
Abstract
To investigate the expressions of key markers in the homologous recombination (HR) pathway and the correlation with clinicopathological parameters in serous ovarian cancer (SOC). We analyzed the protein expression of MRE11, MDC1, ATM, ATR and BRCA1 by immunohistochemistry (IHC) in 97 SOC samples, and correlated with clinical parameters including age, tumor grades, clinical stage, status of menstruation and chemotherapy. Low expression of MRE11 and MDC1 was detected in 14.4 % and 3.1 % of the patient samples, and negative expression of ATM, ATR and BRCA1 was found in 11.3 %, 6.3 % and 29.9 % of the patient samples, respectively. ATR deficiency was significantly associated with menopause (P = 0.025), strong expression of ATM (P = 0.017) and MRE11 (P = 0.040) with pre-menopausal SOC, strong expression of MRE11 (P = 0.016) with low tumor grade, and strong expression of BRCA1 (P = 0.015) with early clinical stage. In addition, low expression of MRE11 was strongly associated with negativity of ATR (P < 0.001) and BRCA1 (P = 0.004) Furthermore, ATR deficiency was associated with low expression of ATM (P = 0.028) and loss expression of BRCA1 (P = 0.009). Our results suggest that reduced expression or loss of proteins in HR pathway is associated with SOC development. Abnormality of MRE11 and BRCA1 are strongly associated with late clinical stage in SOC patients.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
DNA double-strand breaks (DSBs) are particularly hazardous to the cell since they cause base pair mismatch,[1] which is strongly associated with cancer susceptibility. Three mechanisms exist to repair DSBs: non-homologous end joining (NHEJ), microhomology-mediated end joining (MMEJ), and homologous recombination (HR).[2] HR-mediated repair requires one homologous sequence to accurately repair breaks. In contrast, MMEJ requires a 5–25 base pair microhomologous sequence, whilst NHEJ can function to directly religate broken ends in the absence of a homologous template. Moreover, HR repairs DSBs in the late S and G2 phases of the cell cycle when sister chromatids are readily available,[3] as opposed to MMEJ which occurs in S phase and NHEJ in the G0/G1 and early S phases. Sister chromatids are ideal templates for repair as they provide identical copies of the same chromosome. Therefore, HR plays an important role in the fidelity of DNA replication,[4] which is vital to the integrity and stability of the genome.
During an HR repair process, DNA lesions are first identified and several key repair proteins are recruited. These include checkpoint mediator proteins like the MRN complex (MRE11-NBS1-RAD50) and MDC1. The MRN complex slows down crossover progression in mitosis [5] and MDC1 facilitates signal transmission to downstream proteins.[6, 7] Next, ATM and the RAD3-related ATR kinase, part of the Phosphatidyl-Inositol 3 Kinase-like protein Kinase (PIKK) family, cascade signals which arrest cell cycle progression [8] and thus allow DNA repair to occur. RAD51 (RAD50 forms MRE11-NSB1-RAD50 complex) also interacts with breast cancer susceptibility genes 1 and 2 (BRCA1/BRCA2). Finally, ATM and ATR then directly phosphorylate BRCA1 and BRCA2 to enable activation of DNA repair [9].
Many reports associate the risk of tumorgenesis with alterations in the HR pathway.[10–12] Aberrations in MDC1 and MRE11 have been strongly linked to breast carcinogenesis [13, 14] and also reported in other cancers.[15–17] Mutations and loss of ATM can contribute to lymphoid malignancies [18] and familial breast and ovarian cancers.[19, 20] Hypomorphic mutations of ATR have been linked to BC and OC development,[21] and mutation and loss of the BRCA1 gene is widely reported to increase the risk of breast and ovarian cancer.[22, 23]
Ovarian cancer (OC) is one of the most common hereditary cancers in women and results in more annual deaths than any other gynecological malignancy.[24] In recent years, reports have emerged suggesting that HR deficiency is strongly linked to the development of OC.[25, 26] 3.7 % deleterious and 4.8 % missense mutations of ATM have been reported in familial OC.[20] ATR mutations have been analyzed in familial breast cancer (BC) and OC, and 23 nucleotide substitution variants discovered.[27] Mutations in BRCA1/2, the two most widely studied genes in the HR pathway, have shown a strong linkage with OC in numerous reports.[28] The majority of the deaths associated with these mutations were from ovarian cancer of the serous histological type (SOC).[29] Moreover, SOC is often detected at an advanced stage, at which time it has already become highly lethal.[30] To better understand the alteration of protein expression in HR pathway, and to provide potential patient selection biomarker for HR inhibitor in SOC, we have analyzed the immunohistochemical expression of MRE11, MDC1, ATM, ATR and BRCA1 in 97 serous ovarian cancers and association with clinicopathological parameters of the patients.
Materials and Methods
Patients and Treatment
The current study was approved by the ethics committee of Chinese PLA General Hospital in Beijing. This study analyzed samples from 97 patients who underwent ovarian cancer complete resection at from Nov. 2005 to Nov. 2009. All 97 patients were diagnosed with serous ovarian cancer (SOC) by 2 qualified pathologists which accounts for more than 40 % of ovarian malignancies. The median patient age was 55 years old (range 35–77). Among the 97 cases, 56.7 % (55/97) of patients had received chemotherapy after surgery and 68.1 % (66/97) of patients had entered into menopause.
Immunohistochemistry (IHC) Study
All tumor samples were collected immediately after surgery, fixed in 10 % buffered formalin and then embedded in paraffin. Four μm-thick tissue sections were cut for IHC study. The slides were baked at 56 ºC for 1 h, followed by de-paraffinized in xylene and rehydrated through a graded series of ethanol concentrations. Antigen retrieval was performed in a pressure cooker for 5 mins using Target Retrieval Solution (Dako). Endogenous peroxidase activity was blocked by Peroxidase Blocking Reagent (Dako) for 5 mins. Primary antibodies (ATM, Epitomics, 1:50; ATR, Santa-cruz Technology, 1:100; BRCA1, Merck, 1:100; MDC1, Sigma, 1:500; MRE11, Abcam, 1:200) were then applied to the specimen for 1 h at room temperature, followed by incubation with labeled polymer-HRP anti rabbit or anti mouse secondary antibody (Dako) for 30 mins at room temperature. Thorough rinsing with TBST was performed after incubation with each reagent. The slides were visualized using DAB substrate-chromagen (Dako) and washed with deionized water before counterstaining with haematoxylin (Sigma). The slides were then dehydrated through a graded series of ethanol concentrations, cleared in xylene and coverslipped in DPX mounting medium. Cases with positive staining of aforementioned 5 biomarkers in previous study were used as positive control. Isotype-matched immunoglobulin fraction instead of primary antibodies used in the experiment served as negative control.
Interpretation of IHC
The intensity of the staining as well as the percentage of positive cells was recorded. Staining intensity was scored from 0, 1+, 2+ to 3+ following the criteria: 0, if absence of staining was observed; 1+, if >10 % of the tumor cells had weak staining; 2+, if >10 % tumor cells had moderate staining; and 3+, if >10 % tumor cells had strong staining. Tumors with 1+, 2+, and 3+ expression were interpreted as positive and tumors with no expression (0 score) were interpreted as negative.
Statistical Analysis
Logistic regression was used to assess the association of homologous recombination deficiency (HRD) expression with clinicopathological parameters and P values were computed from log-likelihood ratio test. P values <0.05 were considered to be statistically significant. Pearson correlation coefficient method was used to assess the correlation of co-expression of HRD pathway genes. The data analysis was performed using R version 2.11.0 on Unix.
Results
Expression of ATM, ATR, MDC1, MRE11 and BRCA1 in SOC
In our SOC samples, negative expression (IHC score ‘0’) of HR pathway proteins was at the following rates; ATM 11.3 % (11/97), ATR 6.3 % (6/96), and BRCA1 29.9 % (29/97). Low expression (IHC score ‘0’ and ‘1+’) of MRE11 was detected in 14.4 % (14/97) and MDC1 in 3.1 % of samples (3/97). (Figs. 1 and 2).

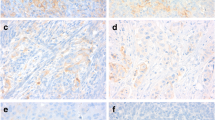
Representative images of ATM IHC staining with scoring 0, 1+, 2+ and 3+ in Chinese SOC. 1a-1d: negative, weak, moderate and strong staining of ATM

Representative images of IHC staining of MRE11, MDC1, ATM, ATR and BRCA1 in Chinese SOC. 2a-2b: Low and strong expression of MRE11; 2c-2d: Low and strong expression of MDC1; 2e-2f: negative and strong staining of ATM; 2G-2H: negative and strong staining of ATR; 2i-2j: negative and strong staining of BRCA1 in different SOC tumors, respectively. (Envision, 40 ×)
Correlation Between Biomarkers Expression and Clinicopathological Parameters
Clinical information of the SOC patients recruited into this study was collected including age, tumor grade, clinical stage, status of menopause and chemotherapy. Statistical analysis of IHC data and clinicopathological parameters are shown in Table 1. Loss of expression of ATM, ATR and BRCA1 and low expression of MDC1 and MRE11 were not associated with age, clinical stage, tumor grade and chemotherapy status. However, loss of ATR expression was significantly correlated with menopause (P = 0.025). Although low or lost expression of these 5 proteins was not strongly associated with clinicopathological parameters, in-depth statistical analysis demonstrated that strong expression (IHC score ‘3+’) of MRE11 (P = 0.016) was significantly associated with low tumor grade (grade I and II). Furthermore, strong expression of ATM (P = 0.017) and MRE11 (P = 0.040), independently, was strongly associated with pre-menopausal SOC, and finally, strong expression of BRCA1 (P = 0.015) was significantly associated with early clinical stage (stage I and II). Detailed analysis results were shown in Table 2.
Overall, 53/96 (55.2 %) SOC cases were identified with negative IHC expression of at least 1 HR pathway protein. Combined biomarker analysis results were shown in Table 3, which indicated that low expression (IHC score 0 and 1+) of MRE11 was strongly associated with loss (IHC score 0) of ATR (P < 0.001) and BRCA1 (P = 0.004). In addition, deficiency of ATR (IHC score 0) was strongly associated with low levels (IHC score 0 and 1+) of ATM (P = 0.028) and loss of expression (IHC score 0) of BRCA1 (P = 0.009). Expression of MDC1 did not show any association with expression of the other 4 HR pathway proteins.
Discussion
Ovarian cancer is the leading cause of death in gynecological malignancies. Although platinum-based chemotherapy regimens are widely used to treat OC patients, these are non-specific treatment modalities which can lead to severe side effects and poor tolerance due to effects on normal tissues. Recently, several reports have shown that deficiencies in the levels of key HR pathway proteins are associated with OC. However, this association has not been fully understood. To explore this further in a Chinese population, we measured the expression of key HR pathway proteins (MRE11, MDC1, ATR, ATM, and BRCA1) and analyzed this data for associations with clinicopathological parameters in 97 Chinese SOC patients.
MRE11 and MDC1 genetic abnormalities can contribute to cancer susceptibility.[15–17] Our data shows that the incidence of low expression of MDC1 and MRE11 was 3.1 % and 14.4 %, respectively. After statistical analysis, we found strong expression of MRE11 (P = 0.016) to be significantly associated with low tumor grade. Conversely, low levels of MRE11 appear to be associated with SOC differentiation. Previous reports have indicated genetic MRE11 abnormalities to be linked to development of various cancers, but protein levels have rarely been studied in SOC. To our knowledge, this study is the first to report an association of MRE11 protein expression with SOC development. Although MDC1 expression failed to show significant correlations with clinical stage and menopause, a strong trend was observed between tumor grade (P = 0.09) and chemotherapy treatment status (P = 0.06), warranting further investigations of this protein as a potential SOC prognostic biomarker.
ATM and ATR are important signaling kinases that activate a complex network of DNA damage response pathways. These coordinate cell cycle checkpoint and DNA repair functions.[8] Deficiencies in ATM and ATR can result in DNA damage sensitivity and cancer predisposition.[10–12] In the current study, loss of expression of ATM and ATR were detected in 11.3 % and 6.3 % of patient samples, respectively. Moreover, statistical analysis showed that loss of expression of ATR (P = 0.025) was significantly correlated with menopause, whilst strong expression of ATM (P = 0.017) correlated with tumors derived from pre-menopausal women.
BRCA1 and BRCA2 protein complexes play critical roles in halting cell cycle progression and enabling DNA repair. Mutations in BRCA1 and BRCA2, which have been reported in various studies, account for 70-80 % and 15 % of families with a history of OC, respectively.[31, 32] Women with a familial history of OC are more likely to develop a deleterious mutation in BRCA1/2.[28] BRCA1 mutations typically confer a higher risk of BC and OC than BRCA2 mutations. Besides OC, deleterious BRCA1 mutations may also increase a woman’s risk of developing cervical, uterine, pancreatic, and colon cancer.[33, 34] A recent study suggested that promoter hypermethylations, somatic mutations, and genomic deletions of BRCA1 might be responsible for the loss or reduced expression of BRCA1 protein.[35] Here, we analyzed BRCA1 protein expression in SOC patients and found a negativity rate of 29.9 % (29/97). Strong expression was significantly correlated with early clinical stage (P = 0.015). Thus, our data indicates that loss of BRCA1 protein is associated with serous ovarian cancer progression.
In the co-expression analysis of all 5 proteins, we found the following associations: 1) low expression of MRE11 with loss of ATR (P < 0.001) and BRCA1 (P = 0.004) and 2) deficiency of ATR with low levels of ATM (P = 0.028) and expression loss of BRCA1 (P = 0.009). These results showed that reduced expression of MRE11 can influence the downstream expression levels of ATR and ATM. Furthermore, the expression level of BRCA1, a direct phosphorylation target of ATM, was significantly lower in patients with ATM loss.
To conclude, by profiling key HR pathway proteins in SOC patients we have demonstrated that protein expression changes of MRE11 and BRCA1 are strongly associated with serous ovarian cancer in late clinical stage, suggesting their potential utility as prognostic tools in the analysis of tumor biopsies or circulating tumor cells. Moreover, as SOC represents an especially lethal cancer with limited therapeutic options, we believe that these association studies further underpin the HR pathway as a novel area of potential therapeutic intervention for SOC.
References
Van Gent DC, Hoeijmakers JHJ, Kanaar R (2001) Chromosomal stability and the DNA double-stranded break connection. Nat Rev Genet 2(3):196–206
Watson JD (1970) Molecular biology of the gene. Molecular biology of the gene (2nd edn)
Haber JE (2000) Partners and pathways: repairing a double-strand break. Trends Genet 16(6):259–264
Shrivastav M, De Haro LP, Nickoloff JA (2007) Regulation of DNA double-strand break repair pathway choice. Cell Res 18(1):134–147
Carson CT, Schwartz RA, Stracker TH, Lilley CE, Lee DV, Weitzman MD (2003) The Mre11 complex is required for ATM activation and the G2/M checkpoint. The EMBO J 22(24):6610–6620
Goldberg M, Stucki M, Falck J, D’Amours D, Rahman D, Pappin D, Bartek J, Jackson SP (2003) MDC1 is required for the intra-S-phase DNA damage checkpoint. Nature 421(6926):952–956
Stewart GS, Wang B, Bignell CR, Taylor AMR, Elledge SJ (2003) MDC1 is a mediator of the mammalian DNA damage checkpoint. Nature 421(6926):961–966
Abraham RT (2001) Cell cycle checkpoint signaling through the ATM and ATR kinases. Genes Dev 15(17):2177–2196
Cortez D, Wang Y, Qin J, Elledge SJ (1999) Requirement of ATM-dependent phosphorylation of brca1 in the DNA damage response to double-strand breaks. Science 286(5442):1162–1166
Helleday T Homologous recombination in cancer development, treatment and development of drug resistance. Carcinogenesis 31 (6):955–960
Kastan MB, Bartek J (2004) Cell-cycle checkpoints and cancer. Nature 432(7015):316–323
McCabe N, Turner NC, Lord CJ, Kluzek K, Bialkowska A, Swift S, Giavara S, O’Connor MJ, Tutt AN, Zdzienicka MZ (2006) Deficiency in the repair of DNA damage by homologous recombination and sensitivity to poly (ADP-ribose) polymerase inhibition. Cancer Res 66(16):8109
Hsu HM, Wang HC, Chen ST, Hsu GC, Shen CY, Yu JC (2007) Breast cancer risk is associated with the genes encoding the DNA double-strand break repair Mre11/Rad50/Nbs1 complex. Cancer Epidemiol Biomark Prev 16(10):2024–2032
Bartkova ZH J (2007) DNA damage response mediators MDC1 and 53BP1: constitutive activation and aberrant loss in breast and lung cancer, but not in testicular germ cell tumours. Oncogene 26(53):7414–7422
Giannini G, Rinaldi C, Ristori E, Ambrosini MI, Cerignoli F, Viel A, Bidoli E, Berni S, D’Amati G, Scambia G (2004) Mutations of an intronic repeat induce impaired MRE11 expression in primary human cancer with microsatellite instability. Oncogene 23(15):2640–2647
Giannini G, Ristori E, Cerignoli F, Rinaldi C, Zani M, Viel A, Ottini L, Crescenzi M, Martinotti S, Bignami M (2002) Human MRE11 is inactivated in mismatch repair-deficient cancers. EMBO Rep 3(3):248–254
Liu ZK, Zhu SC, Wang YX (2007) Expression of DNA damage checkpoint mediator 1 and p53-binding protein 1 in human esophageal cancer cell lines TE-1, TE-13, and Eca109]. Nan fang yi ke da xue xue bao=. J of South Med Univ 27(9):1314
Shiloh Y, Kastan MB (2001) ATM: genome stability, neuronal development, and cancer cross paths. Adv Cancer Res 83:209–254
Chenevix-Trench G, Spurdle AB, Gatei M, Kelly H, Marsh A, Chen X, Donn K, Cummings M, Nyholt D, Jenkins MA (2002) Dominant negative ATM mutations in breast cancer families. J Natl Cancer Inst 94(3):205–215
Thorstenson YR, Roxas A, Kroiss R, Jenkins MA, Yu KM, Bachrich T, Muhr D, Wayne TL, Chu G, Davis RW (2003) Contributions of ATM mutations to familial breast and ovarian cancer. Cancer Res 63(12):3325
Durocher F, Labrie Y, Soucy P, Sinilnikova O, Labuda D, Bessette P, Chiquette J, Laframboise R, Lépine J, Lespérance B (2006) Mutation analysis and characterization of ATR sequence variants in breast cancer cases from high-risk French Canadian breast/ovarian cancer families. BMC Cancer 6(1):230
Geisler JP, Hatterman-Zogg MA, Rathe JA, Buller RE (2002) Frequency of BRCA1 dysfunction in ovarian cancer. J Natl Cancer Inst 94(1):61–67
Welcsh PL, King MC (2001) BRCA1 and BRCA2 and the genetics of breast and ovarian cancer. Hum Mol Genet 10(7):705–713
Jemal A, Siegel R, Ward E, Murray T, Xu J, Smigal C, Thun MJ (2006) Cancer statistics, 2006. CA Cancer J clin 56(2):106–130
Auranen A, Song H, Waterfall C, DiCioccio RA, Kuschel B, Kjaer SK, Hogdall E, Hogdall C, Stratton J, Whittemore AS (2005) Polymorphisms in DNA repair genes and epithelial ovarian cancer risk. Int J Cancer 117(4):611–618
Powell SN, Willers H, Xia F (2002) BRCA2 Keeps Rad51 in Line: High-Fidelity Homologous Recombination Prevents Breast and Ovarian Cancer? Mol Cell 10(6):1262–1263
Heikkinen K, Mansikka V, Karppinen SM, Rapakko K, Winqvist R (2005) Mutation analysis of the ATR gene in breast and ovarian cancer families. Breast Cancer Res 7(4):R495–501
Frank TS, Manley SA, Olopade OI, Cummings S, Garber JE, Bernhardt B, Antman K, Russo D, Wood ME, Mullineau L (1998) Sequence analysis of BRCA1 and BRCA2: correlation of mutations with family history and ovarian cancer risk. J Clin Oncol 16(7):2417–2425
Brown PO, Palmer C (2009) The preclinical natural history of serous ovarian cancer: defining the target for early detection. PLoS Med 6(7):e1000114
Ries LAG, Young JL, Keel GE, Eisner MP, Lin YD, Horner MJ (2007) SEER Survival Monograph: Cancer Survival Among Adults: US SEER Program, 1988–2001, Patient and Tumor Characteristics. National Cancer Institute, SEER Program, NIH Pub (07–6215)
Esteban Cardeñosa E, Bolufer Gilabert P, de Juan Jiménez I, Palanca Suela S, Barragán González E, González Anguix V, Lerma Alejos E, Chirivella González I, Segura Huerta Á, Guillén Ponce C, Relationship of BRCA1 and BRCA2 mutations with cancer burden in the family and tumor incidence. Familial cancer 9 (3):291–295
Lux MP, Fasching PA, Beckmann MW (2006) Hereditary breast and ovarian cancer: review and future perspectives. J Mol Med 84(1):16–28
Kadouri L, Hubert A, Rotenberg Y, Hamburger T, Sagi M, Nechushtan C, Abeliovich D, Peretz T (2007) Cancer risks in carriers of the BRCA1/2 Ashkenazi founder mutations. J Med Genet 44(7):467–471
Thompson D, Easton DF (2002) Cancer incidence in BRCA1 mutation carriers. J Natl Cancer Inst 94(18):1358–1365
W-b L, Ao L, Z-h C, Z-y Z, Y-h Z, X-y Y, Y-l X, Cao J, J-y L (2011) Molecular analysis of DNA repair gene methylation and protein expression during chemical-induced rat lung carcinogenesis. Biochem Biophys Res Commun 408(4):595–601
Acknowledgments
The study was wholly funded by AstraZeneca company group and this submission was approved by the company and Chinese PLA General Hospital in Beijing. The authors disclose no potential conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Qingqing Ye and Li Chen contributed equally to this study.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article

Cite this article
Ye, Q., Chen, L., Yin, X. et al. Development of Serous Ovarian Cancer is Associated with the Expression of Homologous Recombination Pathway Proteins. Pathol. Oncol. Res. 20, 931–938 (2014). https://doi.org/10.1007/s12253-014-9776-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12253-014-9776-8