Iran (Kryštufek 1999).
Regarding the habitat of Chionomys nivalis, the species is present in open rocky areas, typically above the tree line and with scarce vegetation cover (Amori 1999).
Chionomys sp.
Material: one isolated tooth. Unit II: one isolated left lower m1.
We opted for the classification Chionomys sp. because of the lack of discriminant characters in the broken tooth.
Genus Microtus Schrank, 1798
The molars identified in the genus Microtus are hypsodont and arhizodont, with crown cementum in the re-entrant angles and Microtus-like enamel differentiation. Eight species of Microtus are currently recognized in the southern Caucasus: Microtus arvalis, Microtus daghestanicus, Microtus levis, Microtus majori, Microtus nasarovi, Microtus schelkovnikovi, Microtus schidlovskii and Microtus socialis (Aşan Baydemir and Duman 2009; Golenishchev et al. 2019; Firouz 2005). Microtus m1s are characterized by four outer and five inner re-entrant angles, with a posterior lobe (PL), seven closed triangles (T) and an anterior cap (AC). In most of our material, triangles T4 and T5 are closed, ruling out the subgenus Terricola.
Microtus is the most abundant taxon in all the sequence, but identifying the species was difficult because we lack good reference collections and comparative data from the literature. However, we were able to differentiate two groups, based on the morphology of the lower m1.
Microtus gr. arvalis-socialis
Material: 296 isolated teeth. Unit II: 135 isolated teeth; 75 right lower m1, 60 left lower m1. Units III–IV: seven isolated teeth; five left lower m1 and two right lower m1. Unit V: 154 isolated teeth; 77 right lower m1 and 77 left lower m1.
Description and discussion: the species (Fig. 3.7) included in this group are Microtus arvalis, Microtus socialis and Microtus guentheri. The triangles from T1 to T5 are closed and opposite one another. T6–T7 are confluent and parallel to BRA4 and LRA5 (Kryštufek and Vohralík 2005).
Habitat and distribution: Microtus species belonging to the group arvalis-socialis mostly occur in steppe, shrubland or semi-desert habitats (Fernandez-Jalvo et al. 2016). According to the IUCN Red List, all of them are currently present in the southern Caucasus.
Microtus ( Terricola ) spp.
Material: 21 isolated teeth. Unit II: seven isolated teeth; three right lower m1 and four left lower m1. Units III–IV: one isolated tooth; one left lower m1. Unit V: 13 isolated teeth; seven right lower m1 and six left lower m1.
Description and discussion: the second group is Microtus (Terricola) spp., (Fig. 3.8 and 3.9) characterized by broadly confluent T4 and T5, forming the pitymyan rhombus (Kryštufek and Vohralík 2005). In our sample, we identified at least two species, based on the size and the morphology of the AC. There are several Terricola species in the region: Microtus daghestanicus, Microtus nasarovi, Microtus majori and Microtus schidlovskii. All of them are very close to one another in their molar morphology, and without a good reference collection, it is not possible to identify them to species level.
Habitat and distribution: Caucasian voles, belonging to the Microtus (Terricola) spp. group, are found in a quite wide range of habitats: Microtus daghestanicus and Microtus nasarovi prefer pastures, alpine meadows and steppe; Microtus majori favours clearings in forests and shrubland, as well as alpine pastures; and Microtus schidlovskii is more closely associated with xerophytic steppes and meadow steppes. All of them are currently present in the southern Caucasus, according to the IUCN Red List.
Genus Ellobius Fischer, 1814
Ellobius cf. lutescens Thomas,
Material: 36 isolated teeth. Unit II: 22 isolated teeth; 10 right lower m1 and 12 left lower m1. Units III–IV: one isolated tooth; one left lower m1. Unit V: 13 isolated teeth; seven right lower m1 and six left lower m1.
Description and discussion: 36 isolated teeth show the typical traits of the genus Ellobius (Miller 1896; Hinton 1962; Kretzoi 1969; Coşkun 2016; Kryštufek & Shenbrot 2016; Kryštufek and Vohralík 2005; Rusin 2017; Kryštufek & Molur 2016; Tesakov 2016).
The Ellobius lower m1 is composed of the anterior cap (AC), five triangles (T) with three buccal and four labial re-entrant angles, and one posterior lobe (PL). Ellobius molars are notably characterized by the presence of roots that are clearly visible in adults and old individuals. Moreover, Ellobius molars lack cement in the re-entrant angles (Coşkun 2016).
For modern representatives, most of the discriminant characters are based on skull morphology (Kaya et al. 2018) and external characters (Kryštufek and Vohralík 2005), whereas in an archaeological context we mostly rely on isolated molars or broken maxillae and mandibles. The lower m1 is quite similar among the different current Ellobius species (Ellobius fuscocapillus, Ellobius lutescens and Ellobius talpinus), but some specific morphological characters have been pointed out in previous studies: the AC is broad in Ellobius lutescens, narrow in Ellobius talpinus and elongated in Ellobius fuscocapillus (Maul et al. 2015); the distance between T4 and T5 (W) and the total length (L) differ among the species, Ellobius fuscocapillus showing the largest teeth and Ellobius talpinus the smallest (Rey-Rodríguez et al. 2020). However, these morphological and biometric characters are not always clear or reliable, and the use of more powerful methods is necessary to obtain reliable identifications. To this end, we applied geometric morphometric analyses to m1 molars following the methodology established by Rey-Rodríguez et al. (2021) for Ellobius. We used specimens from the modern reference collections of the Natural History Museum of London, the Field Museum of Chicago and the American Museum of Natural History of New York, and the archaeological sample is from Azokh 1 Cave (Unit II and Unit V).
The PCA performed on the normalized landmarks and sliding semi-landmarks of the right lower molar reveals significant differences among the analysed species, the first two principal components (PCs) accounting for 57.3% of the total variance (Fig. 4a).

a Principal component analysis of the normalized landmarks and sliding semi-landmarks and shape configuration at the extreme ends of the PCs. b First principal component of size and shape including the reference collection and Azokh 1 Cave material
The main variation along PC1 (41.4%) relates to the morphology of the anterior cap, which is more flattened for the positive values and more rounded for the negatives ones. Ellobius talpinus occurs in the positive part of the PC1 axis whereas Ellobius fuscocapillus and Ellobius lutescens are located in the negative part, reflecting a broader and more rounded AC. Along the PC2 (15.9%) scores, the positive values reflect a rounded and less developed AC turned to the labial side, whereas the negative values show an elongated AC turned to the buccal side. In PC2, there is no clear differentiation between the three species. However, it is worth noting that the Ellobius lutescens specimens are located principally in the upper half of the graphs, showing negative values in PC2. We can thus confirm the species identifications and corroborate that Ellobius talpinus is not present in the archaeological samples.
To look at possible allometric effects on the samples, we performed a linear regression of the PCs onto the log of the size of the centroid (Mitteroecker et al. 2015). Including size allows us to discriminate E. lutescens from E. fuscocapillus, the latter showing greater dimensions (Fig. 4b). E. talpinus presents a wide size range overlapping the two latter species and is only distinguished by the molar conformation (PC1). The archaeological remains from Azokh 1 Cave are distributed among E. lutescens and E. fuscocapillus (Fig. 4b).
Other criteria can allow species to be identified, such as the configuration of the anterior cap (Maul et al. 2015), with the transition between T4 and T5 being narrower in E. lutescens than in E. fuscocapillus, leading to a smaller and more closed AC in E. lutescens. By combining GMM and PCA analyses with morphological criteria (AC and distribution of the triangles), we were able to attribute the material from Azokh 1 to the species Ellobius cf. lutescens (Fig. 3.10).
Habitat and distribution: Ellobius species frequent steppes, grasslands and semi-deserts in eastern Europe and central Asia; these fossorial species are specialized in subterranean life (Coşkun 2016; Kryštufek and Vohralík 2005). Ellobius lutescens (western mole vole) is distributed in northwestern Iran, Iraq, Azerbaijan, Armenia and eastern Anatolia (Thomas 1905; Ellerman and Morrison-Scott 1951; Darlington 1957; Osborn 1962; Walker 1964; Lay 1967; Hassinger 1973; Roberts 1977; Corbet 1978; Corbet and Hill 1991; Coşkun 1997; Nowak 1999; Wilson and Reeder 2005, 2017; Kryštufek and Shenbrot 2016).
Subfamily Gerbillinae Gray, 1825
Genus Meriones Illiger, 1811
The genus Meriones is one of the most diverse among the tribe Gerbillini in the Palaearctic region, particularly in arid regions of Asia (Darvish 2011; Denys 2017). The Meriones species currently reported in the southern Caucasus are Meriones dahli, Meriones libycus, Meriones persicus, Meriones tristrami and Meriones vinogradovi (Dianat et al. 2017; Fernández-Jalvo et al. 2016b; Kryštufek and Vohralík 2009). In our archaeological material from Azokh 1 unit V, III–IV and II we have identified at least two different groups of Meriones species based on their molar size and morphology, as well as the number of roots on the molars.
Meriones gr. persicus- tristrami
Material: nine isolated teeth. Unit II: two isolated teeth; two right lower m1. Units III-IV: one isolated tooth; one left upper M1. Unit V: six isolated teeth; three right lower m1 and three left lower m1.
Description and discussion: the part of the material from Azokh 1 Cave attributed to the genus Meriones (Fig. 3.11) displays the typical morphology of this group, including semi-hypsodont molars with prismatic enamel triangles linked by a longitudinal crest and with no trace of cusps. In our sample, we identify first upper molars (M1) with three roots, which is characteristic of Meriones persicus and Meriones tristrami. Unfortunately, the dental morphology of these two species is very similar; their m1 and M1 display three roots, the second molars (m2 and M2) have two transverse plates and two roots, whereas the third molars (m3 and M3) are simple and rounded with a single root (Coşkun 2016; Kryštufek and Vohralík 2009).
Habitat and distribution: the genus Meriones is distributed across North Africa, Central Asia, Turkey and Pakistan (Darvish et al. 2014; Wilson et al. 2017). It lives mostly in dry steppes of short or tall grass, on open hillside, among rocky outcrops in steppes, or in open dry meadows.
The distribution of Meriones persicus ranges from the Caucasus (including the southeastern foothills of the Lesser Caucasus and the Talysh Plateau in Azerbaijan) in the west, through northeastern Iraq and Iran to Turkmenistan, Afghanistan (Habibi 2004) and Pakistan, where it is widely distributed. The species generally occurs in arid, rocky or mountainous regions (Kryštufek and Vohralík 2009; Molur & Sozen 2016).
The habitat of Meriones tristrami is limited to areas with 100 mm of rainfall annually; the species needs well-drained soil although it avoids rocky conditions. It has been found in Armenia, Azerbaijan, Georgia, Iran, the Islamic Republic of Iraq, Israel, Jordan, Lebanon, the Syrian Arab Republic and Turkey. Its habitat is mainly shrubland and desert (Kryštufek and Vohralík 2009; Sozen et al. 2016).
Meriones gr. dahli - libycus
Material: five isolated teeth. Unit II: one isolated tooth; one right upper M1. Unit V: four isolated teeth; one right lower m1 and three left upper M1.
Description and discussion: in our sample, we also identified upper first molars with two roots, which are characteristic of Meriones dahli and Meriones libycus. We provisionally attribute our specimens to this group (Fig. 3.12) in the light of the number of roots and the morphology of M1, pending a revision of the Middle Eastern species of the genus.
Habitat and distribution: Meriones dahli is known in the border area of Armenia and Turkey, and possibly Azerbaijan. It has been found only in local desert habitats (Kefelioglu et al. 2008).
Meriones libycus has a wide global range, occurring in North Africa (from Western Sahara and Mauritania to Egypt) and in Asia (from the Eastern Arabian Peninsula to China). M. libycus occupies semi-desert and desert habitats; it is most abundant in unflooded river plains and is often found close to wadis and dayas (Granjon 2016).
Meriones sp.
Material: nine isolated teeth. Unit II: four isolated teeth; one right lower m1, two right upper M1 and one left lower m1. Unit V: five isolated teeth; three right lower m1 and two left lower m1.
Description and discussion: we identify as Meriones sp. broken or digested molars without clear discriminating characters.
Family Muridae Illiger, 1811
Genus Apodemus Kaup, 1829
Seven Apodemus species are currently recognized in the southern Caucasus: A. hyrcanicus, A. flavicollis, A. witherbyi, A. ponticus, A. agrarius, A. mystacinus and A. uralensis, all belonging to the Sylvaemus subgenus (Jangjoo et al. 2011).
Apodemus ( Sylvaemus ) sp.
Material: 10 isolated teeth. Unit II: seven isolated teeth; two right lower m1, one left mandible with an m1, m2 and m3, one left upper M1 and three left lower m1. Unit V: three isolated teeth; one right upper M1, one right maxilla with an M1 and M2 and one right lower m1.
Description and discussion: the first lower molars (m1) from our material show the traits characteristic of the genus Apodemus (Fig. 3.13): the occlusal surface is low, with six main cusps and a small anterior mesial tubercle; the anterolabial and posterolabial cusps of m1 converge in an X-shape; the posterior cusp of m1 is low, rounded and well developed, with two or three secondary cusps in the labial part. Identifications to the species level are difficult due to a lack of documented and genetically typed reference collections. Moreover, comparative morphological data available in the literature (López-García 2008; Siahsarvie and Darvish 2008; Kryštufek and Vohralík 2009; Bogicevic et al. 2011; Darvish et al. 2015; Amori et al. 2016; Knitlová and Horáček 2017) are sometimes unclear and/or display high variability, and some species are not documented.
Habitat and distribution: the genus Apodemus has a large distribution range extending from Great Britain across much of continental Europe to the Urals. It also extends east through Turkey to western Armenia, the Zagros Mountains of Iran and south to Syria, Lebanon and Israel. It inhabits a variety of woodland and bushy habitats (Amori et al. 2016).
Family Dipodidae Fischer, 1817
In the Caucasus, three species are currently represented: Allactaga williamsi, Allactaga major and Allactaga elater.
Genus Allactaga Cuvier, 1837
Allactaga cf. williamsi
Material: four isolated teeth. Unit II: two isolated teeth; one right lower m1 and one left upper M1. Unit V: two isolated teeth; one right upper M3 and one left lower m2.
Discussion: this rodent group is poorly known in the Middle East (Shenbrot 2009; Naderi et al. 2011; Dianat et al. 2013), and sometimes there is a size overlap between the species. In our sample, we only found six remains attributed to Allactaga. The remains are not damaged and were compared with specimens from the reference collection of the Natural History Museum of London and with data from the literature. We have an M1 with four roots, a lower m1 with two roots and an M3 with three roots. All of them share the same morphological features as Allactaga williamsi (lower molars narrower than upper ones, third molars less reduced, first lower molar with a small fold in front and two folds on either side). In comparison, Allactaga major and Allactaga elater have an M1 with only three or two roots (Kryštufek and Vohralík 2009; Markova 1982). We thus tentatively attribute the remains from Azokh 1 to Allactaga cf. williamsi (Fig. 3.14), awaiting further studies on the molar morphology among the species of this genus.
Habitat and distribution: Allactaga williamsi is distributed in Anatolia (Turkey), the Caucasus (Armenia, Azerbaijan, Iran and Turkey) and northwestern Iran, with an isolated population in central Afghanistan. As regards its habitat, this species occurs in steppes with sparse vegetation (Eken et al. 2016).
Allactaga sp.
Material: two isolated teeth. Unit V: two isolated teeth; two lower m3.
Description and discussion: we classified as Allactaga sp. broken teeth whose roots could not be counted or whose molar morphology could not be seen properly.
Taphonomic study
For the taphonomic study, we focused on a subsample composed of 380 incisors and molars (100 incisors and 90 molars for each unit). We considered that it was necessary to identify the origin of the accumulation and the potential influence of this on any interpretation before a palaeoecological study could be undertaken.
A considerable percentage of the molars and incisors show evidence of digestion: 26.83% in Unit V and 22.22% in Unit II (Fig. 3, Table 6). According to Andrews (1990) and Fernández-Jalvo et al. (2016a), the intensities of digestion are mostly light (14.53% in Unit V and 17.50% in Unit II) or moderate (5.77% in Unit V and 8.37% in Unit II) in both units (Table 6), suggesting that the remains were probably accumulated by a category 1 predator such as the barn owl (Andrews 1990; Fernández-Jalvo et al. 2016a). Such owls have opportunistic hunting habits and are sedentary, so their prey spectrum is assumed to be a good representation of the ecosystem in which they live, taking into account their nocturnal habits and their size.
These results are in agreement with previous taphonomic work published by Andrews et al. (2016), where the molars and incisors showed similar levels of digestion. The digestion levels are mainly light or moderate, indicating that the sample was probably accumulated by Tyto alba, which produces low degrees of digestion except at its nest site (Andrews et al. 2016).
Note that Parfitt (2016) found Rattus sp. remains in Unit V (Table 7), which may raise some taphonomic issues. However, we were unable to verify the identification of these remains, so we cannot draw any conclusion on this point for now.
Palaeoecological reconstruction of Azokh 1 Cave Sequence 2
The small-mammal sequence of Azokh 1 Cave is quite diverse, with at least 13 taxa identified in this work (Table 3). Compared to previous studies (Parfitt 2016), the new taxa we identified include (Table 7) Mesocricetus brandti, Arvicola ex. gr. persicus, Ellobius cf. lutescens, Meriones gr. persicus-tristrami, Meriones gr. dahli-libycus and Allactaga cf. williamsi. Conversely, some species identified by Parfitt were not found in our samples: Allocricetus sp., Rattus sp., Mus cf. macedonicus and Dryomys nitedula.
There is no significant turnover between the studied units, which contain similar rodent assemblages (Table 3). The most abundant taxon in all the units is Microtus gr. arvalis-socialis (77 individuals in Unit V, seven in Units III–IV and 75 in Unit II), followed by Ellobius cf. lutescens (seven individuals in Unit V and 12 individuals in Unit II) and Microtus (Terricola) spp. (seven individuals in Unit V and four individuals in Unit II). Most of the species identified at Azokh 1 Cave are still present in the area today, but there are some exceptions, such as Chionomys gud, Ellobius cf. lutescens and Clethrionomys glareolus.
Units III–IV do not yield enough material to draw palaeoecological inferences (MNI < 30). For Units V and II, the bioclimatic model shows similar results (Table 8). The BM based on the small mammals from Azokh 1 suggests mean annual temperatures and precipitation similar to the present (Table 8), it being a bit colder and drier in Unit II than in Unit V. The climate seems to be relatively warm-temperate, with continental conditions in both units.
The habitat weighting method (Fig. 5) shows an environment mainly composed of steppes and shrublands, notably indicated by the presence of Meriones spp. and Mesocricetus brandti. In Unit V, grasslands (Allactaga cf. williamsi), desert (Cricetulus migratorius) and rocky habitats (Chionomys gud, C. nivalis) are also well represented. Unit II presents a higher representation of wetland habitat than Unit V, due to the presence of Arvicola ex. gr. persicus. The proportion of forest in Unit II is also greater than in Unit V, as indicated by the presence of Clethrionomys glareolus and Apodemus (Sylvaemus) sp.

Results obtained by the habitat weighting method for Azokh 1 Cave (Unit V and Unit II)

a Location of Qesem Cave, Misliya Cave and Azokh 1. b Climatic map of the studied region, obtained from Zittis (2015)
The current vegetation map of Azokh shows the presence of a broad belt of semi-xerophilous woodland in the lowlands (Manuk 2010). Further to the east are belts of sagebrush steppe and sagebrush desert, and both would have formed part of the habitat ranges of larger mammals and birds of prey (Andrews et al. 2016).
In summary, the BM does not reveal significant palaeoclimatic differences between Unit V, Unit II and the current values. The HW shows some differences, especially in the proportions of forest and wetlands. However, the environmental data from both Units V and II are consistent with steppe environments with shrubland, rocky areas and arid conditions.
Comparisons with other palaeoenvironmental proxies from Azokh 1 Cave
The results we obtained with the rodent assemblages were compared with previous studies carried out at Azokh 1 Cave, where palaeoenvironmental proxies were estimated using large mammals, small vertebrates and archaeobotanical data (Table 9). We paid special attention to the study undertaken by Parfitt (2016), who analysed most of the small mammals collected between 2002 and 2009 in Sediment Sequence 2 from Unit V to Unit I, as well as to the palaeoecological interpretations proposed by Andrews et al. (2016).
Most of the previous studies of the small mammals from Azokh have been based on Unit V, which yielded the most abundant material. Parfitt’s results showed subtle differences in faunal composition throughout the Middle (Unit V) to Late Pleistocene (Units III and II) sequence of Azokh, indicating changes in aridity and temperature, combined with fluctuations in woodland cover and the proximity of trees to the site (Parfitt 2016). Our study made it possible to add one new species (Arvicola ex. gr. persicus) to the faunal list established by Parfitt (2016) and to specify the identifications for Ellobius cf. lutescens, Meriones gr. persicus-tristrami, Meriones gr. dahli-libycus and Allactaga cf. williamsi, which were previously limited to the genus level. Moreover, our study allowed us to provide more consistent data for Unit II, which was the least representative unit among the samples in Parfitt (2016).
According to Marin-Monfort et al. (2016), the large mammals were accumulated both by carnivores and humans. Taxonomic identifications have been performed by Van der Made et al. (2016), with at least 29 species represented in the sequence (Table 9). Some of the species, such as Cervus elaphus, are present in the whole sequence. Ursus spelaeus was identified in all the Pleistocene units, and a taphonomic study revealed that cave bear remains were relatively complete, with some bones in anatomical connection, suggesting that the bears living in the cave were using it as a den (Marin-Monfort et al. 2016). The remains of other mammals are, in most units, extremely fragmented, and mainly represented by teeth, horn/antler and foot bones. According to Andrews et al. (2016), using the habitat weighting method, Units V–II have the highest index value for deciduous woodland and also for Mediterranean evergreen woodland, as indicated by Stephanorhinus kirchbergensis, Sus scrofa and Dama sp. Unit I differs from the others by its more steppic, arid signal.
The herpetofauna of Azokh 1 is composed exclusively of extant genera and species (Table 9), most of them belonging to thermophilous and xerophilous forms (e.g. Pelobates syriacus, Agamidae, Pseudopus apodus, Ophisops elegans, Eryx jaculus, Elaphe sauromates, among others). The anuran Pseudepidalea viridis has a wide ecological tolerance. Most taxa frequent open wooded or bushy areas, such as Pseudopus apodus or Ophisops elegans (Blain 2016). According to Blain (2016), the faunas from Units II to I reflect a drier period, including a representative of the Vipera berus complex (probably V. ursinii) and the small colubrine Coronella austriaca. In Unit V, the environment seems to be more consistent with a meadow steppe. The results obtained with amphibians and squamates are consistent with those obtained with rodents and indicate an environment mainly composed of desert and steppes, especially in Unit V.
The bat fauna from Azokh 1 is composed of extant genera (Table 9), and the main habitats that it reflects are mountains steppes, followed by mountain grassland. Unit V is dominated by species with Mediterranean or humid affinities, indicating woodland conditions. The bat assemblage indicates colder conditions for Unit II, with a higher proportion of steppe environment. We can thus see some discrepancies with respect to the results obtained from rodents, which indicate more steppic-arid conditions in Unit V and more woody-temperate conditions in Unit II.
Archaeobotanical data (Table 9) obtained from charcoal studies (Allué 2016) of Units V and II show that wood could have been carried into the cave by humans. However, no hearth or other intentionally constructed feature has been identified in the site at the rear of the cave where these fossil assemblages were deposited during the Pleistocene. Wood could have also been carried in by animals or brought in by natural surface fires (Andrews et al. 2016). The main taxa identified from the charcoal remains are Prunus, Acer and Quercus, a combination of large woodland trees and small trees and shrubs. The results of the phytolith analysis of coprolites (Scott et al. 2016) for Unit II can also be included, indicating a temperate steppe mosaic with grassy conditions, but the density of woody components cannot be determined. This interpretation is consistent with our rodent analysis, which shows a wide spectrum of species suggesting an open or semi-open landscape formed mainly by woody trees and shrubs.
In summary, the large mammals and the charcoal data reflect woodland environments, whereas the small vertebrates and coprolites mainly depict arid environments, mostly steppes and shrubland. These differences were at least partly explained by Andrews et al. (2016), who argued that they indicate different taphonomic trajectories, or origins of the accumulations. Both the large mammals and wood could have been selected in specific habitats and introduced to the site by humans (Allué 2016). By contrast, the small vertebrates and coprolites were accumulated by non-human predators, which could have hunted over a wider diversity of habitats, especially open areas (Andrews et al. 2016).
Azokh 1 Cave and the Southern Caucasus context
Several other Pleistocene sites have provided small-mammal studies in the Middle East: Qesem Cave (Smith et al. 2015; Maul et al. 2016), Hummal (Maul et al. 2015), Aghitu-3 Cave (Kandel et al. 2017; Nishiaki and Akazawa 2018; Frahm 2019), Nesher Ramla (Weissbrod and Zaidner 2014), Amud Cave (Belmaker and Hovers 2011), Misliya Cave (Weissbrod and Weinstein-Evron 2020), Dzudzuana Cave (Belmaker et al. 2016), Kaldar Cave (Rey-Rodríguez et al. 2020), Shanidar Cave (Tilby et al. 2020), Kudaro Caves (Baryshnikov 2002) and Karain Cave (Demirel et al. 2011).
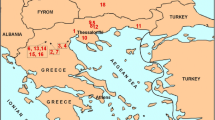
Most of these studies have highlighted the problems of identifying rodent species in this region, especially the Middle Pleistocene species. We focus on Qesem Cave and Misliya Cave, both of them located in Israel (Table 10, Fig. 6). We are aware that Israel is quite a long way from our study area for a comparative analysis; the sites do not belong to the same climatic zones (Fig. 6), and the faunas may be very different. However, these sites are present well-studied rodent assemblages that could be chronologically compared with Azokh 1 Cave.
Qesem Cave is an archaeological site located near Tel Aviv (32° 06′ 36″ N, 34° 58′ 48″ E) at 90 m a.s.l. It can be compared with Azokh Unit V (Table 9) in terms of its chronological framework. The palaeoecology of Qesem Cave was explored using two methods. The first was the nearest living relative (NRL) approach, and the second was the coexistence approach, both of which are based on the assumption that at least since the Neogene taxa have had climatic requirements similar to their NRLs, allowing inferences to be drawn on past climatic conditions. The modern climatic requirements were taken from the IUCN (Maul et al. 2015). The results show a climate of Mediterranean type during the accumulation of the two microvertebrate concentrations. A large proportion of the annual precipitation occurred in winter, whereas summers were dry. The temperatures were lower than today during the deposition of Concentration 2, with especially harsh winters, and winter precipitation was lower, resulting in lower precipitation seasonality. The landscape was a mosaic of open habitats with sparse vegetation, shrubland, Mediterranean forest, rocky areas and riverbanks (Maul et al. 2015; Smith et al. 2015). Both concentrations were accumulated by the barn owl Tyto alba (Smith et al. 2015).
Misliya Cave is located on the western slope of Mount Carmel (32° 44′ 29″ N, 34° 58′ 21″ E), facing the coastal plain and situated at an elevation of about 90 m a.s.l. It is chronologically comparable with Azokh Unit II (Table 10). The environmental conditions of Misliya Cave are characterized by a heterogeneous landscape, or habitat mosaics (Weissbrod and Weinstein-Evron 2020).
Misliya Cave is characterized by a combination of African rodent taxa, such as Mastomys and Arvicanthis, and Euro-Siberian taxa, such as Ellobius. This could suggest that the Transcaucasia region (like the Central Mountains in Israel and the Caucasus mountains), the Zagros and Taurus Mountains may have acted more as a barrier than as a migration corridor for the Azokh rodents. This hypothesis, combined with the greater Mediterranean influence on the Israeli sites, could explain the differences in the palaeoclimatic signal observed between Qesem and Misliya Cave on the one hand, and Azokh 1 Cave on the other.
Figure 6 shows the location of Qesem Cave and Misliya Cave in relation to the different main climates in present-day Israel according to Köppen (1936, modified by Beck et al. 2018). The climate ranges mainly from temperate (Misliya) to an arid limit (Qesem), with a hot summer and steppe environment. For Azokh 1 the climate is classified as “snowy”, with dry winters and warm summers.
The rodent assemblages of Azokh 1 show a strong Asiatic influence, whereas the large mammals (Van der Made 2016) are dominated by Western Eurasian species, also with African, Indian and central Asian elements. The rodent fauna did not record any African species, whereas some were found at the Israeli sites.
The identified rodent species present several biogeographic affinities but are mainly from Europe and Asia. Misliya also recorded the presence of African rodent taxa, such as Mastomys and Arvicanthis (Maul et al. 2016; Weissbrod and Weinstein-Evron 2020). Such species were not found at Azokh, indicating that the Caucasus, Zagros and Taurus Mountains probably acted as a barrier for small mammals coming from Africa. Moreover, most of the species found at Azokh display an Asiatic origin, suggesting the possible importance of the Black and Caspian seas as barriers to small mammals coming from Europe, as was previously indicated by Yanina (2014).
Conclusions
-
We have identified 434 rodent remains, corresponding to a minimum number of 237 individuals. The rodent assemblages from Azokh 1 Cave are composed of at least 13 taxa: seven arvicoline (Clethrionomys glareolus, Microtus gr. arvalis-socialis, Microtus (Terricola) spp., Arvicola ex. gr. persicus, Chionomys nivalis, Chionomys gud and Ellobius cf. lutescens), two cricetine (Cricetulus migratorius and Mesocricetus brandti), two gerbilline (Meriones gr. persicus-tristrami, Meriones gr. dahli-libycus), one dipodid (Allactaga cf. williamsi) and one murine species (Apodemus spp.).
-
The palaeoclimatic parameters obtained with the bioclimatic model suggest mean annual temperatures and precipitation similar to today, although the climate seems to be relatively warm-temperate in both units.
-
The palaeoenvironmental reconstruction, based on the habitat weighting method, shows an environment mainly composed of shrubland and steppe, with patches of deciduous forests and desert, similar to that currently found in the area.
-
Whereas large-mammal and charcoal studies indicate a woodland environment, small vertebrates and phytoliths from coprolites mainly reflect arid environments, such as steppes and desert. These differences can be partly explained by the origin of the accumulations.
-
A comparison between Azokh Cave on one hand and Qesem Cave and Misliya Cave (Israel) on the other indicates that there is no exact correspondence between the rodent faunas, both because the sites belong to different climatic regions and because the Israeli sites record African influences, which were not observed in Azokh 1.
Change history
24 August 2022
Missing Open Access funding information has been added in the Funding Note.
References
Abbate E, Sagri M (2012) Early to Middle Pleistocene Homo dispersals from Africa to Eurasia: geological, climatic and environmental constraints. Quatern Int 267:3–19. https://doi.org/10.1016/j.quaint.2011.02.043
Adams DC, Collyer ML, Kaliontzopoulou A (2020) Geomorph: software for geometric morphometric analyses. R Package Version 3(2):1
Agustí J, Lordkipanidze D (2019) An alternative scenario for the first human dispersal out of Africa. L’anthropologie 123:682–687. https://doi.org/10.1016/j.anthro.2019.102727
Allué E (2016) Charcoal remains from Azokh 1 Cave: preliminary results. In: Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Springer, Dordrecht, pp 297–304
Amori G, Hutterer R, Kryštufek B, et al (2016) Apodemus flavicollis The IUCN Red List of Threatened Species 2016: eT1892A115058023 8235:. https://doi.org/10.2305/IUCN.UK.2016-3.RLTS.T1892A22423256.en
Andrews P (1990) Owls, caves and fossils. University of Chicago, Chicago
Andrews P (2006) Taphonomic effects of faunal impoverishment and faunal mixing. Palaeogeogr Palaeoclimatol Palaeoecol 241:572–589. https://doi.org/10.1016/j.palaeo.2006.04.012
Andrews P, Hixson S, King T (2016) Paleoecology of Azokh 1. In: Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Springer, Dordrecht, pp 305–320
Asryan L (2015) Azokh Cave lithic assemblages and their contextualization in the Middle and Upper Pleistocene of Southwest Asia (PhD thesis).
Asryan L, Ollé A, Moloney N et al (2017) Chemical alteration of lithic artefacts: an experimental case study on the effect of guano on stone flakes and its contextualization in the archaeological assemblage of Azokh Cave (Southern Caucasus). Archaeometry 59:981–999. https://doi.org/10.1111/arcm.12300
Asryan L, Ollé A, Moloney N, King T (2020) Occupying cave-sites: a case study from Azokh 1 Cave (Southern Caucasus). In: Short-Term ocupations in Paleolithic Archaeology. pp 149–181
Baryshnikov GF (2002) Local biochronology of the Middle and Late Pleistocene mammals from Caucasus. Russ J Theriol: 61–67
Beck HE, Zimmermann NE, Mcvicar TR et al (2018) Data Descriptor : Present and future Köppen-Geiger climate classification maps at 1 -km resolution. Nature Publ Group 1–12. https://doi.org/10.1038/sdata.2018.214
Belmaker M, Bar-Yosef O, Belfer-Cohen A et al (2016) The environment in the Caucasus in the Upper Paleolithic (Late Pleistocene): evidence from the small mammals from Dzudzuana cave, Georgia. Quatern Int 425:4–15. https://doi.org/10.1016/j.quaint.2016.06.022
Belmaker M, Hovers E (2011) Ecological change and the extinction of the Levantine Neanderthals: implications from a diachronic study of micromammals from Amud Cave, Israel. Quatern Sci Rev 30:3196–3209. https://doi.org/10.1016/j.quascirev.2011.08.001
Bergl; R.A.; Dunn; A.; Fowler; A.; Imong; I.; Ndeloh; D.; Nicholas; A. & Oates; J.F. (2017) Myodes glareolus. The IUCN Red List of Threatened Species 2016: e.T4973A115070929. 8235:8
Bermúdez de Castro JM, Martinón-Torres M (2013) A new model for the evolution of the human Pleistocene populations of Europe. Quatern Int 295:102–112. https://doi.org/10.1016/j.quaint.2012.02.036
Blain H-A (2016) Amphibians and squamate reptiles from Azokh 1. In: Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Springer, Dordrecht, pp 191–210
Blain H-A, Bailon S, Cuenca-Bescós G (2008) The Early-Middle Pleistocene palaeoenvironmental change based on the squamate reptile and amphibian proxies at the Gran Dolina site, Atapuerca, Spain. Palaeogeogr Palaeoclimatol Palaeoecol 261:177–192. https://doi.org/10.1016/j.palaeo.2008.01.015
Bogicevic K, Nenadic D, Mihailovic D et al (2011) Late Pleistocene rodents (mammalia: Rodentia) from the Baranica Cave near Knjazevac (Eastern Serbia): systematics and paleoecology. Riv Ital Paleontol Stratigr 117:2
Chevret P, Renaud S, Helvaci Z et al (2020) Genetic structure, ecological versatility, and skull shape differentiation in Arvicola water voles (Rodentia, Cricetidae). J Zool Syst Evol Res 58:1323–1334. https://doi.org/10.1111/jzs.12384
Coil R, Tappen M, Ferring R et al (2020) Spatial patterning of the archaeological and paleontological assemblage at Dmanisi, Georgia: an analysis of site formation and carnivore-hominin interaction in Block 2. J Hum Evol 143:102773. https://doi.org/10.1016/j.jhevol.2020.102773
Coşkun Y (1999) Morphological characteristics of Meriones tristrami Thomas, 1892 (Rodentia: Gerbillinae) from Diyarbakır, Turkey. Turk J Zool 23:345–355
Coşkun Y (2016) Review of unique odd chromosome-numbered underground rodent species of the Palearctic region: Ellobius Lutescens Thomas 1897 (Rodentia: Cricetidae). Turk J Zool 40:831–841. https://doi.org/10.3906/zoo-1509-53
Darvish J, Mohammadi Z, Mahmoudi A, Siahsarvie R (2014) Faunistic and taxonomic study of rodents from northwestern Iran. Iran J Anim Biosyst (IJAB) 10:119–136
Darvish J (2011) Morphological comparison of fourteen species of the genus Meriones Illiger, 1811 (Rodentia: Gerbillinae) from Asia and North Africa. Iran J Anim Biosyst 7:49–74
Darvish J, Mohammadi Z, Ghorbani F et al (2015) Phylogenetic relationships of Apodemus Kaup, 1829 (Rodentia: Muridae) species in the Eastern Mediterranean inferred from mitochondrial DNA, with emphasis on Iranian species. J Mamm Evol 22:583–595. https://doi.org/10.1007/s10914-015-9294-9
Demirel A, Andrews P, Yalçinkaya I, Ersoy A (2011) The taphonomy and palaeoenvironmental implications of the small mammals from Karain Cave, Turkey. J Archaeol Sci 38:3048–3059. https://doi.org/10.1016/j.jas.2011.07.003
Denys C (2017) Subfamily Deomyinae, Gerbillinae, Leimacomyinae, lophiomyinae species accounts. In: Wilson DE, Lacher TE, Mittermeier RA (eds) Handbook of the Mammals of the World. Lynxs Edic, Barcelona, pp 598–650
Dianat M, Aliabadian M, Darvish J, Akbarirad S (2013) Molecular phylogeny of the Iranian Plateau five-toed jerboa, Allactaga (Dipodidea: Rodentia), inferred from mtDNA. mammalia 77:. https://doi.org/10.1515/mammalia-2012-0011
Dianat M, Darvish J, Cornette R et al (2017) Evolutionary history of the Persian Jird, Meriones persicus, based on genetics, species distribution modelling and morphometric data. J Zool Syst Evol Res 55:29–45. https://doi.org/10.1111/jzs.12145
Eken G, Bozdogan M, Molur S (2016) Allactaga williamsi (errata version published in 2017). The IUCN Red List of Threatened Species 2016: e.T136326A115205783. 8235:
Evans EMN, Van Couvering JAH, Andrews P (1981) Paleoecology of Miocene sites in Western Kenya. J Hum Evol 10:99–116
Fernández-Jalvo Y, King T, Andrews P et al (2010) The Azokh Cave complex: Middle Pleistocene to Holocene human occupation in the Caucasus. J Hum Evol 58:103–109. https://doi.org/10.1016/j.jhevol.2009.07.005
Fernandez-Jalvo Y, Andrews P (2016) Atlas of taphonomic identifications. Springer
Fernández-Jalvo Y, Andrews P, Denys C et al (2016a) Taphonomy for taxonomists: Implications of predation in small mammal studies. Quatern Sci Rev 139:138–157. https://doi.org/10.1016/j.quascirev.2016.03.016
Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (2016) Azokh Cave and the Transcaucasian Corridor. Springer, Dordrecht
Fernandez-Jalvo Y, King T, Yepiskoposyan L, Andrews P (2016c) Introduction: Azokh Cave and the Transcaucasican Corridor. In: Fernandez-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Springer, Dordrecht, pp 1–26
Frahm E (2019) Upper Palaeolithic settlement and mobility in the Armenian highlands : agent-based modeling, obsidian sourcing, and lithic analysis at Aghitu-3 Cave. J Paleolithic Archaeol 2:418–465
Granjon L (2016) Meriones libycus (errata version published in 2017). The IUCN Red List of Threatened Species 2016: e.T13164A115110005. 8235:
Guseinov M (2010) Early Paleolithic of Azerbaijan. TekNur, Baku, 234pp (in Russian with and English summary).
Habibi K (2004) Mammals of Afghanistan. Zoo Outreach Organisation/USFWS, Coimbatore, India
Heinrich W-D (1987) Neue Ergebnisse zur Evolution und Biostratigraphie von Arvicola (Rodentia, Mammalia) im Quatär Europas. Z Geol Wiss 15:389–406
Hernández Fernández M (2001) Bioclimatic discriminant capacity of terrestrial mammal faunas. Glob Ecol Biogeogr 10:189–204. https://doi.org/10.1046/j.1466-822x.2001.00218.x
Hernández Fernández M, Álvarez Sierra MÁ, Peláez-Campomanes P (2007) Bioclimatic analysis of rodent palaeofaunas reveals severe climatic changes in Southwestern Europe during the Plio-Pleistocene. Palaeogeogr Palaeoclimatol Palaeoecol 251:500–526. https://doi.org/10.1016/j.palaeo.2007.04.015
Hershkovitz I, May H, Sarig R et al (2021) A Middle Pleistocene Homo from Nesher Ramla, Israel. Science 372:1424–1428
Hershkovitz I, Weber GW, Quam R et al (2018) The earliest modern humans outside Africa. Science 359:456–459. https://doi.org/10.1126/science.aap8369
Jangjoo M, Darvish J, Vign JD (2011) Aplication of outline analysis on fossil and modern specimens of Apodemus. Iran J Anim Biosyst 7(2):143–155
Kandel AW, Gasparyan B, Allué E et al (2017) The earliest evidence for Upper Paleolithic occupation in the Armenian Highlands at Aghitu-3 Cave. J Hum Evol 110:37–68. https://doi.org/10.1016/j.jhevol.2017.05.010
Karami M, Hutterer R, Benda P et al (2008) Annotated check-list of the mammals of Iran. Lynx (praha) 39:63–102
Kasimova RM (2001) Anthropological research of Azykh man osseous remains. Hum Evol 16:37–44. https://doi.org/10.1007/BF02438921
Kaya A, Gharakhloo MM, Coşkun Y (2018) Geographic variation in the skull morphology of Ellobius lutescens Thomas, 1897 (Mammalia: Rodentia) by geometric morphometric analyses. Vertebrate Zool 68:157–164
Kefelioglu H, Yigit N, Kryštufek B (2008) Meriones dahli. The IUCN Red List of Threatened Species 2008: e.T13162A3415328. 8235:
King T, Compton T, Rosas A (2016) Azokh Cave Hominin remains. In: Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Springer, Dordrecht, pp 103–116
Knitlová M, Horáček I (2017) Late Pleistocene-Holocene paleobiogeography of the genus Apodemus in Central Europe. PLoS ONE 12:1–23. https://doi.org/10.1371/journal.pone.0173668
Kryštufek B., Vohralík V. (2005) Mammals of Turkey and Cyprus. Rodentia I: Sciuridae, Dipodidae, Gliridae, Arvicolinae.
Kryštufek B, Vohralík V (2009) Mammals of Turkey and Cyprus, Rodentia II: Cricetinae, Muridae, Spalacidae, Calomyscidae, Capromyidae, Hystricidae, Castoridae
Krystufek B (2017) Chionomys nivalis , European Snow Vole. The IUCN Red List of Threatened Species 2016: eT4659A115069366 8235:. https://doi.org/10.2305/IUCN.UK.2016-3.RLTS.T4659A22379147.en
Kryštufek B (1999) Snow voles, genus Chionomys, of Turkey. Mammalia 63(3):323–339
Kryštufek B, Bukhnikashvili A, Sozen M, Isfendiyaroglu S (2017) Cricetulus migratorius. The IUCN Red List of Threatened Species 2016: eT5528A115073390 8235:. https://doi.org/10.2305/IUCN.UK.2016-3.RLTS.T5528A22391440.en
Kryštufek B, Shenbrot G (2016) Ellobius lutescens, Transcaucasian mole vole. The IUCN Red List of Threatened Species 2016: eT7655A22340006 8235:. https://doi.org/10.2305/IUCN.UK.2016- 2.RLTS.T7655A22340006.en
Kryštufek B, Tesakov AS, Lebedev VS et al (2020) Back to the future: the proper name for red-backed voles is Clethrionomys Tilesius and not Myodes Pallas. Mammalia 84:214–217. https://doi.org/10.1515/mammalia-2019-0067
Kryštufek B, Yigit N, Amori G (2015) Mesocricetus brandti , Brandt ’ s Hamster. The IUCN Red List of Threatened Species 2008: eT13220A3421550 8235:. https://doi.org/10.2305/IUCN.UK.2008.RLTS.T13220A3421550.en
López-García JM (2008) Evolución de la diversidad taxonómica de los micromamíferos en la Península Ibérica y cambios Paleoambientales durante el Pleistoceno Superior (PhD thesis).
López-García JM, Blain H-A, Cuenca-Bescós G et al (2011) Small vertebrates (Amphibia, Squamata, Mammalia) from the late Pleistocene-Holocene of the Valdavara-1 cave (Galicia, northwestern Spain). Geobios 44:253–269. https://doi.org/10.1016/j.geobios.2010.10.001
López-García JM, Blain HA, Cuenca-Bescós G et al (2010) Palaeoenvironmental and palaeoclimatic reconstruction of the Latest Pleistocene of El Portalón Site, Sierra de Atapuerca, northwestern Spain. Palaeogeogr Palaeoclimatol Palaeoecol 292:453–464. https://doi.org/10.1016/j.palaeo.2010.04.006
Lordkipanidze D, De Ponce León MS, Margvelashvili A et al (2013) A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science 342:326–331. https://doi.org/10.1126/science.1238484
Lozano-Fernández I, Agustí J, Bescós GC, et al (2013) Pleistocene evolutionary trends in dental morphology of Mimomys savini ( Rodentia , Mammalia ) from Iberian peninsula and discussion about the origin of the genus Arvicola. 24:. https://doi.org/10.4000/quaternaire.6587
Mahmoudi LC, Maul M, Khoshyar et al (2020) Evolutionary history of water voles revisited: confronting a new phylogenetic model from molecular data with the fossil record. Mammalia 1–14 https://doi.org/10.1515/mammalia-2018-0178
Manuk V (2010) Atlas of the Ngorno-Karabahg republic. Stepanakert
Marin-Monfort MD, Caceres I, Andrews P (2016) Taphonomy and site formation of Azokh 1. In: Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Springer, Dordrecht, pp 211–249
Markova AK (1982) Small mammals fauna from Paleolithic Cave site Azykh. Paleontologucheskii Sbornik 19:14–28
Maul LC, Bruch AA, Smith KT et al (2016) Palaeoecological and biostratigraphical implications of the microvertebrates of Qesem Cave in Israel. Quatern Int 398:219–232. https://doi.org/10.1016/j.quaint.2015.04.032
Maul LC, Rabinovich R, Biton R, Maul LC (2020) At the southern fringe : extant and fossil water voles of the genus Arvicola ( Rodentia, Cricetidae, Arvicolinae ) from Israel, with the description of a new species. Hist Biol 00:1–21. https://doi.org/10.1080/08912963.2020.1827240
Maul LC, Smith KT, Shenbrot G et al (2015) Microvertebrates from unit G/layer 17 of the archaeological site of Hummal (El Kowm, Central Syria): preliminary results. Anthropologie (france) 119:676–686. https://doi.org/10.1016/j.anthro.2015.10.010
Mitteroecker P, Windhager S, Müller GB, Schaefer K (2015) The morphometrics of “masculinity” in human faces. PLoS ONE 10(2):e0118374. https://doi.org/10.1371/journal.pone.0118374
Molur, S. & Sozen M (2016) Meriones persicus, Persian Jird. The IUCN Red List of Threatened Species 2016: eT13166A22433231 8235:. https://doi.org/10.2305/IUCN.UK.2016-2.RLTS.T13166A22433231.en
Moradi Gharkheloo M (2003) A study on the morphology, karyology and distribution of Ellobius Fisher, 1814 ( Mammalia : Rodentia ) in Iran. Turk J Zool 27:281–292
Murray J, Domínguez-alonso P, Fernández- Y et al (2010) Pleistocene to Holocene stratigraphy of Azokh 1 Cave, Lesser Caucasus. Irish J Earth Sci 28:75–91. https://doi.org/10.3318/IJES.2010.28.75
Murray J, Lynch EP, Dominguez-Alonso P, Barham M (2016) Stratigraphy and sedimentology of Azokh Caves, South Caucasus. In: Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Springer, Dordrecht, pp 27–54
Nadachowski a, (1991) Systematics, geographic variation, and evolution of snow voles (Chionomys) based on dental characters. Acta Theriol 36:1–45
Naderi G, Hemami MR, Mohammadi S (2011) Investigation of habitat preferences of Iranian jerboa (Allactaga firouzi Womochel 1978). Mammalia 75. https://doi.org/10.1515/mamm.2011.008
Neumann K, Yiğit N, Fritzsche P et al (2017) Genetic structure of the Turkish hamster (Mesocricetus brandti). Mamm Biol 86:84–91. https://doi.org/10.1016/j.mambio.2017.06.004
Nishiaki Y, Akazawa T (2018) The Middle and Upper Paleolithic Archeology of the Levant and Beyond. Replacement of Neanderthals by Modern Humans Series 35–47. https://doi.org/10.1007/978-981-10-6826-3_3
Parfitt SA (2016) Rodents, lagomorphs and insectivores from Azokh Cave. In: Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Springer, Dordrecht, pp 163–176
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing
Rey-Rodríguez I, Arnaud J, López-García JM, et al (2021) Distinguishing between three modern Ellobius species (Rodentia, Mammalia) and identification of fossil Ellobius from Kaldar Cave (Iran) using geometric morphometric analyses of the first lower molar. Palaeontologia Electronica 24 (1):a01:1–18. https://doi.org/10.26879/1122
Rey-Rodríguez I, López-García J-M, Blain H-A et al (2020) Exploring the landscape and climatic conditions of Neanderthals and anatomically modern humans in the Middle East: the rodent assemblage from the late Pleistocene of Kaldar Cave (Khorramabad Valley, Iran). Quatern Sci Rev 236:106278. https://doi.org/10.1016/j.quascirev.2020.106278
Rohlf FJ (2016) tpsDig, digitize landmarks and outlines, Version 2.32. Department of Ecology and Evolution, State University of New York at Stony Brook, New York
Royer A, García BA, Laffont R, et al (2020) New bioclimatic models for the quaternary palaearctic based on insectivore and rodent communities. 560:
Rusin M (2017) Ellobius talpinus , Northern mole vole. The IUCN Red List of Threatened Species 2016: eT7656A115085720 8235:. https://doi.org/10.2305/IUCN.UK.2016-3.RLTS.T7656A22339917.en
Sandor AD (2018) Rediscovered after half a century: a new record of the grey dwarf hamster, Cricetulus migratorius (Mammalia: Cricetidae), in Romania. Turk J Zool 42:495–498. https://doi.org/10.3906/zoo-1712-15
Schlager S (2017) Morpho and Rvcg Shape Analysis. In: Guoyan Zheng and Shuo Li and Gabor Szekely (ed) Statistical Shape and Deformation Analysis. pp 217–256
Scott L, Rossouw L, Cordova C, Risberg J (2016) Palaeoenvironmental Context of Coprolites and Plant Microfossils from Unit II. Azokh 1. In: Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Vertebrate Paleobiology and Paleoanthropology. Springer, Cham. https://doi.org/10.1007/978-3-319-24924-7_13
Sevilla P (2016) Bats from Azokh Cave. In: Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Springer, Dordrecht, pp 177–189
Shenbrot G (2009) On the conspecifity of Allactaga hotsoni Thomas, 1920 and Allactaga firouzi Womochel, 1978 (Rodentia: Dipodoidea). Mammalia 73:231–237. https://doi.org/10.1515/MAMM.2009.043
Shenbrot G, Kryštufek B, Molur S (2016) Ellobius fuscocapillus , Southern mole vole. The IUCN Red List of Threatened Species 2016: eT7654A22339730 8235:. https://doi.org/10.2305/IUCN.UK.2016- 2.RLTS.T7654A22339730.en
Siahsarvie R, Darvish J (2008) Geometric morphometric analysis of Iranian wood mice of the genus Apodemus (Rodentia, Muridae). Mammalia 72. https://doi.org/10.1515/MAMM.2008.020
Smith KT, Christian L, Flemming F et al (2015) The microvertebrates of Qesem Cave : A comparison of the two concentrations. Quat Int 398:233–245. https://doi.org/10.1016/j.quaint.2015.04.047
Sozen M, Bukhnikashvili A, Shenbrot G, et al (2016) Meriones tristrami (errata version published in 2017). The IUCN Red List of Threatened Species 2016: e.T13170A115110701. 8235:
Sözen M, Özkurt SO, Karataş A et al (2009) On the karyology, morphology and biology of Chionomys gud (Satunin, 1909) (Mammalia: Rodentia) in Turkey. North Western J Zool 5(1):121–129
Stoetzel E, Cornette R, Lalis A et al (2017) Systematics and evolution of the Meriones shawii/grandis complex (Rodentia, Gerbillinae) during the Late Quaternary in northwestern Africa: exploring the role of environmental and anthropogenic changes. Quatern Sci Rev 164:199–216. https://doi.org/10.1016/j.quascirev.2017.04.002
Tesakov AS (2016) Early Middle Pleistocene Ellobius ( Rodentia , Cricetidae , Arvicolinae ) from Armenia. 15:151–158
Tilby E, Miracle P, Barker G (2022) The microvertebrates of Shanidar Cave: preliminary Taphonomic Findings. Quaternary 5:4. https://doi.org/10.3390/quat5010004
Van der Made J, Torres T, Ortiz JE (2016) The new material of large mammals from Azokh and comments on the older collections. In: Fernández-Jalvo Y, King T, Yepiskoposyan L, Andrews P (eds) Azokh Cave and the Transcaucasian Corridor. Springer, Dodrecht, pp 117–162
Weissbrod L, Weinstein-Evron M (2020) Climate variability in early expansions of Homo sapiens in light of the new record of micromammals in Misliya Cave. Israel J Human Evol 139:102741. https://doi.org/10.1016/j.jhevol.2020.102741
Weissbrod L, Zaidner Y (2014) Taphonomy and paleoecological implications of fossorial microvertebrates at the Middle Paleolithic open-air site of Nesher Ramla, Israel. Quatern Int 331:115–127. https://doi.org/10.1016/j.quaint.2013.05.044
Yanina TA (2014) The Ponto-Caspian region: Environmental consequences of climate change during the Late Pleistocene. Quatern Int 345:88–99. https://doi.org/10.1016/j.quaint.2014.01.045
Acknowledgements
We would like to thank Roberto Portela Miguez, Senior Curator in Charge of Mammals, for his help with the reference collection in the Natural History Museum of London; Lawrence Heaney, Adam Ferguson and Lauren Smith of the Chicago Field Museum; and Marisa Surovy, Judith Galkin and Carl Mehling of the American Museum of Natural History of New York. We would like to thank Rupert Glasgow for reviewing the English language of the manuscript. We also want to thank the Guest Editor Angel Blanco, as well as Dr. Alexey S. Tesakov and the anonymous reviewer for their comments and suggestions, which greatly improved the final version of the manuscript.
Funding
Open access funding provided by Universitat Rovira i Virgili. I. Rey-Rodriguez is the beneficiary of a Margarita Salas postdoctoral scholarship (2021URV‐MS‐03) at Universitat Rovira i Virgili funded by the European Union – NextGenerationEU. J.M. López-García was supported by a Ramón y Cajal contract (RYC-2016–19386) with financial sponsorship from the Spanish Ministry of Science, Innovation and Universities.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Microvertebrate Studies in Archaeological Contexts: Middle Paleolithic to early Holocene past environments
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visithttp://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rey-Rodríguez, I., López-García, J.M., Stoetzel, E. et al. Palaeoecological reconstructions of the Middle to Late Pleistocene occupations in the Southern Caucasus using rodent assemblages. Archaeol Anthropol Sci 14, 96 (2022). https://doi.org/10.1007/s12520-022-01555-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-022-01555-w