
Abstract
Silvicultural practices applied in managed forest plantations may help counteract the effects of climate change by influencing soil surface CO2 efflux (Fs). Understanding the effects of silvicultural practices on Fs will provide unbiased estimates of carbon fluxes and allow better silvicultural decisions for carbon sequestration. Therefore, we assessed how Fs differed seasonally across silvicultural practices (i.e., stocking levels, clone, fertilization and weed control treatments) and evaluated the effects of soil temperature (Ts) and soil volumetric water content (θv) on Fs across these practices for a mid-rotation (14 year-old) Pinus radiata plantation in the Canterbury region of New Zealand. There were significant differences in Fs (p < 0.05) over the four seasons, three levels of stocking, and five clones. The effects of fertilization and weed control applied 12 years previously on Fs were insignificant. Annual estimate of Fs (mean ± 1 standard deviation) from the study site was 22.7 ± 7.1 t ha−1 a−1 in the form of CO2 (6.2 ± 2.1 t ha−1 a−1 in the form of C). Fs values were consistently higher in plots with 1250 stems ha−1 compared to 2500 stems ha−1, which may be related to a strong soil resource limitation because of the close spacing in the latter plantation. Significant differences in Fs across clones suggest that variations in carbon partitioning might explain their growth performance. Silvicultural treatments influenced Fs response to soil temperature (p < 0.05), resulting in models explaining 28–49% of the total variance in Fs. These findings provide insights into how silvicultural management decisions may impact Fs in mid-rotation radiata pine plantations, contributing towards developing more precise and unbiased plantation carbon budgets.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Forests store some 75% of all carbon in terrestrial ecosystems accounting for up to 40% of carbon exchange between the biosphere and the atmosphere (Raich and Schlesinger 1992). Soil CO2 efflux is the second largest flux in the global carbon cycle, which is a magnitude greater than emissions from the burning of fossil fuel (Raich and Schlesinger 1992). Soil CO2 efflux changes due to anthropogenic factors may therefore accelerate global warming by speeding up global carbon cycling (Raich and Potter 1995; Schlesinger and Andrews 2000). For example, land clearing and utilization, and deforestation increases soil CO2 efflux as a result of increased rates of decomposition (Rapp 2014). Given the extent of global forest cover, minor changes in disturbance regimes and forest management practices may considerably affect soil carbon fluxes (Maier and Kress 2000).
Radiata pine (Pinus radiata D. Don) plantations occupy more than four million hectares worldwide (Mead 2013), exhibiting greater productivity and plasticity than many other tree species. In New Zealand, radiata pine is the predominantly planted species, accounting for about 90% of 1.73 million ha (NZFOA 2019). Intensively managed plantations represent an important pool in the global carbon cycle (Templeton et al. 2015). The productivity of radiata pine plantations can be greatly altered by silvicultural management practices (Hollinger et al. 1993). In intensively managed plantations, these practices may contribute to counteract the effects of climate change. Total soil surface CO2 efflux (Fs) consists of two parts (Tyree et al. 2014): (1) autotrophic respiration, which is root respiration resulting from maintenance, growth and ion uptake; and, (2) heterotrophic respiration from the decomposition of soil organic matter by micro and macro fauna. Environmental factors significantly influence forest productivity and decomposition of soil organic matter, explaining seasonal variation in Fs (Schlesinger and Andrews 2000). For example, Fs was higher in summer and lower in winter in North Florida’s slash pine (Pinus elliottii Engelm) plantation, corresponding to changes in ambient temperatures and soil moisture (Ewel et al. 1987).
Fertilization, weed control, planting at different stocking levels and with different genotypes are common silvicultural practices for radiata pine plantations throughout New Zealand (Mason and Milne 1999). The purpose of these practices is to produce healthy and vigorous trees by enhancing growth and productivity (Mason 1992). A previous study indicated that silvicultural practices have long term impacts on Fs in intensively managed plantations (Tyree et al. 2006). These practices can significantly influence Fs by altering the microclimate of the site, including light, soil moisture (θv), soil temperature (Ts), and the soil microbial community. For example, fertilization has been shown to decrease (Samuelson et al. 2004), be independent of (Tyree et al. 2006; Templeton et al. 2015), and increase (Bracho et al. 2018) Fs in loblolly pine (Pinus taeda L.) plantations. Moreover, two contrasting loblolly pine clones differed in their Fs responses to nutrient manipulations in terms of both partitioning and physiology (Tyree et al. 2009). Furthermore, stocking levels significantly affected both heterotrophic and autotrophic soil CO2 efflux in lodgepole pine (Pinus contorta Dougl.) (Litton et al. 2003a).
The main research topics for radiata pine plantations in New Zealand are silviculture practices, genetics (Mason and Kirongo 1999; Lasserre et al. 2005; Mason 2008), growth and yield modelling, fertilization, and weed control practices (Mason et al. 1996; Mason and Milne 1999), and carbon cycling and partitioning (KC et al. 2020). However, understanding how these practices affect the dynamics of Fs in radiata pine plantations is limited. Therefore, the objectives of this study were: (a) to assess the influence of stocking, fertilization, weed control, and clone selection on Fs; and, (b) to determine the influence of soil temperature and soil volumetric water on Fs across silvicultural treatments in a young radiata pine plantation. It was hypothesized that: (1) Fs would increase with stocking; (2) faster growing clones would exhibit greater Fs; (3) Fs would be independent of fertilization carried out 12 years before; (4) Fs would be independent of weed control carried out 12 years before; and, (5) both Ts and θv would explain most of the variations in Fs across the silvicultural treatments. This study will contribute to better understanding of differences among silvicultural management decisions on Fs when developing stand-level carbon budget models in forest plantations.
Materials and methods
Study site and experiment
The experiment was located in Rolleston, Canterbury, New Zealand (43° 37.2′ S and 172° 20.4′ E) (Fig. 1a). It was established by the School of Forestry, University of Canterbury on land owned by the Selwyn District Council, comprising 7.5 ha of radiata pine planted in 2005. The site is approximately 45 m a.s.l. on a plain (Fig. 1a). Typical summer daytime maximum temperatures range from 18 to 26 °C (Fig. 1b), but may rise above 30 °C. Winters have daytime maximum temperatures from 7 to 14 °C (Salekin et al. 2019; NIWA 2020) (Fig. 1b). Mean annual rainfall is approximately 618 mm with a monthly range of 38 to 68 mm (NIWA 2020) (Fig. 1b). Northeasterly winds occur most frequently in coastal sites throughout the year while southwesterly winds are more frequent during winter (NIWA 2020). The site had formerly been livestock pasture. The soil is a Lismore stony silt loam with aggradation gravel as a parent material and also includes partial glacial gravel (Rennie and Bennett 1981; Hewitt 2010).

Location (a), ombrothermic diagram–January–December 2017 (b) and layout of the experimental trial (c)
The experimental design consisted of 48 permanent plots with a randomized complete block factorial split plot with four complete blocks (Mason 2008), with an arrangement of factors within each block. Plots 1–12, 13–24, 25–36, and 37–48 corresponded to blocks one, two, three, and four respectively (Fig. 1c). The main plots consisted of three levels of stocking (625, 1250 and 2500 stems ha−1). A first split consisted of four levels of follow-up weed control and fertilization treatments (fertilization, F; herbicide, H; both, FH; and no chemicals). Fertilization was carried out once in year 1 and once in year 3 (Nitrogen, Phosphorus, Potassium, Sulphur, and trace elements at a rate of 80 g per tree). Strip weed control was applied in years 1 and 2, and a follow-up herbicide treatment with complete clearance in year 3 and in subsequent years when required. A second split consisted of five different clones randomly allocated to all plots, with 1, 2, 3, 4 and 5 indicating different clonal genotypes (Mason 2008). Measurements of soil surface CO2 efflux rates (Fs) were carried out when the plantation was 14 years-old, 12 years after the last fertilization and herbicide applications were performed.
Measurement of soil surface CO2 efflux
Efflux rates (Fs) were measured using an infrared gas analyzer (EGM-4, PP Systems, Hitchin, Hertfordshire, UK) equipped with a soil respiration chamber (SRC-1) with a 10- cm inner diameter. Two hundred and forty PVC collars (10 cm wide × 6 cm high) were placed into the soil and left undisturbed for one year before Fs measurements were made. Fs was measured at the centre within the two rows for each clone in each plot. Nine hundred and sixty measurements were made (48 plots × 5 clones × 4 seasons). Measurements were carried out between 9:00 AM and 4:00 PM during autumn (13–16 April 2017), winter (7–10 July 2017), spring (3–6 October 2017), and summer (13–16 January 2018).
Measurement of soil temperature and soil water content
Soil temperature (Ts) and soil volumetric water (θv) were measured simultaneously to Fs, within 10 cm of the PVC collar. Ts values were determined using a built-in temperature probe (STP-1) of the EGM-4 at a 10-cm soil depth. The θv was measured with a portable moisture meter SM150T (Delta-T Devices Ltd., Burwell, Cambridge, UK), at 10 cm. Measurements of the SM150T were calibrated using samples analyzed by a gravimetric method consisting of oven drying soil samples of a known volume (Walker et al. 2004).
Scaling to annual soil surface CO2 efflux
Fs values were scaled into t ha−1 a−1 in the form of CO2 using the exponential equations of Fs vs Ts and θv developed for this experiment (Eq. 4) and continuous measurements of Ts and θv recorded by in-site micrometeorological stations.
Statistical analysis
The influence of stocking, specific clone, fertilization, weed control, and season on Fs, Ts, and θv were examined using linear mixed effects models of the nlme package in R statistical software (R Core Team 2018), considering blocks as random effects. Fixed effects consisted of stocking, clone, fertilization, and weed control and their two-way interactions. A mathematical representation of the linear mixed-effects model in matrix form is given in Eq. (1) (Zuur et al. 2009).
where Yi is the response variable (Fs, Ts, and θv) for different treatment levels i (i.e., three levels of stocking, two levels of fertilization, two levels of weed control, and five levels of type of clone), \(X_{i} \times \beta\) is the fixed term, \(Z_{i} \times b_{i}\) is the random term, and \(\varepsilon_{i}\) is error term.
To determine the influence of Ts and θv on Fs across silvicultural treatments, the following commonly used exponential functions were tested: Ts-based model [Eq. (2)] (Lloyd and Taylor 1994), and the combined Ts and θv-based model [Eq. (4)] (Lavigne et al. 2004). Temperature sensitivity (Q10), i.e., the response of Fs to a 10 °C change in Ts values, was estimated across all silvicultural treatments using Eq. (3) based on the model of Lloyd and Taylor (1994). Fitted models were evaluated using two of the most commonly used goodness-of-fit statistics (Huber-Carol et al. 2012): the root mean square error (RMSE), and the coefficient of determination (R2). Regression models were fitted in the R statistical software (R Core Team 2018).
where Fs is the measured soil surface CO2 efflux rate (µmol CO2 m−2 s−1), Ts is the measured soil temperature at 10 cm soil depth (°C), Q10 is the temperature sensitivity response of Fs, θv is the measured volumetric water content (%) at 10 cm soil depth; and a, b, and c are fitted parameters of the regression.
Results
Influence of T s and θ v on F s across silvicultural treatments
For the period of the experiment, Ts was strongly controlled by the season (F3,889 = 9677, p < 0.001), and specific clone (F4,889 = 9.37, p < 0.001) (Table 1). Stocking, fertilization, and follow-up herbicide treatments had no significant effects on Ts (p > 0.05). When examined by season, Ts was significantly influenced by the main clonal effect in all seasons (F4,172 > 3.27, p < 0.05) (Table 2). Volumetric water content (θv) followed an opposite trend to Ts (soil temperature), being lower in summer (7.3% ± 1%) compared to spring (14.6% ± 1.6%), winter (17.1% ± 1%), and autumn (17.7% ± 1%) (Table 3). For the period of the experiment, θv was strongly affected by the interacting effects of stocking × herbicide (F2,29 = 8.06, p < 0.01) (Table 1). Significant interactions of stocking × fertilization (F2,29 = 5, p < 0.05) and herbicide × clone (F4,172 = 2.54, p < 0.05) were observed in autumn and spring, respectively (Table 2). No significant (p > 0.05) effects of silvicultural treatments on θv were observed in summer (Table 2).
The combined Ts and θv-based model [Eq. (4)] was selected as best to predict Fs as it yielded highest R2 values and lowest RMSE compared to Ts-based models across all levels of treatments (Table 4). The Q10 values from the combined model ranged from 2.91 to 5.23 and were marginally higher than the ones estimated from the Ts-based model, with the highest Q10 observed with clone 3 and the lowest in clone 2. Comparing across silvicultural treatments, Q10 values were higher with clone 3, a stocking level of 1250 stems ha−1, and in unfertilized plots without weed control than in other treatments (Table 4). The slope between Ts and Fs increased exponentially and was higher in fertilized plots (Fig. S1-A), and in plots without weed control (Fig. S1-B) than in unfertilized plots and in plots with weed control. The slope of the relationship between Ts and Fs tended to be higher for the 1250 stems ha−1 stocking than for stocking levels of 625 and 2500 stems ha−1 (Fig. S1-C). For the model with clones, the slope of Ts with Fs was significantly higher in clone 3 than in the other four clones (Fig. S1-D). The slope of the Fs-Ts was positive but that of the Fs-θv was negative across all levels of treatments (Table 4). There was a strongly negative correlation between the Ts and θv (r = − 0.89) (Fig. S2), indicating that soil temperatures were high when soil volumetric water was low, and vice versa. Therefore, in the regression models for the relationship between Fs against Ts, and θv, the estimated values of parameters a and b were always positive, and values of c always negative (Table 4).
Influence of season and silvicultural treatments on surface CO2 efflux
Values of Fs were strongly controlled by the effects of season (F3,889 = 319.68, p < 0.001), stocking (F2,6 = 9.92, p < 0.05), and clone (F4,889 = 4.32, p < 0.01) (Table 1). No significant effects of fertilization or follow-up weed control, or two-way interaction between stocking levels, clone, fertilization, and weed control treatments on Fs were observed (p > 0.05) (Table 1). Annual Fs estimated from the study site was on average 22.7 ± 7.1 t ha−1 a−1 for CO2 (range 15.6–27.8). The rate of Fs was highest in autumn (27.7 ± 7.1 t ha−1 a−1 for CO2, range 25.2–32.5) and lowest in winter (15.6 ± 7.0 t ha−1 a−1 for CO2, range 14.4–17.4) (Table 3).
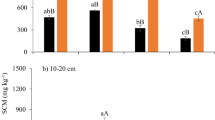
When seasonal data were examined separately, the clone significantly affected Fs for summer (F4,172 = 2.68, p < 0.05), and autumn (F4,172 = 7.74, p < 0.001) (Table 2). There were no significant differences between mean soil efflux rates for any clones during winter and spring. Clone 3 exhibited a consistently higher Fs compared to other clones (Table 3). For all clones, values of Fs were at their highest in autumn and lowest in winter, while spring and summer showed intermediate rates between these two extremes (Fig. 2a). Stocking levels significantly influenced Fs in spring (F2,6 = 17.69, p < 0.01) and summer (F2,6 = 7.75, p < 0.05) (Table 2). In spring, Fs was highest (29.5 t ha−1 a−1 for CO2) for the 1250 stems ha−1 stocking level and lowest (20.9 t ha−1 a−1 for CO2) for the 2500 stems ha−1 stocking level (Fig. 2b, Table 3). Fs values were consistently higher at a stocking level of 1250 stems ha−1compared to 625 (by 22%) and 2500 (by 18%) stems ha−1 over all seasons (Table 3). There were no significant main or interactive effects of fertilization and herbicide treatments on Fs (p > 0.05) (Table 2).

Seasonal dynamics of Fs (µmol CO2 m−2 s−1) across silvicultural treatments across five clones (a), and three stockings (b). Values are presented as least square mean (± 1 SE) of Fs, by season. Treatment means within a season followed by the same letter do not differ significantly at α = 0.05 level using Tukey’s HSD test
Discussion
The first hypothesis that Fs would increase with stocking because of greater root and microbial biomass and competition, was only partially supported by the results of this study. Fs values were higher at a medium (1250 stems ha−1) stocking level compared to low (625 stems ha−1) and high (2500 stems ha−1) stocking level across all silvicultural treatments. Observed values of Fs (20.6–26.3 t ha−1 a−1 for CO2) were within the range reported by Noh et al. (2010) for a Korean red pine (Pinus densifolia Siebold & Zucc.) forest in Korea (22.8–27.3 t ha−1 a−1 for CO2) with the highest value at a medium stocking level, similar to this study. Other studies, however, have found that surface CO2 efflux increased with stand density (Litton et al. 2003b, 2004) which might be explained by greater root and microbial biomass and higher litter production and decomposition (Litton et al. 2001) at higher densities. There are several possible causes why Fs was greatest at a medium (1250 stems ha−1) level of stocking. First, surface CO2 efflux may increase from a low to medium stocking level brought about by increases in root and microbial biomass. In addition, Fs may decrease from a medium to a high level of stocking associated with (1) lower soil temperatures and less available water at high stocking levels or, (2) light becoming the most limiting growth factor triggering greater carbon allocation aboveground at the expense of belowground processes. The former may be more likely as our results showed that soil volumetric water content was significantly lower at 2500 stems ha−1 by 2.6 and 1.6% compared to 625 and 1250 stems ha−1, respectively (Table 3).
The second hypothesis was that faster growing clones would show higher surface CO2 efflux levels, which is fully supported by this study. Clone 3, the fastest growing, had the highest Fs average values (3–10%) compared with the other clones. This indicates that clone 3, having overall greater carbon assimilation, also allocated a greater proportion of fixed carbon belowground compared to the other clones (Bown et al. 2009).
The third hypothesis for this study was that fertilization would not influence surface CO2 efflux as it was carried out 12 years before the measurements. There were no significant effects of fertilization on surface CO2 emissions in any season during the year which is consistent with research on radiata pine in Chile (Bown and Watt 2016), slash pine in Florida, USA (Shan et al. 2001), and loblolly pine in Virginia (Tyree et al. 2006) and North Carolina, USA (Maier and Kress 2000). Our study, however, contains only one-year data and the absence of inter-annual variability may prevent finding Fs responses to fertilization (Tyree et al. 2006). However, the insignificant effects of fertilization on surface CO2 efflux in this study may be because the last fertilization was applied three years after planting and its effects after nine years might have ceased. It may also be because the fertilization level was too small to trigger a growth response (Tyree et al. 2006). At the same time, our temperate study site experiences strong water limitations, particularly during summer, and therefore water rather than nutrients might be the major limiting factor affecting plant growth (Davidson et al. 1998; Lavigne et al. 2004).
The fourth hypothesis that there would be no effects of weed control on soil surface CO2 efflux was supported by this study. In contrast, research by Shan et al. (2001) in slash pine plantations in Florida, USA, indicated that the elimination of the understory significantly affected surface CO2 effluxes. This suggests that weed control reduced Fs by decreasing below-ground biomass. However, we believe the insignificant effects of weed control on Fs in this study is the result of the time elapsed. Weed control was applied to all plots during the first two years, while total weed control was carried out prior to canopy closure in weed control plots only (Mason 2008). However, surface CO2 efflux measurements in this study were carried out at age 12, eight years after the follow up weed control when the effects of weeds were likely to have ceased.
The fifth hypothesis that both soil temperature and soil water would control CO2 effluxes across all silvicultural treatments is supported by this study. Collectively, both explain 27.9–48.9% of the efflux variance as being significantly better to the model that considered only soil temperature (Table 4). Other studies have also reported that the combination of soil temperature and water can explain most of the variations in CO2 efflux as being the most important controlling factors (Gough and Seiler 2004; Liu et al. 2011). Soil temperatures contributed positively to Fs, while soil water contributed negatively (Table 4). However, there was a strong, negative correlation between sol temperature and soil water (Fig. S2), indicating that soil temperatures were high when soil water was low and vice versa, which may confound the interpretation of the model. The temperature sensitivity of surface CO2 efflux (Q10) values in this study (2.9 to 5.2) across all silvicultural treatments were in the 1–10 range of other reports (Raich and Schlesinger 1992; Davidson et al. 1998; Gulledge and Schimel 2000; Xu and Qi 2001a, b) depending on the type of ecosystem and climatic conditions. The Q10 value in this study was highest for the fast-growing clone 3 at a stocking level of 1250 stems ha−1.
This study provides insight into how silvicultural management decisions may impact soil surface CO2 effluxes and may contribute towards better estimates of carbon budgets in plantations. Surface CO2 emissions varied with plantation density and clone, and with soil temperatures and water content, both contributing to the differences in fluxes across seasons. Higher efflux rates at a medium (1250 stems ha−1) stocking level compared to low (625 stems ha−1) and high (2500 stems ha−1) stocking levels were observed. There were no effects of fertilization and weed control after 12 years, indicating that the effects of these treatments might have ceased. Efflux levels varied with clones, being higher in faster growing clones. In conclusion, surface CO2 effluxes changed with silvicultural practices which might have implications for carbon accounting.
References
Bown H, Watt M (2016) Stem and soil CO2 efflux responses of P. radiata plantations to temperature, season, age, time (day/night) and fertilization. Cienc Investig Agrar 43:95–109
Bown H, Watt M, Clinton P, Mason E, Whitehead D (2009) The influence of N and P supply and genotype on carbon flux and partitioning in potted Pinus radiata plants. Tree Physiol 29:857–868
Bracho R, Vogel JG, Will RE, Noormets A, Samuelson LJ, Jokela EJ, Gonzalez-Benecke CA, Gezan SA, Markewitz D, Seiler JR, Strahm BD, Teskey RO, Fox TR, Kane MB, Laviner MA, McElligot KM, Yang J, Lin W, Meek CR, Cucinella J, Akers MK, Martin TA (2018) Carbon accumulation in loblolly pine plantations is increased by fertilization across a soil moisture availability gradient. For Ecol Manag 424:39–52
Davidson EA, Belk E, Boone RD (1998) Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob Change Biol 4:217–227
Ewel KC, Cropper WP, Gholz HL (1987) Soil CO2 evolution in Florida slash pine plantations. I. Changes through time. Can J For Res 17:325–329
Gough CM, Seiler JR (2004) Belowground carbon dynamics in loblolly pine (Pinus taeda) immediately following diammonium phosphate fertilization. Tree Physiol 24:845–851
Gulledge J, Schimel JP (2000) Controls on soil carbon dioxide and methane fluxes in a variety of taiga forest stands in interior Alaska. Ecosystems 3:269–282
Hewitt AE (2010) New Zealand soil classification, Landcare research science series no. 1, 3rd edn. Manaaki Whenua Press, Lincoln, Canterbury
Hollinger D, Maclaren J, Beets P, Turland J (1993) Carbon sequestration by New Zealand’s plantation forests. N Z J For Sci 23:194–208
Huber-Carol C, Balakrishnan N, Nikulin MS, Mesbah M (eds) (2012) Goodness-of-fit tests and model validity. Springer, New York
KC M, Mason EG, Bown HE (2020) Linking above-ground biomass production to below-ground carbon fluxes across stocking, clone, fertilization, and understory elimination in Pinus radiata D. Don plantations, New Zealand. For Ecol Manag 477:118–469
Lasserre JP, Mason E, Watt M (2005) The effects of genotype and spacing on Pinus radiata D. Don corewood stiffness in an 11-year old experiment. For Ecol Manag 205:375–383
Lavigne M, Foster R, Goodine G (2004) Seasonal and annual changes in soil respiration in relation to soil temperature, water potential and trenching. Tree Physiol 24:415–424
Litton CM, Knight DH, Ryan MG (2001) Above-and belowground carbon allocation in post-fire lodgepole pine forests: effects of tree density and stand age. Univ Wyom Natl Park Serv Res Cent Ann Rep 25:123–129
Litton CM, Ryan MG, Knight DH (2004) Effects of tree density and stand age on carbon allocation patterns in postfire lodgepole pine. Ecol Appl 14:460–475
Litton CM, Ryan MG, Knight DH, Stahl PD (2003a) Soil-surface carbon dioxide efflux and microbial biomass in relation to tree density 13 years after a stand replacing fire in a lodgepole pine ecosystem. Glob Change Biol 9:680–696
Litton CM, Ryan MG, Tinker DB, Knight DH (2003b) Belowground and aboveground biomass in young postfire lodgepole pine forests of contrasting tree density. Can J For Res 33:351–363
Liu J, Jiang PK, Wang HL, Zhou GM, Wu JS, Yang F, Qian XB (2011) Seasonal soil CO2 efflux dynamics after land use change from a natural forest to Moso bamboo plantations in subtropical China. For Ecol Manag 262:1131–1137
Lloyd J, Taylor J (1994) On the temperature dependence of soil respiration. Funct Ecol 315–323
Maier CA, Kress L (2000) Soil CO2 evolution and root respiration in 11 year-old loblolly pine (Pinus taeda) plantations as affected by moisture and nutrient availability. Can J For Res 30:347–359
Mason E (1992) Decision-support systems for establishing radiata pine plantations in the Central North Island of New Zealand. PhD thesis, University of Canterbury, Christchurch, New Zealand
Mason E (2008) Influences of silviculture, genetics and environment on radiata pine corewood properties: results from recent studies and a future direction. N Z J For Sci 53:26–31
Mason E, Kirongo B (1999) Responses of radiata pine clones to varying levels of pasture competition in a semiarid environment. Can J For Res 29:934–939
Mason E, Milne P (1999) Effects of weed control, fertilization, and soil cultivation on the growth of Pinus radiata at midrotation in Canterbury, New Zealand. Can J For Res 29:985–992
Mason E, South DB, Weizhong Z (1996) Performance of Pinus radiata in relation to seedling grade, weed control, and soil cultivation in the central North Island of New Zealand. N Z J For Sci 26:173–183
Mead D (2013) Sustainable management of Pinus radiata plantations: FAO Forestry Paper No. 170. FAO, Rome
NIWA (2020) Overview of New Zealand's climate. https://niwa.co.nz/education-and-training/schools/resources/climate/overview. Accessed 28 Oct 2020
Noh NJ, Son Y, Lee SK, Yoon TK, Seo KW, Kim C, Lee WK, Bae SW, Hwang J (2010) Influence of stand density on soil CO2 efflux for a Pinus densiflora forest in Korea. J Plant Res 123:411–419
NZFOA (2019) Facts & Figures 2018/19. New Zealand Plantation Forest Industry. New Zealand, https://www.nzfoa.org.nz/resources/publications/facts-and-figures
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Raich JW, Potter CS (1995) Global patterns of carbon dioxide emissions from soils. Glob Biogeochem Cycles 9(1):23–36
Raich JW, Schlesinger WH (1992) The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B Chem Phys Meteorol 44:81–99
Rapp D (2014) Anthropogenic influences on climate change. In: Assessing climate change. Springer Praxis Books. Springer, Cham, pp 533–595. https://doi.org/10.1007/978-3-319-00455-6_7
Rennie WF, Bennett CM (1981) Soils of the Rolleston new town site, Canterbury, New Zealand. N.Z. soil survey report 61. New Zealand Soil Bureau, Department of Scientific and Industrial Research, Wellington. https://doi.org/10.7931/DL1-SSR-61
Salekin S, Mason EG, Morgenroth J, Bloomberg M, Meason DF (2019) Modelling the effect of microsite influences on the growth and survival of juvenile Eucalyptus globoidea (Blakely) and Eucalyptus bosistoana (F. Muell) in New Zealand. Forests 10:857
Samuelson LJ, Johnsen K, Stokes T, Lu W (2004) Intensive management modifies soil CO2 efflux in 6-year-old Pinus taeda L. stands. For Ecol Manag 200:335–345
Schlesinger WH, Andrews JA (2000) Soil respiration and the global carbon cycle. Biogeochemistry 48:7–20
Shan JP, Morris LA, Hendrick RL (2001) The effects of management on soil and plant carbon sequestration in slash pine plantations. J Appl Ecol 38:932–941
Templeton BS, Seiler JR, Peterson JA, Tyree MC (2015) Environmental and stand management influences on soil CO2 efflux across the range of loblolly pine. For Ecol Manag 355:15–23
Tyree MC, Seiler JR, Aust WM, Sampson DA, Fox TR (2006) Long-term effects of site preparation and fertilization on total soil CO2 efflux and heterotrophic respiration in a 33-year-old Pinus taeda L. plantation on the wet flats of the Virginia Lower Coastal Plain. For Ecol Manag 234:363–369
Tyree MC, Seiler JR, Maier CA (2014) Contrasting genotypes, soil amendments, and their interactive effects on short-term total soil CO2 efflux in a 3-year-old Pinus taeda L. plantation. Soil Biol Biochem 69:93–100
Tyree MC, Seiler JR, Maier CA, Johnsen KH (2009) Pinus taeda clones and soil nutrient availability: effects of soil organic matter incorporation and fertilization on biomass partitioning and leaf physiology. Tree Physiol 29:1117–1131
Walker JP, Willgoose GR, Kalma JD (2004) In situ measurement of soil moisture: a comparison of techniques. J Hydrol 293:85–99
Xu M, Qi Y (2001a) Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in Northern California. Glob Change Biol 7:667–677
Xu M, Qi Y (2001b) Spatial and seasonal variations of Q10 determined by soil respiration measurements at a Sierra Nevadan forest. Global Biogeochem Cycles 15:687–696
Zuur A, Ieno E, Walker N, Saveliev A, Smith G (2009) Mixed effects models and extensions in ecology with R. Springer, New Work
Acknowledgements
Thanks to the New Zealand School of Forestry, University of Canterbury, for permission to access the experimental site and for arranging the necessary equipment during the research. The corresponding author thanks the NZ Ministry of Foreign Affair and Trade (MFAT), as this research was partially supported by MFAT's “NZAID Programme” as a postgraduate research allowance to the primary author, which facilitated the seasonal data collection. We would like to thank undergraduate as well as postgraduate students at the School of Forestry who contributed to field measurements. We are grateful to anonymous reviewers and editors for their insightful comments on the manuscript.
Author information
Authors and Affiliations
Contributions
Mohan KC, Euan G. Mason and Horacio E. Bown developed the concept for this manuscript. Mohan KC carried out field measurements, conducted data analysis, and drafted the manuscript under the supervision of Euan G. Mason and Horacio E. Bown. All authors contributed to subsequent writing, reviewed, and approved the final version of the manuscript.
Corresponding author
Additional information
Corresponding editor: Tao Xu.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: The work was supported by the NZ Ministry of Foreign Affair and Trade (MFAT)’s NZAID Programme.
The online version is available at https://www.springerlink.com.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
KC, M., Mason, E.G. & Bown, H.E. The influence of stocking levels, clone, fertilization, and weed control on surface CO2 efflux in a mid-rotation Pinus radiata D. Don plantation in Canterbury, New Zealand. J. For. Res. 32, 2567–2575 (2021). https://doi.org/10.1007/s11676-021-01298-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-021-01298-9