
Abstract
Background
Recent studies have demonstrated a direct involvement of B7-H1, PD-1 and FOXP3 molecules in the immune escape of cancer. B7-H1 is an inhibitory molecule that binds to PD-1 on T lymphocytes, while FOXP3 is a marker for regulatory T cells (Tregs). We have previously demonstrated the association of B7-H1-expressing T infiltrating lymphocytes (TIL) with high-risk breast cancer patients while other studies reported the involvement of FOXP3+ Tregs as a bad prognostic factor in breast tumors. Although the co-existence between the two types of cells has been demonstrated in vitro and animal models, their relative infiltration and correlation with the clinicopathological parameters of cancer patients have not been well studied. Therefore, we investigated TIL-expressing the B7-H1, PD-1, and FOXP3 molecules, in the microenvironment of human breast tumors and their possible association with the progression of the disease.
Methods
Using immunohistochemistry, tumor sections from 62 breast cancer patients were co-stained for B7-H1, PD-1 and FOXP3 molecules and their expression was statistically correlated with factors known to be involved in the progression of the disease.
Results
A co-existence of B7-H1+ T lymphocytes and FOXP3+ Tregs was evidenced by the highly significant correlation of these molecules (P < .0001) and their expression by different T lymphocyte subsets was clearly demonstrated. Interestingly, concomitant presence of FOXP3+ Tregs, B7-H1+ and PD-1+ TIL synergistically correlated with high histological grade (III) (P < .001), estrogen receptor negative status (P = .017), and the presence of severe lymphocytic infiltration (P = .022).
Conclusion
Accumulation of TIL-expressing such inhibitory molecules may deteriorate the immunity of high-risk breast cancer patients and this should encourage vigorous combinatorial immunotherapeutic approaches targeting Tregs and B7-H1/PD-1 molecules.
Similar content being viewed by others

Background
It is widely believed that dysfunction in the immune system of cancer patients allows the tumor cells to escape a process termed immunosurveillance [1–3]. A T lymphocytes inhibitory molecule named B7-H1 (also called PD-L1) expressed by antigen presenting cells has been shown to induce T lymphocyte anergy and/or apoptosis after ligation to its T lymphocytes receptor PD-1 [4–7]. B7-H1 has been shown to be directly involved in the protection of cancer cells from activated T lymphocytes [8]. Indeed, the expression of this molecule and its receptor has been described in several malignancies [9–17] where a strong link between its expression by the cancer cells and the patient clinicopathological status has been demonstrated in some of these malignancies [9, 11, 13, 16]. We have shown previously that B7-H1 (PD-L1) is expressed in the tumor tissues of 50% of breast cancer patients and its expression was significantly associated with some important prognostic factors linked to high-risk patients [18].
Regulatory T cells (Tregs) are a subset of T lymphocytes that regulate the immune response by suppressing the proliferation and cytokines production of effector T lymphocytes [19, 20]. Tregs are thus important for protecting our body by suppressing auto-reactive T lymphocytes. FOXP3, a forkhead/winged helix transcription factor was found to be essential for the development and control of Tregs [21, 22]. It has been used recently as a biomarker and a prognostic factor for malignant human tumor as reviewed in [23]. A direct link between the presence of Tregs and progression of ovarian carcinoma has been demonstrated where human tumor FOXP3+ Tregs were found to suppress tumor specific immunity and contribute to reduced survival of these patients [24]. In breast cancer, an increase in Tregs population both in peripheral blood and tumor tissues was also reported [25] and a recent study demonstrated a significant intratumoral infiltration of FOXP3+ Tregs in high-risk breast cancer patients and those at risk of late relapse [26].
Compelling evidences indicate a key role for the involvement of B7-H1 molecule in the development of Tregs. Induction of adaptive CD4+CD25+ Tregs which develop in the periphery from CD4+CD25- naïve T lymphocytes (in contrast to natural Tregs formed in the thymus) could not be formed in mice which are B7 -/- [27]. In addition, mice vascular endothelial cells were able to induce CD4+CD25+FOXP3+ Tregs only in the presence of B7-H1 [28]. Another study has demonstrated the involvement of B7-H1 as an essential element for the induction of Tregs after intra-tracheal delivery of an alloantigen [29]. Furthermore, B7-H1 was found to affect the function of Tregs in which its blockade decreased the inhibitory effect of such cells [30] and blockade of B7-H1/PD-1 interactions abrogated Tregs-mediated immunoregulation [31]. Although these studies have demonstrated the interaction between B7-H1 molecule and Tregs in animal models their co-existence in cancer patients was not evaluated. This is the first study to investigate the correlation between FOXP3+ Tregs and TIL-expressing B7-H1 and PD-1 in breast cancer patients. We have shown that their co-infiltration is strongly associated with high-risk prognostic factors and those T lymphocytes expressing FOXP3, B7-H1 and PD-1 molecules are of different T lymphocytes subsets.
Methods
Patients and samples collection
This study was conducted in accordance with Helsinki Declaration and all patients signed a consent form approved by the Research Ethics Committee of King Faisal Specialist Hospital and Research Center (KFSH&RC). The study was approved by the Research Advisory Council (RAC) of KFSH&RC (RAC# 2030 034).
Breast cancer specimens were collected from primary tumors of 68 patients including the 44 patients reported in our previous study [18] (median age 44 years) who were seeking treatments and had to undergo surgery (breast conservative surgery or total mastectomy) at KFSH&RC from 2003 to 2006. From the selected patients, 6 patients were removed from the study as they had no detectable TIL. Normal breast tissues were also obtained from 2 healthy women undergoing a plastic surgery and designated as BP. Upon excision of tissues by a surgeon, an anatomical pathologist obtained sample of the tumor tissue, denoted T, and an adjacent normal breast tissue from the same breast having the tumor, denoted N. Tissues from both T and N were processed as described before [18]. Briefly, they were fixed in formalin and embedded in paraffin for routine histopathological analysis while other piece was snap frozen in liquid nitrogen, preserved at -80°C and sectioned using a cryostat.
Immunohistochemistry
Routine formalin-fixed, paraffin-embedded hospital tests were evaluated by immunohistochemistry for Her/2neu, Estrogen receptor, and progesterone receptor status as described before [18] while immunohistochemistry staining of frozen tissue sections were carried out as follows:
(a) Single staining
B7-H1 and PD-1 detection were carried out as described previously on fresh cryogenic sections with slight modifications [18, 32] in which a very brief fixation in 2% paraformaldehyde (Fisher scientific) in PBS at RT (4 minutes for B7-H1 and 10 minutes for PD-1) was initially applied to improve the morphology of section. Briefly, sections were incubated for 15 minutes in 0.3% hydrogen peroxide solution and 0.1% sodium azide (Sigma, Saint Louis, MO, USA). Sections were blocked with 10% goat serum (DAKO, Denmark) for 30 minutes followed by B7-H1 antibody (MIH1 clone, ebioscience, San Diego, CA, USA) or PD-1 antibody (J116 clone, ebioscience,) diluted at 1:50 overnight. After washing, sections were stained for 30 minutes at RT, with Labeled Polymer (EnVision+) horseradish peroxidase (HRP) detection kit (DAKO). Colors were developed using DAB (Novocastra, Newcastle upon Tyne, UK) or AEC (Sigma) and sections were counterstained with instant hematoxylin (Shandon, Pittsburgh, PA, USA).
(b) Double staining
Sections were washed in PBS, incubated with paraformaldehyde for 4 minutes and blocked in H2O2 and goat serum as described above. The primary and secondary antibodies were added as follows: For CD3/FOXP3, CD8/FOXP3, CD8/PD-1 or CD8/B7-H1 double staining the 2 primary antibodies from two different species, mouse anti-FOXP3 (236A/E7 clone, ebioscience) diluted at 1:100, mouse anti-B7-H1 (clone MIH1, ebioscience) diluted at 1:25 or mouse anti-PD-1 antibody (J116 clone, ebioscience) diluted at 1:50 were mixed with rabbit anti-CD3 (Dako) at 1:50 dilution or rabbit anti-CD8 (Abcam, Cambridge, UK) at 1:75 dilution, together in 10% human AB serum (Cambrex) and incubated for 2 h at RT or overnight at 4°C. Sections were washed 3 times in PBS and incubated for 30 minutes at RT with a secondary antibody: swine anti-rabbit AP (Dako) diluted at 1:50 in a ready-to-use polymer (Envision+, Dako) linked to HRP. After washing 3 times with PBS, substrates were added starting with fuschin red (Dako) followed by DAB (Novocastra). Slides were counterstained with instant hematoxylin (Shandon). FOXP3 and B7-H1 double staining were carried out as described above except the primary antibodies used were rabbit anti-FOXP3 (polyclonal, Abcam) at 1:500 dilution mixed with mouse anti-B7-H1 (clone MIH1, ebioscience) at 1:25 dilution and overnight incubation at 4°C. FOXP3 and PD-1 double staining were started with a single staining of PD-1 as before, however after color development with DAB, sections were boiled for 3 minutes in an antigen retrieval citrate solution (pH = 6.0) to remove previously attached antibodies. FOXP3 antigen was then stained with mouse anti-FOXP3 antibody (236A/E7 clone, ebioscience) diluted at 1:100 for 2 hours at RT followed by goat anti-mouse IgG1 AP (Southern Biotech, Birmingham, Alabama, USA) for 30 minutes at RT. The color for FOXP3 staining was developed with Fuschin Red (Dako) and sections were counterstained with instant hematoxylin (Shandon).
Interpretation of stained tumor sections
The B7-H1 and PD-1 staining were scored by an anatomical pathologist (AT) as described previously [18]. FOXP3+ T lymphocytes were scored in 5–10% increments as a percentage of total CD3+ staining cells in several high magnification fields. Intratumoral FOXP3+/CD3+ cells were only considered and counted while intrastromal FOXP3+ cells were disregarded. FOXP3+ cells with moderate or high intensity (++ or +++) were considered Tregs while weak staining (+) were ignored. Due to the co localization of the CD3 and PD-1 molecules in the T lymphocyte membranes, double staining of PD-1 or B7-H1 and CD3 were assumed based on single-staining of sequential slides. The cut off point for positive and negative for FOXP3, B7-H1 and PD-1 was 5% (5% of total CD3+ T-lymphocytes).
Statistical Analysis
Statistical analyses were used to determine the association between the B7-H1 expression in TIL or FOXP3+ Tregs infiltration and the patients' clinico-pathological parameters. Nominal parameters were analyzed using the fisher exact test. Comparisons of the independent variables with the dependent variable were performed using a simple logistic regression analysis. Significance was defined as the probability of a type one error of < 5%, and 95% confidence intervals were included. The software package SAS was used for these analyses.
Results
Frequencies of Tregs, B7-H1+ and PD-1+ T lymphocytes in normal and breast cancer tissues
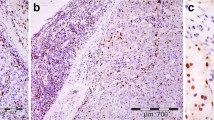
In order to determine the frequencies of Tregs, B7-H1 and PD-1 expressing-T lymphocytes in normal breast tissues we stained sections obtained from breast patients undergoing plastic surgery (BP) as well as normal breast tissues adjacent to breast cancer tumors (N) for the expression of FOXP3, B7-H1 and PD-1 molecules. There were a few T lymphocytes in both N and BP in which FOXP3+ Tregs represented <5% of the total CD3+ TIL (Figure 1A). Similarly, T lymphocytes in normal tissues were negative for B7-H1 (Figure 1B). However, PD-1+ T lymphocytes were abundant in normal tissues with up to 30% of CD3+ cells co-expressing the PD-1 molecule (Figure 1C). In contrast to normal breast tissues, 56% of breast cancer patients had a higher Tregs frequency in their tumor tissues in which 5–50% of CD3+ TILs were Tregs (Figure 1D). Similarly, B7-H1 was expressed in TILs of 54% of breast cancer patients' tumor tissues in which 5–80% of TILs express this molecule. B7-H1 expression was not restricted to T lymphocytes as 30% of breast cancer patients showed also B7-H1 expression in their tumor cells (Figure 1E). This result was also described in our previous study [18]. The PD-1 molecule was expressed in TILs in 60% of the patients in which 5–70% of TIL express this molecule (Figure 1F). The expression pattern of the PD-1 molecule was membranous and restricted to T lymphocytes only.

Immunohistochemical staining of FOXP3+ T regs B7-H1 +and PD-1+ in T lymphocytes of breast tissues. Representative micrographs at × 530 magnification of (A&D) CD3/FOXP3 double staining (red color, membranous for CD3 and brown nuclear color for FOXP3 expression). (B&E) B7-H1 single staining (brown color, membranous/cytoplasmic expression). (C&F) PD-1 single staining (brown color, membranous expression). Upper panel (A-C) is sections for normal breast duct and lower panel (D-F) is sections for infiltrating ductal carcinoma of the breast.
Both intratumoral and intrastromal FOXP3+CD3+ cells were present in the patients tissues (Figure 2A) and the expression level of FOXP3 in the intratumoral Tregs ranged from weak to intense (Figure 2B). Interestingly, most of FOXP3 intensely stained Tregs were found in close proximity to tumor cells. PD-1+ T lymphocytes were found both intrastromal and intratumoral, where they commonly seen in clumps, or as clumps/single cells respectively.

Immunohistochemical staining showing intratumoral and intrastromal FOXP3+ T regs in breast cancer tissues. Representative micrographs at × 530 magnification of CD3+ (red color membranous expression) and FOXP3+ (brown color nuclear expression) TIL of (A) intrastromal (left) and intratumoral (right) sections. Solid arrow indicates a CD3+/FOXP3+ Treg cell and dashed arrow indicates a CD3+/FOXP3- cell. (B) Intratumoral FOXP3+ TIL with different staining intensity (Intense, Medium, and weak). Cells were counterstained with hematoxylin.
Correlation of FOXP3, B7-H1 and PD-1 expression in TIL with clinicopathological parameters of patients
Breast cancer tissues infiltrated with intratumoral FOXP3+ Tregs were found to be significantly associated with patients who had bad prognostic factors such as large tumor size (P = .029), histological grade III (P < .001), estrogen receptor negative status (P = .010) and severe lymphocytic infiltration (P = .051). Intratumoral FOXP3+ Tregs were found also to be associated with progesterone receptor negative status and lymph node metastasis although they did not reach statistical significance (Table 1). The correlation of the expression B7-H1 in tumor cells and FOXP3+ Tregs was also studied. There was a correlation between B7-H1 in tumor cells and FOXP3+ Tregs (P < .019) (Table 1).
We have reported previously on the expression of B7-H1 in 50% of 44 breast cancer patients; and found its expression in TIL to be significantly associated with large tumor size, high histological grade III, Her2/neu positive status and severe lymphocytic infiltration [18]. We further examined the B7-H1 expression in TIL of 62 breast cancer patients and correlated its expression with the same prognostic factors. Interestingly, very similar correlation was recorded for B7-H1+ TIL, and this was significantly associated with high histological grade III (P = .002), estrogen receptor negative status (P = .029), and sever lymphocytic infiltration (P = .017). There was also a correlation with large tumor size with a borderline significance (P = .072) (Table 1). There was also a correlation between B7-H1 in tumor cells and B7-H1+ TIL (P < .002) (Table 1). Although PD-1 molecule was also expressed by T lymphocytes infiltrating normal breast tissues lacking B7-H1, its presence in tumor tissues is considered essential for the inhibitory effect of B7-H1 molecule which is abundant in the tumor tissues. Therefore, we tested whether PD-1 is concomitantly expressed in the same patients along with B7-H1+ TIL and FOXP3+ Tregs. We have found that PD-1+ TILs were abundant in patients with high histological grade III (P = .001), estrogen receptor negative status (P = .001) and progesterone receptor negative status (P < .001) (Table 1). There was also a correlation between B7-H1 in tumor cells and PD-1+ TIL (P < .002) (Table 1).
We have found that B7-H1+, PD-1+ TIL and FOXP3+ Tregs were significantly associated with similar bad prognostic factors. Therefore, we further investigated the correlation between B7-H1+ TIL or PD-1+ TIL and FOXP3+ Tregs infiltration in breast tumors. There was a highly significant correlation between the expression of B7-H1 in TIL and intratumoral FOXP3+ Tregs (P < .0001) as shown by ANOVA test (Figure 3A) and by linear regression analysis (P < .004 and R2 = 0.3) (Figure 3B). There was also a significant correlation between FOXP3+ Tregs infiltration and PD-1 expression in TIL (P = .007) as shown by ANOVA (Figure 3C).

Correlations between FOXP3+ T regs , B7-H1 + TIL and PD-1 + TIL. (A) statistical analysis of FOXP3+ Tregs infiltration in tumor tissues with B7-H1+ TIL, as analyzed with ANOVA of three groups of breast cancer patients with different FOXP3+ Tregs infiltration (Low = 0 to 4%, Medium = 5 to 14% and High 15% and above of total CD3+ TIL), (B) their linear correlation analysis and (C) statistical analysis of FOXP3+ Tregs infiltration with PD-1+ TIL in breast cancer as analyzed with ANOVA of three groups of breast cancer patients with different FOXP3+ Tregs infiltration (Low = 0 to 4%, Medium = 5 to 14% and High 15% and above of total CD3+ TIL).
Correlation of combined FOXP3, and B7-H1 expression in TIL with clinicopathological parameters of patients
We next tested whether a combination of FOXP3 and B7-H1 molecules will have a synergistic effect on the correlation with the patients' clinicopathological parameters. We used a combination of the two factors; FOXP3+ Tregs and B7-H1 expression in TIL in the presence of PD-1 expression (only patients with B7-H1-expressing TIL were considered positive when PD-1+ TIL were present in the same tissue). The combined molecules maintained a significant correlation with the same prognostic factors (Table 2).
Expression of FOXP3, B7-H1 and PD-1 molecules by different subsets of TIL
We next asked whether FOXP3, PD-1 and B7-H1 molecules are expressed by different T lymphocyte subsets. Double-staining on 6 selected samples which had high FOXP3+ Tregs, high B7-H1+ TIL and PD-1+ TIL was performed. The selections were based on sections that had a good morphology to make the interpretation easier. Double-staining assays showed that B7-H1 and FOXP3 molecules were generally expressed by different T lymphocyte subsets (most B7-H1+ TIL were FOXP3- while only very few FOXP3+ Tregs co-express the B7-H1 molecule). In addition, the distribution pattern of B7-H1+ TIL was different from that of FOXP3+ Tregs; Tregs were single cells distributed over the section of the tumor tissues while B7-H1+ TIL aggregated in clumps (Figure 4A). Similarly, PD-1 and FOXP3 molecules were expressed by different T lymphocyte subsets (Figure 4B). Furthermore, double staining assays demonstrated that FOXP3 and CD8 were mainly expressed by two different T lymphocyte subsets (Figure 4C) and only very few CD8+ TIL (< 5%) were FOXP3+. Therefore, it seems that most FOXP3+ Tregs described here are of the CD4+ subset since they were restricted to CD3+ T lymphocytes. On the other hand PD-1 molecule was mainly expressed by CD8+ T lymphocytes with only few CD8- population were positive for PD-1 (Figure 4D). B7-H1 was mainly expressed by CD8- T lymphocytes (Figure 4E) consistent with our previous observation of its expression by CD4+ T lymphocytes [18].

Immunohistochemical staining showing the expression of FOXP3, B7-H1 and PD-1 molecules by different subsets of TIL. Representative micrographs at × 530 magnification of (A) double staining of B7-H1 (brown color, membranous/cytoplasmic) and FOXP3 (red color, nuclear) in an area rich in TIL of tumor section. (B) Double staining of PD-1 (brown color, membranous) and FOXP3 (red color, nuclear) in sections from the same tumor as in A. (C) double staining of FOXP3 (brown color, nuclear) and CD8 (red color, membranous). (D) double staining of PD-1 (brown color, membranous) and CD8 (red color, membranous). Solid arrows indicate CD8+/PD-1+ T lymphocytes and dashed arrow indicates a CD8-/PD-1+ T lymphocyte. (E) Double staining of B7-H1 (brown color, membranous/cytoplasmic) and CD8 (red color, membranous). Solid arrow indicates a CD8+/B7-H1+ T lymphocyte and dashed arrow indicates a CD8-/B7-H1+ T lymphocyte.
Discussion
The immune system use intricate balance between positive and negative signals to protect the body from foreign agents while preventing autoimmunity [33]. This balance seems to be disturbed in various pathological conditions resulting in either inhibition of the immune system allowing invasion by tumor cells, or stimulation of the immune system to cause autoimmune diseases. For example, although interaction of B7-H1, a coinhibitory molecule, with its PD-1 ligand is important for prevention of autoimmunity [34], tumor cells use this interaction as a mechanism of immune evasion [35]. Furthermore, while the presence of Tregs is important for suppression of host immune response to prevent autoimmune diseases [36], tumor cells can recruit Tregs to inhibit anti-tumor immunity in cancer patients [37].
We have shown previously an abundant expression of B7-H1 molecule in high-risk breast cancer patients [18] who have highly proliferative tumor cells [32] and suggested the involvement of this molecule in the progression of the disease. In the present report we expanded our study to investigate the co-involvement of Tregs with B7-H1 in the immune evasion of breast cancer. Although the interaction between the two types of cells has been demonstrated in vitro and animal models, their relative infiltration and correlation with the clinicopathological parameters of cancer patients have not been well studied. In this study, we investigated FOXP3+ Tregs and TIL-expressing B7-H1 and its ligand PD-1 in breast cancer patients. We have used FOXP3 as a detection marker for Tregs as it became recently a biomarker for studying Tregs in malignant human cancers [23]. We have found that FOXP3+ Tregs are abundant in tumor tissues of 56% of breast cancer patients but absent in normal tissues adjacent to the tumors. FOXP3+ Tregs infiltrating tumors have been reported by other studies [25, 26, 38]. Abundant accumulation of Tregs in tumor tissues might be due to the induction of CD4+CD25+ Tregs from peripheral CD4+CD25- T lymphocytes [39, 40] and/or migration of Tregs from other parts of the body to the tumor area by chemotactic factors like CCL22 [24, 41]. This hypothesis is supported by increase of Tregs in peripheral blood of cancer patients [25, 42].
Standard prognostic factors linked to high-risk breast cancer patients include: young age, large tumor size, high histological grade, positive lymph node metastasis and negative hormonal receptors status [43]. We have demonstrated that FOXP3+ Tregs infiltrating tumor tissues correlates significantly with important bad prognostic factors: large tumor size (P = .029), high histological grade (P < .001) and estrogen receptor negative status (P = .010). Similar findings have been recently reported by Bates et al showing an association between high Tregs numbers, and high histological grade and estrogen receptor negative status [26]. Estrogen receptor negative breast tumors are generally associated with poor prognosis which is usually attributed to the association with high proliferative rate and lack of differentiation [44], that are features of progenitors of stem-like cells. Indeed it has been shown recently that stem cells obtained from both normal and cancer breast tissues lack the estrogen receptor [45]. Such stem cells were found to produce TGFβ1 [45] which is known to induce Tregs [46]. The association of FOXP3+ Tregs with bad prognostic factors may suggest a contribution of Tregs infiltrating breast cancer tissues to tumor escape from the immune system as their depletion lead to tumor rejection in animal models [47]. We have also confirmed the B7-H1 expression in TIL and its correlation with bad prognostic factors reported in our previous study [18]. The association of B7-H1 expression in TIL and FOXP3+ Tregs infiltration with the same bad prognostic factors suggests that both molecules correlate with each other. Indeed, there was a highly significant correlation between the expression of FOXP3+ Tregs and B7-H1 (P < .0001).
We have also found that PD-1 expression in TIL correlated with bad prognostic factors. PD-1 has been reported to be expressed in normal tissues to regulate self-reactive T cell responses [48]. In the present study PD-1 was expressed in up to 30% of T lymphocytes from normal breast tissues while up to 80% of TIL were PD-1+. The abundant expression of PD-1 in TIL together with B7-H1 (absent in normal tissues) seen in the present study may contribute to downregulation of the patients' immune response [4, 6]. Furthermore, combination of FOXP3+ Tregs, B7-H1+ TIL and PD-1+ TIL were found to be highly associated with patients who had high histological grade III (P < .001) and estrogen receptor negative tumors (P < .007) suggestive of synergistic contribution to bad prognosis. It seems that FOXP3, B7-H1 and PD-1 molecules co-expressed in the TIL of the same tumor tissues may have synergistic effect in weakening the immune response of patients. IL-2 may play a fundamental role in regulating the interaction between FOXP3+ Tregs, B7-H1+ TIL and PD-1+ TIL. B7-H1/PD-1 interaction contributes to induction of T lymphocyte anergy [4] and this can be restored in the presence of IL-2 [49]. On the other hand, Tregs are dependent on IL-2 for their expansion and function, and consumption of IL-2 present in the microenvironment may increase the inhibitory effect of B7-H1/PD-1 interaction [50].
We also investigated in the present study whether FOXP3, B7-H1 and PD-1 molecules are expressed by different T lymphocyte subsets. We have demonstrated that FOXP3+ Tregs is a separate population from PD-1+ and B7-H1+ TIL. Similar results have been recently reported in non-Hodgkin's lymphoma by Yang et al [41] who showed that PD-1 and B7-H1 expression were mainly found in a subset of non-Tregs T lymphocytes. The expression of FOXP3 was confirmed to be linked to CD4+ T lymphocytes as it has been reported by Roncador et al [51]. We have also confirmed our previous findings showing that B7-H1 was expressed mainly in CD4+ T lymphocytes [18]. On the other hand, we have shown that PD-1 molecules were expressed mainly by CD8+ T lymphocytes. It is important to mention that exhausted CD8+ T lymphocytes have been reported to express the PD-1 molecule and its blockade restored the function of these T lymphocytes [52].
One of the most important findings of this study is the co-localization of immunosuppressive molecules expressed by different subsets of T lymphocytes in a group of high-risk breast cancer patients. Indeed, induction of immunosuppressive molecules such as B7-H1, PD-1 and Tregs have been recently shown to counteract the anti-tumor effect of IL-12-based gene therapy in a transgenic mouse model of liver cancer [53]. Therefore, current interests in breast cancer immunotherapy should be focused on designing immunological tools to block B7-H1 and its PD-1 ligand and to deplete Tregs in addition to cancer vaccination [54]. Blocking B7-H1 and PD-1 molecules with monoclonal antibodies or soluble ligands has been shown to enhance cancer immunity in animal models [55]. In addition, elimination of Tregs by an anti-CD25 mAb enhanced anti-tumor immunity and induced tumor regression in animal models [47]. Furthermore, elimination of Tregs by IL-2-conjugated to diphtheria-toxin (ONTAK) enhanced vaccine-mediated anti-tumor immunity in cancer patients [56]. Interestingly, a very recent study, in an animal model of renal cell carcinoma, has shown that only triple treatment consisting of tumor vaccine, B7-H1 blockade, and Tregs depletion can result in a complete tumor regression and long lasting protective immunity [57] supporting the use of triple therapy treatment of cancer patients.
Conclusion
In conclusion, we have shown a concurrent and abundant infiltration of different immune suppressive subsets of T lymphocytes in the microenvironment of high-risk breast cancer patients. This interesting observation suggests the development of a new therapeutic modalities aiming at targeting B7-H1/PD-1 and Tregs in addition to still-developing immunotherapy.
References
Ahmad M, Rees RC, Ali SA: Escape from immunotherapy: possible mechanisms that influence tumor regression/progression. Cancer Immunology, Immunotherapy. 2004, 53 (10): 844-854. 10.1007/s00262-004-0540-x.
Rivoltini L, Canese P, Huber V, Iero M, Pilla L, Valenti R, Fais S, Lozupone F, Casati C, Castelli C, Parmiani G: Escape strategies and reasons for failure in the interaction between tumour cells and the immune system: how can we tilt the balance towards immune-mediated cancer control?. Expert Opinion on Biological Therapy. 2005, 5 (4): 463-476. 10.1517/14712598.5.4.463.
Seliger B: Strategies of tumor immune evasion. Biodrugs. 2005, 19 (6): 347-354. 10.2165/00063030-200519060-00002.
Freeman GJ, Long AJ, Iwai Y, Bourque K, Chernova T, Nishimura H, Fitz LJ, Malenkovich N, Okazaki T, Byrne MC, Horton HF, Fouser L, Carter L, Ling V, Bowman MR, Carreno BM, Collins M, Wood CR, Honjo T: Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J Exp Med. 2000, 192 (7): 1027-1034. 10.1084/jem.192.7.1027.
Dong H, Zhu G, Tamada K, Chen L: B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion. Nat Med. 1999, 5 (12): 1365-1369. 10.1038/70932.
Dong H, Strome SE, Salomao DR, Tamura H, Hirano F, Flies DB, Roche PC, Lu J, Zhu G, Tamada K, Lennon VA, Celis E, Chen L: Tumor-associated B7-H1 promotes T-cell apoptosis: a potential mechanism of immune evasion. Nat Med. 2002, 8 (8): 793-800.
Selenko-Gebauer N, Majdic O, Szekeres A, Hofler G, Guthann E, Korthauer U, Zlabinger G, Steinberger P, Pickl WF, Stockinger H, Knapp W, Stockl J: B7-H1 (programmed death-1 ligand) on dendritic cells is involved in the induction and maintenance of T cell anergy. J Immunol. 2003, 170 (7): 3637-3644.
Iwai Y, Ishida M, Tanaka Y, Okazaki T, Honjo T, Minato N: Involvement of PD-L1 on tumor cells in the escape from host immune system and tumor immunotherapy by PD-L1 blockade. Proc Natl Acad Sci USA. 2002, 99 (19): 12293-12297. 10.1073/pnas.192461099.
Konishi J, Yamazaki K, Azuma M, Kinoshita I, Dosaka-Akita H, Nishimura M: B7-H1 expression on non-small cell lung cancer cells and its relationship with tumor-infiltrating lymphocytes and their PD-1 expression. Clin Cancer Res. 2004, 10 (15): 5094-5100. 10.1158/1078-0432.CCR-04-0428.
Nomi T, Sho M, Akahori T, Hamada K, Kubo A, Kanehiro H, Nakamura S, Enomoto K, Yagita H, Azuma M, Nakajima Y: Clinical significance and therapeutic potential of the programmed death-1 ligand/programmed death-1 pathway in human pancreatic cancer. Clin Cancer Res. 2007, 13 (7): 2151-2157. 10.1158/1078-0432.CCR-06-2746.
Ohigashi Y, Sho M, Yamada Y, Tsurui Y, Hamada K, Ikeda N, Mizuno T, Yoriki R, Kashizuka H, Yane K, Tsushima F, Otsuki N, Yagita H, Azuma M, Nakajima Y: Clinical significance of programmed death-1 ligand-1 and programmed death-1 ligand-2 expression in human esophageal cancer. Clin Cancer Res. 2005, 11 (8): 2947-2953. 10.1158/1078-0432.CCR-04-1469.
Strome SE, Dong H, Tamura H, Voss SG, Flies DB, Tamada K, Salomao D, Cheville J, Hirano F, Lin W, Kasperbauer JL, Ballman KV, Chen L: B7-H1 blockade augments adoptive T-cell immunotherapy for squamous cell carcinoma. Cancer Res. 2003, 63 (19): 6501-6505.
Thompson RH, Gillett MD, Cheville JC, Lohse CM, Dong H, Webster WS, Krejci KG, Lobo JR, Sengupta S, Chen L, Zincke H, Blute ML, Strome SE, Leibovich BC, Kwon ED: Costimulatory B7-H1 in renal cell carcinoma patients: Indicator of tumor aggressiveness and potential therapeutic target. Proc Natl Acad Sci U S A. 2004, 101 (49): 17174-17179. 10.1073/pnas.0406351101.
Thompson RH, Kuntz SM, Leibovich BC, Dong H, Lohse CM, Webster WS, Sengupta S, Frank I, Parker AS, Zincke H, Blute ML, Sebo TJ, Cheville JC, Kwon ED: Tumor B7-H1 is associated with poor prognosis in renal cell carcinoma patients with long-term follow-up. Cancer Res. 2006, 66 (7): 3381-3385. 10.1158/0008-5472.CAN-05-4303.
Wintterle S, Schreiner B, Mitsdoerffer M, Schneider D, Chen L, Meyermann R, Weller M, Wiendl H: Expression of the B7-related molecule B7-H1 by glioma cells: a potential mechanism of immune paralysis. Cancer Res. 2003, 63 (21): 7462-7467.
Hamanishi J, Mandai M, Iwasaki M, Okazaki T, Tanaka Y, Yamaguchi K, Higuchi T, Yagi H, Takakura K, Minato N, Honjo T, Fujii S: Programmed cell death 1 ligand 1 and tumor-infiltrating CD8+ T lymphocytes are prognostic factors of human ovarian cancer. Proc Natl Acad Sci U S A. 2007, 104 (9): 3360-3365. 10.1073/pnas.0611533104.
Thompson RH, Dong H, Lohse CM, Leibovich BC, Blute ML, Cheville JC, Kwon ED: PD-1 is expressed by tumor-infiltrating immune cells and is associated with poor outcome for patients with renal cell carcinoma. Clin Cancer Res. 2007, 13 (6): 1757-1761. 10.1158/1078-0432.CCR-06-2599.
Ghebeh H, Mohammed S, Al-Omair A, Qattan A, Lehe C, Al-Qudaihi G, Elkum N, Alshabanah M, Bin Amer S, Tulbah A, Ajarim D, Al-Tweigeri T, Dermime S: The B7-H1 (PD-L1) T lymphocyte-inhibitory molecule is expressed in breast cancer patients with infiltrating ductal carcinoma: correlation with important high-risk prognostic factors. Neoplasia. 2006, 8 (3): 190-198. 10.1593/neo.05733.
Thornton AM, Shevach EM: CD4+CD25+ immunoregulatory T cells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2 production. Journal of Experimental Medicine. 1998, 188 (2): 287-296. 10.1084/jem.188.2.287.
Ng WF, Duggan PJ, Ponchel F, Matarese G, Lombardi G, Edwards AD, Isaacs JD, Lechler RI: Human CD4(+)CD25(+) cells: a naturally occurring population of regulatory T cells. Blood. 2001, 98 (9): 2736-2744. 10.1182/blood.V98.9.2736.
Hori S, Nomura T, Sakaguchi S: Control of regulatory T cell development by the transcription factor Foxp3.[see comment]. Science. 2003, 299 (5609): 1057-1061. 10.1126/science.1079490.
Yagi H, Nomura T, Nakamura K, Yamazaki S, Kitawaki T, Hori S, Maeda M, Onodera M, Uchiyama T, Fujii S, Sakaguchi S: Crucial role of FOXP3 in the development and function of human CD25+CD4+ regulatory T cells. International Immunology. 2004, 16 (11): 1643-1656. 10.1093/intimm/dxh165.
Schreiber TH: The Use of FoxP3 as a Biomarker and Prognostic Factor for Malignant Human Tumors. Cancer Epidemiol Biomarkers Prev. 2007, 16 (10): 1931-1934. 10.1158/1055-9965.EPI-07-0396.
Curiel TJ, Coukos G, Zou L, Alvarez X, Cheng P, Mottram P, Evdemon-Hogan M, Conejo-Garcia JR, Zhang L, Burow M, Zhu Y, Wei S, Kryczek I, Daniel B, Gordon A, Myers L, Lackner A, Disis ML, Knutson KL, Chen L, Zou W: Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nature Medicine. 2004, 10 (9): 942-949. 10.1038/nm1093.
Liyanage UK, Moore TT, Joo HG, Tanaka Y, Herrmann V, Doherty G, Drebin JA, Strasberg SM, Eberlein TJ, Goedegebuure PS, Linehan DC: Prevalence of regulatory T cells is increased in peripheral blood and tumor microenvironment of patients with pancreas or breast adenocarcinoma. J Immunol. 2002, 169 (5): 2756-2761.
Bates GJ, Fox SB, Han C, Leek RD, Garcia JF, Harris AL, Banham AH: Quantification of regulatory T cells enables the identification of high-risk breast cancer patients and those at risk of late relapse. Journal of Clinical Oncology. 2006, 24 (34): 5373-5380. 10.1200/JCO.2006.05.9584.
Liang S, Alard P, Zhao Y, Parnell S, Clark SL, Kosiewicz MM: Conversion of CD4+ CD25- cells into CD4+ CD25+ regulatory T cells in vivo requires B7 costimulation, but not the thymus. Journal of Experimental Medicine. 2005, 201 (1): 127-137. 10.1084/jem.20041201.
Krupnick AS, Gelman AE, Barchet W, Richardson S, Kreisel FH, Turka LA, Colonna M, Patterson GA, Kreisel D: Murine vascular endothelium activates and induces the generation of allogeneic CD4+25+Foxp3+ regulatory T cells. J Immunol. 2005, 175 (10): 6265-6270.
Aramaki O, Shirasugi N, Takayama T, Shimazu M, Kitajima M, Ikeda Y, Azuma M, Okumura K, Yagita H, Niimi M: Programmed death-1-programmed death-L1 interaction is essential for induction of regulatory cells by intratracheal delivery of alloantigen. Transplantation. 2004, 77 (1): 6-12. 10.1097/01.TP.0000108637.65091.4B.
Baecher-Allan C, Brown JA, Freeman GJ, Hafler DA: CD4+CD25high regulatory cells in human peripheral blood. J Immunol. 2001, 167 (3): 1245-1253.
Kitazawa Y, Fujino M, Wang Q, Kimura H, Azuma M, Kubo M, Abe R, Li XK: Involvement of the programmed death-1/programmed death-1 ligand pathway in CD4+CD25+ regulatory T-cell activity to suppress alloimmune responses. Transplantation. 2007, 83 (6): 774-782. 10.1097/01.tp.0000256293.90270.e8.
Ghebeh H, Tulbah A, Mohammed S, Elkum N, Amer SM, Al-Tweigeri T, Dermime S: Expression of B7-H1 in breast cancer patients is strongly associated with high proliferative Ki-67-expressing tumor cells. Int J Cancer. 2007
Armstrong A, Dermime S: Developing effective cancer vaccines: design and monitoring are critical. Br J Cancer. 2001, 84 (11): 1433-1436. 10.1054/bjoc.2001.1839.
Keir ME, Liang SC, Guleria I, Latchman YE, Qipo A, Albacker LA, Koulmanda M, Freeman GJ, Sayegh MH, Sharpe AH: Tissue expression of PD-L1 mediates peripheral T cell tolerance. J Exp Med. 2006, 203 (4): 883-895. 10.1084/jem.20051776.
Blank C, Gajewski TF, Mackensen A: Interaction of PD-L1 on tumor cells with PD-1 on tumor-specific T cells as a mechanism of immune evasion: implications for tumor immunotherapy. Cancer Immunol Immunother. 2005, 54 (4): 307-314. 10.1007/s00262-004-0593-x.
Bala KK, Moudgil KD: Induction and maintenance of self tolerance: the role of CD4+CD25+ regulatory T cells. Arch Immunol Ther Exp (Warsz). 2006, 54 (5): 307-321. 10.1007/s00005-006-0035-x.
Beyer M, Schultze JL: Regulatory T cells in cancer. Blood. 2006, 108 (3): 804-811. 10.1182/blood-2006-02-002774.
Leong PP, Mohammad R, Ibrahim N, Ithnin H, Abdullah M, Davis WC, Seow HF: Phenotyping of lymphocytes expressing regulatory and effector markers in infiltrating ductal carcinoma of the breast. Immunol Lett. 2006, 102 (2): 229-236. 10.1016/j.imlet.2005.09.006.
Chen W, Jin W, Hardegen N, Lei KJ, Li L, Marinos N, McGrady G, Wahl SM: Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J Exp Med. 2003, 198 (12): 1875-1886. 10.1084/jem.20030152.
Walker MR, Carson BD, Nepom GT, Ziegler SF, Buckner JH: De novo generation of antigen-specific CD4+CD25+ regulatory T cells from human CD4+CD25- cells. Proc Natl Acad Sci U S A. 2005, 102 (11): 4103-4108. 10.1073/pnas.0407691102.
Yang ZZ, Novak AJ, Stenson MJ, Witzig TE, Ansell SM: Intratumoral CD4+CD25+ regulatory T-cell-mediated suppression of infiltrating CD4+ T cells in B-cell non-Hodgkin lymphoma. Blood. 2006, 107 (9): 3639-3646. 10.1182/blood-2005-08-3376.
Miller AM, Lundberg K, Ozenci V, Banham AH, Hellstrom M, Egevad L, Pisa P: CD4+CD25high T cells are enriched in the tumor and peripheral blood of prostate cancer patients. J Immunol. 2006, 177 (10): 7398-7405.
Chang J, Clark GM, Allred DC, Mohsin S, Chamness G, Elledge RM: Survival of patients with metastatic breast carcinoma: importance of prognostic markers of the primary tumor. Cancer. 2003, 97 (3): 545-553. 10.1002/cncr.11083.
Hilsenbeck SG, Ravdin PM, de Moor CA, Chamness GC, Osborne CK, Clark GM: Time-dependence of hazard ratios for prognostic factors in primary breast cancer. Breast Cancer Res Treat. 1998, 52 (1–3): 227-237. 10.1023/A:1006133418245.
Shipitsin M, Campbell LL, Argani P, Weremowicz S, Bloushtain-Qimron N, Yao J, Nikolskaya T, Serebryiskaya T, Beroukhim R, Hu M, Halushka MK, Sukumar S, Parker LM, Anderson KS, Harris LN, Garber JE, Richardson AL, Schnitt SJ, Nikolsky Y, Gelman RS, Polyak K: Molecular definition of breast tumor heterogeneity. Cancer Cell. 2007, 11 (3): 259-273. 10.1016/j.ccr.2007.01.013.
Marie JC, Letterio JJ, Gavin M, Rudensky AY: TGF-beta1 maintains suppressor function and Foxp3 expression in CD4+CD25+ regulatory T cells. J Exp Med. 2005, 201 (7): 1061-1067. 10.1084/jem.20042276.
Onizuka S, Tawara I, Shimizu J, Sakaguchi S, Fujita T, Nakayama E: Tumor rejection by in vivo administration of anti-CD25 (interleukin-2 receptor alpha) monoclonal antibody. Cancer Res. 1999, 59 (13): 3128-3133.
Keir ME, Freeman GJ, Sharpe AH: PD-1 Regulates Self-Reactive CD8+ T Cell Responses to Antigen in Lymph Nodes and Tissues. J Immunol. 2007, 179 (8): 5064-5070.
Carter L, Fouser LA, Jussif J, Fitz L, Deng B, Wood CR, Collins M, Honjo T, Freeman GJ, Carreno BM: PD-1:PD-L inhibitory pathway affects both CD4(+) and CD8(+) T cells and is overcome by IL-2. Eur J Immunol. 2002, 32 (3): 634-643. 10.1002/1521-4141(200203)32:3<634::AID-IMMU634>3.0.CO;2-9.
Ahmadzadeh M, Rosenberg SA: IL-2 administration increases CD4+ CD25(hi) Foxp3+ regulatory T cells in cancer patients. Blood. 2006, 107 (6): 2409-2414. 10.1182/blood-2005-06-2399.
Roncador G, Brown PJ, Maestre L, Hue S, Martinez-Torrecuadrada JL, Ling KL, Pratap S, Toms C, Fox BC, Cerundolo V, Powrie F, Banham AH: Analysis of FOXP3 protein expression in human CD4+CD25+ regulatory T cells at the single-cell level. Eur J Immunol. 2005, 35 (6): 1681-1691. 10.1002/eji.200526189.
Barber DL, Wherry EJ, Masopust D, Zhu B, Allison JP, Sharpe AH, Freeman GJ, Ahmed R: Restoring function in exhausted CD8 T cells during chronic viral infection. Nature. 2006, 439 (7077): 682-687. 10.1038/nature04444.
Zabala M, Lasarte JJ, Perret C, Sola J, Berraondo P, Alfaro M, Larrea E, Prieto J, Kramer MG: Induction of immunosuppressive molecules and regulatory T cells counteracts the antitumor effect of interleukin-12-based gene therapy in a transgenic mouse model of liver cancer. J Hepatol. 2007, 1: 1-10.1007/s12072-007-5002-z.
Curigliano G, Spitaleri G, Dettori M, Locatelli M, Scarano E, Goldhirsch A: Vaccine immunotherapy in breast cancer treatment: promising, but still early. Expert Rev Anticancer Ther. 2007, 7 (9): 1225-1241. 10.1586/14737140.7.9.1225.
Hirano F, Kaneko K, Tamura H, Dong H, Wang S, Ichikawa M, Rietz C, Flies DB, Lau JS, Zhu G, Tamada K, Chen L: Blockade of B7-H1 and PD-1 by monoclonal antibodies potentiates cancer therapeutic immunity. Cancer Res. 2005, 65 (3): 1089-1096.
Dannull J, Su Z, Rizzieri D, Yang BK, Coleman D, Yancey D, Zhang A, Dahm P, Chao N, Gilboa E, Vieweg J: Enhancement of vaccine-mediated antitumor immunity in cancer patients after depletion of regulatory T cells. J Clin Invest. 2005, 115 (12): 3623-3633. 10.1172/JCI25947.
Webster WS, Thompson RH, Harris KJ, Frigola X, Kuntz S, Inman BA, Dong H: Targeting molecular and cellular inhibitory mechanisms for improvement of antitumor memory responses reactivated by tumor cell vaccine. J Immunol. 2007, 179 (5): 2860-2869.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1471-2407/8/57/prepub
Acknowledgements
We are very grateful to the administration of the Research Centre and the Office of Research Affairs (ORA) for their support. This work was sponsored by KFSH&RC (RAC # 2030 034). We would like to thank Drs. Monther Al-Alwan and Ayodele Alaiya for critical review for the manuscript, and Zuha Al-Mukhlafi and Manogaran Pulicat for analyzing the FACS data.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The author(s) declare that they have no competing interests.
Authors' contributions
HG designed the study, carried out the immunohistochemistry for B7-H1, coordinated the work and wrote the manuscript. EB carried out immunohistochemistry of PD-1, FOXP3, and CD8 molecules. AT (anatomical pathologist) read and interpreted the sections. NE (statistician) carried out the statistical analysis. TA (medical oncologist) participated in conceiving the study and provided the clinical data. SD (principal investigator) wrote the proposal, conceived, supervised the study, and wrote the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Ghebeh, H., Barhoush, E., Tulbah, A. et al. FOXP3+ Tregs and B7-H1+/PD-1+T lymphocytes co-infiltrate the tumor tissues of high-risk breast cancer patients: Implication for immunotherapy. BMC Cancer 8, 57 (2008). https://doi.org/10.1186/1471-2407-8-57
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2407-8-57