
Abstract
T cells in tumors—the so-called tumor infiltrating lymphocytes (TIL) have been studied intensively over the past years. Compelling evidence point to a clinical relevance for high numbers of T cells at the tumor site with CD8 memory T cells as a key denominator for overall survival (OS) in patients with colo-rectal cancer (CRC), and also for others solid cancers. These data goes hand in hand with studies of clonality of TIL showing the T cells among TIL are expanded clonally, and also that tumor specific T cells of CD4 as well as CD8 type are enriched at the tumor site. The tumor microenvironment is hostile to T cell function e.g., due to expression of enzymes that depletes the amino acids tryptophan and arginine, high concentration of tumor secreted lactate, and presence innate cells or regulatory T cells both with suppressive activity. Analyses of the specificity of TILs in melanoma demonstrate that quite few known antigens are in fact recognized by these cultures underscoring patient unique and/or mutated antigens may represent important target for recognition.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
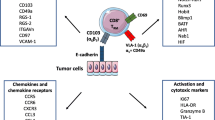
Tremendous focus has been on elucidating the biological properties of cancer cells. To this end, for many cancers we have detailed information on the genetic and epi-genetic alterations in the cancerous cells and the associated changes in signaling pathways, cell cycle regulation, etc. However, it has also become realized that the tumor is a complex entity comprising cancer cells and stromal cells, tumor infiltrating cells—both cells of the immune system and cells not by convention being of the immune system, as well as an extracellular matrix mainly of proteins and carbohydrates. Strikingly, cancer cells may comprise as little as approx. 30 % of the cells in the tumor. Moreover, cancer cells are in most cases not autonomous—they need stroma cells to survive and grow, which in turn implies that non-cancerous cells in the tumor have an intimate relationship to the cancer cells. The intra-tumor cell types may play various roles in the natural life of the tumor, being pro-tumorigenic or the opposite. In this part we discuss the role of CD4 and CD8 effector T cells in the tumor microenviroment.
Tumor Infiltrating Lymphocytes in Solid Tumors and Impact on Course of Disease
Mainly using immune histochemistry (IHC), the presence of T cells in tumor biopsies and their potential impact on prognosis have been studied for decades. Early data suggested that a brisk infiltration of T cells in primary melanoma lesions was a positive prognostic factor [20]. More recently similar data has been found in other cancers including ovarian cancer [92], renal cell carcinoma (RCC) [73], bladder cancer [96], and several other solid cancers. Obviously, this goes well hand in hand with the data now available on the presence of tumor specific T cells among TIL as discussed in more detail in later sections. As given above the main incentive for studying T cell infiltration has been to identify any impact on clinical course. However, when it comes to a potential clinical significance many studies are rather small and thus of limited statistical strength. In a recent meta-analysis including studies in which clinical significance of TIL were studied in solid tumors (CD3, CD4, CD8, FoxP3 and rations between these), the presence of CD3, CD8, as well as a high CD8/FoxP3 ration had a positive effect on survival [44]. In addition, the assessment of TIL density and distribution was shown to independently predict sentinel lymph node status and survival in patients with melanoma [10]. Some studies have been more detailed in term of T cell subtypes and the compiled data described by Fridman and colleagues show that CD8/CD45RO and Th1 T cells are both found to be a positive prognostic factor in the vast majority of studies [36]. Thus, memory T cells and/or T cells with a Th1 biased phenotype are associated with a better prognosis. To this end, Galon and colleagues have studied approx. 400 patient samples and shown that CD8, CD45RO T cells in the core of the tumor is a superior prognostic factor to the conventional clinical staging in colorectal cancer [39]. Strikingly, even patients with localized disease had a very poor prognosis, similar to that of patients with concomitant distant metastasis if their tumors showed low frequency of T cells in the tumor core. Thus, overall survival of these patients is governed to a large extent by the local adaptive—and supposedly tumor specific - immune response. Obviously, this underscores that the quantity and quality of T cells participating is key, with CD45RO T cells being crucially important. Moreover, the capacity of the T cells to infiltrate deep into the core of the tumor is fundamental in terms of anti-tumor activity that translate into benefit in survival benefit [36].
Although there have now been conducted numerous studies that suggest a positive prognostic impact of TIL, some of which are quite large, this still need to be verified in large multi-center studies recently initiated [40]. Provided that the data from this study support the impact of immune infiltrating cells in CRC lesions, this will completely change the prognostic tools in this disease to the benefit of the patients. To this end, quite a substantial fraction of patients with local spread of disease are treated with chemotherapy. However, available data suggests that most of these patients have a very limited risk of relapse if the tumor has a pronounced infiltration of memory T cells in the tumor core. As mentioned, other cancers have been studied as well, and if the prognostic impact of T cell infiltration is verified in CRC there is no question that similar studies will be conducted in other indications.
At present there is very limited knowledge as to why some tumors are heavily infiltrated by T cells whereas others are not. Very elegant studies in murine models from the lab of Robert Schreiber have suggested the “Three Es of cancer immunoediting” - that the interplay between tumor and the immune system goes through three phases characterized by immune-elimination (of cancer cells), immune equilibrium (between cancer cells and cells of the immune system) and immune escape (by cancer cells). Along those lines, it could be speculated that some lesions progress faster and reach the “escape” phase even as a primary or early metastatic tumors [30]. However, the notion of the “Three Es of cancer immunoediting” and its relevance in human tumor immunology is still unclear. Moreover, such very rapidly progressing lesions would still have a biological background. To this end, one would expect that the induction of a clinically relevant T cell response - capable of extending the life-expectancy of the patients, is induced via cells of the innate immune system. Consequently, the quality and quantity of the induced response may be influenced by numerous factors in the micro-environment of the tumor and the draining lymph nodes; cytokines and chemokines in turn influencing the cell types attracted and their function, availability of nutrients, oxygen, lactate, expression of danger signals, etc. Quite likely, a rational approach to reach a higher level of understanding of these important issues will require a systems biology approach with large numbers of patient’s samples with associated clinical data to be studied at the genomic, proteomic, molecular, and cellular level and the collected data to be integrated. The data may provide us with tools that enable the therapeutic transition of a tumor from a “non-inflammatory” to an “inflammatory tumor” with the associated impact on overall survival. Moreover, although T cell infiltration into tumors may have a significant impact on patient survival, most cancer patients with metastatic disease eventually die from their cancer, which underscores that elucidation of the mechanisms of immune escape may lead to development of new strategies that extend the immunological control of tumor progression.
It remains to be seen if T cells—as suggested by the available data so far – are important players in the majority of solid cancers. To this end, in renal cell carcinoma (RCC) some data suggests that TILs is only a good prognostic factor if the cells are proliferating [73]. In pancreatic cancer, immune therapy with agonistic CD40 antibody was shown to be depended on macrophages rather than the expected T cells [12]. Thus, each organ is characterized by unique immune characteristics and similarly each tumor entity is likely to possess unique infiltrates of immune cells.
This proposed clinical impact of CD8 T memory cells among TIL is striking and obviously suggest their in situ function to be direct cancer cell killers. Accordingly, our current knowledge suggests that CD8 T cells seem to be more “simplified” in function compared to the CD4 T cells, and the reason for a stronger consensus in clinical significance lies in the fact that the presence of a simple killer at the tumor site really only have one outcome. However, it should be kept in mind that suppressor CD8 T cells have been described. These CD8 suppressors detected in quite high frequencies in cancer patients were characterized by very potent suppressive capacity, and the detection of such cells at the tumor site could therefore reveal yet another layer of complexity [6]. Unfortunately, there are as of yet no surface markers that enable IHC of these cells.
As given above data are accumulating that point to a role of CD8 T cells in spontaneous tumor control. Available data on CD4 T cells are more dubious, which could be taken as support for the notion that CD4 T cells are more plastic and play dual roles; thus CD4 T cells can convert from anti-tumor to pro-tumor [95, 111]. Moreover, concerning Treg some organs are characterized by large numbers of Treg, underscoring the role of the tissue in immune responses as recently suggested by Polly Matzinger [67]. Herein lies probably the reason that prognostic significance of CD4 T cells—irrespective of subtype—is controversial despite numerous and quite large studies [36]. Early data in studies of Treg mainly studied the expression of CD3 FoxP3 cells alone. However, more recent studies include staining for CD8 as well and the ratio of these cell types have been shown in numerous studies to be significant, with more CD8/few FoxP3 being described as a positive prognostic factor in many studies. Thus, despite the fact that FoxP3 may not be expressed solely by Treg these data does suggest a predominant expression of FoxP3 in cells that are pro-tumor.
CD4 T cells will in many cases not be able to recognize cancer cells directly due to the lack MHC class II on most (solid) cancers. Obviously, hematological cancer represents an exception since many cell types of the immune system may express class II molecules e.g., B and T cells. However, solid cancers may express class II molecules constitutively, such as over 40 % of melanomas [68] or upon induction with IFN-g. In addition, TILs in melanoma, breast and ovarian cancer comprise CD4 T cells specific for class II restricted tumor antigens [25, 26, 88]. CD4+ T cells may indeed exert anticancer actions by providing help to CD8+ T cells or, in some cases, directly recognizing endogenously processed antigens presented on the surface of cancers eventually followed by secretion of type 1 cytokines [27, 37] or direct tumor killing [86] To this end, a CD4 T cell clone specific for a class II restricted NY-ESO-1 peptide mediated anti-tumor function when administered to a melanoma patient [50] and some patients undergoing TIL based adoptive T cell therapy with infusion products largely dominated by CD4+ T cells nonetheless obtained clinical regressions either in our trial [clinicaltrials.gov identifier : NCT00937625] or elsewhere [15, 84]. Along this line of research, Friedman and colleagues have studied 44 cultures of CD8 depleted TILs and found that 8 of these (18 %) secreted IFN-γ that could be blocked by class II antibody upon co-culture with autologous established melanoma cell lines or fresh tumor digests. In another study, we have detected specific production of two type 1 cytokines (TNF-α and IFN-γ) by CD4+ T cells in 6 out of 12 TILs (50 %), upon co-culture with autologous short-term cultured melanoma cells in which class II expression was pre-induced by IFN-γ [27]. Thus, these data point out that a significant fraction of tumors are indeed naturally infiltrated by CD4 T cells which are tumor specific and class II restricted. However, their function is hard to scrutinize due the more plastic nature of these cells—and the lack of appropriate markers to identify different subsets.
Clonally Expanded T Cell Clonotypes Among TIL
T cells recognize peptide antigens in the context of MHC molecules through the clonally distributed T-cell receptor (TCR). Accordingly, activation of a T cells and the subsequent proliferation leads to elevated numbers of mRNA encoding that particular TCR alpha and beta chains. Thus, detection of expanded TCR clonotypes is a strong indication that an HLA restricted T-cell responses is ongoing [74, 106]. A limited number of studies have been conducted in various murine tumor models e.g. the widely used murine melanoma model B16. T cells infiltrating B16 tumors were shown to comprise numerous clonal TCR transcripts none of which could be recurrently found in different animals [70]. Even in different tumors in the same animal most TCR clonotypes are unique i.e. none-public at least concerning spontaneous responses [70]. In a study in which IL-2 was targeted to the tumor site identical TCR clonotypes could be found infiltrating tumors on both flanks of the animals, even if only one tumor had been targeted with IL-2 [102, 104]. Thus, at least during therapy T-cell clonotypes may travel across the blood stream and home to distant tumor sites.
In human tumors data from several studies demonstrate that TILs are clonally expanded. To this end, the presence of expanded T-cell clonotypes have been demonstrated in melanoma [101, 103], and these findings have been corroborated by others and – importantly – also been found in other cancer lesions, e.g., seminoma and breast cancer [107].
Melanoma has been most intensively studied, and at least as judged by the more limited data available from studies of other cancer biopsies, melanoma seem to be characterized by an exceedingly high number of clonally expanded T cells. Thus, subcutaneous lesions may comprise up to 60 or 80 unique expanded T cell clonotypes [103]. This supports the notion of an ongoing anti-tumor immune response in situ, however, standing alone these analyses are merely descriptive and reveals no insight into the type, function and specificity of the T cells.
The clonal distribution of the TCR offers the means to detect and track specific T cells based upon detection of the unique TCR. In this respect, identical TCR clonotypes can be detected in different lesions from the same patient and treatment induced responses can be tracked to the tumor site from the blood [3]. Also, in some studies it has been possible to track T-cell clonotypes from the tumor even before vaccination and monitor the clonotype upon treatment both in blood and tumor site [13]. Similarly, TCR clonotype analyses may shed light on the TIL in situ and after in vitro growth. To this end, we have shown that although clonotype composition may change during culture [105] the use of high-dose IL-2 for expansion of TILs for adoptive transfer seem to maintain a substantial fraction of in situ TIL clonotypes during expansion, and as a consequence studies of IL-2 expanded TILs bear relevance for the TILs in situ at least in terms of specificity [54].
The specificity of TIL will be discussed in some more detail below, but in a number of studies the connection between clonality and specificity has been verified providing evidence that the clonally expanded cells at the tumor site may indeed be tumor specific. Thus, clonal responses against the HLA-A2 restricted melanocyte differentiation antigen MART-1 has been studied in detail with respect to clonotype composition [91, 110], and Ferradini and colleges characterized a cytotoxic T-cell clone derived from the regressive part of a melanoma tumor, however the antigen was not characterized [34]. The general findings of such studies is that responses may involve numerous TCR clonotypes, and although there may be structural similarities, quite different TCRs may recognize the same HLA/peptide complex [65, 89].
Specificity and Function of TIL
We and others have also used IHC with specific HLA-peptide multimers to demonstrate the presence of tumor specific T cells at the tumor site, e.g., using multimers to detect CD8 T cells specific for Mart-1, survivin and HO-1 [4–6]. Indeed using confocal microscopy it was possible to demonstrate that specific cells also expressed markers associated with cytotoxic activity suggesting in situ tumor cell killing. Others have used a different approach and taken advantage of needle biopsies to demonstrate an enrichment of tumor specific CD8 T cells intratumorally compared to peripheral blood [61, 62, 77, 78]. These data demonstrate that numerous antigens are recognized by melanoma TIL and also that these antigens fall in different groups. Thus, although it seems that differentiation antigens gp100 [11] and Mart-1 [57] are quite dominant, cancer testis antigens [21], over expressed antigens [2], and mutated antigens are recognized as well [115].
CD8 T Cells in Melanoma—Lessons from in vitro Expanded TIL Cultures and Their Clinical Application
CD8 T cells among TILs have shown great promise when used for ACT in melanoma [90]. This therapy has been pioneered by the group of Steve Rosenberg at NIH and is currently established in a small number of cancer-centers worldwide [14, 32, 87]. TILs are cultured from resected melanoma lesions and after in-vitro expansion in the presence of IL-2 transferred back to the lymphodepleted patient together with supplement IL-2 treatment. As given above, in situ TCR clonotypes are maintained during the expansion and studies of these cultures may therefore provide valuable new knowledge with regards to the antigens recognized and the therapeutic potential of the cells.
The Antigen Specificities Recognized by Melanoma Tumor Infiltrating Lymphocytes
Melanoma TILs used for adoptive cell therapy has shown both autologous and allogenic tumor cell recognition, but until recently very little was known about the antigen specific reactivity of these TIL preparations. Two publications by Andersen et al. [7] and Kvistborg et al. [60] have recently demonstrated that TILs comprise T cells reactive against only a minor fraction of the previously described T-cell epitopes relevant for melanoma. Moreover, most identified peptide-specific T-cell populations were of quite low frequency (< 1 % of total CD8+ T cells). In these studies, melanoma TILs were screened for reactivity against a library of all published T cell epitopes of relevance for melanoma, including 175 MHC-class I peptides restricted to HLA-A1, A2, A3, A11 and B7 [7]. Screening of peptide-specific T-cell responses was conducted by MHC-multimers, generated by peptide exchange from conditional ligand-HLA complexes and combinatorially encoded with different fluorescence molecules to generate unique two-color codes allowing parallel detection of large numbers of different antigen specific T cells [46, 108]. Based on TILs from three different centers data from these studies showed that T-cell populations recognizing described T-cell epitopes are low-frequentcy and only a small fraction of the described melanoma-associated antigens are indeed recognized. The most prominently recognized groups of antigens were differentiation antigens, with MART-1 and gp100 together accounting for more than half of the responses. Cancer-testis antigen-responses were also observed, but strikingly few epitopes from the group of over expressed antigens were recognized, and the majority of these were encoded in alternative open reading frames (ORFs).
An obvious limitation in our knowledge about T-cell recognition relates to the HLA-restriction of the described epitopes. In the database generated of all described tumor associated T-cell epitopes 57 % of all epitopes (326 of 576) are restricted to HLA-A2. Although this allele is frequently expressed in many different population, even for an HLA-A2 positive individual the responses determined by the additional 5 HLA class I loci may be of equal importance for the tumor recognition as the HLA-A2 restricted recognition.
Recognition of de-novo Sequences/Mutated Antigen
There is no question a significant number of tumor associated antigens remains to be identified. An important group of antigens, not included in from the study described above is the personalized de-novo epitopes—antigens arising from mutations in the individual tumor. It has recently been demonstrated in mice that mutated antigens may be a target for immune recognition, with the potential to eradicate a developing tumor [66], and that the growing tumor undergo an immunoediting process, including changes in the mutation pattern, and thereby may escape immunological recognition [31]. Based on these data, it could be speculated that a large fraction of the tumor reactive CD8 T cells among TILs recognize specific mutations formed in the individual patient’s tumor.
Since the mutation landscape of human tumors is very diverse, a personalized approach may be required to describe the entire range of T-cell reactivity against mutated antigens. The vast majority of mutations found in human tumors is patient specific and will differ among different patients, even with the same cancer type. Furthermore, there may even be differences among lesions from the same patient, as well as intra-lesional heterogeneity [42]. It has been documented that mutated sequences may be recognized by the immune system and that these can be therapeutically targeted to induce efficient anti-tumor reactivity even in tumors developed in an immunocompetent host [18, 63]. Also among TILs used for adoptive cell therapy recognition of mutated sequences has been described [116]. I has been predicted that a large number (~8000) of HLA-binding peptides may exists around functionally relevant missense mutation sites with significantly increased HLA-binding affinity of the mutated peptide versus the cognate wildtype peptide. This would imply that a large number of peptide sequences not present in normal cells may be presented to the immune system as a consequence of mutations in tumors. Since these sequences are foreign to the patient’s immune system, they serve as ideal targets for T-cell recognition, as the repertoire of T cells recognizing these are not limited by tolerance, as is often the case with peptide sequences from shared antigens.
TILs in Non-melanoma Solid Cancers and Potential Clinical Application
In ovarian cancer, infiltration of CD3 and CD8 T cells is of prognostic significance as evident in a recent meta-analyses [51]. Further supporting the importance of a stimulatory intratumoral T-cell response, regulatory cell populations has been correlated with poor clinical outcome in ovarian cancer, such as Tregs [22] and immunosuppressive B7-H4 expressing macrophages [59]. Also, and PD-L1 expression and overexpression of the endothelin B receptor (ETBR) on ovarian cancer cells is correlated to decreased numbers of tumor infiltrating lymphocytes, and associated with poor prognosis [48]. For these reasons, TIL adoptive transfer as a therapeutic approach may seem obvious in ovarian cancer, and a number of pilot clinical trials support the feasibility of adoptive TIL therapy for ovarian cancer [55].
Renal cell carcinoma is one of the additional tumors considered immunogenic and with proven clinical benefit for immune modulatory treatments such as IL-2 or PD1 and CTLA-4 blockade [16, 28, 49, 109], however tumor-infiltrating lymphocytes has not yet proven feasible for adoptive T cell therapy in this disease. RCC TILs comprise effector T cells alongside regulatory T cells [9], and some data suggest that T cells in RCC TILs may differ from melanoma in being more differentiated and exhausted, suggesting that other means of culturing may be required to re-activating and expansion of RCC TILs in vitro [112].
We recently studied the clonotype composition in RCC and strikingly, most lesions studied were characterized by absence or very few clonally expanded T cells (Sittig et al. Manuscript in preparation). RCC is a more vascularized tumor than e.g., melanoma for which reason blood T cells may be a more prominent component of the TILs. However, as judged by phenotype analyses these TILs are very different from PBMC T cells e.g., by expression of PD-1 in a high frequency suggesting that these are not simply PBMC T cells. Moreover, it seems that cytoxicity even against autologous tumor cell lines is rarely detectable [8].
Head and neck squamous cell carcinoma (HNSCC) comprise TILs and it has been shown that TILs in HNSCC may be capable of killing autologous HNSCC cells [47]. We recently conducted a pre-clinical study on HNSCC TIL and demonstrated that these can be expanded with high dose IL-2, and were capable of killing autologous as well as allogeneic tumor cell lines [54]. Although not tested in the clinic these data suggests that ACT in HNSCC using TILs could be clinically relevant.
Phenotypes of Tumor Infiltrating Lymphocytes and Manipulation of Immune Checkpoints
Tumor infiltrating T cells may display a wide range of different phenotypes. In general, however, several studies have shown that CD8 T cells at the tumor site display markers of T cell exhaustion to a higher extend than do T cells in the blood or T cells from normal adjacent tissue [1, 112]. In melanomas CD8 tumor infiltrating T cells display a high expression of PD1 and CTLA-4, and are often co-expressing these inhibitory molecules. At the same time they have a high expression of HLA-DR, a marker for T cell activation and low expression of CD127, the IL-7 receptor alpha chain, as well as CD25 [1]. Also CD4 T cells showed high expression of the immune inhibitory molecules PD1 and CTLA-4. Furthermore it was evident that the PD1 positive fraction of the TILs display impaired effector functions [1]. Also in prostate cancer, PD1 is significantly expressed by CD8 TILs [94], and this marker has shown prognostic value in breast cancer [43], as well as in several other cancers as indicated above. Recent studies in breast cancer patients show evidence of exhaustion in both blood and tumor already at early stage disease, with upregulation of PD1 and CD69. With disease progression exhaustion became even more evident and TILs displayed more terminally differentiated markers than T cell from peripheral blood [85]. Thus, it appears that the tumor induce a general immune suppression already at very early stage of disease.
The immune regulatory checkpoint PD1 and CTLA-4 play a major role in the exhausted phenotype of TILs, and blockade of these has shown to not only increase effector functions but also increase infiltration into tumor lesions [23, 24]. One mechanism by which PD-1 blockade may promote tumor-infiltration as through increased levels of IFN-γ inducible chemokines [82]. Also PD-L1, which is primarily expressed by tumor cells or tumor associated fibroblasts, may regulate T-cell infiltration. PD-L1 expression has been correlated with decreased CD8 infiltration, but the correlation to overall survival or disease progression remains controversial and may differ with regard to different malignancies [19, 38, 41]. However, overall there is clear association between the manipulation of these immune-blockade pathways, increased T-cell functionality, increased tumor infiltration and the clinical effect of these novel immunotherapies.
T Cell Functionality in the Hostile Tumor Microenviroment
As described elsewhere T cells may express inhibitory surface molecules the blockade of which may lead to clinically relevant anti-tumor T-cell responses. A detailed description of local tumor suppression of T cell function is beyond the scope of this review and a number of mechanisms may be at plays that render T cells in tumors non-functional. As already mentioned Treg may play a role in inhibiting CD8 T cell responses by various mechanisms [22], and MDSC and other cells on the innate immune system may play a similar role [28, 69, 76, 93]. For instance, expression of the enzymes Indoleamine-2,3-dioxygenase 1 (IDO-1) [72], IDO-2 or tryptophan 2,3-dioxygenase (TDO) [114], or tryptophan hydroxylase-1 [75] all metabolize tryptophan and thereby deplete Trp in the microenvironment. These enzymes may be expressed by cancer cells or by cells of the innate immune system; e.g. dendritic cells. Interestingly, we recently identified IDO derived peptides to be recognized by cytotoxic T cells in cancer patients suggesting a counter response against cells expressing immune suppressive IDO [97–100]. Arginine (Arg) may be depleted at the tumor site similarly due to expression of arginase-1 which is expressed by myeloid derived suppressor cells (MDSC) [17]. Lack of the amino acids Trp and Arg leads to activation of the GCN2 signaling pathway [71] which renders the T cell non-functional and eventually the T cell will go into apoptosis. Other cell types—including cancer cells express the IMPACT protein which is inhibiting GCN2 signaling thereby leaving these cells more resistant to amino acid deprivation [45].
Cancer cells are characterized by an altered metabolism – glycolysis – which metabolize glucose to lactate which is secreted to the microenvironment rather than further metabolized in the mitocondria. This altered metabolism is governed by activated oncogenes and/or hypoxia. Lactate impact negatively on the function of cells of the immune systems and lactate is detrimental to T cell function; cytokine production and cytotoxic capacity [29, 35].
Several other mechanisms have been described. To this end, some cancer cells secrete cytokines that are directly immune suppressive e.g., interleukin 10 and Transforming growth factor β (TGF-β), or cytokines that attract cells that diverge the anti-cancer immune response in a more pro-tumor direction (e.g., interleukin 6 and 8, and thymic stromal lymphopoietin (TSLP)) [81].
T Cell Infiltration in Virus Induced Cancers
Several types of cancers are known to have a viral origin. These include – as mentioned above – a fraction of the head and neck cancers - which similarly to cervical cancer is induced by Human Papillomavirus (HPV). In addition, Kaposi Sarcoma induced by Epstein Barr Virus, adult T-cell leukemia induced by Human T-Lymphotropic virus and Merkel cell carcinoma induced by Merkel cell polyomavirus [33] are viral induced cancers. Worldwide, the WHO International Agency for Research on Cancer estimated that in 2002 17.8 % of human cancers were caused by infection, with 11.9 % being caused by one of seven different viruses [79]. The importance of this is that these cancers might be easily prevented through vaccination (as recently developed for human papilloma virus), diagnosed with simple blood tests, and treated with less-toxic antiviral compounds. However, when the cancer has developed virus particles are no longer formed, and the anti-viral agents are by itself inefficient to combat the cancer – still therapeutic strategies for immunological recognition of the integrated oncogenes of viral origin may still be very relevant. For several of these cancers immune reactivity and immune-infiltration is associated with favourable clinical outcome, well in line with the observation that most virus-induced cancers develop preferentially in immune suppressed individuals.
Cervical cancer induced by human papillomavirus infection is the most frequently observed virus induced cancer. T cell infiltration to cervical cancer with higher CD8/CD4 ratio and higher CD8/Treg ratio is correlated with absence of lymph node metastasis in patients with large early-stage cervical cancer, and even further supported by the presence of a strong systemic anti-vial T cell response [83]. One of the cell-types affecting T cell infiltration in cervical cancer is the tumor associated macrophages, shown to induce regulatory phenotype characterized by IL10 production and Foxp3 expression in both CD4 and CD8 T cells. Furthermore, the tumor associated macrophages reduced the antigen-specific effector functions and led to decreased T-cell infiltration [64]. Along this line, CXCL12 expression in cervical cancer is associated with increased infiltration of Foxp3 positive T cells and correlates with disease progression [53]. Furthermore, as for tumor infiltrating lymphocytes in many indications CD8 T cells infiltrating cervical cancer is frequently expressing the inhibitory molecule PD-1, indicating that these patients could benefit from PD-1 blockade [56]. Indeed, therapeutic vaccination therapy seems to benefit patients with HPV-associated neoplastic lesions [58, 113], and immunotherapy has great clinical promise for HPV induced cervical cancer.
For the more recently discovered virus induced cancer, Merkel Cell Carcinoma (Feng et al., 2008), a clear correlation between CD8 T-cell infiltration to tumor lesions and favorable clinical outcome has been established, indicating a role for host T-cells in recognition of tumor cells [80]. Also for this cancer virus specific T cells has been identified in cancer patients [52], and clinic strategies to boost these T cell responses and improve infiltration is likely to benefit clinical outcome.
In general the group of virus-induced cancers are very attractive in terms of immunotherapy and especially adoptive T cell therapy as defined foreign antigens are expressed by these cancers, allowing for clear distinguish in immune recognition of healthy and malignant tissue.
Concluding Remarks and Perspectives
A tumor is a very complex structure, and as evident from this review not only the type of infiltrating cells are important by also the location, and moreover the specific phenotype and function of that cells in the particular environment. This is a major challenge for our understanding of lymphocyte infiltration and the impact on clinical outcome. Furthermore signatures of infiltration differ between different cancer types – and it is likely that infiltration patterns are as heterogeneous as recently observed for the mutation pattern in RCC. There seems to be a consensus that CD8 infiltration is a good prognostic marker in most malignancies analysed – however the impact of CD8 T cells on clinical outcome may differ and is difficult to quantify. New initiatives like the immunoscore assessment will enable us to better understand this impact.
Lymphocyte infiltration has gained increasing interest with the approval/testing of anti-CTLA4, PD1, and PD-L1 antibodies, as these may not only put a block on immune suppression but also influence the microenvironment to increase T-cell infiltration. Clinical efficacy of these therapies is likely to depend on the ability to induce T-cell infiltration – and as a potential biomarker for response it will likely be critical to evaluate these criteria in future therapeutic trials.
The new developments in the field; immunoscope, ACT based on TIL, as well as of the use of antibodies to unleash T cell reactivity to tumors bring hope that increased insight into the complex cellular and molecular interactions in the tumor will pave the way for development of new treatment strategies that build on the biological characteristics of the tumor microenviroment.
References
Ahmadzadeh M, Johnson LA, Heemskerk B, Wunderlich JR, Dudley ME, White DE, Rosenberg SA (2009) Tumor antigen-specific CD8 T cells infiltrating the tumor express high levels of PD-1 and are functionally impaired. Blood
Andersen MH, Becker JC, thor Straten P (2005) Regulators of apoptosis: Suitable targets for immune therapy of cancer. Nat Rev Drug Discov 4:399–409
Andersen MH, Gehl J, Reker S, Ostergaard Pedersen L, Becker JC, Geertsen PF, thor Straten P (2003) Dynamic changes of specific T cell responses to melanoma correlate with IL-2 administration. Semin Cancer Biol 13:449–459
Andersen MH, Ostergaard Pedersen L, Capeller B, Bröcker EB, Becker JC, thor Straten P (2001) Spontaneous cytoxic T-cell responses against survivin-derived MHC class I restricted T-cell epitopes in situ as well as ex vivo in cancer patients. Cancer Res 61:5964–5968
Andersen MH, Reker S, Becker JC, thor Straten P (2004) The melanoma inhibitor of apoptosis protein ML-IAP: a target for spontaneous cytotoxic T-cell responses. J Invest Dermatol 122:392–399
Andersen MH, Sorensen BS, Brimnes M, Svane IM, Becker JC, thor Straten P (2009) Identification of heme Oxygenase-1-specific regulatory CD8+ T cells in cancer patients. J Clin Investig 119:2245–2256
Andersen RS, Albaek Thrue C, Junker N, Lyngaa R, Donia M, Ellebaek E, Svane IM, Schumacher TN, thor Straten P, Hadrup SR (2012) Dissection of T cell antigen specificity in human melanoma. Cancer Res
Angevin E, Kremer F, Gaudin C, Hercend T, Triebel F (1997) Analysis of T-cell immune response in renal cell carcinoma: polarization to type 1-like differentiation pattern, clonal T-cell expansion and tumor-specific cytotoxicity. Int J Cancer 72:431–440
Attig S, Hennenlotter J, Pawelec G, Klein G, Koch SD, Pircher H, Feyerabend S, Wernet D, Stenzl A, Rammensee HG, Gouttefangeas C (2009) Simultaneous infiltration of polyfunctional effector and suppressor T cells into renal cell carcinomas. Cancer Res 69:8412–8419
Azimi F, Scolyer RA, Rumcheva P, Moncrieff M, Murali R, McCarthy SW, Saw RP, Thompson JF (2012) Tumor-infiltrating lymphocyte grade is an independent predictor of sentinel lymph node status and survival in patients with cutaneous melanoma. J Clin Oncol 30:2678–2683
Bakker AB, Schreurs MW, de Boer AJ, Kawakami Y, Rosenberg SA, Adema GJ, Figdor CG (1994) Melanocyte lineage-specific antigen gp100 is recognized by melanoma-derived tumor-infiltrating lymphocytes. J Exp Med 179:1005–1009
Beatty GL, Chiorean EG, Fishman MP, Saboury B, Teitelbaum UR, Sun W, Huhn RD, Song W, Li D, Sharp LL, Torigian DA, O’Dwyer PJ, Vonderheide RH (2011) CD40 agonists alter tumor stroma and show efficacy against pancreatic carcinoma in mice and humans. Science 331:1612–1616
Berger TG, Haendle I, Schrama D, Lüftl M, Bauer N, Ostergaard Pedersen L, Schuler-Thurner B, Hohenberger W, thor Straten P, Schuler G, Becker JC (2004) Circulation and homing of melanoma-reactive T cell clones to both cutaneaus and visceral metastases. Int J Cancer 111:229–237
Besser MJ, Shapira-Frommer R, Treves AJ, Zippel D, Itzhaki O, Hershkovitz L, Levy D, Kubi A, Hovav E, Chermoshniuk N, Shalmon B, Hardan I, Catane R, Markel G, Apter S, Ben-Nun A, Kuchuk I, Shimoni A, Nagler A, Schachter J (2010) Clinical responses in a phase II study using adoptive transfer of short-term cultured tumor infiltration lymphocytes in metastatic melanoma patients. Clin Cancer Res 16:2646–2655
Besser MJ, Shapira-Frommer R, Treves AJ, Zippel D, Itzhaki O, Schallmach E, Kubi A, Shalmon B, Hardan I, Catane R, Segal E, Markel G, Apter S, Nun AB, Kuchuk I, Shimoni A, Nagler A, Schachter J (2009) Minimally cultured or selected autologous tumor-infiltrating lymphocytes after a lympho-depleting chemotherapy regimen in metastatic melanoma patients. J Immunother 32:415–423
Brahmer JR, Tykodi SS, Chow LQ, Hwu WJ, Topalian SL, Hwu P, Drake CG, Camacho LH, Kauh J, Odunsi K, Pitot HC, Hamid O, Bhatia S, Martins R, Eaton K, Chen S, Salay TM, Alaparthy S, Grosso JF, Korman AJ, Parker SM, Agrawal S, Goldberg SM, Pardoll DM, Gupta A, Wigginton JM (2012) Safety and Activity of Anti-PD-L1 Antibody in Patients with Advanced Cancer. N Engl J Med
Bronte V, Zanovello P (2005) Regulation of immune responses by L-arginine metabolism. Nat Rev Immunol 5:641–654
Castle JC, Kreiter S, Diekmann J, Lower M, van de Roemer N, de Graaf J, Selmi A, Diken M, Boegel S, Paret C, Koslowski M, Kuhn AN, Britten CM, Huber C, Tureci O, Sahin U (2012) Exploiting the mutanome for tumor vaccination. Cancer Res 72:1081–1091
Cho YA, Yoon HJ, Lee JI, Hong SP, Hong SD (2011) Relationship between the expressions of PD-L1 and tumor-infiltrating lymphocytes in oral squamous cell carcinoma. Oral Oncol 47:1148–1153
Clark WH (1991) Tumour progression and the nature of cancer. Br J Cancer 64:631–644
Coulie PG, Somville M, Lehmann F, Hainaut P, Brasseur F, Devos R, Boon T (1992) Precursor frequency analysis of human cytolytic T lymphocytes directed against autologous melanoma cells. Int J Cancer 50:289–297
Curiel TJ, Coukos G, Zou L, Alvarez X, Cheng P, Mottram P, Evdemon-Hogan M, Conejo-Garcia JR, Zhang L, Burow M, Zhu Y, Wei S, Kryczek I, Daniel B, Gordon A, Myers L, Lackner A, Disis ML, Knutson KL, Chen L, Zou W (2004) Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat Med 10:942–949
Curran MA, Kim M, Montalvo W, Al-Shamkhani A, Allison JP (2011) Combination CTLA-4 blockade and 4-1BB activation enhances tumor rejection by increasing T-cell infiltration, proliferation, and cytokine production. PLoS One 6:e19499
Curran MA, Montalvo W, Yagita H, Allison JP (2010) PD-1 and CTLA-4 combination blockade expands infiltrating T cells and reduces regulatory T and myeloid cells within B16 melanoma tumors. Proc Natl Acad Sci U S A 107:4275–4280
Dadmarz R, Sgagias MK, Rosenberg SA, Schwartzentruber DJ (1995) CD4+ T lymphocytes infiltrating human breast cancer recognise autologous tumor in an MHC-class-II restricted fashion. Cancer Immunol Immunother 40:1–9
Dadmarz RD, Ordoubadi A, Mixon A, Thompson CO, Barracchini KC, Hijazi YM, Steller MA, Rosenberg SA, Schwartzentruber DJ (1996) Tumor-infiltrating lymphocytes from human ovarian cancer patients recognize autologous tumor in an MHC class II-restricted fashion. Cancer J Sci Am 2:263–272
Donia M, Hansen M, Sendrup SL, Iversen TZ, Ellebaek E, Andersen MH, thor Straten P, Svane IM (2012) Methods to Improve Adoptive T-Cell Therapy for Melanoma: IFN-gamma Enhances Anticancer Responses of Cell Products for Infusion. J Invest Dermatol 10
Donskov F, von der Maase H (2006) Impact of immune parameters on long-term survival in metastatic renal cell carcinoma. J Clin Oncol 24:1997–2005
Droge W, Roth S, Altmann A, Mihm S (1987) Regulation of T-cell functions by L-lactate. Cell Immunol 108:405–416
Dunn GP, Old LJ, Schreiber RD (2004) The Three Es of Cancer Immunoediting. Annu Rev Immunol 22:329–360
DuPage M, Mazumdar C, Schmidt LM, Cheung AF, Jacks T (2012) Expression of tumour-specific antigens underlies cancer immunoediting. Nature 482:405–409
Ellebaek E, Iversen TZ, Junker N, Donia M, Engell-Noerregaard L, Met O, Holmich LR, Andersen RS, Hadrup SR, Andersen MH, thor Straten P, Svane IM (2012) Adoptive cell therapy with autologous tumor infiltrating lymphocytes and low-dose Interleukin-2 in metastatic melanoma patients. J Transl Med 10:169
Feng H, Shuda M, Chang Y, Moore PS (2008) Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science 319:1096–1100
Ferradini L, Mackensen A, Genevee C, Bosq J, Duvillard P, Avril MF, Hercend T (1993) Analysis of T cell receptor variability in tumor-infiltrating lymphocytes from a human regressive melanoma. Evidence for in situ T cell clonal expansion. J Clin Invest 91:1183–1190
Fischer K, Hoffmann P, Voelkl S, Meidenbauer N, Ammer J, Edinger M, Gottfried E, Schwarz S, Rothe G, Hoves S, Renner K, Timischl B, Mackensen A, Kunz-Schughart L, Andreesen R, Krause SW, Kreutz M (2007) Inhibitory effect of tumor cell-derived lactic acid on human T cells. Blood 109:3812–3819
Fridman WH, Pages F, Sautes-Fridman C, Galon J (2012) The immune contexture in human tumours: impact on clinical outcome. Nat Rev Cancer 12:298–306
Friedman KM, Prieto PA, Devillier LE, Gross CA, Yang JC, Wunderlich JR, Rosenberg SA, Dudley ME (2012) Tumor-specific CD4+ melanoma tumor-infiltrating lymphocytes. J Immunother 35:400–408
Gadiot J, Hooijkaas AI, Kaiser AD, van Tinteren H, van Boven H, Blank C (2011) Overall survival and PD-L1 expression in metastasized malignant melanoma. Cancer 117:2192–2201
Galon J, Costes A, Sanchez-Cabo F, Kirilovsky A, Mlecnik B, Lagorce-Pages C, Tosolini M, Camus M, Berger A, Wind P, Zinzindohoue F, Bruneval P, Cugnenc PH, Trajanoski Z, Fridman WH, Pages F (2006) Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science 313:1960–1964
Galon J, Franck P, Marincola FM, Angell HK, Thurin M, Lugli A, Zlobec I, Berger A, Bifulco C, Botti G, Tatangelo F, Britten CM, Kreiter S, Chouchane L, Delrio P, Hartmann A, Asslaber M, Maio M, Masucci GV, Mihm M, Vidal-Vanaclocha F, Allison JP, Gnjatic S, Hakansson L, Huber C, Singh-Jasuja H, Ottensmeier C, Zwierzina H, Laghi L, Grizzi F, Ohashi PS, Shaw PA, Clarke BA, Wouters BG, Kawakami Y, Hazama S, Okuno K, Wang E, O’Donnell-Tormey J, Lagorce C, Pawelec G, Nishimura MI, Hawkins R, Lapointe R, Lundqvist A, Khleif SN, Ogino S, Gibbs P, Waring P, Sato N, Torigoe T, Itoh K, Patel PS, Shukla SN, Palmqvist R, Nagtegaal ID, Wang Y, D’Arrigo C, Kopetz S, Sinicrope FA, Trinchieri G, Gajewski TF, Ascierto PA, Fox BA (2012) Cancer classification using the Immunoscore: a worldwide task force. J Transl Med 10:205
Gao Q, Wang XY, Qiu SJ, Yamato I, Sho M, Nakajima Y, Zhou J, Li BZ, Shi YH, Xiao YS, Xu Y, Fan J (2009) Overexpression of PD-L1 significantly associates with tumor aggressiveness and postoperative recurrence in human hepatocellular carcinoma. Clin Cancer Res 15:971–979
Gerlinger M, Rowan AJ, Horswell S, Larkin J, Endesfelder D, Gronroos E, Martinez P, Matthews N, Stewart A, Tarpey P, Varela I, Phillimore B, Begum S, McDonald NQ, Butler A, Jones D, Raine K, Latimer C, Santos CR, Nohadani M, Eklund AC, Spencer-Dene B, Clark G, Pickering L, Stamp G, Gore M, Szallasi Z, Downward J, Futreal PA, Swanton C (2012) Intratumor heterogeneity and branched evolution revealed by multiregion sequencing. N Engl J Med 366:883–892
Ghebeh H, Barhoush E, Tulbah A, Elkum N, Al-Tweigeri T, Dermime S (2008) FOXP3+ Tregs and B7-H1+/PD-1+ T lymphocytes co-infiltrate the tumor tissues of high-risk breast cancer patients: Implication for immunotherapy. BMC Cancer 8:57
Gooden MJ, de Bock GH, Leffers N, Daemen T, Nijman HW (2011) The prognostic influence of tumour-infiltrating lymphocytes in cancer: a systematic review with meta-analysis. Br J Cancer 105:93–103
Habibi D, Jalili RB, Forouzandeh F, Ong CJ, Ghahary A (2010) High expression of IMPACT protein promotes resistance to indoleamine 2,3-dioxygenase-induced cell death. J Cell Physiol 225:196–205
Hadrup SR, Bakker AH, Shu CJ, Andersen RS, van Veluw J, Hombrink P, Castermans E, thor Straten P, Blank C, Haanen JB, Heemskerk MH, Schumacher TN (2009) Parallel detection of antigen-specific T-cell responses by multidimensional encoding of MHC multimers. Nat Methods 6:520–526
Hald J, Rasmussen N, Claesson MH (1995) Tumour-infiltrating lymphocytes mediate lysis of autologous squamous cell carcinomas of the head and neck. Cancer Immunol Immunother 41:243–250
Hamanishi J, Mandai M, Iwasaki M, Okazaki T, Tanaka Y, Yamaguchi K, Higuchi T, Yagi H, Takakura K, Minato N, Honjo T, Fujii S (2007) Programmed cell death 1 ligand 1 and tumor-infiltrating CD8+ T lymphocytes are prognostic factors of human ovarian cancer. Proc Natl Acad Sci U S A 104:3360–3365
Hodi FS, O’Day SJ, McDermott DF, Weber RW, Sosman JA, Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel JC, Akerley W, van den Eertwegh AJ, Lutzky J, Lorigan P, Vaubel JM, Linette GP, Hogg D, Ottensmeier CH, Lebbe C, Peschel C, Quirt I, Clark JI, Wolchok JD, Weber JS, Tian J, Yellin MJ, Nichol GM, Hoos A, Urba WJ (2010) Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med 363:711–723
Hunder NN, Wallen H, Cao J, Hendricks DW, Reilly JZ, Rodmyre R, Jungbluth A, Gnjatic S, Thompson JA, Yee C (2008) Treatment of metastatic melanoma with autologous CD4+ T cells against NY-ESO-1. N Engl J Med 358:2698–2703
Hwang WT, Adams SF, Tahirovic E, Hagemann IS, Coukos G (2012) Prognostic significance of tumor-infiltrating T cells in ovarian cancer: a meta-analysis. Gynecol Oncol 124:192–198
Iyer JG, Afanasiev OK, McClurkan C, Paulson K, Nagase K, Jing L, Marshak JO, Dong L, Carter J, Lai I, Farrar E, Byrd D, Galloway D, Yee C, Koelle DM, Nghiem P (2011) Merkel cell polyomavirus-specific CD8(+) and CD4(+) T-cell responses identified in Merkel cell carcinomas and blood. Clin Cancer Res 17:6671–6680
Jaafar F, Righi E, Lindstrom V, Linton C, Nohadani M, Van NS, Lloyd T, Poznansky J, Stamp G, Dina R, Coleman DV, Poznansky MC (2009) Correlation of CXCL12 expression and FoxP3+ cell infiltration with human papillomavirus infection and clinicopathological progression of cervical cancer. Am J Pathol 175:1525–1535
Junker N, Andersen MH, Wenandy L, Dombernowsky SL, Kiss K, Sorensen CH, Therkildsen MH, Von BC, Andersen E, thor Straten P, Svane IM (2011) Bimodal ex vivo expansion of T cells from patients with head and neck squamous cell carcinoma: a prerequisite for adoptive cell transfer. Cytotherapy
Kandalaft LE, Powell DJ Jr, Singh N, Coukos G (2011) Immunotherapy for ovarian cancer: what’s next? J Clin Oncol 29:925–933
Karim R, Jordanova ES, Piersma SJ, Kenter GG, Chen L, Boer JM, Melief CJ, van der Burg SH (2009) Tumor-expressed B7-H1 and B7-DC in relation to PD-1+ T-cell infiltration and survival of patients with cervical carcinoma. Clin Cancer Res 15:6341–6347
Kawakami Y, Eliyahu S, Sakaguchi K, Robbins PF, Rivoltini L, Yannelli JR, Appella E, Rosenberg SA (1994) Identification of the immunodominant peptides of the MART-1 human melanoma antigen recognized by the majority of HLA-A2- restricted tumor infiltrating lymphocytes. J Exp Med 180:347–352
Kenter GG, Welters MJ, Valentijn AR, Lowik MJ, der Meer DM B-v, Vloon AP, Essahsah F, Fathers LM, Offringa R, Drijfhout JW, Wafelman AR, Oostendorp J, Fleuren GJ, van der Burg SH, Melief CJ (2009) Vaccination against HPV-16 oncoproteins for vulvar intraepithelial neoplasia. N Engl J Med 361:1838–1847
Kryczek I, Wei S, Zhu G, Myers L, Mottram P, Cheng P, Chen L, Coukos G, Zou W (2007) Relationship between B7-H4, regulatory T cells, and patient outcome in human ovarian carcinoma. Cancer Res 67:8900–8905
Kvistborg P, Shu CJ, Heemskerk B, Fankhauser M, Thrue CA, Toebes M, van Rooij N, Linnemann C, van Buuren MM, Urbanus JH, Beltman JB, thor Straten P, Li YF, Robbins PF, Besser MJ, Schachter J, Kenter GG, Dudley ME, Rosenberg SA, Haanen JB, Hadrup SR, Schumacher TN (2012) TIL therapy broadens the tumor-reactive CD8(+) T cell compartment in melanoma patients. Oncoimmunology 1:409–418
Le Gal FA, Widmer VM, Dutoit V, Rubio-Godoy V, Schrenzel J, Walker PR, Romero PJ, Valmori D, Speiser DE, Dietrich PY (2006) Tissue Homing and Persistence of Defined Antigen-Specific CD8(+) Tumor-Reactive T-Cell Clones in Long-Term Melanoma Survivors. J Invest Dermatol
Lee KH, Panelli MC, Kim CJ, Riker AI, Bettinotti MP, Roden MM, Fetsch P, Abati A, Rosenberg SA, Marincola FM (1998) Functional dissociation between local and systemic immune response during anti-melanoma peptide vaccination. J Immunol 161:4183–4194
Lennerz V, Fatho M, Gentilini C, Frye RA, Lifke A, Ferel D, Wolfel C, Huber C, Wolfel T (2005) The response of autologous T cells to a human melanoma is dominated by mutated neoantigens. Proc Natl Acad Sci U S A 102:16013–16018
Lepique AP, Daghastanli KR, Cuccovia IM, Villa LL (2009) HPV16 tumor associated macrophages suppress antitumor T cell responses. Clin Cancer Res 15:4391–4400
Mason D (1998) A very high level of crossreactivity is an essential feature of the T-cell receptor. Immunol Today 9:395–404
Matsushita H, Vesely MD, Koboldt DC, Rickert CG, Uppaluri R, Magrini VJ, Arthur CD, White JM, Chen YS, Shea LK, Hundal J, Wendl MC, Demeter R, Wylie T, Allison JP, Smyth MJ, Old LJ, Mardis ER, Schreiber RD (2012) Cancer exome analysis reveals a T-cell-dependent mechanism of cancer immunoediting. Nature 482:400–404
Matzinger P, Kamala T (2011) Tissue-based class control: the other side of tolerance. Nat Rev Immunol 11:221–230
Mendez R, Aptsiauri N, Del CA, Maleno I, Cabrera T, Ruiz-Cabello F, Garrido F, Garcia-Lora A (2009) HLA and melanoma: multiple alterations in HLA class I and II expression in human melanoma cell lines from ESTDAB cell bank. Cancer Immunol Immunother 58:1507–1515
Meyer C, Sevko A, Ramacher M, Bazhin AV, Falk CS, Osen W, Borrello I, Kato M, Schadendorf D, Baniyash M, Umansky V (2011) Chronic inflammation promotes myeloid-derived suppressor cell activation blocking antitumor immunity in transgenic mouse melanoma model. Proc Natl Acad Sci U S A 108:17111–17116
Moerch U, Schrama D, Guldberg P, Seremet T, Zeuthen J, Becker JC, thor Straten P (2000) Comparative delineantion of T cell clonotypes in coexisting syngeneic B16 melanoma. Cancer Immunol Immunother 49:426–432
Munn DH, Sharma MD, Baban B, Harding HP, Zhang Y, Ron D, Mellor AL (2005) GCN2 kinase in T cells mediates proliferative arrest and anergy induction in response to indoleamine 2,3-dioxygenase. Immunity 22:633–642
Munn DH, Sharma MD, Lee JR, Jhaver KG, Johnson TS, Keskin DB, Marshall B, Chandler P, Antonia SJ, Burgess R, Slingluff CL Jr, Mellor AL (2002) Potential regulatory function of human dendritic cells expressing indoleamine 2,3-dioxygenase. Science 297:1867–1870
Nakano O, Sato M, Naito Y, Suzuki K, Orikasa S, Aizawa M, Suzuki Y, Shintaku I, Nagura H, Ohtani H (2001) Proliferative activity of intratumoral CD8(+) T-lymphocytes as a prognostic factor in human renal cell carcinoma: clinicopathologic demonstration of antitumor immunity. Cancer Res 61:5132–5136
Nitta T, Oksenberg JR, Rao NA, Steinman RM (1990) Predominant expression of T cell receptor V alpha 7 in tumor- infiltrating lymphocytes of uveal melanoma. Science 249:672–674
Nowak EC, de Vries VC, Wasiuk A, Ahonen C, Bennett KA, Le Mercier I, Ha DG, Noelle RJ, (2012) Tryptophan hydroxylase-1 regulates immune tolerance and inflammation. J Exp Med
Ostrand-Rosenberg S, Sinha P, Beury DW, Clements VK (2012) Cross-talk between myeloid-derived suppressor cells (MDSC), macrophages, and dendritic cells enhances tumor-induced immune suppression. Semin Cancer Biol
Panelli MC, Bettinotti MP, Lally K, Ohnmacht GA, Li Y, Robbins P, Riker A, Rosenberg SA, Marincola FM (2000) A tumor-infiltrating lymphocyte from a melanoma metastasis with decreased expression of melanoma differentiation antigens recognizes MAGE-12. J Immunol 164:4382–4392
Panelli MC, Riker A, Kammula U, Wang E, Lee KH, Rosenberg SA, Marincola FM (2000) Expansion of tumor-T cell pairs from fine needle aspirates of melanoma metastases. J Immunol 164:495–504
Parkin DM (2006) The global health burden of infection-associated cancers in the year 2002. Int J Cancer 118:3030–3044
Paulson KG, Iyer JG, Tegeder AR, Thibodeau R, Schelter J, Koba S, Schrama D, Simonson WT, Lemos BD, Byrd DR, Koelle DM, Galloway DA, Leonard JH, Madeleine MM, Argenyi ZB, Disis ML, Becker JC, Cleary MA, Nghiem P (2011) Transcriptome-wide studies of merkel cell carcinoma and validation of intratumoral CD8+ lymphocyte invasion as an independent predictor of survival. J Clin Oncol 29:1539–1546
Pedroza-Gonzalez A, Xu K, Wu TC, Aspord C, Tindle S, Marches F, Gallegos M, Burton EC, Savino D, Hori T, Tanaka Y, Zurawski S, Zurawski G, Bover L, Liu YJ, Banchereau J, Palucka AK (2011) Thymic stromal lymphopoietin fosters human breast tumor growth by promoting type 2 inflammation. J Exp Med 208:479–490
Peng W, Liu C, Xu C, Lou Y, Chen J, Yang Y, Yagita H, Overwijk WW, Lizee G, Radvanyi L, Hwu P (2012) PD-1 Blockade Enhances T-cell Migration to Tumors by Elevating IFN-gamma Inducible Chemokines. Cancer Res 72:5209–5218
Piersma SJ, Jordanova ES, van Poelgeest MI, Kwappenberg KM, van der Hulst JM, Drijfhout JW, Melief CJ, Kenter GG, Fleuren GJ, Offringa R, van der Burg SH (2007) High number of intraepithelial CD8+ tumor-infiltrating lymphocytes is associated with the absence of lymph node metastases in patients with large early-stage cervical cancer. Cancer Res 67:354–361
Pilon-Thomas S, Kuhn L, Ellwanger S, Janssen W, Royster E, Marzban S, Kudchadkar R, Zager J, Gibney G, Sondak VK, Weber J, Mule JJ, Sarnaik AA (2012) Efficacy of Adoptive Cell Transfer of Tumor-infiltrating Lymphocytes After Lymphopenia Induction for Metastatic Melanoma. J Immunother 35:615–620
Poschke I, De BJ, Mao Y, Kiessling R (2012) Tumor-induced changes in the phenotype of blood-derived and tumor-associated T cells of early stage breast cancer patients. Int J Cancer 131:1611–1620
Quezada SA, Simpson TR, Peggs KS, Merghoub T, Vider J, Fan X, Blasberg R, Yagita H, Muranski P, Antony PA, Restifo NP, Allison JP (2010) Tumor-reactive CD4(+) T cells develop cytotoxic activity and eradicate large established melanoma after transfer into lymphopenic hosts. J Exp Med 207:637–650
Radvanyi LG, Bernatchez C, Zhang M, Fox P, Miller P, Chacon J, Wu RC, Lizee G, Mahoney S, Alvarado G, Glass M, Johnson V, McMannis JD, Shpall EJ, Prieto VG, Papadopoulos NE, Kim KB, Homsi J, Bedikian AY, Hwu WJ, Patel S, Ross MI, Lee JE, Gershenwald JE, Lucci A, Royal R, Cormier JN, Davies MA, Mansaray R, Fulbright OJ, Toth C, Ramachandran R, Wardell S, Gonzalez A, Hwu P (2012) Specific lymphocyte subsets predict response to adoptive cell therapy using expanded autologous tumor-infiltrating lymphocytes in metastatic melanoma patients. Clin Cancer Res
Robbins PF, El-Gamil M, Li YF, Zeng G, Dudley ME, Rosenberg SA (2002) Multiple HLA class II-restricted melanocyte differentiation antigens are recognized by tumor-infiltrating lymphocytes from a patient with melanoma. J Immunol 169:6036–6047
Romero P, Pannetier C, Herman J, Jongeneel CV, Cerottini JC, Coulie PG (1995) Multiple specificities in the repertoire of a melanoma patients cytolytic T lymphocytes directed against tumor antigen MAGE-1.A1. J Exp Med 182:1019–1028
Rosenberg SA, Yang JC, Sherry RM, Kammula US, Hughes MS, Phan GQ, Citrin DE, Restifo NP, Robbins PF, Wunderlich JR, Morton KE, Laurencot CM, Steinberg SM, White DE, Dudley ME (2011) Durable complete responses in heavily pretreated patients with metastatic melanoma using T-cell transfer immunotherapy. Clin Cancer Res 17:4550–4557
Salvi S, Segalla F, Rao S, Arienti F, Sartori M, Bratina G, Caronni E, Anichini A, Clemente C, Parmiani G, Sensi M (1995) Overexpression of the T-cell receptor beta-chain variable region TCRBV14 in HLA-A2-matched primary human melanomas. Cancer Res 55:3374–3379
Sato E, Olson SH, Ahn J, Bundy B, Nishikawa H, Qian F, Jungbluth AA, Frosina D, Gnjatic S, Ambrosone C, Kepner J, Odunsi T, Ritter G, Lele S, Chen YT, Ohtani H, Old LJ, Odunsi K (2005) Intraepithelial CD8+ tumor-infiltrating lymphocytes and a high CD8+/regulatory T cell ratio are associated with favorable prognosis in ovarian cancer. Proc Natl Acad Sci U S A 102:18538–18543
Schmidt H, Suciu S, Punt CJ, Gore M, Kruit W, Patel P, Lienard D, von der Maase H, Eggermont AM, Keilholz U (2007) Pretreatment levels of peripheral neutrophils and leukocytes as independent predictors of overall survival in patients with American Joint Committee on Cancer Stage IV Melanoma: results of the EORTC 18951 Biochemotherapy Trial. J Clin Oncol 25:1562–1569
Sfanos KS, Bruno TC, Meeker AK, De Marzo AM, Isaacs WB, Drake CG (2009) Human prostate-infiltrating CD8+ T lymphocytes are oligoclonal and PD-1+. Prostate 69:1694–1703
Sharma MD, Hou DY, Liu Y, Koni PA, Metz R, Chandler P, Mellor AL, He Y, Munn DH (2009) Indoleamine 2,3-dioxygenase controls conversion of Foxp3+ Tregs to TH17-like cells in tumor-draining lymph nodes. Blood 113:6102–6111
Sharma P, Shen Y, Wen S, Yamada S, Jungbluth AA, Gnjatic S, Bajorin DF, Reuter VE, Herr H, Old LJ, Sato E (2007) CD8 tumor-infiltrating lymphocytes are predictive of survival in muscle-invasive urothelial carcinoma. Proc Natl Acad Sci U S A 104:3967–3972
Sorensen RB, Berge-Hansen L, Junker N, Hansen CA, Hadrup SR, Schumacher TN, Svane IM, Becker JC, thor Straten P, Andersen MH (2009) The immune system strikes back: cellular immune responses against indoleamine 2,3-dioxygenase. PLoS One 4:e6910
Sorensen RB, Hadrup SR, Svane IM, Hjortso MC, thor Straten P, Andersen MH (2011) Indoleamine 2,3-dioxygenase specific cytotoxic T cells as immune regulators. Blood 117:2200–2210
Sorensen RB, Kollgaard T, Andersen RS, van den Berg J, Svane IM, thor Straten P, Andersen MH (2011) Spontaneous cytotoxic T-cell reactivity against indoleamine 2-dioxygenase-2. Cancer Res 71:2038–2044
Sorensen RB, thor Straten P, Andersen MH (2009) Comment on “Reduced cytotoxic function of effector CD8+ T cells is responsible for indoleamine 2,3-dioxygenase-dependent immune suppression”. J Immunol 183:6040
thor Straten P, Becker JC, Guldberg P, Zeuthen J (1999) In Situ T cells in Melanoma. Cancer Immunol Immunother 48:386–395
thor Straten P, Guldberg P, Andersen MH, Moerch U, Seremet T, Siedel C, Zeuthen J, Reisfeld RA, Becker JC (2001) In situ cytokine therapy: redistribution of clonally expanded therapeutic T-cell clones. Eur J Immunol 31:250–258
thor Straten P, Guldberg P, Grønbæk K, Zeuthen J, Becker JC (1999) In Situ T-Cell Responses against Melanoma Comprise High Numbers of Locally Expanded T-Cell Clonotypes. J Immunol 163:443–447
thor Straten P, Guldberg P, Seremet T, Reisfeld RA, Zeuthen J, Becker JC (1998) Activation of pre-existing T-cell clones by targeted interleukin 2 therapy. Proc Natl Acad Sci U S A 95:8785–8790
thor Straten P, Kirkin AF, Siim E, Dahlstrom K, Drzewiecki KT, Seremet T, Zeuthen J, Becker JC, Guldberg P (2000) Tumor infiltrating lymphocytes (TIL) in melanoma comprise high numbers of T-cell clonotypes that are lost during in vitro culture. Clin Immunol 96:94–99
thor Straten P, Schøller J, Hou Jensen K, Zeuthen J (1994) Preferential usage of T-cell receptor alpha beta variable regions among tumor-infiltrating lymphocytes in primary human malignant melanomas. Int J Cancer 56:78–86
thor Straten P, Schrama D, Andersen MH, Becker JC (2004) T cell clonotypes in cancer. Transl Med 2:1–11
Toebes M, Coccoris M, Bins A, Rodenko B, Gomez R, Nieuwkoop NJ, van de Kasteele W, Rimmelzwaan GF, Haanen JB, Ovaa H, Schumacher TN (2006) Design and use of conditional MHC class I ligands. Nat Med 12:246–251
Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, Powderly JD, Carvajal RD, Sosman JA, Atkins MB, Leming PD, Spigel DR, Antonia SJ, Horn L, Drake CG, Pardoll DM, Chen L, Sharfman WH, Anders RA, Taube JM, McMiller TL, Xu H, Korman AJ, Jure-Kunkel M, Agrawal S, McDonald D, Kollia GD, Gupta A, Wigginton JM, Sznol M (2012) Safety, Activity, and Immune Correlates of Anti-PD-1 Antibody in Cancer. N Engl J Med
Valmori D, Dutoit V, Lienard D, Lejeune F, Speiser DE, Rimoldi D, Cerundolo V, Dietrich PY, Cerottini JC, Romero P (2000) Tetramer-guided analysis of TCR beta-chain usage reveals a large repertoire of melan-A-specific CD8+ T cells in melanoma patients. J Immunol 165:533–538
Vukmanovic-Stejic M, Zhang Y, Cook JE, Fletcher JM, McQuaid A, Masters JE, Rustin MH, Taams LS, Beverley PC, Macallan DC, Akbar AN (2006) Human CD4+ CD25hi Foxp3+ regulatory T cells are derived by rapid turnover of memory populations in vivo. J Clin Invest 116:2423–2433
Wang QJ, Hanada KI, Robbins PF, Li YF, Yang JC (2012) Distinctive Features of the Differentiated Phenotype and Infiltration of Tumor-reactive Lymphocytes in Clear Cell Renal Cell Carcinoma. Cancer Res
Welters MJ, Kenter GG, Piersma SJ, Vloon AP, Lowik MJ, der Meer DM B-v, Drijfhout JW, Valentijn AR, Wafelman AR, Oostendorp J, Fleuren GJ, Offringa R, Melief CJ, van der Burg SH (2008) Induction of tumor-specific CD4+ and CD8+ T-cell immunity in cervical cancer patients by a human papillomavirus type 16 E6 and E7 long peptides vaccine. Clin Cancer Res 14:178–187
Witkiewicz AK, Costantino CL, Metz R, Muller AJ, Prendergast GC, Yeo CJ, Brody JR (2009) Genotyping and expression analysis of IDO2 in human pancreatic cancer: a novel, active target. J Am Coll Surg 208:781–787
Wölfel T, Hauer M, Schneider J, Serrano M, Wolfel C, Klehmann-Hieb E, De Plaen E, Hankeln T, Meyer zum Büschenfelde K, Beach D (1995) A p16INK4a-insensitive CDK4 mutant targeted by cytolytic T lymphocytes in a human melanoma. Science 269:1281–1284
Zhou J, Dudley ME, Rosenberg SA, Robbins PF (2005) Persistence of multiple tumor-specific T-cell clones is associated with complete tumor regression in a melanoma patient receiving adoptive cell transfer therapy. J Immunother 28:53–62
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Hadrup, S., Donia, M. & thor Straten, P. Effector CD4 and CD8 T Cells and Their Role in the Tumor Microenvironment. Cancer Microenvironment 6, 123–133 (2013). https://doi.org/10.1007/s12307-012-0127-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12307-012-0127-6