
Abstract
Iron deficiency (ID) is the most common gestational micronutrient deficiency in the world, targets the fetal hippocampus and striatum and results in long-term behavioral abnormalities. These structures primarily mediate spatial and procedural memory, respectively, in the rodent but have interconnections that result in competition or cooperation during cognitive tasks. We determined whether ID-induced impairment of one alters the function of the other by genetically inducing a 40% reduction of hippocampus iron content in late fetal life in mice and measuring dorsal striatal gene expression and metabolism and the behavioral balance between the two memory systems in adulthood. Slc11a2hipp/hipp mice had similar striatum iron content, but 18% lower glucose and 44% lower lactate levels, a 30% higher phosphocreatine:creatine ratio, and reduced iron transporter gene expression compared to wild type (WT) littermates, implying reduced striatal metabolic function. Slc11a2hipp/hipp mice had longer mean escape times on a cued task paradigm implying impaired procedural memory. Nevertheless, when hippocampal and striatal memory systems were placed in competition using a Morris Water Maze task that alternates spatial navigation and visual cued responses during training, and forces a choice between hippocampal and striatal strategies during probe trials, Slc11a2hipp/hipp mice used the hippocampus-dependent response less often (25%) and the visual cued response more often (75%) compared to WT littermates that used both strategies approximately equally. Hippocampal ID not only reduces spatial recognition memory performance but also affects systems that support procedural memory, suggesting an altered balance between memory systems.
Similar content being viewed by others


Introduction
Iron deficiency is the most common nutrient deficiency in the world, and affects the developing fetal brain in humans in the context of gestations complicated by maternal iron deficiency, diabetes mellitus, or hypertension (Georgieff et al. 1990; Lozoff and Georgieff 2006). Behaviorally, iron deficient infants of diabetic mothers have abnormal recognition memory at birth (Siddappa et al. 2004), a finding that remains present at 3.5 years of age and is a function of the degree of ID at birth (Riggins et al. 2009). Fetal/neonatal ID has adverse long-term effects (despite iron repletion after birth) on both the hippocampus and the striatum, which are among the primary brain regions that contribute to spatial navigation and procedural memory, respectively (Felt and Lozoff 1996; Beard et al. 2002; Felt et al. 2006; Schmidt et al. 2007; Ward et al. 2007; Carlson et al. 2009). ID reduces energy metabolism, glutamatergic and GABAergic neurotransmission, neuronal dendrite extension, and pre- and post-synaptic function in the rodent hippocampus (Jorgenson et al. 2003; Rao et al. 2003; Jorgenson et al. 2005; Carlson et al. 2009). Early ID also profoundly and permanently alters monoamine and energy metabolism in the rodent striatum in spite of iron treatment (Youdim et al. 1980; Beard et al. 2003; Beard et al. 2006; Felt et al. 2006; Ward et al. 2007).
Optimal cognitive development and subsequent performance rely on the interactions among multiple memory systems including recognition memory based largely in the hippocampus, and procedural memory linked to the dorsal striatum (White and McDonald 2002). In the Multiple Parallel Memory Systems model (White and McDonald 2002), these systems are juxtaposed as competitive, while in the Interactive Memory Systems Theory these systems act cooperatively to mediate behavior (McDonald et al. 2004). Important connections among these systems allow detection of novelty and facilitate cognitive flexibility (McDonald et al. 2002; Lisman and Grace 2005; McDonald et al. 2008). Dampening of one input system can result in dominance of another, consequently eliciting a maladapted behavioral phenotype (McDonald et al. 2004; Lee et al. 2008).
A working framework of how these individually affected areas work together to produce abnormal early life and adult behavioral phenotypes seen in ID has not been explored despite an extensive literature on the adverse effects of early ID on the primary circuitry of each memory system (Lozoff and Georgieff 2006). One approach to create such a framework is to selectively cause ID in one memory system (and not the others) to understand the role of the first system in regulating the others. This has not been possible with the maternal dietary ID model because it induces ID throughout the brain and thus affects structures in both systems (deUngria et al. 2000). Recently we reported a model that generates early ID specifically in hippocampal neurons at embryonic day 18.5 by deleting the iron transporter Slc11a2 in a conditional manner in neurons of the hippocampus (Carlson et al. 2009). The model has several advantages. It allows a reductionistic approach to examine the effects of ID in a tissue specific manner. It avoids several potential confounds of dietary models, such as anemia (which can cause fatigue and tissue hypoxia), altered maternal behavior, and increased uptake of other divalent metals. The targeted tissue knock-out obviates the problem with whole-animal Slc11a2 knockouts, which are lethal in the first week of life (Gunshin et al. 2005).
Slc11a2 encodes a channel protein involved in the transfer of ferrous iron from a clathrin-coated endosome to the cytosol, after it has been taken up into the cell on the transferrin-transferrin receptor-1 complex (Gunshin et al. 2005). Slc11a2, in addition to other iron-sensitive mRNA transcripts, are upregulated in hippocampus after spatial navigation learning, suggesting that neuronal iron uptake is critical for such learning (Carlson et al. 2009; Haeger et al. 2009). Cre-recombinase mediated deletion of this gene specifically in neurons of the developing hippocampus reduces iron content and impairs spatial navigation learning and memory in two versions of place learning in the Morris Water Maze. The effect is accompanied by altered hippocampal gene expression, energy metabolism and neuronal structure (Carlson et al. 2009). Here, we use this unique model to probe the effects of hippocampal-neuron specific ID on other brain regions and functions, in order to understand the interactions between developing spatial and procedural memory systems.
Methods
Animals
All experiments were performed in accordance with the NRC Guide for the Care and Use of Laboratory Animals, and approved by the Institutional Animal Care and Use Committee of the University of Minnesota. Slc11a2hipp/hipp mice and Slc11a2WT/WT littermates were bred as described previously; briefly: Slc11a2 flox/flox mice (Gunshin et al. 2005) were crossed with CaMKIIa-cre (L7ag#13 line, (Dragatsis and Zeitlin 2000)) transgenic mice to generate hippocampal-neuron specific knockout of Slc11a2 (Carlson et al. 2009). All animals were bred and housed in a specific pathogen free facility, and only male littermates were analyzed. Individual mouse tail DNA was genotyped by PCR, with previously published cycling conditions and primers (Carlson et al. 2009).
Tissue dissection, iron concentration analysis, and RNA collection
Male mice (not used in other experiments) at age 3 months were killed by an intraperitoneal injection of Beuthanasia (10 mg/kg) for both iron content experiments and mRNA isolation as previously described (Carlson et al. 2009). The striatum, nucleus accumbens (NA), PFC, ventral midbrain (VMB, substantia nigra and ventral tegmentum), and hippocampus were dissected and flash-frozen in liquid nitrogen. Iron concentration was analyzed with atomic absorption spectroscopy as described previously (Beard et al. 2006). Total RNA was isolated and concentrations were measured as previously described (Carlson et al. 2009).
Quantitative real-time PCR
Messenger RNA levels from the five regions were measured for 14 transcripts from P90 mice by real-time, quantitative PCR (qPCR) (Taqman) (Supplemental Table 1). Thirteen mRNA transcripts were used to assess iron metabolism, the neurometabalome, and dendritic structure. Reverse transcription was carried out using SuperScript III (Invitrogen) and random hexamers as described previously (Carlson et al. 2009).
In vivo 1H NMR spectroscopy
All spectroscopic experiments were performed as previously described (Carlson et al. 2009) on a horizontal bore 9.4 T/31 cm magnet (Varian/Magnex, Oxford, UK) equipped with a 15-cm I.D. gradient coil insert (450 mT/m, 200 μs) which included a strong second- order shim system (Resonance Research, Inc., Billerica, MA) interfaced to a Varian INOVA console (Varian, Inc., Palo Alto, CA, USA) (Tkac et al. 2004).
In vivo 1H NMR spectra were collected using a previously described protocol (Rao et al. 2003; Tkac et al. 2004; Rao et al. 2007; Ward et al. 2007; Carlson et al. 2009). Briefly, all first- and second-order shims were automatically adjusted using FASTMAP (Gruetter 1993; Gruetter and Tkac 2000). Ultra-short echo-time STEAM (echo time TE = 2 ms, repetition time TR = 5 s, number of transients NT = 240) combined with outer volume suppression and VAPOR water suppression (Tkac et al. 1999; Tkac et al. 2004) was used to acquire spetral data from 5.2 μL (1.6 × 1.8 × 1.8 mm3) volume of interest (VOI) centered in the left dorsal striatum. Multi-slice coronal and sagittal RARE imaging technique (echo train length ETL = 8, echo spacing ESP = 15 ms, TE = 60 ms, matrix = 256 × 128, FOV = 20 mm × 20 mm, slice thickness = 1 mm) was used for the selection of the VOI.
Metabolite concentrations were quantified from 1H NMR spectra using LCModel software package (Provencher 1993). The LCModel analysis calculates the best fit to the experimental spectrum as a linear combination of model solution spectra of brain metabolites. The spectrum of fast relaxing macromolecules was also included in the LCModel basis set as previously described (Pfeuffer et al. 1999; Rao et al. 2003; Tkac et al. 2004; Rao et al. 2007; Ward et al. 2007; Carlson et al. 2009). Unsuppressed water signal was used as an internal reference assuming brain water content of 80%. Concentrations of the following 14 metabolites were consistently quantified from striatal 1H NMR spectra: alanine (Ala), creatine (Cr), phosphocreatine (PCr), γ-aminobutyric acid (GABA), glucose (Glc), glutamate (Glu), glutamine (Gln), glutathione (GSH), the sum of glycerophosphorylcholine and phosphocholine (GPC + PC), myo-inositol (myo-Ins), lactate (Lac), N-acetylaspartate + N-acetylaspartylglutamate (NAA + NAAG), phosphoethanolamine (PE), and taurine (Tau). Estimated errors of metabolite concentrations (Cramer-Rao lower bounds) were 0.3–0.5 µmol/g.
Behavioral experiments
Apparatus
The Morris Water Maze (MWM) consisted of a white circular water tank 120 cm in diameter and 45 cm high, filled to 30 cm. A transparent platform (10 cm in diameter) was placed so that its surface was either 1.5 cm below the water line (hidden escape platform) or 0.5 cm above the water line and locally cued with a visible flag. The pool was located in a large test room where there were many cues external to the maze (e.g., pictures, lamps): these were visible from the pool and could be used by the mice for spatial orientation. These extra-maze cues were kept constant throughout the testing period. Water temperature was kept at 21–23°C. Animal behaviors were videotaped and analyzed by the Topscan system (Clever Systems, Reston, VA). For descriptive data collection, the pool was subdivided into four equal quadrants formed by imaginary lines.
Procedural memory experiment
This experiment was performed in animals not used in the Competition Experiment. A single habituation trial was performed 1 day before the first water maze test, by placing each mouse on the hidden platform for 30 s. If the mouse fell or jumped from the platform, it was placed back on the platform. Four cued trials, in which the escape platform protruded above the water surface at different locations within the target quadrant, were performed a day after the habituation trial.
Competition experiment
This experiment was performed in animals not used in the Procedural Memory Experiment. As with the first MWM experiment, a single habituation trial was performed 1 day before the first water maze test (Carlson et al. 2009). In this experiment, each mouse received four training trials per day for 8 consecutive days. On the first day of the experiment, mice were trained to swim to the visible platform located in the center of the target quadrant equivalent to the procedural memory experiment described above. Each daily training session consisted of four trials on which each mouse was released once from each of four start points. The trial began by placing a mouse in the pool facing the wall, at a randomly selected start position and ended when the mouse climbed onto the visible platform, or after 30 s had elapsed. If the mouse had not escaped after 30 s, it was gently placed on the platform. Each mouse was left on the platform for 30 s, and returned to its home cage. For each mouse, there was a 20–25 min delay between trials within a daily session. During the delay, the remaining mice were run on the same trial.
On the next day after the day of training on the visible platform, each mouse received a four-trial session in which the visible platform was replaced with the submerged platform in the same location. Identical training procedures were used during these hidden platform trials. Subsequently, this 2-day sequence of one visible platform session followed by one hidden platform session was repeated three more times, for a total of 32 training trials (16 visible, 16 hidden) over a total of 8 days. Escape latency (time to reach the available platform) were measured on these acquisition trials.
On day 6 and day 8, competition probes were given. The visible platform was moved to the center of the quadrant directly opposite the target quadrant used in the acquisition trials. Two trials were given from start positions equidistant to the center of the both the target quadrant used during training and the new location of the visible platform. Video recordings were used to determine whether the mice swam within 5 cm of the perimeter of the former platform location in the target quadrant before escaping to the visible platform, now located in the opposite quadrant. A point was scored for each animal each time it crossed within 5 cm of the perimeter of the former platform location in the target quadrant.
Statistical analysis
For statistical comparisons between data from Slc11a2hipp/hipp and Slc11a2WT/WT mice in iron content, gene expression, and 1H MRS studies, significance was determined using unpaired, two-tailed Student’s t tests. For behavioral data, we performed two-way ANOVA with presence or absence of Cre recombinase expression (Cre +/− status), trial number, and their interaction as variables to examine the response of each group for each trial in the visual cued task and for analysis of competition probe trials. We analyzed the aggregate responses of competition trials between Slc11a2hipp/hipp and Slc11a2WT/WT mice with unpaired, two-tailed Student’s t-test.
Results
Iron concentration in striatum and nucleus accumbens
The iron concentration was not significantly reduced in either striatum (Slc11a2hipp/hipp: 18.0 ± 0.8 μg iron/g tissue ± SEM, n = 8; Slc11a2WT/WT: 20.9 ± 1.0 μg iron/g tissue ± SEM, n = 12, P > 0.05) or NA (Slc11a2hipp/hipp: 13.5 ± 0.6 μg iron/g tissue ± SEM, n = 8, Slc11a2WT/WT: 15.0 ± 0.4 μg iron/g tissue ± SEM, n = 12, P > 0.05) in Slc11a2hipp/hipp mice at 3 months old.
Gene expression in five selected brain regions
Three month old male mice (n = 5/group) were used for all gene expression results. Slc11a2 showed decreased expression in all brain regions measured (Fig. 1a). Since Cre is not expressed in all regions of the brain of Slc11a2hipp/hipp mice, the question arose of whether the decrease in gene expression in regions of the brain besides hippocampus was due to deletion of the gene by Cre recombinase or down-regulation, potentially due to decreased demand. Thus, we directly compared expression of deleted region/floxed Slc11a2 (detected by primer set “ex6-7”) with nonfloxed/nondeleted Slc11a2 (detected by primer set “ex15-6”) in each brain region of each animal. In areas where Slc11a2 was conditionally knocked out (hippocampus, PFC, and NA), there was significant decrease in the floxed region of expression, relative to the unfloxed region (Fig. 1c). In areas (e.g., striatum, VMB) where iron demand was lower due to reduced metabolism (see 1H NMR spectroscopy results below), there were no significant relative decreases in the floxed to unfloxed region of expression (Fig. 1b–c).

Gene expression of Slc11a2 exon 6–7 (floxed region) (a) Slc11a2 exon 15–16 (unfloxed region) (b) and ratio of floxed to unfloxed expression (c) in selected brain regions. Values are means ± SEM, n = 5 for each group. *Significantly different from Slc11a2hipp/hipp mice at P < 0.05, ** P < 0.01, *** P < 0.001
Twelve additional mRNA transcripts relevant to iron metabolism, neurometabolome, and dendritic and synaptic structure and function (Carlson et al. 2009) were different in Slc11a2hipp/hipp mice compared with Slc11a2WT/WT mice in at least one brain region (Table 1).
1H NMR spectroscopy in striatum
In vivo 1H NMR spectra measured from striata of Slc11a2hipp/hipp (n = 6) and Slc11a2WT/WT mice (n = 7) are shown in Fig. 2a–b and were markedly different than spectra measured from the hippocampus in the same animals (Carlson et al. 2009). Neurochemical profiles quantified from 1H NMR spectra of Slc11a2hipp/hipp and Slc11a2WT/WT mice are compared in Fig. 3. Among the 14 neurometabolites that could be reliably quantified in the striatum, significant differences were observed in concentrations of metabolites involved in energy production (glucose, lactate, and phosphocreatine) and membrane metabolism (GPC + PC). Concentration of PCr was increased by 17% and concentrations of glucose and lactate were decreased in Slc11a2hipp/hipp mice by 18%, and 45% respectively (P < 0.05) (Fig. 3). The PCr/Cr ratio in Slc11a2hipp/hipp mice was increased by 30% (P < 0.05). Choline compounds (GPC + PC), a marker of membrane metabolism, was decreased in Slc11a2hipp/hipp mice by 19% (P < 0.05) (Fig. 3). Markers of myelination (PE, myo-inositol), GABAergic neurotransmission, and neuronal integrity (NAA + NAAG) were unchanged.

In vivo 1H NMR spectra measured from the left striatum of 3-month old Slc11a2hipp/hipp mice (n = 6) and Slc11a2WT/WT mice (n = 7). Inset: MR Images with the selected volume of interest centered in the left dorsal striatum. Abbreviations: Cr, creatine; Gln, glutamine; Glu, glutamate; GPC, glycerophosphocholine; GSH, glutathione; Lac, lactate; Ins, myo-inositol; NAA, N-acetylaspartate; PC, phosphorylcholine; PCr, phosphocreatine; Tau, taurine

Striatal neurochemical profiles of Slc11a2hipp/hipp mice (n = 6) and Slc11a2WT/WT mice (n = 7) at 3 months of age. Values are means ± SD. *Different from Slc11a2hipp/hipp mice, P < 0.05. alanine (Ala), creatine (Cr), phosphocreatine (PCr), γ-aminobutyric acid (GABA), glucose (Glc), glutamate (Glu), glutamine (Gln), glutathione (GSH), the sum of glycerophosphorylcholine and phosphocholine (GPC + PC), myo-inositol (myo-Ins), lactate (Lac), N-acetylaspartate + N-acetylaspartylglutamate (NAA + NAAG), phosphoethanolamine (PE), and taurine (Tau)
Behavioral assessment
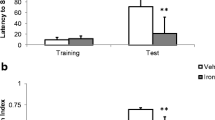
We evaluated procedural memory function using the visual cue task of the MWM. Initially, a version of the visual cued task in which the visible platform is moved between training trials to random places within the target quadrant was used. Slc11a2hipp/hipp mice (n = 17) had greater mean swim distances (ANOVA F-value = 5.65 for Cre +/− status, P < 0.05, F-value = 3.1 for trial number P < 0.05, and interaction, P > 0.05) over all, compared to Slc11a2WT/WT mice (n = 13), suggesting altered acquisition of procedural memory (Fig. 4). Slc11a2hipp/hipp and Slc11a2WT/WT mice had swim velocities that did not differ in this task (data not shown). We then determined relative contributions of spatial navigation memory and procedural memory deficits in Slc11a2hipp/hipp mice and WT littermates not used in the first experiment. We used a modified version of the MWM in which the visible platform and hidden platform training trials were alternated on a daily basis, and a competition test to probe for place and/or cued responses (McDonald and White 1994; Devan and White 1999). This version puts spatial navigation and procedural memory into direct competition (McDonald and White 1994). During training, there were no significant differences in escape latency (aggregated by training day) (data not shown) or distance traveled (aggregated by training day) between Slc11a2hipp/hipp (n = 8) and Slc11a2WT/WT (n = 6) mice on either the visible (cued) platform or the hidden platform (spatial navigation place learning) task suggesting that there was no difference between Slc11a2hipp/hipp and Slc11a2WT/WT mice in terms of vision, motivation, or fatigue in the learning of this task (Fig. 5a). However, faced with the choice of escaping to the visible platform or searching the location for the previously hidden platform on the memory system competition probes, Slc11a2hipp/hipp mice showed fewer place responses during each trial (ANOVA F-value = 4.11 for Cre +/− status, P < 0.05; trial number and interaction P > 0.05) (Fig. 5b) and in aggregate across trials (Slc11a2WT/WT had 0.58 ± 0.18 place responses/competition probe trial vs 0.25 ± 0/10 place responses/competition probe trial for Slc11a2hipp/hipp mice, P < 0.05). Therefore, they spent significantly less time (ANOVA F-value = 4.35 for Cre +/− status, P < 0.05; trial number and interaction, P > 0.05) and traveled a shorter distance (ANOVA F-value = 4.17 Cre +/− status, P < 0.05; trial number and interaction P > 0.05) in the maze than Slc11a2WT/WT mice before escaping to the visible platform (Fig. 5c), all of which are consistent with a strategy of escaping to the visible platform. They Slc11a2hipp/hipp and Slc11a2WT/WT mice had similar swim velocities during the probe trials (data not shown).

Procedural memory performance in 3-month old Slc11a2hipp/hipp mice (n = 17) and Slc11a2WT/WT mice (n = 13) in the MWM visual (cued) task. Values are distance traveled (cm) ± SEM. Performance was significantly different between Slc11a2hipp/hipp mice and Slc11a2WT/WT mice by two-way ANOVA (F-value = 5.65 for Cre +/− status, P < 0.05, F-value = 3.1 for trial number P < 0.05, interaction P > 0.05). *Significantly different from Slc11a2hipp/hipp mice on individual trial at P < 0.05 by Bonferroni’s post hoc test

Performance in memory system competition task experiment. Performance for 3-month old Slc11a2hipp/hipp mice (n = 8) and Slc11a2WT/WT mice (n = 6) in each training trial block (a) are expressed as distance traveled (cm) ± SEM. (b) Performance was significantly different between Slc11a2hipp/hipp mice and Slc11a2WT/WT mice over all probe trials by two-way ANOVA as measured by distance traveled (cm) ± SEM (Source of variation = Cre +/− status, F-value = 4.17, P < 0.05, for trial number and interaction, P > 0.05). (c) Performance was significantly different between Slc11a2hipp/hipp mice and Slc11a2WT/WT mice over all probe trials by two-way ANOVA as measured by place responses/trial (Source of variation = Cre +/− status, F-value = 4.11, P < 0.05, for trial number and interaction, P > 0.05)
Discussion
In this study, Slc11a2hipp/hipp mice, which have a 40% reduction in hippocampal iron content and abnormal spatial navigation memory (Carlson et al. 2009), disproportionately resorted to procedural memory solutions to escape the MWM, when the spatial navigation memory and procedural memory systems were placed in competition. This occurred despite the finding that Slc11a2hipp/hipp mice appear to have reduced striatal metabolism and gene expression, and altered performance in a visual cued task that relies heavily on a normally functioning dorsal striatum when compared to wild type control mice. Since iron content in the striatum was not different between Slc11a2hipp/hipp mice and their wild type littermates, we interpret the striatal changes seen in this animal to be due to impaired hippocampal input resulting from hippocampal-neuron specific ID. These abnormalities appear to index a disturbance of the balance between two cooperating systems and demonstrate the dependence of optimal striatal performance on an intact hippocampus.
Approaches to dissociate multiple parallel memory systems through stereotaxically-lesioning either the dorsal lateral striatum or hippocampus were validated in the 1990s using similar water maze competition tasks to the ones used in the current experiment (McDonald and White 1994; Devan and White 1999). A similar cue/place competition water maze task was recently validated with a transgenic mouse model and showed bidirectional competition between hippocampus and striatum during learning (Lee et al. 2008). Rats with sham surgeries show a striking balance between these two memory systems when these systems are forced to compete (McDonald and White 1994; Devan and White 1999). This balance can be altered experimentally by stereotaxically ablating one region or the other. Rats with a dorsal striatal lesion but a functioning hippocampus choose spatial navigation strategies, whereas rats with a hippocampal lesion but intact dorsal striatum choose cued strategies to solve water mazes (Packard and McGaugh 1992; McDonald and White 1994; Devan and White 1999).
Nutritional deficiencies early in life affect the developing brain because of its high demand for substrates that support the metabolic requirements of growth and differentiation (Fugelstad et al. 2008). Early deficiencies can cause brain dysfunction during the period of deficiency but importantly can also alter the developmental trajectories of rapidly developing regions such that the functions subserved by those areas remain abnormal throughout the lifespan. Unlike stereotaxic lesions, nutritional deficiencies dampen but do not obliterate neural circuitry and thus result in impairment, but not complete loss of performance. Our model of 40% ID that is specific to the hippocampus is particularly relevant to the degree of ID seen in human neonates (Petry et al. 1992). ID, the most common single nutrient deficiency in the world, has been extensively studied in humans as well as rodent and monkey models. The effects of late fetal and early postnatal ID on the brain are protean and occur when several processes of archicortical and neocortical development are underway. Early brain-wide ID leads to impairments of learning and memory, mood and affect, and speed of processing through its effects on the hippocampus, on monoamine-rich areas like the striatum and on myelin. All three processes are undergoing rapid development in the late fetal and early neonatal period (Thompson and Nelson 2001; Georgieff and Innis 2005; Ward et al. 2007), but fetal ID tends to compromise the hippocampus disproportionately (deUngria et al. 2000; Golub et al. 2007) resulting in long term spatial learning and memory abnormalities (Schmidt et al. 2007). However, prior models that induced pan-brain ID were unable to provide any understanding of the relative contributions of the three processes to altered behavioral function during ID. Thus, it was unclear whether the extensive striatal dysfunction noted in early ID (Beard et al. 2003; Ward et al. 2007) is solely due to alterations in dopaminergic neurotransmission because tyrosine hydroxylase is iron dependent or whether hippocampal inputs also compromise this region’s performance. By combining the temporal- and tissue-specific genetic manipulation with a set of molecular, biochemical and behavioral experiments designed to dissociate these memory systems, we were able in this study to differentiate the primary effects of neuronal ID as described in our previous report (Carlson et al. 2009) from secondary or downstream effects on extra-hippocampal brain regions subserving other memory systems. The current study suggests that ID-induced pathology in striatum is not solely due to direct effects (on dopamine) within that region (Beard et al. 2003), but is in part due to a direct, downstream impact from ID in another area of the brain involved in a complimentary/competitive type of cognitive function (the hippocampus). Therapeutically, strategies to shore up dopaminergic neurotransmission in the striatum are not likely to reverse all the behavioral manifestations of the disease.
Iron has an important role in neuronal metabolism through its incorporation into cytochromes (Dallman 1986). A 40% reduction in brain iron content leads to a 40% reduction in cytochrome c oxidase activity in the hippocampus, altered dendritic arborization, reduced long-term potentiation and poorer performance on spatial memory tasks (deUngria et al. 2000). Genes that are responsible for cellular iron uptake, including Slc11a2 (encoding DMT1), Tfrc, Aco1 (IRP-1), and Ireb2 (IRP-2), are upregulated in hippocampus during spatial navigation learning tasks in a dose dependent manner, consistent with a response to the increased neuronal metabolic demands induced by learning (Carlson et al. 2009). Thus, it was interesting to note that despite its normal iron content, the dorsal striatum of Slc11a2hipp/hipp mice nevertheless showed reduced expression of genes related to iron uptake (e.g., Slc11a2, Slc40a1, and Tf), energy metabolism (ckb and Slc2a3, the neuronal glucose transporter), and dendrite formation/synaptic activity compared to wild type mice. We interpret these findings to be consistent with hypoactive neural circuits in the striatum. Reductions in the transcript for tyrosine hydroxylase, the rate-limiting (and iron-requiring) enzyme in dopamine synthesis, are also evident in the striatum of Slc11a2hipp/hipp mice and suggest potential alterations in that neurotransmitter system that are not directly due to ID in the that brain region. Mbp, a marker of myelination, is downregulated in the striatum and VMB of Slc11a2hipp/hipp mice, but not in hippocampus. Myelination is a neuronal activity-dependent process (Demerens et al. 1996; Zalc and Fields 2000), and in Slc11a2 mice, decreased Mbp expression in striatum may reflect decreased basal maintenance of myelin due to suppressed basal electrical activity due to the dampening of hippocampal activity. Expression of cfos, an immediate early gene whose expression is induced in an activity-dependent manner (Herdegen and Leah 1998) was downregulated in all regions examined except NA, with its greatest reduction in striatum. Basal levels of Camk2a are elevated, and basal levels of Dlg4 (encoding PSD-95) are decreased in Slc11a2hipp/hipp mouse striatum. Camk2a and PSD-95 act in a differential manner to regulate the stability of NR2A and NR2B containing NMDA receptors in hippocampus (Park et al. 2008), and the altered basal differential expression of Dlg4 and Camk2a maybe an adaptive response to maintain long term potentiation-type plasticity in this structure, at the cost of long term depression-type stability.
The NMR spectroscopy findings support the genomic findings and indicate dorsal striatal alterations in critical metabolites such as lactate, glucose, phosphocreatine, and GPC + PC in Slc11a2hipp/hipp mice. Decreased lactate concentration and increased PCr/Cr ratio in Slc11a2hipp/hipp mouse striatum likely reflect decreased energy demands (Jost et al. 2002; Bartlett et al. 2004), and are consistent with a hypometabolic state (Mangia et al. 2007a; Mangia et al. 2007b). Lower glucose levels in the striatum are consistent with observed regional decreases in gene expression for neuronal and endothelial glucose transporters. Reductions in GPC + PC observed in Slc11a2hipp/hipp mice may represent decreased synthesis of membrane phospholipids (Ledeen 1984; Vance et al. 1994), which are required for synaptic plasticity and dendrite growth (Ehlers 2007).
A potential confounding factor in the model is that the PFC also shows significant cre deletion of Slc11a2. PFC-striatal connections are important in learning (McDonald et al. 2008), and cre-recombinase mediated reduction of Slc11a2 in PFC could influence striatal metabolism, gene expression and behavior. However, the robustness of our behavioral dissociation of the hippocampal and striatal memory systems remains compelling. Furthermore, electrophysiologic coherence between striatum and hippocampus is observed in rats that are successful in a procedural memory task, evidence of the coordinated, dynamic interactions in hippocampus and striatum during experience-dependent acquisition and performance of learned behaviors despite relatively indirect connectivity of these structures (DeCoteau et al. 2007). Since the primary lesion in Slc11a2hipp/hipp mice is in the hippocampus, we interpret our results to indicate that the dorsal striatal system does not need to expend as much metabolic activity to overcome the hippocampal system. A future experiment could use a mouse model in which the hippocampal iron deficiency was repletable to determine whether the formerly iron deficient hippocampus induced metabolic changes in the striatum returns to normal with subsequent normalization of procedural memory behaviors.
These studies were designed to better understand the role of hippocampal iron in early cognitive development. However, the neurobehavioral consequences of early ID in humans appear to extend beyond disordered cognition. Fetal or early postnatal ID also results in abnormal motor development, slower auditory and visual system neurotransmission, altered sleep architecture, poorer maternal ratings of neonatal social and emotional behaviors, and an increased incidence of behavioral problems during childhood (Lozoff and Georgieff 2006; Burden et al. 2007; Lozoff et al. 2007; Peirano et al. 2007; Lozoff et al. 2008; Corapci et al. 2009; Shafir et al. 2009). Early ID is also linked to complex behavioral abnormalities later in life such as attention deficit disorder and schizophrenia (Insel et al. 2008; Van Lieshout and Voruganti 2008; Dopheide and Pliszka 2009; Sorensen et al. 2010). These latent effects are of particular interest because their fundamental pathologies may lie outside primary target areas of early ID and their symptoms may instead represent a failure of primary areas to interconnect and regulate each other in a cooperative manner from early life onward. Hippocampal dampening may predispose an organism to a schizophrenic endophenotype, with alterations in dopamine-related behavior being a secondary consequence of prenatal or neonatal alterations in hippocampus (McDonald et al. 2004). Hyper-dopaminergic states in one animal model of schizophrenia are dependent on hippocampal dysfunction induced prenatally (Lodge and Grace 2007; Lisman et al. 2008). Another major heuristic model of the etiology of schizophrenia induces a transient disruption of neural activity in the ventral hippocampus during the neonatal period, and results in several endophenotypic features of schizophrenia later in the life of the organism (Lipska 2004; Tseng et al. 2009). In a recent large cohort study, maternal gestational ID was associated with an increased risk of schizophrenia in offspring in a dose dependent manner (Insel et al. 2008). Diabetes mellitus (DM) during gestation also increases the risk of schizophrenia in the offspring seven-fold (Cannon et al. 2002; Van Lieshout and Voruganti 2008). A nutritional hallmark of DM during pregnancy is altered fetal iron metabolism that results in a 65% rate of ID and a 40% reduction in brain iron content in the offspring (Georgieff et al. 1990; Petry et al. 1992). Maternal ID and DM during gestation increase the risk of neonatal hypoxic-ischemic injury (Rao et al. 1999; Rao et al. 2007), a condition that also increases the risk of schizophrenia in adulthood (Cannon et al. 2002).Taken together, the altered behaviors observed in children with ID are remarkably similar to the motor, cognitive, and behavioral deficits that are observed in children who go on to develop schizophrenia (Erlenmeyer-Kimling 2000; Isohanni et al. 2004). We speculate that ID induced alterations of hippocampal, prefrontal cortex (PFC) and striatal development and function in early life may induce significant imbalances in developing memory systems that may contribute to the cognitive and behavioral abnormalities similar to those seen in pre-schizophrenic children.
The hope of identifying early disruptions of memory function is that intervention is most effective when instituted during maximal brain plasticity early in life. Nutritional deficiencies and glucose intolerance during pregnancy are eminently treatable. The critical periods for developing brain regions that are particularly susceptible to ID appear to span the late fetal and early neonatal periods, suggesting that both maternal and neonatal nutritional status and metabolic health are paramount to cognitive health. Specifically, screening pregnant women for ID and DM during the last trimester and both recognizing and treating iron deficient newborns may reduce the risk to developing memory systems and of developing psychopathology later in life.
References
Bartlett DJ, Rae C, Thompson CH, Byth K, Joffe DA, Enright T, et al. Hippocampal area metabolites relate to severity and cognitive function in obstructive sleep apnea. Sleep Med. 2004;5:593–6.
Beard J, Erikson KM, Jones BC. Neonatal iron deficiency results in irreversible changes in dopamine function in rats. J Nutr. 2003;133:1174–9.
Beard JL, Erikson KM, Jones BC. Neurobehavioral analysis of developmental iron deficiency in rats. Behav Brain Res. 2002;134:517–24.
Beard JL, Felt B, Schallert T, Burhans M, Connor JR, Georgieff MK. Moderate iron deficiency in infancy: biology and behavior in young rats. Behav Brain Res. 2006;170:224–32.
Burden MJ, Westerlund AJ, Armony-Sivan R, Nelson CA, Jacobson SW, Lozoff B, et al. An event-related potential study of attention and recognition memory in infants with iron-deficiency anemia. Pediatrics. 2007;120:e336–45.
Cannon M, Jones PB, Murray RM. Obstetric complications and schizophrenia: historical and meta-analytic review. Am J Psychiatry. 2002;159:1080–92.
Carlson ES, Tkac I, Magid R, O’Connor MB, Andrews NC, Schallert T, et al. Iron is essential for neuron development and memory function in mouse hippocampus. J Nutr. 2009;139:672–9.
Corapci F, Calatroni A, Kaciroti N, Jimenez E, Lozoff B. Longitudinal evaluation of externalizing and internalizing behavior problems following iron deficiency in infancy. J Pediatr Psychol. 2010;35:296–305.
Dallman PR. Biochemical basis for the manifestations of iron deficiency. Annu Rev Nutr. 1986;6:13–40.
deUngria M, Rao R, Wobken JD, Luciana M, Nelson CA, Georgieff MK. Perinatal iron deficiency decreases cytochrome c oxidase (CytOx) activity in selected regions of neonatal rat brain. Pediatr Res. 2000;48:169–76.
DeCoteau WE, Thorn C, Gibson DJ, Courtemanche R, Mitra P, Kubota Y, et al. Learning-related coordination of striatal and hippocampal theta rhythms during acquisition of a procedural maze task. Proc Natl Acad Sci USA. 2007;104:5644–9.
Demerens C, Stankoff B, Logak M, Anglade P, Allinquant B, Couraud F, et al. Induction of myelination in the central nervous system by electrical activity. Proc Natl Acad Sci USA. 1996;93:9887–92.
Devan BD, White NM. Parallel information processing in the dorsal striatum: relation to hippocampal function. J Neurosci. 1999;19:2789–98.
Dopheide JA, Pliszka SR. Attention-deficit-hyperactivity disorder: an update. Pharmacotherapy. 2009;29:656–79.
Dragatsis I, Zeitlin S. CaMKIIalpha-Cre transgene expression and recombination patterns in the mouse brain. Genesis. 2000;26:133–5.
Ehlers MD. Secrets of the secretory pathway in dendrite growth. Neuron. 2007;55:686–9.
Erlenmeyer-Kimling L. Neurobehavioral deficits in offspring of schizophrenic parents: liability indicators and predictors of illness. Am J Med Genet. 2000;97:65–71.
Felt BT, Beard JL, Schallert T, Shao J, Aldridge JW, Connor JR, et al. Persistent neurochemical and behavioral abnormalities in adulthood despite early iron supplementation for perinatal iron deficiency anemia in rats. Behav Brain Res. 2006;171:261–70.
Felt BT, Lozoff B. Brain iron and behavior of rats are not normalized by treatment of iron deficiency anemia during early development. J Nutr. 1996;126:693–701.
Fugelstad A, Rao R, Georgieff MK. The role of nutrition in cognitive development. Cambridge: MIT Press; 2008.
Georgieff MK, Innis SM. Controversial nutrients that potentially affect preterm neurodevelopment: essential fatty acids and iron. Pediatr Res. 2005;57:99R–103.
Georgieff MK, Landon MB, Mills MM, Hedlund BE, Faassen AE, Schmidt RL, et al. Abnormal iron distribution in infants of diabetic mothers: spectrum and maternal antecedents. J Pediatr. 1990;117:455–61.
Golub MS, Hogrefe CE, Germann SL. Iron deprivation during fetal development changes the behavior of juvenile rhesus monkeys. J Nutr. 2007;137:979–84.
Gruetter R. Automatic, localized in vivo adjustment of all first- and second-order shim coils. Magn Reson Med. 1993;29:804–11.
Gruetter R, Tkac I. Field mapping without reference scan using asymmetric echo-planar techniques. Magn Reson Med. 2000;43:319–23.
Gunshin H, Fujiwara Y, Custodio AO, Direnzo C, Robine S, Andrews NC. Slc11a2 is required for intestinal iron absorption and erythropoiesis but dispensable in placenta and liver. J Clin Invest. 2005;115:1258–66.
Haeger P, Alvarez A, Leal N, Adasme T, Nunez MT, Hidalgo C. Increased Hippocampal Expression of the Divalent Metal Transporter 1 (DMT1) mRNA Variants 1B and + IRE and DMT1 Protein After NMDA-Receptor Stimulation or Spatial Memory Training. Neurotox Res. 2010;17:238–247.
Herdegen T, Leah JD. Inducible and constitutive transcription factors in the mammalian nervous system: control of gene expression by Jun, Fos and Krox, and CREB/ATF proteins. Brain Res Brain Res Rev. 1998;28:370–490.
Insel BJ, Schaefer CA, McKeague IW, Susser ES, Brown AS. Maternal iron deficiency and the risk of schizophrenia in offspring. Arch Gen Psychiatry. 2008;65:1136–44.
Isohanni M, Murray GK, Jokelainen J, Croudace T, Jones PB. The persistence of developmental markers in childhood and adolescence and risk for schizophrenic psychoses in adult life. A 34-year follow-up of the Northern Finland 1966 birth cohort. Schizophr Res. 2004;71:213–25.
Jorgenson LA, Sun M, O’Connor M, Georgieff MK. Fetal iron deficiency disrupts the maturation of synaptic function and efficacy in area CA1 of the developing rat hippocampus. Hippocampus. 2005;15:1094–102.
Jorgenson LA, Wobken JD, Georgieff MK. Perinatal iron deficiency alters apical dendritic growth in hippocampal CA1 pyramidal neurons. Dev Neurosci. 2003;25:412–20.
Jost CR, Van Der Zee CE, In’t Zandt HJ, Oerlemans F, Verheij M, Streijger F, et al. Creatine kinase B-driven energy transfer in the brain is important for habituation and spatial learning behaviour, mossy fibre field size and determination of seizure susceptibility. Eur J Neurosci. 2002;5:1692–706.
Ledeen RW. Lipid-metabolizing enzymes of myelin and their relation to the axon. J Lipid Res. 1984;25:1548–54.
Lee AS, Duman RS, Pittenger C. A double dissociation revealing bidirectional competition between striatum and hippocampus during learning. Proc Natl Acad Sci USA. 2008;105:17163–8.
Lipska BK. Using animal models to test a neurodevelopmental hypothesis of schizophrenia. J Psychiatry Neurosci. 2004;29:282–6.
Lisman JE, Coyle JT, Green RW, Javitt DC, Benes FM, Heckers S, et al. Circuit-based framework for understanding neurotransmitter and risk gene interactions in schizophrenia. Trends Neurosci. 2008;31:234–42.
Lisman JE, Grace AA. The hippocampal-VTA loop: controlling the entry of information into long-term memory. Neuron. 2005;46:703–13.
Lodge DJ, Grace AA. Aberrant hippocampal activity underlies the dopamine dysregulation in an animal model of schizophrenia. J Neurosci. 2007;27:11424–30.
Lozoff B, Clark KM, Jing Y, Armony-Sivan R, Angelilli ML, Jacobson SW. Dose-response relationships between iron deficiency with or without anemia and infant social-emotional behavior. J Pediatr. 2008;152:696–702. 702.31-3.
Lozoff B, Corapci F, Burden MJ, Kaciroti N, Angulo-Barroso R, Sazawal S, et al. Preschool-aged children with iron deficiency anemia show altered affect and behavior. J Nutr. 2007;137:683–9.
Lozoff B, Georgieff MK. Iron deficiency and brain development. Semin Pediatr Neurol. 2006;13:158–65.
Mangia S, Tkac I, Gruetter R, Van de Moortele PF, Maraviglia B, Ugurbil K. Sustained neuronal activation raises oxidative metabolism to a new steady-state level: evidence from 1H NMR spectroscopy in the human visual cortex. J Cereb Blood Flow Metab. 2007a;27:1055–63.
Mangia S, Tkac I, Logothetis NK, Gruetter R, Van de Moortele PF, Ugurbil K. Dynamics of lactate concentration and blood oxygen level-dependent effect in the human visual cortex during repeated identical stimuli. J Neurosci Res. 2007b;85:3340–6.
McDonald RJ, Devan BD, Hong NS. Multiple memory systems: the power of interactions. Neurobiol Learn Mem. 2004;82:333–46.
McDonald RJ, King AL, Foong N, Rizos Z, Hong NS. Neurotoxic lesions of the medial prefrontal cortex or medial striatum impair multiple-location place learning in the water task: evidence for neural structures with complementary roles in behavioural flexibility. Exp Brain Res. 2008;187:419–27.
McDonald RJ, Ko CH, Hong NS. Attenuation of context-specific inhibition on reversal learning of a stimulus-response task in rats with neurotoxic hippocampal damage. Behav Brain Res. 2002;136:113–26.
McDonald RJ, White NM. Parallel information processing in the water maze: evidence for independent memory systems involving dorsal striatum and hippocampus. Behav Neural Biol. 1994;61:260–70.
Packard MG, McGaugh JL. Double dissociation of fornix and caudate nucleus lesions on acquisition of two water maze tasks: further evidence for multiple memory systems. Behav Neurosci. 1992;106:439–46.
Park CS, Elgersma Y, Grant SG, Morrison JH. alpha-Isoform of calcium-calmodulin-dependent protein kinase II and postsynaptic density protein 95 differentially regulate synaptic expression of NR2A- and NR2B-containing N-methyl-d-aspartate receptors in hippocampus. Neuroscience. 2008;151:43–55.
Peirano PD, Algarin CR, Garrido MI, Lozoff B. Iron deficiency anemia in infancy is associated with altered temporal organization of sleep states in childhood. Pediatr Res. 2007;62:715–9.
Petry CD, Eaton MA, Wobken JD, Mills MM, Johnson DE, Georgieff MK. Iron deficiency of liver, heart, and brain in newborn infants of diabetic mothers. J Pediatr. 1992;121:109–14.
Pfeuffer J, Tkac I, Provencher SW, Gruetter R. Toward an in vivo neurochemical profile: quantification of 18 metabolites in short-echo-time (1)H NMR spectra of the rat brain. J Magn Reson. 1999;141:104–20.
Provencher SW. Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn Reson Med. 1993;30:672–9.
Rao R, de Ungria M, Sullivan D, Wu P, Wobken JD, Nelson CA, et al. Perinatal brain iron deficiency increases the vulnerability of rat hippocampus to hypoxic ischemic insult. J Nutr. 1999;129:199–206.
Rao R, Tkac I, Townsend EL, Ennis K, Gruetter R, Georgieff MK. Perinatal iron deficiency predisposes the developing rat hippocampus to greater injury from mild to moderate hypoxia-ischemia. J Cereb Blood Flow Metab. 2007;27:729–40.
Rao R, Tkac I, Townsend EL, Gruetter R, Georgieff MK. Perinatal iron deficiency alters the neurochemical profile of the developing rat hippocampus. J Nutr. 2003;133:3215–21.
Riggins T, Miller NC, Bauer PJ, Georgieff MK, Nelson CA. Electrophysiological indices of memory for temporal order in early childhood: implications for the development of recollection. Dev Sci. 2009;12:209–19.
Schmidt AT, Waldow KJ, Grove WM, Salinas JA, Georgieff MK. Dissociating the long-term effects of fetal/neonatal iron deficiency on three types of learning in the rat. Behav Neurosci. 2007;121:475–82.
Shafir T, Angulo-Barroso R, Su J, Jacobson SW, Lozoff B. Iron deficiency anemia in infancy and reach and grasp development. Infant Behav Dev. 2009;32:366–75.
Siddappa AM, Georgieff MK, Wewerka S, Worwa C, Nelson CA, Deregnier RA. Iron deficiency alters auditory recognition memory in newborn infants of diabetic mothers. Pediatr Res. 2004;55:1034–41.
Sorensen HJ, Nielsen PR, Pedersen CB, Mortensen PB. Association between prepartum maternal iron deficiency and offspring risk of schizophrenia: population-based cohort study with linkage of danish national registers. Schizophr Bull. 2010. doi:10.1093/schbul/sbp167
Thompson RA, Nelson CA. Developmental science and the media. Early brain development. Am Psychol. 2001;56:5–15.
Tkac I, Henry PG, Andersen P, Keene CD, Low WC, Gruetter R. Highly resolved in vivo 1H NMR spectroscopy of the mouse brain at 9.4 T. Magn Reson Med. 2004;52:478–84.
Tkac I, Starcuk Z, Choi IY, Gruetter R. In vivo 1H NMR spectroscopy of rat brain at 1 ms echo time. Magn Reson Med. 1999;41:649–56.
Tseng KY, Chambers RA, Lipska BK. The neonatal ventral hippocampal lesion as a heuristic neurodevelopmental model of schizophrenia. Behav Brain Res. 2009;204:295–305.
Van Lieshout RJ, Voruganti LP. Diabetes mellitus during pregnancy and increased risk of schizophrenia in offspring: a review of the evidence and putative mechanisms. J Psychiatry Neurosci. 2008;33:395–404.
Vance JE, Pan D, Campenot RB, Bussiere M, Vance DE. Evidence that the major membrane lipids, except cholesterol, are made in axons of cultured rat sympathetic neurons. J Neurochem. 1994;62:329–37.
Ward KL, Tkac I, Jing Y, Felt B, Beard J, Connor J, et al. Gestational and lactational iron deficiency alters the developing striatal metabolome and associated behaviors in young rats. J Nutr. 2007;137:1043–9.
White NM, McDonald RJ. Multiple parallel memory systems in the brain of the rat. Neurobiol Learn Mem. 2002;77:125–84.
Youdim MB, Green AR, Bloomfield MR, Mitchell BD, Heal DJ, Grahame-Smith DG. The effects of iron deficiency on brain biogenic monoamine biochemistry and function in rats. Neuropharmacology. 1980;19:259–67.
Zalc B, Fields RD. Do action potentials regulate myelination? Neuroscientist. 2000;6:5–13.
Acknowledgements
Supported by the grants: NIH R21 HD054490, NIH R01 HD29421, Minnesota Medical Foundation at the University of Minnesota, Vikings Children’s Fund at the University of Minnesota to M.K.G.; NIH: NINDS Individual Kirschstein-NRSA F31 NS047876, a seed grant from the American Medical Association Foundation, and the Dr. Warren J. Warwick and Henrietta Holm Warwick Fellowship Award to E.S.C; and NIH P-01 HD039386 awarded to M.K.G., T.S, R.R. and E.U; NIH grants P41 RR008079, P30 NS057091 and the Keck Foundation to I.T.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
SUPPLEMENTAL TABLE 1
Genes examined by quantitative PCR in Slc11a2hipphipp mice. (DOC 102 kb)
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Carlson, E.S., Fretham, S.J.B., Unger, E. et al. Hippocampus specific iron deficiency alters competition and cooperation between developing memory systems. J Neurodevelop Disord 2, 133–143 (2010). https://doi.org/10.1007/s11689-010-9049-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11689-010-9049-0