
Abstract
A discrete-time predator–prey model is investigated in this paper. In considered model, the population is assumed to follow the model suggested by Ricker 1954. Existence and stability of equilibria are studied. Numerical simulations reveal that, depending on the parameters, the system has complicated and rich dynamics and can exhibit complex patterns. Also the bifurcation diagrams are presented.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The predator–prey interaction among the population is well known to be one of the most difficult inquiry areas for the biology and ecology population. Many ecologists, mathematicians, and biologists have studied this during the last few decades [1,2,3,4,5,6]. The differential and difference equations are used to explain the majority of dynamic population models. When populations have non-overlapping generations, the discrete-time models are more reasonable than the continuous-time models [2, 7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23]. Due to its universal existence and importance, a large number of investigations have already examined the predator–prey models (see [24,25,26,27,28,29,30,31,32,33,34,35], for examples). Volterra established a model in 1931 to represent the connection between predators and their preys in an ecological system by developing a system of two autonomous ordinary differential equations [36]. Based on linear per capita growth rates, the Volterra system is the simplest model of predator–prey interactions. The following system of ordinary differential equations represents the famous Volterra prey–predator model.
where X stands for the number of prey and Y stands for the number of predators. Furthermore, the parameters a, b, c, and d are all positive. It has been demonstrated that in the absence of predators, the number of prey increases exponentially, while the number of predators declines exponentially in the absence of a prey population. The terms bXY and dXY describe predator–prey interactions that are beneficial to predators but deadly to prey. A good update for this model is to replace the term aX by the logistic growth \(aX(1-X)\) where population’s per capita growth rate decreases as population size approaches a maximum imposed by limited resources. Stability, bifurcations and chaos control of the improved system studied by [37]. However, we are quite interested in studying the effect of using Ricker map \(r\,X_{n}\,e^{1-\frac{X_{n}}{k}}\) instead of the logistic map to describe the dynamics of preys. In Ricker map, the population growth is virtually exponential; but, as population size grows, the instantaneous growth rate decreases linearly; and finally, population size plateaus and fluctuates around a mean.
The outline of this paper is as follows: The construction of the model is discussed in Sect. 2. Section 3 investigates the existence of equilibria and local stability of the model (5) with various topological types. We explore the existence of bifurcations around equilibria in Sect. 4. Numerical simulations using MATLAB are applied in Sect. 5 to support the theoretical analysis and visualize the newly observed complex dynamics of the system. Section contains the conclusion and discussion.
2 Derivation of the model
This paper deals with the study of stability and bifurcations of a discrete-time predator–prey model. Let \(X_{n}\) and \(Y_{n}\) represent the population density of prey and predator at time n, respectively. We impose the following assumptions to formulate the difference equations which describe the required system.
Assumption 1
In the absence of predator Y, the population of prey X would grow in a classical discrete Ricker population map manner.
where \(r\,>\,0\) is per-capita birth or growth rate in absence of predators r is the prey finite rate of increase in the absence of predators and \(k>0\) is the carrying capacity parameter.
Assumption 2
The population of prey X will drop in the presence of the predator Y. If we suppose that the interaction between the prey and the predator has the mass action principle response due to the prey’s defensive ability, then Eq. (1) can be expressed as
where \(b>0\) is the biomass conversion parameter.
Assumption 3
Prey X are favourite food for predator Y, the population density of predator Y will increase following the mass action principle response manner in the presence of favourite food. Hence we have
where \(d>0\) denotes the predator’s biomass conversion efficiency.
Hence the proposed model is from Eqs. (1)–(3)
with positive initial condition \(X_{0}>0\) and \(Y_{0}>0\). Let \(X_{n}=k\,x_{n}\) and \(Y_{n}=\frac{y_{n}}{b}\). We rewrite model Eq. (4) as
where \(\alpha = d\,k\,>0.\)
3 Fixed points: existence and stability
In this section, we study the existence and stability of fixed points of the system (5) in \({\mathbb {R}}_{+}^{2}\). Results about the existence of fixed points are summarized as follows:
Proposition 1
For model (5), we can have at most three fixed points:
-
(i)
The trivial fixed point \(E_{0}(0, 0)\) always exists;
-
(ii)
The predator-free fixed point \(E_{1}\left( 1+\ln {(r)},0\right) \) exists if \( r\,>\,\mathrm {e}^{-1}\);
-
(iii)
The interior fixed \(E_{2}\left( \frac{1}{\alpha },r\,\mathrm {e}^{\frac{\alpha -1}{\alpha }}-1\right) \) exists if \(r\,>\, \mathrm {e}^{\frac{1-\alpha }{\alpha }}\).
Proof
The fixed points correspond to the steady states of the model population (5) can be obtained by solving the algebraic system
then all non-negative fixed points of the system (5) are \(E_{0}(0, 0),\) \(E_{1}\left( 1+\ln {(r)},0\right) , \) which is positive if \( r\,>\,\mathrm {e}^{-1}\) and \(E_{2}\left( \frac{1}{\alpha },r\,\mathrm {e}^{\frac{\alpha -1}{\alpha }}-1\right) ,\) which is positive if \(r\,>\,\mathrm {e}^{\frac{1-\alpha }{\alpha }}\). \(\square \)
The Jacobian matrix J(x, y) of the model (5) about the fixed point (x, y) is given by
Its characteristic equation is
The biological interruption for three fixed points is as follows: The fixed point \(E_{0}\) represents a situation in which there is no prey and no predators. The fixed point \(E_{1}\) reflects the case where there is prey but no predators. \(E_{2}\) refers to the coexistence of fixed nonzero numbers of prey and predators.
3.1 Dynamic behavior of the model
Now we will study the behavior of the solutions of (5) about \( E_{0} \), \(E_{1}\) and \(E_{2}\). To do this we recall the following definitions:
Definition 1
Let E(x, y) be a fixed point of (5) and let \(\mu _1\) and \(\mu _2\) are the eigenvalues of (7).
-
(i)
E is called a sink (locally asymptotic stable) if \(\mid \mu _1\mid < 1\) and \(\mid \mu _2\mid < 1\);
-
(ii)
E is called a source if \(\mid \mu _1\mid > 1\) and \(\mid \mu _2\mid > 1\). A source is locally unstable;
-
(iii)
E is called a saddle if \(\mid \mu _1\mid < 1\) and \(\mid \mu _2\mid > 1\) (or \(\mid \mu _1\mid > 1\) and \(\mid \mu _2\mid < 1\));
-
(iv)
E is called non-hyperbolic if either \(\mid \mu _1\mid =1\) and \(\mid \mu _2\mid = 1\).
The relations between roots and coefficients of the quadratic equation is given by the following Lemma.
Lemma 1
Let \(R(\mu ) = \mu ^2 + A\mu + B\). Suppose that \(R(1) > 0\), \(\mu _1\) and \(\mu _2\) are the roots of \(R(\mu ) = 0\). Then
-
(i)
\(\mid \mu _1\mid < 1\) and \(\mid \mu _2\mid < 1\) if and only if \(R(-1) > 0\) and \(B < 1\);
-
(ii)
\(\mid \mu _1\mid < 1\) and \(\mid \mu _2\mid > 1\) (or \(\mid \mu _1\mid > 1\) and \(\mid \mu _2\mid < 1\)) if and only if \(R(-1) < 0\);
-
(iii)
\(\mid \mu _1\mid > 1\) and \(\mid \mu _2\mid > 1\) if and only if \(R(-1) > 0\) and \(B > 1\);
-
(iv)
\(\mu _1 =-1\) and \(\mid \mu _2\mid \ne 1\) if and only if \(R(-1) = 0\) and \(B \ne 0,2\);
-
(v)
\(\mu _1\) and \(\mu _2\) are complex and \(\mu _1 =\mid \mu _2\mid = 1\) if and only if \(A^2-4B < 0\) and \(B = 1\). \(\square \)
The following Proposition confirms the stability of fixed points of system (5) under some conditions.
Proposition 2
For the fixed point \(E_{0}\) of the system (5), the following statements are true:
-
(i)
\(E_{0}\) is sink if \(0\,<\,r\,<\,{\mathrm {e}^{-1}}\);
-
(ii)
\(E_{0}\) is never a source;
-
(iii)
\(E_{0}\) is an unstable saddle if \(r\,>\,{\mathrm {e}^{-1}}\);
-
(iv)
\(E_{0}\) is non-hyperbolic if \(r\,=\,\mathrm {e}^{-1}\).
Proof
The Jacobian matrix associated with (5) at \(E_{0}\) is \(J\left( E_{0}\right) \) and is given by
the eigenvalues of \(J\left( E_{0}\right) \) are \(\lambda _{1}=0\) and \(\lambda _{2}=r\,\mathrm {e}\). So, the fixed point
-
(i)
\(E_{0}\) is a sink if and only if \(0\,<\,r\,<\,\mathrm {e}^{-1}\);
-
(ii)
\(E_{0}\) is never a source since one of the eigenvalues is always zero;
-
(iii)
\(E_{0}\) is a saddle if and only if \(r\,>\,\mathrm {e}^{-1}\);
-
(iv)
\(E_{0}\) is non-hyperbolic if only if \(r\,=\,\mathrm {e}^{-1}\). \(\square \)
Proposition 3
The fixed point \(E_{0}\) of the system (5) is globally asymptotically stable if \(0\,<\,r\,<\,{\mathrm {e}^{-1}}\).
Proof
It was shown in Proposition 2 that \(E_{0}\) is locally asymptotically stable if \(0\,<\,r\,<\,{\mathrm {e}^{-1}}\). So, proving that \(\lim _{n\rightarrow \infty } x_{n}=0\) and \(\lim _{n\rightarrow \infty } y_{n}=0\) is sufficient to prove the global stability of \(E_{0}\).
It follows from the first equation of system (5) that
Then
Then
Note that
Since \(x < \mathrm {e}^{x}\) for all \(x \ge 0\), then \(x_{n}y_{n}< y_{n} \mathrm {e}^{x_{n}} < r \mathrm {e}\). Then it follows from second equation of system (5) that \( y_{n+1} \le \alpha r \mathrm {e}\). That is \(y_{n}\) is bounded from above. Again we see from the second equation of system (5) and Eq. (8) that
Thus
Since \(y_{n}\) is bounded and \(r \mathrm {e}<1\) therefore
Then the proof is completed. \(\square \)
Proposition 4
For the fixed point \(E_{1}\) of the system (5), the following statements are true:
-
(i)
\(E_{1}\) is sink if \(\mathrm {e}^{-1}\,<\,r\,<\,\min \{\mathrm {e},\mathrm {e}^{\frac{1-\alpha }{\alpha }}\}\);
-
(ii)
\(E_{1}\) is a source if \(r\,>\,\max \{\mathrm {e},\mathrm {e}^{\frac{1-\alpha }{\alpha }}\}\);
-
(iii)
\(E_{1}\) is an unstable saddle point if \(\min \{\mathrm {e},\mathrm {e}^{\frac{1-\alpha }{\alpha }}\}\,<\,r\,<\,\max \{\mathrm {e},\mathrm {e}^{\frac{1-\alpha }{\alpha }}\}\);
-
(iv)
\(E_{1}\) is non-hyperbolic if \(r\in \{\mathrm {e},\mathrm {e}^{-\frac{\alpha -1}{\alpha }}\}\).
Proof
In the same manner as in Proposition (2). The Jacobian matrix associated with (5) at \(E_{1}\) is \(J\left( E_{1}\right) \) and is given by
hence, the eigenvalues of \(J\left( E_{1}\right) \) are \(\lambda _{1}=-\ln (r)\) and \(\lambda _{2}=\alpha \,(1+\ln (r))\). Then, by using Lemma (1) we get the results of Proposition (4). \(\square \)
We are interested to analyze the dynamical behavior of the system around interior fixed point \(E_{2}\) for its importance. In order to determine conditions that guarantee that the characteristic roots of Jacobian matrix about \(E_{2}\) are inside the unit disk one can use Lemma (1).
Proposition 5
The positive equilibrium point \(E_{2}\) of the model (5) is sink if and only if the following conditions hold:
-
(i)
\(\mathrm {e}^{\frac{1-\alpha }{\alpha }}\,<\,r<\,\frac{3\,\alpha }{2-\alpha }\,\mathrm {e}^{\frac{1-\alpha }{\alpha }}\) and \(\frac{1}{2}< \alpha \,\le \frac{5}{4}\);
-
(ii)
\(\mathrm {e}^{\frac{1-\alpha }{\alpha }}\,<\,r<\,\frac{\alpha }{\alpha -1}\,\mathrm {e}^{\frac{1-\alpha }{\alpha }}\) and \( \alpha \,>\,\frac{5}{4}\);
Proof
The Jacobian matrix (6) at \(E_{2}\) reads as
and the characteristic equation of \(J(E_{2})\) is
where
according to Lemma (1), the positive equilibrium point \(E_{2}\) is locally asymptotically stable if
-
(i)
\(1+P+Q>0\) and this means \(r>\mathrm {e}^{\frac{1-\alpha }{\alpha }}\);
-
(ii)
\(1-P+Q>0\) and this means \(r\,(2-\alpha )\mathrm {e}^{\frac{\alpha -1}{\alpha }}<3\alpha \); and
-
(iii)
\(Q<1\) and this means \(r(\alpha -1)\mathrm {e}^{\frac{\alpha -1}{\alpha }}<\alpha \),
or, equivalently
-
(i)
\(\mathrm {e}^{\frac{1-\alpha }{\alpha }}\,<\,r<\,\frac{3\,\alpha }{2-\alpha }\,\mathrm {e}^{\frac{1-\alpha }{\alpha }}\) and \(\frac{1}{2}< \alpha \,\le \frac{5}{4}\);
-
(ii)
\(\mathrm {e}^{\frac{1-\alpha }{\alpha }}\,<\,r<\,\frac{\alpha }{\alpha -1}\,\mathrm {e}^{\frac{1-\alpha }{\alpha }}\) and \( \alpha \,>\,\frac{5}{4}\).\(\square \)
The nonhyperbolic fixed point case is more involved. According to the eigenvalues of 1 or \(-\,1\), there are several scenarios. It is common to use centre manifold theory to determine the stability of the fixed point when one of the eigenvalues is on the unit circle and the other eigenvalue is inside the unit circle [38,39,40]. In order to determine conditions that guarantee that the characteristic roots in the unit circle, one can use Theorem 4.5 (Elaydi [40], p. 203). \(\square \)
Proposition 6
\(E_{2}\) loses stability:
-
(i)
via a flip point when \(1 / 2<\alpha <5 / 4\) and \(r=\frac{3 \alpha }{ 2-\alpha }e^{\frac{1-\alpha }{\alpha }}\);
-
(ii)
via a Neimark–Sacker point when \(5/ 4<\alpha \) and \(r=\frac{ \alpha }{ \alpha -1}e^{\frac{1-\alpha }{\alpha }}\);
-
(iii)
via a branch point when \(\alpha >1 / 2\) and \(r=e^{\frac{1-\alpha }{\alpha }}\).
Proof
By Proposition (4) the stability boundary of \(E_{2}\) consists of parts of three curves, namely,
-
(1)
Curve 1: \(r=\frac{3 \alpha }{2-\alpha }e^{\frac{1-\alpha }{\alpha }}\),
-
(2)
Curve 2:\(r=\frac{ \alpha }{\alpha -1}e^{\frac{1-\alpha }{\alpha }}\),
-
(3)
Curve 3: \(r=e^{\frac{1-\alpha }{\alpha }}\), the points of Curve 1 on the stability boundary of \(E_{2}\) meet \(1+p+q=0\), implying that they have an eigenvalue of \(-1\) and are hence period doubling points. The points on the stability boundary of Curve 2 satisfy \(q=1\), i.e. they have two eigenvalues with product 1 and are hence Neimark–Sacker points. Curve 3 points on the stability boundary satisfy \(1-p+q=0\), implying that they have an eigenvalue of 1. It is simple to verify that \(E_{2}\) then branches to \(E_{1}\). When this is combined with Proposition (4), the interior points of the boundary parts of Curves 1,2, and 3 form the sets specified in Proposition (4) parts (i), (ii), and (iii) respectively. \(\square \)

Stability regions in (\(\alpha ,r\))-space
4 Bifurcations analysis
The existence of bifurcations about equilibria is investigated in this section, which is based on theoretical studies in Sect. 3. It is well known that the classification of bifurcation types depends mainly on the natural of the eigenvalues about the equilibrium points. The following is a summary of the presence of related bifurcations around the equilibria \(E_{0}\), \(E_{1}\), and \(E_{2}\):
-
(i)
From Proposition (2), we can find out that when \(r =\mathrm {e}^{-1}\), one of the eigenvalues about the equilibrium \(E_{0}\) is 1. So when the parameter varies in the small neighborhood of \(r =\mathrm {e}^{-1}\), a fold bifurcation may occur.
-
(ii)
From Proposition (2), we can see that if \(r = \mathrm {e}^{\frac{1-\alpha }{ \alpha }}\) holds then one of the eigenvalues about \(E_{1}\) is 1 . So a fold bifurcation may occurs when the parameter varies in a small neighborhood of \( r = \mathrm {e}^{\frac{1-\alpha }{\alpha }}\). And we denote the parameters satisfying \(r = \mathrm {e}^{\frac{1-\alpha }{\alpha }}\) as
$$\begin{aligned} F_{E_{1}}=\left\{ \left( \alpha , r\right) : r = \mathrm {e}^{\frac{1-\alpha }{ \alpha }}, \alpha , r>0\right\} . \end{aligned}$$ -
(iii)
If (i) holds in Proposition (6), we can observe that one of the eigenvalues of \(J_{E_{2}}\) about \(E_{2}\) is \(-1\), while the other is neither 1 nor \(-1\) . As a result, a period-doubling bifurcation exists due to parameter variation in a small neighborhood of \(r=\frac{3\alpha }{2-\alpha }\mathrm {e}^{ \frac{1-\alpha }{\alpha }}\). More precisely we can also represent the parameters satisfying \(r=\frac{3\alpha }{2-\alpha }\mathrm {e}^{\frac{1-\alpha }{ \alpha }}\) as
$$\begin{aligned} P_{E_{2}}=\left\{ \left( \alpha , r\right) : r=\frac{3\alpha }{2-\alpha }\mathrm {e} ^{\frac{1-\alpha }{\alpha }}\quad \text {and} \quad \frac{1}{2}<\alpha <\frac{5}{ 4} \right\} . \end{aligned}$$ -
(iv)
If (ii) holds in Proposition (6), we see that the eigenvalues of \(J_{E_{2}}\) about \(E_{2}\) are a pair of complex conjugate with modulus 1 . So a Neimark–Sacker bifurcation exists by the variation of parameter in a small neighborhood of \(r=\frac{\alpha }{\alpha -1}\mathrm {e}^{\frac{1-\alpha }{\alpha } } \). Precisely we represent the parameters satisfying \(r=\frac{\alpha }{ \alpha -1}\mathrm {e}^{\frac{1-\alpha }{\alpha }} \) as
The stability regions (i), (ii) and (iii) of \(E_{2}\) obtained in Proposition 3 are depicted as \(\Omega _{E 2}^{S}\) in Fig. 1. \(\Omega _{E 0}^{S}\) and \(\Omega _{E 1}^{S}\) in Fig. 1 represent the stability regions of \(E_{0}\) and \(E_{1}\), respectively.The following is the biological interpretation of Fig. 1. For all parameter values, the situation with no prey and no predators exists, but it is only stable in \(\Omega _{E 0}^{S}\), that is, for \(r<\frac{1}{e}\). For every \(r>\frac{1}{e}\), the situation with a fixed number of prey but no predators exists, but only in \(\Omega _{E 2}^{S}\) is stable. When \(r>\frac{1}{ e}\) and \(\alpha >\frac{1}{1+\ln (r)}\), coexistence of fixed nonzero numbers of prey and predators is possible, but only in \(\Omega _{E 2}^{S}\) is stable.
4.1 Neimark–Sacker bifurcation about \(E_{2}\)
We first discuss the Neimark–Sacker bifurcation of the model (5) about \(E_{2}\). Consider the parameter r in a neighborhood of \(r^{*}\), i.e., \(r=r^{*}+\epsilon \) where \(\epsilon \ll 1\), then the model (5) becomes
the characteristic equation of \(J_{E_{2}(\frac{1}{\alpha },(r^{*}+\epsilon ) \mathrm {e}^{\frac{\alpha -1}{\alpha }}-1)}\) about \(E_{2}(\frac{1}{\alpha } ,(r^{*}+\epsilon )\mathrm {e}^{\frac{\alpha -1}{\alpha }}-1)\) of the model (9) is
where
the roots of characteristic equation of \(J_{E_{2}(\frac{1}{\alpha } ,(r^{*}+\epsilon )\mathrm {e}^{\frac{\alpha -1}{\alpha }}-1)}\) about \(E_{2}( \frac{1}{\alpha },(r^{*}+\epsilon )\mathrm {e}^{\frac{\alpha -1}{\alpha }}-1)\) are
and
additionally, we required that when \(\epsilon =0, \lambda _{1,2}^{m} \ne 1, m=1,2,3,4\), which corresponds to \(p(0) \ne -2,0,1,2\), which is true by computation.Let \(u_{n}=x_{n}-x^{*}, v_{n}=y_{n}-y^{*}\) then the equilibrium \(E_{2}\) of the model (5) transforms into O(0, 0).By manipulation, one gets
where \(x^{*}=\frac{1}{\alpha }\), \( y^{*}=r\mathrm {e}^{\frac{\alpha -1}{\alpha }}-1\). Hereafter when \(\epsilon =0\), the normal form of system (10) is studied. Expanding (10) up to third order about \(\left( u_{n}, v_{n}\right) =(0,0)\) by Taylor series, we get
where
Now, let
and the invertible matrix T defined by
Using the following translation:
(10) gives
where
and
In addition,
and
In order for (11) to undergo a Neimark–Sacker bifurcation, it is mandatory that the following discriminatory quantity, i.e., \(\chi \ne 0\) (see [38, 40, 41]),
where
after calculating, we get
Based on this analysis and the Neimark–Sacker bifurcation Proposition discussed in [38, 42,43,44,45], we arrive at the following Proposition.
Proposition 6
If \(\chi \ne 0\) then the model (5) undergoes a Neimark–Sacker bifurcation about \(E_{2}\) as \(\left( r, \alpha \right) \) go through \(N_{E_{2}} .\) Additionally, an attracting (resp. repelling) closed curve bifurcates from \(E_{2}\) if \(\chi <0(\) resp. \(\chi >0)\).
Remark
According to bifurcation theory discussed in [43], the bifurcation is called a supercritical Neimark–Sacker bifurcation if the discriminatory quantity \(\chi <0 .\) In the following section, numerical simulations guarantee that a supercritical Neimark–Sacker bifurcation occurs for the model (5).
4.2 Period-doubling bifurcation
Here we study the period-doubling bifurcation of model (5) at \(E_{2}\) when parameters vary in a small neighborhood of \( P_{E_{2}}\). Select arbitrary parameters \(\left( r^{*},\alpha \right) \) from \( P_{E_{2}}\). We consider the parameter \(r^{*}\) as a new dependent variable, and we can get
let \(u_{n}=x_{n}-x^{*}\), \(v_{n}=y_{n}-y^{*}\) then the equilibrium \(E_{2}\) of the discrete-time model (13) transforms into O(0, 0). By calculating we get
where
Now, construct an invertible matrix T
and use the translation
(14) gives
where
hereafter we determine the center manifold \(W^{c}(0,0)\) of (15) about (0, 0) in a small neighborhood of \(r^{*}\) [6, 38, 43, 46]. By center manifold theorem, there exists a center manifold \(W^{c}(0,0)\) that can be represented as follows:
where \(O\left( \left( \left| X_{n}\right| +\left| r^{*}\right| \right) ^{3}\right) \) is a function with order at least three in their variables \(\left( X_{n}, r^{*}\right) \), and
Therefore, we consider the map (15) restricted to \(W^{c}(0,0)\) as follows:
where
in order for the map (16) to undergo a period-doubling bifurcation, we require that the following discriminatory quantities are non-zero:
after calculating we obtain
and
from the above analysis in [37] and theorem in [38, 42,43,44,45] ,we have the following Proposition
Proposition 7
If \(\Lambda _{2} \ne 0\), the map (13) undergoes a period-doubling bifurcation about the unique positive equilibrium \(E_{2}\) when \(r^{*}\) varies in a small neighborhood of O(0, 0) . Moreover, if \( \Lambda _{2}>0\left( \right. \) resp \(\left. . \Lambda _{2}<0\right) \), then the period-2 points that bifurcate from \(E_{2}\) are stable (resp. unstable).
5 Numerical simulations
We use numerical simulations to create bifurcation diagrams and the phase portraits of model (5) to support the previous theoretical analysis and highlight new complicated dynamical behaviors. We will use the MATCONTM package [43, 47] to carry out the numerical continuation of equilibria in order to obtain bifurcation curves of system (5) for different sets of parameters and locate points of interest. The bifurcation parameters are considered in the following four cases.

Phase portrait of equilibrium point \(E_{0}\) and its bifurcating to \(E_{1}\)
Case 1 Choosing \(r=0.2, \alpha =1\), with initial value \(\left( x_{0}, y_{0}\right) =(0.1,0.1)\), We see that model (5) has stable equilibrium point \(E_{0}(0,0)\), there are no prey or predator in the area. Figure 2 shows the correctness of discussion about equilibrium \(E_{0}=(0,0)\) in Sect. 3. From Fig. 2d we see that equilibrium \(E_{0}=(0,0)\) loses its stability and bifurcates to \(E_{1}\) when \(r = \mathrm {e}^{-1}\). The MATCONTM report is
Case 2 Choosing \(r=1, \alpha =0.4\), with initial value \(\left( x_{0}, y_{0}\right) =(0.5,0.2)\), model (5) has stable equilibrium point \(E_{1}(1,0)\). It can be observed from Fig. 3a that the prey density x is fixed at 1 but the predator population y quickly reduces to zero. In stability region of \(E_{1}\) with r free, we can see that \(E_{1}\) is stable when \(\mathrm {e}^{-1}<r<\mathrm {e}\). It loses its stability via a supercritical period doubling point (PD) when \(r=\mathrm {e}\), and via a branch point when \(r=\mathrm {e}^{-1}\) (see Fig. 3b). Figure 3 demonstrates the correctness of discussion about equilibrium \(E_{1}\left( 1+\ln {r},0\right) \) in Sect. 3. The MATCONTM report is

Phase portraits of equilibrium point \(E_{1}\) and its bifurcation curve
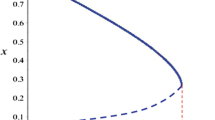
Case 3 Varying \(\alpha \) in range \(1.2 \le \alpha \le 1.54\) and fixing \( r=2.1\). When \(1.2 \le \alpha < 1.513333\) model (5) has a unique positive stable equilibrium point. By calculating, we see that this unique equilibrium point is (0.7142857143, 1.794495614) for \(\left( \alpha , r\right) =(1.4,2.1)\). The corresponding phase portraits for various values of \(\alpha \) in this range are shown in Fig. 4. Neimark–Sacker bifurcation occurs at \(\alpha =1.513333\), as shown in Fig. 5f, with \(\sqrt{\frac{\alpha -1}{4 \alpha r}}e^{(\frac{\alpha -1}{2\alpha } )}=0.2380951456>0\) hold. Moreover, for \(\alpha =1.513333\) the eigenvalues of \( J_{E_{2}}\) about \(E_{2}\) are
after some calculations from Maple one gets
in view of (17) and (18) the value of the discriminatory quantity is \(\chi =-0.4373638855<0\). Therefore if \(\alpha =1.52>1.51333\) the model (5) undergoes a supercritical Neimark–Sacker bifurcation and a stable invariant close curve appears, which is presented in Fig. 5. The MATCONTM report is

Phase portraits for various values of \(\alpha \) from 1.2 to 1.51

Phase portraits for various values of \(\alpha \) from 1.5142 to 1.54

Phase portraits for various values of r from 2.3 to 3.89
The numerical solution in Figs. 4 and 5 reveals that the stable fixed point \(E_{2}\) becomes unstable, allowing preys and predators to coexist by a persistent positive periodic oscillation as time passes.Case 4 Varying r in range \(2.3 \le r \le 3.89\) and fixing \(\alpha =0.63\). Figure 6 shows that prey and predator densities are fixed at the stable equilibrium point \(E_{2}\) for \(r<2.482007420\), and \(E_{2}\) loses its stability when \(r=2.482007420\). Further, when \(r>2.482007420\), the numbers of prey and predator oscillate in a stable period-2 orbit then period 4-orbit and so on, that is a period-doubling bifurcation. Moreover, a chaotic set is emerged with the increasing of r.
6 Conclusion
The dynamical behaviors of model (5) is discussed in this study. Model (5) contains two boundary equilibria, \(E_{0}, E_{1}\), and a positive equilibria \(E_{2}\) when \(r>max\{\mathrm {e}^{-1}, \mathrm {e}^{\frac{1-\alpha }{\alpha }}\}\). We explored local stability along topological types about \(E_{0}\), \(E_{1}\) and \( E_{2}\) by using the linearization method. From the discussion in Sect. 4.2, we know there is period-doubling bifurcation and chaos near equilibria \(E_{2}\) as the parameter r varies in the small neighbourhood of \(r=2.5\). Moreover, if \( \alpha \) varies in the small neighborhood when \((a, r) \in N_{E_{2}}\), there exists Neimark–Sacker bifurcation.Numerical simulations have shown not only the correctness of the theoretical results but also the complexity of the behavior of the model (5) such as the period-2, -4, and -8 orbits. Comparing with the discrete-time model (4) in [37] where the author used the logistic growth \(rX_{n}(1-X_{n})\) in the first equation of the model. This logistic map has the unrealistic fact that the parabola drops below the horizontal axis as we move off to the right. A more reasonable choice would be the Ricker map \(r X_{n}\left( e^{1-X_{n}}\right) \) which produces very small, but still positive, values as \(X_{n}\) becomes very large [2]. The global qualitative analysis of model(5) has not been obtained yet, we will leave it for our future work.
Data availability
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
References
Chapman, R.: The struggle for existence. Ecology 16(4), 656–657 (1935)
Allman, E.S., Rhodes, J.A.: Mathematical Models in Biology: An Introduction. Cambridge University Press, Cambridge (2004)
Edelstein-Keshet, L.: Mathematical Models in Biology. SIAM, Philadelphia (2005)
Hoppensteadt, F.C.: Mathematical Methods of Population Biology. Cambridge University Press, Cambridge (1982)
Fulford, G., Forrester, P., Forrester, P.J., Jones, A.: Modelling with Differential and Difference Equations, vol. 10. Cambridge University Press, Cambridge (1997)
Zhang, W.-B.: Discrete Dynamical Systems. Bifurcations and Chaos in Economics. Elsevier, Amsterdam (2006)
Murray, J.D.: Mathematical Biology: I. An Introduction. Interdisciplinary Applied Mathematics. Springer, Berlin (2002)
Zeb, A., Alzahrani, E., Erturk, V.S., Zaman, G.: Mathematical model for coronavirus disease (Covid-19) containing isolation class. Biomed. Res. Int. 2020, 2020 (2019)
Zhang, Z., Zeb, A., Hussain, S., Alzahrani, E.: Dynamics of Covid-19 mathematical model with stochastic perturbation. Adv. Differ. Equ. 2020(1), 1–12 (2020)
Bushnaq, S., Saeed, T., Torres, D.F., Zeb, A.: Control of Covid-19 dynamics through a fractional-order model. Alex. Eng. J. 60(4), 3587–3592 (2021)
Nazir, G., Zeb, A., Shah, K., Saeed, T., Khan, R.A., Khan, S.I.U.: Study of Covid-19 mathematical model of fractional order via modified Euler method. Alex. Eng. J. 60(6), 5287–5296 (2021)
Zhang, Z., Gul, R., Zeb, A.: Global sensitivity analysis of Covid-19 mathematical model. Alex. Eng. J. 60(1), 565–572 (2021)
Zhang, Z., Zeb, A., Alzahrani, E., Iqbal, S.: Crowding effects on the dynamics of Covid-19 mathematical model. Adv. Differ. Equ. 2020(1), 1–13 (2020)
Mezouaghi, A., Djillali, S., Zeb, A., Nisar, K.S.: Global proprieties of a delayed epidemic model with partial susceptible protection. Math. Biosci. Eng. 19(1), 209–224 (2022)
Souna, F., Lakmeche, A., Djilali, S.: Spatiotemporal patterns in a diffusive predator–prey model with protection zone and predator harvesting. Chaos Solitons Fractals 140, 110180 (2020)
Bentout, S., Djilali, S., Ghanbari, B.: Backward, hopf bifurcation in a heroin epidemic model with treat age. Int. J. Model. Simul. Sci. Comput. 12(02), 2150018 (2021)
Sitthiwirattham, T., Zeb, A., Chasreechai, S., Eskandari, Z., Tilioua, M., Djilali, S.: Analysis of a discrete mathematical Covid-19 model. Results Phys. 28, 104668 (2021)
Soufiane, B., Touaoula, T.M.: Global analysis of an infection age model with a class of nonlinear incidence rates. J. Math. Anal. Appl. 434(2), 1211–1239 (2016)
Bentout, S., Tridane, A., Djilali, S., Touaoula, T.: Age-structured modeling of Covid-19 epidemic in the USA, UAE and Algeria. Alex. Eng. J. 60, 401–411 (2021)
Bentout, S., Chekroun, A., Kuniya, T.: Parameter estimation and prediction for coronavirus disease outbreak 2019 (Covid-19) in Algeria. AIMS Public Health 7(2), 306 (2020)
Bentout, S., Chen, Y., Djilali, S.: Global dynamics of an Seir model with two age structures and a nonlinear incidence. Acta Appl. Math. 171(1), 1–27 (2021)
Mezouaghi, A., Djilali, S., Bentout, S., Biroud, K.: Bifurcation analysis of a diffusive predator–prey model with prey social behavior and predator harvesting. Math. Methods Appl. Sci. 45(2), 718–731 (2022)
Djilali, S., Bentout, S.: Pattern formations of a delayed diffusive predator–prey model with predator harvesting and prey social behavior. Math. Methods Appl. Sci. 44(11), 9128–9142 (2021)
Agiza, H., ELabbasy, E., EL-Metwally, H., Elsadany, A.: Chaotic dynamics of a discrete prey–predator model with Holling type II. Nonlinear Anal. Real World Appl. 10(1), 116–129 (2009)
Din, Q.: Complexity and chaos control in a discrete-time prey–predator model. Commun. Nonlinear Sci. Numer. Simul. 49, 113–134 (2017)
Elsadany, A., El-Metwally, H., Elabbasy, E., Agiza, H.: Chaos and bifurcation of a nonlinear discrete prey–predator system. Comput. Ecol. Softw. 2(3), 169 (2012)
Freedman, H.: A model of predator–prey dynamics as modified by the action of a parasite. Math. Biosci. 99(2), 143–155 (1990)
Zhao, J., Yan, Y.: Stability and bifurcation analysis of a discrete predator–prey system with modified Holling–Tanner functional response. Adv. Differ. Equ. 2018(1), 1–18 (2018)
Fang, Q., Li, X.: Complex dynamics of a discrete predator–prey system with a strong Allee effect on the prey and a ratio-dependent functional response. Adv. Differ. Equ. 2018(1), 1–16 (2018)
Kangalgil, F., Kartal, S.: Stability and bifurcation analysis in a host-parasitoid model with Hassell growth function. Adv. Differ. Equ. 2018(1), 1–15 (2018)
Berryman, A.A.: The orgins and evolution of predator–prey theory. Ecology 73(5), 1530–1535 (1992)
Smith, J.M., Slatkin, M.: The stability of predator–prey systems. Ecology 54(2), 384–391 (1973)
Hamada, M.Y., El-Azab, T., El-Metwally, H.: Dynamics of a Ricker type sir discrete time system. J. Appl. Anal. Comput. (2022) (to appear)
Hamada, M.Y., El-Azab, T., El-Metwally, H.: Bifurcations and chaos analysis of a two-dimensional discrete-time predator–prey model. Int. J. Dyn. Control (2022) (to appear)
Hamada, M.Y., El-Azab, T., El-Metwally, H.: Bifurcations analysis of a two-dimensional discrete-time predator–prey model. Math. Methods Appl. Sci. (2022) (to appear)
Volterra, V.: Théorie mathématique de la lutte pour la vie. Gauthiers-Villars, Paris (1931)
Khan, A.Q.: Bifurcations of a two-dimensional discrete-time predator–prey model. Adv. Differ. Equ. 2019(1), 1–23 (2019)
Kuznetsov, Y.A.: Elements of Applied Bifurcation Theory, vol. 112. Springer, New York (2004)
Wiggins, S., Golubitsky, M.: Introduction to Applied Nonlinear Dynamical Systems and Chaos, vol. 2. Springer, Berlin (1990)
Elaydi, S.N.: Discrete Chaos: With Applications in Science and Engineering. Chapman and Hall/CRC, Boca Raton (2007)
Liu, X., Xiao, D.: Complex dynamic behaviors of a discrete-time predator–prey system. Chaos Solitons Fractals 32(1), 80–94 (2007)
Guckenheimer, J., Holmes, P.: Nonlinear Oscillations, Dynamical Systems, and Bifurcations of Vector Fields, vol. 42. Springer, Berlin (2013)
Kuznetsov, Y.A., Meijer, H.G.E.: Numerical Bifurcation Analysis of Maps: From Theory to Software. Cambridge University Press, Cambridge (2019)
Iooss, G.: Bifurcation of Maps and Applications. Elsevier, Amsterdam (1979)
Crawford, J.D.: Introduction to bifurcation theory. Rev. Mod. Phys. 63(4), 991 (1991)
Carr, J.: Applications of Centre Manifold Theory, vol. 35. Springer, Berlin (2012)
Govaerts, W., Kuznetsov, Y.A., Ghaziani, R.K., Meijer, H.: Cl MatContM: A Toolbox for Continuation and Bifurcation of Cycles of Maps. Universiteit Gent, Gent (2008)
Acknowledgements
We would like to thank the editor and the anonymous referees for their valuable comments and helpful suggestions, which have led to a great improvement of the initial version.
Funding
Open access funding provided by The Science, Technology &; Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). The authors declare that they got no funding on any part of this research.
Author information
Authors and Affiliations
Contributions
All authors contributed equally in this article. They read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest regarding the publication of this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hamada, M.Y., El-Azab, T. & El-Metwally, H. Bifurcations and dynamics of a discrete predator–prey model of ricker type. J. Appl. Math. Comput. 69, 113–135 (2023). https://doi.org/10.1007/s12190-022-01737-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12190-022-01737-8