
Abstract
In this study, we investigate the phylogeny and taxonomy of the /Camptori clade sensu Soop et al. (2019a). Based on combined nrDNA ITS phylogenetic, morphological, and ecological analyses, the clade includes six species in Europe, four of which are described here as new to science: Cortinarius malavalii, C. marklundii, C. violaceoserrulatus, and C. viridocaelestinus. We also provide a taxonomic revision of the two known species in the clade, C. camptoros and C. viridocoeruleus. In its current concept, the /Camptori clade is mostly represented in frondose woodlands of southern Europe and none of the species is found in the boreal coniferous zones of Northern Europe. Three species are strictly thermophilous Mediterranean-submediterranean species associated mainly with southern Quercus spp., whereas C. camptoros is found mainly in montane Abies forests, and C. marklundii reaches boreonemoral Tilia-Corylus sites in S Scandinavia and alpine Dryas sites in Spain. An identification key to the species is presented.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The huge genus Cortinarius has traditionally been divided into 5–8 subgenera (Moser 1967, Brandrud et al. 1990–2018, Jeppesen et al. 2012), of which subgen. Phlegmacium, Telamonia, and Dermocybe are the largest and most well-known, based on morphology. Molecular phylogenetic studies, however, have shown that these entities do not represent unique lineages and need to be emended or more seriously revised to better reflect evolutionary history within the genus (e.g. Peintner et al. 2004, Garnica et al. 2005). Recently, Liimatainen et al. (2022) have split the genus Cortinarius into 10 genera based on genomic and multi-gene sequence data. The new classification, however, is not yet adopted in the current work, which has been submitted before the latter publication.
The traditional subgenus Phlegmacium, in particular, encompasses three major clades: /Phlegmacium I, II and /Calochroi (Stensrud et al. 2014, Garnica et al. 2016, Soop et al. 2019a). For this reason, members of the morphologically based subgenus Phlegmacium are nowadays collectively referred to as “phlegmacioid species”. The presently treated /Camptori clade s. Soop et al. (2019a) belongs to the major clade Phlegmacium I (Garnica et al. 2016), together with morphologically resembling taxa belonging to /Dionysae clade, sections Glaucopodes, Amoenolentes, and Aureocistophili (Soop et al. 2019a, b). All these groups are characterized by (i) a distinctly duplex structure of pileipellis, including a “subcellular” subcutis/hypoderm of inflated elements, and (ii) citriform-amygdaloid spores with verrucose, discrete (little confluent) ornaments. Associated with the pileipellis structure comes also the ability of pilei to get easy water-soaked, exhibiting dark, hygrophanous spots or zones in moist weather (like many telamonioid species).
During the last 10–15 years, the taxonomy of a number of phlegmacioid groups/sections has been extensively revised in Europe on the basis of combined morphological and phylogenetic evidence: sect. Calochroi (Frøslev 2007, Frøslev et al. 2007; cf. also Garnica et al. 2009), (sub)section Elegantiores (Garnica et al. 2006), sect. Claricolores (Brandrud et al. 2013), sect. Multiformes (Brandrud et al., 2014a), sect. Purpurascentes (Saar et al. 2014), sect. Riederi (Brandrud et al. 2018), sect. Aureocistophili (Fernández-Brime et al. 2014, Soop et al. 2019b), and subsect. Varii (Schmidt-Stohn et al. 2020). Comprehensive taxonomic studies are, however, still lacking for many groups in Europe.
The /Camptori clade, together with the related /Dionysae clade, sect. Amoenolentes and sect. Aureocistophili, consists of mainly southern, Mediterranean, poorly known species that have been little studied until quite recently. There are hardly any “traces” of Camptori species in classical mycological literature, not even in the Phlegmacium monograph of Moser (1961). This situation remained so until C. viridocoeruleus was described by Chevassut & Henry (1975) and C. camptoros by Brandrud & Melot (1983). Over the years, we have been gradually convinced that these two species were both collective, but clarifying these species complexes with additional phylogenetic methods took a longer time. The revision of /Camptori is presented here with detailed species-level taxonomy, including type studies, based on combined molecular, morphological, and ecological data. Four out of six European Camptori species are here presented as new to science. They are all relatively rare and have restricted distributions in poorly studied regions of Europe, so we find it very likely that further taxa are to be discovered, especially in the Mediterranean region.
Materials and methods
Molecular analyses
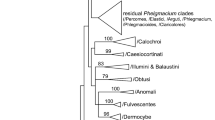
A total of 42 Camptori specimens from eight European countries were studied, including the type specimens of Cortinarius camptoros, C. purpuratus, and C. viridocoeruleus. Additional eight European and four North American Camptori as well as three outgroup sequences were retrieved from GenBank (Table 1). For most of the DNA extractions as well as the PCR and sequencing of the universal fungal DNA barcode region (Schoch et al. 2012), the ribosomal internal transcribed spacer (ITS1–5.8S–ITS2), we followed the protocols described in Liimatainen et al. (2014), Borovička et al. (2015), Dima et al. (2016), and Brandrud et al. (2018). Primer pairs of ITS1F-ITS4, and alternatively, in case of old type materials ITS1F/ITS2 and ITS3/4 (Gardes & Bruns 1993, White et al. 1990) were used to amplify the whole or partial ITS region. For three specimens (types of C. camptoros, C. purpuratus, and C. viridocoeruleus), the amplicons were sequenced with NGS sequencing on Illumina MiSeq using tagged primers and a setup and bioinformatic approach similar to that in Frøslev et al. (2019). GenBank accession numbers of the sequences used in the phylogenetic analyses are listed in Table 1. Multiple sequence alignment was done in MAFFT v7 (http://mafft.cbrc.jp/alignment/server) using the E-INS-I method (Katoh & Standley 2013) with default settings. The alignment was checked and trimmed in SeaView 4 (Gouy et al. 2010). Following Nagy et al. (2012), the phylogenetically informative indels were coded as presence/absence data with FastGap 1.2 (Borchsenius 2009), applying the simple indel coding algorithm (Simmons et al. 2001). The final alignment including nucleotide and binary characters was analysed in RAxML (Stamatakis 2014) and MrBayes 3.1.2 (Ronquist & Huelsenbeck 2003). Maximum likelihood (ML) phylogenetic reconstruction was performed in raxmlGUI (Silvestro & Michalak 2012) using rapid bootstrap analysis with 1,000 replicates. Three nucleotide partitions (ITS1, 5.8S, ITS2) were set to the GTRGAMMA substitution model in addition to one binary partition (indel characters) that was set to default. In the Bayesian inference (BI) phylogeny, the alignment was divided into four partitions (ITS1, 5.8S, ITS2, and indels) as well. The GTR + Γ substitution model was applied to the nucleotide characters, while the two-parameter Markov model was set for the indels. Two independent runs of four Markov chain Monte Carlo (MCMC) were performed each for 5 000 000 generations, sampling every 1000th generation. The first 30% of the trees were discarded as burn-in. For the remaining trees, a 50% majority rule consensus phylogram with posterior probabilities as nodal supports was computed. The best scoring ML tree from Maximum likelihood analysis was further edited in MEGA 7 (Kumar et al. 2016) and Adobe Illustrator CS4 and shown in Fig. 1.

Maximum likelihood phylogenetic tree of the /Camptori clade of the genus Cortinarius based on nrDNA ITS sequence analyses with gap coding. Sequences are labelled with their voucher numbers/GenBank accession numbers and geographical origin. Type sequences are marked with (T). Species treated in this study are highlighted in colour boxes. ML bootstrap values >70% as well as Bayesian posterior probabilities >0.6 are placed above or below branches. Scale bar indicates 0.03 expected change per site per branch
Morphological studies
Fresh material was examined mainly from N, C, W, and S Europe. More collections than those sequenced were studied morphologically, but the character variation described in the taxonomic part is based on the sequenced material. Specimens examined morphologically and sequenced are listed under each species description (see also Table 1); most specimens were examined both macro- and micromorphologically.
The taxonomic descriptions are based on the material studied by the authors. The measurements of macromorphological characters stem from expanded, but never old (and then often aberrant) basidiocarps. The terminology of characters follows Brandrud et al. (1990–2018) and Brandrud (1996).
Microscopic structures were observed partly from fresh material mounted in H2O, often with a drop of 40% KOH subsequently added, and partly from dried material mounted in H2O and then KOH.
Basidiospore measurements were made in L4 solution according to Clémençon (Clémençon 1972, Erb and Matheis, 1983) at 1000× magnification with the Zeiss microscope Axioskop 40 and the Zeiss Axiocam 208 color on a flat screen with the program ZEN core 3.1.88.0000 from Zeiss. The measurements are based on at least 30 spores from each specimen/basidiocarp; in the descriptions, numbers in square brackets refer (in this order) to the number of collections they originate from, the number of basidiocarps, and the number of spores measured, respectively. Spore measurements are given as follows: length range × width range. Q values were calculated as follows: Q = length divided by width. MV/MVs indicate “mean value(s)”. To exclude aberrant spores, the given values in the descriptions are based only on spores within the 95% confidence interval.
The photo micrographs of the spores are created with the method of “focus-stacking” (Schmidt-Stohn 2011). About 25 shots with the Zeiss Axiocam 208 color digital camera and the Zeiss Plan-Apochromat 63×/1.40 Oil, each shot with different focus, are combined with the final picture with the Helicon Focus 7.6.6 software. For such small, reproducible focus steps (each about 0.1–0.2 μm in height), a self-made mechanical equipment, coupled with the focusing fine drive of the microscope, is used (any further questions about the equipment, please direct to the first author). For the correction and the final arrangement of the spores on the plates, Adobe Photoshop version 22.1.0 was used.
The scatter diagrams of the spore dimensions are plotted with Microsoft Excel. For the final presentation, the graphics were copied from Excel and edited with Adobe Photoshop.
Results
Molecular phylogeny
The final dataset is composed of 57 nrDNA ITS sequences. The alignment including the nucleotide and indel characters was 654+54=708 characters long (alignment is included in the 12). Phylogenetic trees from ML and BI analyses showed congruent topologies. The best scoring ML tree is shown in Fig. 1. The /Camptori clade was introduced by Soop et al. (2019a), and the concept is followed here. So far, the circumscription is based mostly on European species and a few North American lineages known from sequence data. The /Camptori clade is well-supported in all phylogenetic analyses (single- and multi-gene as well).
Ten species-level terminal clades were resolved within the /Camptori clade based on our phylogenetic analyses. Four North American lineages (Cortinarius velicopius and three yet nameless species) are not treated further in this work. The remaining, moderately to well-supported six clades represent European species. Holotype sequences of C. camptoros, C. lepistoides, C. purpuratus, and C. viridocoeruleus clustered into two clades, representing C. camptoros and C. viridocoeruleus, with C. purpuratus and C. lepistoides as younger synonyms of the latter name. Four lineages did not match any type sequences, and based on their unique morphological and genetic features, we formally describe them below as C. malavalii, C. marklundii, C. violaceoserrulatus, and C. viridocaelestinus spp. nov. (see “Taxonomy” part). The minimal phylogenetic distance in the nrDNA ITS region between the six identified species ranges from 6 (C. camptoros/C. viridocoeruleus) to 36 (C. malavalii/C. marklundii) nucleotide and indel differences, representing c. 1 to 6% of sequence divergence, respectively. Intraspecific sequence variability is low among the species: 0–2 nucleotide or indel differences, representing maximum of 0.3% of sequence divergence.
Morphological characteristics of the /Camptori clade
The members of the /Camptori clade have some distinctive characters on group level. The pileus is glabrous and often hygrophanous towards margin, or with hygrophanous spots or steaks. The context appears elastic. The lamellae are tinged violaceous blue, often with a pronounced and persistent violaceous edge.
A total of 1113 spores from 26 collections/basidiocarps were measured and partially also documented photographically in this study. The mean values of length (L) × width (W) for different species fall between 8.7–10.4 × 5.1–6.2 μm (Table 2). With such narrow ranges, it is hardly possible to calculate statistically significant differences (Fig. 2). Nevertheless, there are clear differences, e.g., between C. viridocoeruleus and C. violaceoserrulatus (Fig. 3). The shape of the spores is a rather well-characterizing feature of the /Camptori clade as a group, all species possessing distinctly citriform spores with an ornament consisting of isolated to slightly interconnected warts and a more or less visible suprahilar plage (Fig. 4).

Scatter diagram representing the variation of the basidiospores of the species/collections in the /Camptori clade of the genus Cortinarius. Each dot is the MV of one collection

Scatter diagram representing the variation of basidiospores of C. viridocoeruleus and C. violaceoserrulatus. Each dot stands for one spore (only values within the 95% confidence interval)

Basidiospores of selected collections of the species in the /Camptori clade of the genus Cortinarius
The pileipellis is distinctly duplex, with a subcellular subcutis (hypoderm) of inflated, more or less cemented elements, resembling the structure of many telamonioid species, having also similar macrofeatures, i.e. the hygrophanous structure.
Habitat and distribution
Most members of the /Camptori clade are thermophilous, with preference for Mediterranean and lowland temperate regions of Europe. Half of the species (C. malavalii, C. violaceoserrulatus, C. viridicaelestinus) are seemingly strictly southern Quercus associates, whereas C. viridocoeruleus has a wider temperate-Mediterranean Fagus-Quercus (-Carpinus) range. Cortinarius marklundii has a peculiar, patchy W European distribution, with core areas in Spain (Dryas sites), France-Germany (lowland Fagus sites), and S Scandinavia (coastal Tilia-Corylus sites). According to present data, C. camptoros is restricted to montane Abies alba forests in C and S Europe.
Taxonomy
Description of the /Camptori clade
Basidiocarps fleshy and phlegmacioid, pileus glabrous and viscid-glutinous, somewhat elastic (like a Lyophyllum sp.), mostly more or less hygrophanous, pileus colours from violaceous-blue, greyish blue, ochraceous yellow, olivaceous brown to vivid ochraceous redbrown. Lamellae usually tinged violaceous blue to lilaceous when young, often with more or less persistent lilac lamellae edge, otherwise soon fading to greyish brown. Stipe with a (slightly) marginate-bulbous base, glossy, initially violaceous blue, sometimes persistent at apex.
Universal veil remnants sparse to (rarely) abundant and leaving silvery-whitish patches on pileus surface; sometimes initially with distinct and whitish-bluish volva at bulb margin. Context initially bluish in stipe, becoming ochraceous brown tinged in pileus and bulb.
Basidiospores distinctly citriform, finely to medium coarsely verrucose with isolated and also interconnected warts, mostly clearly visible in the outline of the spores, often with suprahilar plage. Pileipellis duplex, with a distinct, subcellular subcutis/hypoderm (resembling that of telamonioid taxa) and above with a thin epicutis of narrow repent-entangled hyphae. Habitat mainly in temperate-Mediterranean frondose forests.
Cortinarius camptoros Brandrud & Melot, Bull. Soc. Myc. Fr. 99(2): 219 (1983)
Fig. 5a–d

Basidiocarps of the species in the /Camptori clade of the genus Cortinarius (voucher numbers and geographical origin in brackets): a, b, c, d C. camptoros (a Melot 81.359 holotype Germany, b GS16-5 Germany, c JB-9819-18 Spain, d GS20-08 Germany); e, f, g, h C. viridocoeruleus (e SSt20-029 Sweden, f SSt04-110 Germany, g TEB984-13 France, h TF2000-056, holotype of C. lepistoides, Denmark). Photos: a J. Melot; b G. Saar; c J. Ballarà; d U. Winkler; e, f G. Schmidt-Stohn; g T.E. Brandrud; h T.S. Jeppesen
MycoBank: MB 108801
Holotype: Germany, Baden-Württemberg, Baar, Trossingen, 10 Oct 1981, J. Melot 81.359 (herb. J. Melot). GenBank ON006470 (ITS).
Description: compiled from the original diagnosis, as well as from own (sequenced) material. Pileus 30–80 mm, hemispherical, soon plano-convex to broadly umbonate, sometimes irregular, margin involute, finally often split, thin, pronounced and significantly elastic (like Lyophyllum decastes), viscid-glutinous, with a greasy appearance, glabrous and glossy when dry, often with a dark, hygrophanous marginal zone or radiate hygrophanous streaks, mature specimens often becoming radially wrinkled upon drying; initially olivaceous grey to grey brown or horn brown, with more ochraceous yellow at centre, often distinctly bicoloured, with ochre to chestnut brown at centre and hygrophanous dark olive grey at margin, becoming darker olivaceous brown at centre, or centre becoming vividly ochre yellow orange (holotype) or remaining ochraceous brown, at margin discolouring to greyish ochraceous, in places with olive tones or also flamed redbrown and sometimes developing redbrown spots; very thin and fugacious frosty-micaceous veil remnants present at centre when young, sometimes silvery-fibrillose from veil also towards margin. Lamellae normally distant to a little bit more crowded, 4–6 mm broad, broadly emarginate, edge initially irregular wavy-serrulate-fimbriate, first bluish grey to whitish grey, with bluish-violaceous tinge towards edge, often with distinctly and persistently violaceous blue edge, finally ochraceous to rusty brown. Stipe 40–100 × 9–20 mm, cylindrical with a marginate, not very distinct bulb (up to 25–30 mm wide), initially distinct greyish violet, especially in upper part and at bulb margin, often persistently with bluish tinge, finally discolouring to whitish, and sometimes with brass brown patches. Veil remnants at bulb margin thin but distinct when young, initially greyish blue to whitish, soon disappearing. Basal mycelium whitish or pale bluish. Context fleshy in the centre and thin at margin, in the pileus and the bulb brownish white, sometimes discolouring to yellow orange in the bulb, greyish violet in the entire stipe, often rather persistent, then fading to (greyish) white in the stipe and slightly yellowish in the base of bulb, sometimes saffron yellow discolouring where eaten by larvae in bulb. Odour distinctly earthy, like C. variecolor, in the bulb sometimes like honey, taste mild. Macrochemical reactions: negative with KOH in the context, light/pale ochraceous on the basal tomentum and reddish brown on the pileus, negative with Guaiac. Exsiccata with a red-brown pileus and a pale brown stipe. Basidiospores [3, 3, 144], 8.1–10.5 × 4.8–6 μm (MV = 9.3 × 5.4 μm); variation of MVs: 8.9–9.6 × 5.3–5.5 μm; Q = 1.56–1.88 (MV = 1.69), brown, distinctly citriform, finely to medium strongly and densely verrucose, warts sometimes isolated but mostly somewhat interconnected, clearly visible in the outline of the spores, suprahilar plage not clearly visible. Lamellar edge partially fertile, with clavate, cylindrical, fusiform or irregularly shaped sterile cells (resembling those of C. dionysae). Basidia with granulate-refractive substance.
Pileipellis duplex, epicutis thin, consisting of 2–5 μm wide, more or less hyaline hyphae, here and there with some yellow brown (probably cytoplasmic) pigment. Hypoderm (subcutis) well developed, elements subcellular, 15–30 μm wide, with (chocolate) brown parietal (thick-wall), amber-like pigment, cementing the elements to a compact structure.
Habitat and distribution: Associated with Abies alba and probably also Picea abies in calcareous, montane coniferous forests. Since lacking from Phlegmacium-rich pure Picea forests, it seems probable that this species has a strict affinity to the calcareous, mixed Abies alba forest type, and that it preferentially forms mycorrhizae with Abies alba. Cortinarius camptoros is mainly verified from the montane Abies alba-Picea abies-Pinus sylvestris forests of the limestone plateaus of Black Forest, SW Germany, and also from the Eastern Prealps in Austria, as well as one collection from the Pyrenees of NE Spain. There are also finds from the Western Prealps in France (Vercors) as well as from the limestone plateaus of French and Swiss Jura that fit with the present species, but these collections are not verified by sequencing.
Comments: Cortinarius camptoros is characterized by its often bicoloured clay grey + ochraceous yellow pileus, turning more vividly redbrown on exposure, the more or less persistent bluish lamellae edge and its habitat in montane, calcareous Abies-Picea forests of C Europe. The (lilac) blue lamellae edge distinguishes this from C. marklundii (described below), which also differs in habitat (associated with Fagus, Tilia, and Corylus), and is phylogenetically well-separated. Cortinarius marklundii was included in the initial, wide concept of C. camptoros in Brandrud & Melot (1983) but with a separate comment (“Récolte….Norvège… sous feuillus”). See further comments under C. marklundii. There is much confusion in the literature around the name C. camptoros. Because of our limited knowledge of the Camptori species, hitherto most hygrophanous collections with similar characters have been erroneously identified as C. camptoros. For instance, Brandrud et al. (1990–2018), following the initial wide concept of C. camptoros in Brandrud & Melot (1983), did not depict the true C. camptoros on plate C24 of the CFP (coll. CFP1024), but an undescribed species at that time (here described as C. marklundii). Moreover, to our current knowledge, C. camptoros is a rare species and considering its initial collective morphological description, it may remain difficult to identify. Habitat may constitute the best distinguishing character, because all sequenced collections of the species originated from sites with coniferous trees (Abies, Picea and Pinus), whereas all other species of the /Camptori clade are found under broadleaved trees. Most confirmed collections of C. camptoros are from a quite restricted geographical area spanning SW Germany to the eastern part of the Black Forest on limestone, and extending to Eastern Austria. There is also one verified collection from the eastern Pyrenees. All other finds mentioned in the literature under the name C. camptoros (including varieties or subspecies) outside the Abies range must be regarded as (highly) uncertain identifications, if not supported by DNA sequence data. Cortinarius camptoros can also be misidentified as C. viridocoeruleus. However, C. viridocoeruleus usually has more bluish tinges on the pileus when young, and in some stages, it may show pronounced greenish tinges, which is never seen in C. camptoros. Cortinarius viridocoeruleus displays a different ecology, growing with Fagus or Quercus. In the protologue of C. camptoros, two collections were described from calcareous Abies alba, Picea abies, and Pinus sylvestris forests of the Black Forest region—the type from Trossingen, Baar, and another collection from Neubulach, Calw (W of Stuttgart, Germany) (Brandrud & Melot 1983). Of these, only the type has been sequenced. Later, we have verified the species from more localities in the Black Forest in calcareous coniferous forests with Abies. Seven localities from the Black Forest region are reported in Krieglsteiner & Gminder (2010). In these sites, C. camptoros often occurs together with other, typical Abies alba-associated species such as C. atrovirens and C. dibaphus, and it seems likely that Abies might be the preferred ectomycorrhizal partner of C. camptoros too. Based on our observations over many years, C. camptoros is one of the rarest phlegmacioid species in these Abies-dominated ecosystems. Consiglio et al. (2007) reported a find of C. viridocoeruleus var. camptoros (Brandrud & Melot) D. Antonini, M. Antonini & Consiglio under Pinus sylvestris and Quercus ilex var. ballota from Morella, NE Spain. Based on photo and morphological description, it may belong to this species, although this record is from a somewhat deviating habitat. Unfortunately, sequencing attempts on this specimen failed, and the identity of this record remains uncertain. Fernández Sasia & Cadiñanos Aguirre (2000) described a C. camptoros f. fuscoilicis Cadiñanos from Quercus faginea and Q. ilex forests of Spain. This has also not been verified by sequencing, and with its quite deviating ecology, it seems highly unlikely to be conspecific with C. camptoros. Cortinarius camptoros differs from the closest species, C. viridocoeruleus (Fig. 1), by 6 substitution and indel positions, representing 1% of sequence divergence in the ITS region.
Additional collections examined: Austria, Niederösterreich, Schneebergdörfl NW, 9 Oct 2017, T.E. Brandrud, I. Krisai-Greilhuber, H. Voglmayr, TEB 901-17 (WU). Germany, Baden-Württemberg, Dunningen, 24 Sept 1995, U. Luhmann, TUB011848; Flözlingen, 9 Oct 2004, G. Saar, TUB019788; Schönbronn, Giffizenmoos, 31 Oct 2016, G. Saar, GS 16-5; Baden-Württemberg, Flözlingen, 17 Oct 2020, U. Winkler, GS 20-08. Spain, Clot Molina, Peguera, Figols, Berguedà, 21 Sept 2018, J. Ballarà, JB-9819-18 (herb. J. Ballarà).
Cortinarius viridocoeruleus Chevassut & Rob. Henry, Doc. Mycol. 5(20): 24 (1975)
Fig. 5e–h
MycoBank: MB 312199
Holotype: France, Hérault, Montpellier, Bois de Maurin, Quercus ilex, 19 Nov 1972, G. Chevassut, Henry3366 (PC), GenBank ON006503 (ITS).
= Cortinarius purpuratus Rob. Henry, Bull. trimest. Soc. mycol. Fr. 101(1): 28 (1985), MycoBank: MB 105185. Holotype: France, Henry84/31 (PC)
= Cortinarius lepistoides T. S. Jeppesen & Frøslev, Mycotaxon 106: 474 (2009). MycoBank: MB 512356. Holotype: Denmark, Jutland, Vosnæs havskov, 30 Sept 2000, TF2000-56 (C).
Description: compiled from the protologue, Frøslev & Jeppesen (2009), Eyssartier (2004), and own findings. Pileus 30–100 mm, hemispherical, then plano-convex, slightly glutinous, glabrous and glossy when dry, often somewhat radially wrinkled upon drying, some specimens towards margin at about half the diameter distinctly innately fibrillose (it is uncertain if these structures are remnants of a superficial veil or from deeper, dark fibres), violaceous blue to violaceous grey (especially towards margin), sometimes with radiate hygrophanous streaks when young, soon becoming more greyish with a greyish brown centre and sometimes rusty spots, with an altogether ± hygrophanous appearance, mostly with a very thin but often clearly visible frosty veil. Single collections from Mediterranean areas develop also greenish tinges. Lamellae crowded, emarginate or almost adnexed at the stipe, edge initially irregular wavy, first greyish-blue to lavender-blue with a distinctly and remaining violaceous edge, finally ochraceous brown with a tendency to turn rusty brown. Stipe 40–80 × 8–18 mm, with a marginate bulb (up to 40 mm), initially greyish-blue to lavender-blue as the young lamellae and finally discolouring whitish from below, with violaceous blue veil at bulb margin. Context white in pileus and bulb but distinctly violaceous blue in the stipe, soon fading whitish in the stipe and slightly yellowish in the bulb. Odour distinctly earth-like, resembles C. variecolor; also a malt/honey-like component noted, especially in the bulb. Macrochemical reactions: KOH yellow brown or red brown on pileus and in pileus context, somewhat weaker in stipe and bulb. Basidiospores [4, 4, 146], 8.3–9.9 × 4.8–5.6 μm (MV = 9.1 × 5.2 μm); variation of MVs: 8.7–9.3 × 5.1–5.3 μm; Q = 1.56–1.92 (MV = 1.74), brown, distinctly citriform, medium strongly and densely verrucose, warts sometimes isolated but mostly interconnected, clearly visible in the outline of the spores, suprahilar plage sometimes clearly visible. Lamellar edge more or less fertile, with sterile cells scattered to sometimes more abundant (in contradiction to Eyssartier 2004). Pileipellis duplex, cutis thin, consisting of 10–15 layers of 2–4 μm wide hyphae, granular intracellular pigment. Hypoderm well developed, consisting of up to 30 μm wide hyphae with a yellow-brown parietal pigment.
Habitat and distribution: Associated mainly with Fagus sylvatica, but also with Quercus spp., Carpinus and possibly Corylus on calcareous or base rich, loamy soils. Cortinarius viridocoeruleus occurs mainly in temperate deciduous forests and is known from Denmark, France, Germany, Hungary, Italy, and Sweden. Mediterranean-submediterranean collections are only verified from Montpellier-Bédarieux area (Hérault) of S France (most reports from S Europe of C. viridocoeruleus apparently belong to other species).
Comments: As with C. camptoros, many references to C. viridocoeruleus (partly also as “viridicoeruleus” or “viridicaeruleus”) can be found in the literature (e.g. Ballarà et al. 2011, Consiglio et al. 2007, Soop 2017). Unfortunately, these records can most often not be unambiguously interpreted without associated DNA sequence data. However, from our wide selection of sequence-verified collections, we can assemble a precise image of what C. viridocoeruleus is. Majority of the photos and descriptive notes from different localities in Europe converge to a quite uniform and consistent representation of this most characteristic and easily recognizable species in the group. This revised concept is well expressed by Frøslev et al. (2009) in the protologue of C. lepistoides: a medium to large Phlegmacium with a violaceous blue to violaceous grey pileus covered partly by a frost-like, white veil, first greyish-blue to lavender-blue lamellae with a distinctly and persisting violaceous edge and a mostly sharp marginated bulb with violaceous blue veil at margin. According to the priority rules, C. lepistoides is a later synonym of C. viridocoeruleus. Cortinarius lepistoides was published because at that time, the authors thought that the name C. viridocoeruleus applied to the species we here describe as C. viridocaelestinus. The protologue and original description of C. viridocoeruleus (Chevassut & Henry 1975) are, however, somewhat in contradiction to the uniform circumscription of the species as it emerges from our synthesis. The French authors describe the pileus as “vividly uniform green then slightly fulvous to tawny in the centre”. Moreover, the line drawing accompanying the protologue (assumingly of the holotype; loc. cit. Fig. 1) depicts a small, delicate species, with stipe not wider than 5 mm, and with very atypical spores. One can say that colours should not be overestimated in this group and that the basidiocarp size and slenderness may be influenced by local ecological constraints. Consistent with this, almost all collections of this species studied by us are from temperate, continental regions and were growing with Fagus and possibly Carpinus or Corylus, whereas the holotype originates from Quercus ilex Mediterranean woodlands of the Montpellier area. However, one of our sequenced collections was found in such a habitat in Bédarieux, 75 km West from Montpellier, and it did display greenish tinges and a less robust habit when compared to extra-Mediterranean material. There is another explanation though, for these observed contradictions. Chevassut & Henry (1975) indicated that their species was known (by Chevassut) from two sites near Montpellier, so it is possible that their concept was not pure, including both C. viridocoeruleus and one or more morphologically similar Camptori species. Consistent with this hypothesis, Henry (1976)—only one year after C. viridocoeruleus—described C. versicolorum which he distinguished from the former, mostly by less vivid green tinges, concolorous and smooth lamellae edge, and a stouter habit. Most later authors placed these two names in synonymy (Bidaud et al. 2008, Eyssartier & Roux 2011) and Chevassut himself later admitted that C. versicolorum probably represents a simple form of C. viridocoeruleus, if not simply identical (Chevassut 1991). Considering what we now know about the morphological features and distribution of Camptori species in the Mediterranean area, it seems likely that Chevassut had in hands C. viridocoeruleus, C. malavalii, and C. violaceoserrulatus, but it also seems very likely that he and Henry would not have been able to discriminate them with the limited taxonomic tools of the pre-sequencing era. It is remarkable though that their descriptions of C. viridocoeruleus and C. versicolorum collectively include all necessary elements to describe the three aforementioned Mediterranean Camptori. We thus conclude that the two names published in 1975 and 1976 are probably both collective and partly redundant. Regardless of the observed contradictions, the successful sequencing of the holotype of C. viridocoeruleus nomenclaturally links this name to the present species. Unfortunately, original material of C. versicolorum could not be located, preventing us to confidently associate this name to either of the three Mediterranean clades.
One collection belonging to the C. viridocoeruleus cluster (GE02.074, Southwest France, Dordogne; see Fig. 1) was initially identified as C. imperialis Bidaud by Eyssartier (2004), suggesting that this name might have represented a later synonym of C. viridocoeruleus. The description and the photo of this collection indeed show great similarities with other sequenced collections of C. viridocoeruleus, however, the original description of C. imperialis shows less similarities with this species (see pl. 100 in Bidaud et al. 1993). Bidaud placed the species in Sect. Caerulescentes, and furthermore it was described as being associated with Picea and depicted as having amygdaliform to subamygdaliform basidiospores. Therefore, C. imperialis is very unlikely a Camptori species, a conclusion that Frøslev et al. (2009) draw too, when comparing it to C. lepistoides. The sequence of the holotype of C. purpuratus Rob. Henry clusters also with C. viridocoeruleus (Fig. 1). This species was described by Henry (1985) as resembling those of sect. Purpurascentes, lacking the typical purple reaction with Tl4. It is not easy to get an exact impression of C. purpuratus because there are no illustrations of basidiocarps or basidiospores in the protologue. Bidaud et al. (1993) later produced a more complete description of this species, based on a collection ecologically, morphologically and microscopically compatible with C. viridicoeruleus, but unfortunately it was not sequenced. According to the priority rules, C. purpuratus is here included as a later synonym of C. viridocoeruleus. Cortinarius viridocoeruleus differs from the closest species, C. camptoros (Fig. 1), by 6 substitution and indel positions, representing 1% of sequence divergence in the ITS region.
Additional collections examined: Denmark, Jylland, Vosnæs Havskov, 30 Sept 2000, T. Froslev, T.S. Jeppesen, TF2000-056 (C, holotype of C. lepistoides). France, Hérault, Bédarieux, Levas, Quercus ilex, 23 Oct 2013, T.E. Brandrud, TEB985-13 (O); Dordogne, Notre-Dame-de-Sanilhac, route de Phébus, under Carpinus with a few Quercus on calcareous soil, 4 Oct 2002, G. Eyssartier, GE02.074 (PC). Germany, Schleswig-Holstein, Bad Malente-Gremsmühlen, Dieksee, 07 Oct 2004, G. Schmidt-Stohn, SSt04-110 (herb. Schmidt-Stohn); Mecklenburg-Vorpommern, Plau, Ziegeleiteich, 4 Oct 2013, G. Schmidt-Stohn, SSt13-130 (herb. Schmidt-Stohn); Bayern, Main-Spessart, Karlstadt, Karlsburg NW, 4 Oct 1989, H. Marklund et al., CFP907 (TEB 43-89) (S). Eschweiler, Quercus spp., Fagus sylvatica, 3 Oct 2001, B. Oertel, F. Röger, TUB 011408. Hungary, Pest County, Visegrádi Mts, 11 Nov 2003, L. Albert, B. Dima, DB628. Sweden, Öland, Halltorps Hage, 25 Sept 2020, G. Schmidt-Stohn, SSt20-029 (herb. G. Schmidt-Stohn).
Cortinarius violaceoserrulatus, L. Albert, Dima, Schmidt-Stohn, C. Rossi & Ballarà, sp. nov.
Fig. 6a–d

Basidiocarps of the species in the /Camptori clade of the genus Cortinarius (voucher numbers and geographical origin in brackets): a, b, c, d C. marklundii (a SSt13-150 Germany, b JB-9727-18 Spain, c TEB354-14 holotype Norway, d TEB452-17 Norway); e, f, g, h C. violaceoserrulatus (e AL14/441 holotype Hungary, f JMB2011112640 France, g CR6374-2018 Spain, h JB9990-18 Spain); Photos: a G. Schmidt-Stohn; b J. Ballarà; c, d B. Dima; e L. Albert; f J.M. Bellanger; g C. Rossi; h J. Ballarà
MycoBank: MB 843228
Holotype: Hungary, Budapest, Mt Széchenyi-hegy, L. Albert, 23 Nov 2014, AL 14/441 (BP111782); GenBank ON006492 (ITS).
Etymology: the epithet refers to the serrulate/crenulated lamellae edge which is beautifully and persistently violaceus.
Diagnosis: Cortinarius violaceoserrulatus differs from other species in the /Camptori clade from the ochraceous to yellow ochraceous pileus with whitish, thick veil patches, the bluish-lilac serrulate lamellae edge, in combination with paler (greyish) lamellae, and the basidiospores MV ≥ 9.7 × 5.5 μm.
Description: Pileus 40–80 mm, convex then applanate, margin incurved, becoming wavy in mature specimens, viscid, smooth to slightly radially fibrillose, often wrinkled, hygrophanous, especially at the margin, with persistent, almost always with thick whitish veil patches at the centre, entirely pale olive greenish when young, then yellow-ochraceous to ochraceous-yellow and later olivaceous brown, greyish ochre, ochraceous brown, sometimes with darker brownish spots. Lamellae crowded, emarginate to adnate, 4–5 mm wide, edge distinctly irregular crenulate to almost serrulate, pale (bluish) grey when young (in Mediterranean collections conspicuously blue colour were also observed), later greyish brown to finally rusty brown, with persistently violaceous blue edge. Stipe 50–100 × 8–12 mm, cylindrical, with a rounded marginated bulb (up to 20 mm wide), fibrillose to almost smooth towards apex, pale violaceous blue at the apex but sometimes also the whole stipe especially when young, whitish to greyish ochraceous with age. Cortina sparse and fugacious. Basal mycelium white. Context firm, thin, elastic as in other species of the group, whitish, more or less violaceous in the cortex, finally slightly yellowing especially in the bulb cortex. Taste mild, slightly sour, odour faintly earth-like. Macrochemical reactions: KOH brownish on the pileus, ochraceous brown in the context. Basidiospores [5, 5, 225], 8.7–11.1 × 5.1–6.3 μm (MV = 9.9 × 5.7 μm); variation of MVs: 9.7–10.3 × 5.5–5.9 μm; Q = 1.54–1.98 (MV = 1.76), brown, distinctly citriform, finely to medium strongly and densely verrucose, warts isolated but also interconnected, clearly visible in the outline of the spores, suprahilar plage often clearly visible. Pileipellis duplex, epicutis thin, consisting of 2–5 μm wide, more or less hyaline, gelified hyphae, occasionally with some yellow brown (probably cytoplasmatic) pigment. Hypoderm (subcutis) well developed, of subcellular, inflated elements 15–25 μm wide, with yellow, parietal (thick-wall) pigment, and a weak yellowish cytoplasmatic pigmentation.
Habitat and distribution: Widespread, known from various southern European countries (France, Italy, Spain) in thermophilous frondose forests dominated by Quercus ilex, while the type locality is from Hungary, where it grows in warm calcareous sites with Quercus pubescens.
Comments: In the past, C. violaceoserrulatus was mostly included in a wide concept of C. viridocoeruleus. Mainly through molecular genetic analysis, we now realize that there are actually three species hidden in this aggregate. Therefore, it seems obvious that Chevassut had a broader concept of his C. viridocoeruleus, most likely including—besides C. viridocoeruleus s. str.—the newly established species C. violaceoserrulatus and C. malavalii. In his ecological papers, Chevassut mentioned C. viridocoeruleus as one of the frequent phlegmacia in Quercus ilex forests around Montpellier, fruiting together with C. aurilicis and C. quercilicis. This ecology matches C. malavalii and C. violaceoserrulatus much better than C. viridocoeruleus, which is apparently very rare in that region (only one recent find near Bédarieux). Cortinarius violaceoserrulatus is known from France, Hungary, Italy, and Spain, in transition between the more northerly distributed C. viridocoeruleus (with most collections known from Sweden, Denmark and Germany) and the currently only southern (France, Spain) C. malavalii. Cortinarius violaceoserrulatus grows in calcareous, thermophilic forests, an assemblage also stated by Chevassut for his Mediterranean collections of C. viridocoeruleus sensu lato. Therefore, C. violaceoserrulatus is perhaps more frequent in the Montpellier area than C. viridocoeruleus and C. malavalii. With regard to morphology, C. violaceoserrulatus is apparently easily distinguishable from C. viridocoeruleus and from C. malavalii. Almost all collections of C. violaceoserrulatus display distinct white velum patches on the pileus, whereas C. malavalii has no or only traces of such a velum. Generally, C. viridocoeruleus has no such velum patches but more often a distinct, very characteristic frosty whitish velum. Moreover, the colours of the three species are somewhat different. In many collections of C. violaceoserrulatus, ochraceous-yellow to yellow-ochraceous tinges are clearly visible, and generally, the basidiocarps make a rather bright impression whereas C. malavalii is usually more green olivaceous, ochraceous-brown to yellow brown and is generally darker. Only a single collection of C. violaceoserrulatus from the Mediterranean area showed greenish tinges. In contrast to most collections of C. viridocoeruleus—especially those from northern regions in Sweden, Denmark, and Germany—young and fresh C. violaceoserrulatus has vividly violaceous blue or violaceous grey colours in lamellae edges, particularly towards pileus margin, and soon becomes greyish with a greyish brown centre. Cortinarius violaceoserrulatus forms a sister clade to C. viridocaelestinus (Fig. 1), from which it differs by 21 substitution and indel positions, representing 4% of sequence divergence in the ITS region.
Additional collections examined: France, Vaucluse, Cadenet, La Royère, under Q. ilex and Q. pubescens, calcareous soil, 26 Nov 2011, J.-M. Bellanger JMB201112640 (herb. J.-M. Bellanger); Essone, Cheptainville under Fagus sylvatica on calcareous soil, 12 Dec 2011, R. Challange, XC2011-236; Region Ile-de-France, in deciduous forest, 30 Oct 2006, X. Carteret, XC2006-161; Hungary, Budapest, Csillebérc, 13 Nov 2014, L. Albert, B. Dima, DB5689 (herb. ELTE); Italy, Livorno, Piombino, Mediterranean forest, 6 Dec 2014, M. Maletti, Maletti 6-12-2014 (herb. M. Maletti); Spain, Cadiz, Grazalema, Llanos de Rabel, calcareous forest with Quercus ilex f. ballota and Q. faginea, 3 Dec 2018, C. Rossi, CR6374-2018 (herb. C. Rossi, G. Schmidt-Stohn); Berguedà, Montmajor, Comarmada, calcareous Quercus ilex forest with Buxus sempervirens, 11 Nov 2018, J. Ballarà, JB-9990-18 (herb. J. Ballarà, G. Schmidt-Stohn), calcareous Q. ilex and Q. humilis forest, 2 Dec 2019, J. Ballarà, JB-10345-19 (herb. J. Ballarà).
Cortinarius marklundii Brandrud, Dima, Saar, Schmidt-Stohn & Ballarà, sp. nov.
Fig. 6e–h
MycoBank MB 843229
Holotype: Norway, Oppland, Gjøvik, Biri, Eriksrud nature reserve, T.E. Brandrud & B. Dima, 4 Sept 2014, TEB354-14 (O-F-259458), GenBank ON006478 (ITS). Isotype: DB5427 (herb. ELTE).
Etymology: the name is in honour to the late photographer of Cortinarius Flora Photographica, Hans Marklund, Härnösand, Sweden.
Diagnosis: Cortinarius marklundii is characterized by the initially olivaceous (green) grey, more or less hygrophanous pileus, bluish lamellae and stipe, and more or less citriform spores. The species can be distinguished from close taxa based on the soon redbrown discolouring pileus, the lack of a bluish-lilac lamellae edge, and its association with Tilia, Fagus, and Dryas.
Description: Pileus 4–8 cm, (hemi-)spherical, then plano-convex (to broadly umbonate), sometimes irregular, somewhat elastic (like a Lyophyllum sp.), viscid-glutinous, glabrous and glossy when dry, often with a dark, hygrophanous marginal zone or with radiate hygrophanous streaks, mature specimens often become radially wrinkled upon drying; initially olivaceous (greenish)grey, even bluish grey tinged at incurved margin when very young, but this tinge fades instantly (on exposure), soon discolouring more vivid ochraceous redbrown from centre, often bicoloured, ochre brown at centre and hygrophanous dark olive grey at margin, finally fulvous or chestnut brown at centre and paler ochraceous yellow brown towards margin, sometimes with redbrown spots. Often with very thin and fugacious frosty-micaceous veil remnants at centre when young, rarely with silvery white veil patches, sometimes radially silvery-fibrillose towards margin. Lamellae crowded, emarginate to adnexed, edge initially irregular wavy-serrulate to white fimbriate, when very young (deep) greyish violet, soon becoming paler, sometimes almost grey, with paler to whitish edge (edge never persistently bluish), finally ochraceous (-rusty) brown. Stipe 3–6(–7) × 0.8–1.6 cm, with a marginate bulb (up to 3 cm wide), initially pale to distinct greyish violet, especially in upper part, usually soon discolouring to whitish, and finally sometimes with brass brown patches. Veil remnants at bulb margin rather thin but distinct and almost volva-like when very young, soon disappearing, initially greyish white or often with a distinctly but very fugacious bluish tinge, soon tinged ochre yellow. Basal mycelium whitish (or when young with a bluish tinge, which usually disappears instantly on exposure). Context white in pileus and bulb, initially greyish violet in the stipe (most distinct at apex, and towards surface, in places concentrated to hygrophanous spots/streaks), soon fading whitish in the stipe and becoming yellowish tinged elsewhere, even brownish in the base of bulb, rather strongly brownish where damaged by insect larvae. Odour earth-like C. variecolor, and with a component of bread dough, or more of honey/malt-like, especially in the bulb. Macrochemical reactions: KOH somewhat red brown on pileus, brownish in context. Basidiospores [9, 9, 387], 8.4–11.0 × 5.0–6.2 μm (MV = 9.8 × 5.6 μm); variation of MVs: 9.1–10.4 × 5.2–6.0 μm; Q = 1.57–1.89 (MV = 1.73), brown, distinctly citriform, finely to medium strongly and densely verrucose, warts sometimes isolated but mostly interconnected, clearly visible in the outline of the spores, suprahilar plage often clearly visible. Lamellar edge often more or less sterile, with clavate, cylindrical or irregularly shaped sterile cells. Pileipellis duplex, epicutis thin, consisting of 2–5 μm wide, more or less hyaline hyphae. Hypoderm (subcutis) well developed, elements subcellular, 15–30 μm wide, with brown parietal (thick-wall), amber-like pigment, cementing the elements into a compact layer.
Habitat and distribution: Associated with Tilia cordata (and Corylus avellana) in S Scandinavia, with Fagus sylvatica in Central/Western Europe, and with Dryas octopetala in the Spanish Pyrenees. Cortinarius marklundii is known from few, scattered sites in calcareous Fagus forests of Germany and France. Larger, but fragmented populations are found in relict calcareous Tilia woodlands (often in scree areas with limestone/shale gravel) of the Oslofjord-Vänern-Gotland areas of S Scandinavia. Finally, the species is also recorded from calcareous alpine Dryas sites of the Pyrenees.
Comments: Cortinarius marklundii is characterized by the pileus soon discolouring vividly ochre red-brownish, lamellae lacking a (persistently) violaceous blue edge, and being associated with Tilia, Fagus, and Dryas. The bluish-violaceous colours are generally weaker or more fugacious than on the other species in the group; the pileus hardly shows any bluish tinges, and veil remnants at bulb margin rarely show bluish (and then very fugacious) tinges. Cortinarius marklundii was initially included in the wide concept of C. camptoros in Brandrud & Melot (1983). In that paper, the morphology of the type and another collection from Abies-Picea forests of the Black Forest, Germany, were described in detail, as well as that of a collection from calcareous Tilia-Corylus forest of Langesund, outer Oslofjord, Norway. The latter represents C. marklundii. In its revised delimitation, C. camptoros is a Central European, montane species of calcareous Abies-Picea forests, that can be distinguished from C. marklundii by distinct lilac lamellae edges, less warm pileus colours and by a phylogenetic distance of 15 substitution and 4 indel differences representing 3% of sequence divergence in the ITS region (Fig. 1). With its Tilia forest populations in S Scandinavia, C. marklundii is the only species of the /Camptori clade that occurs in N Europe. Cortinarius marklundii was first depicted in Cortinarius, Flora Photographica in 1994 (Brandrud et al. 1990–2018; plate C24; as C. camptoros), with material from a calcareous, Tilia-dominated woodland meadow at Gotland, Sweden. Later, in Brandrud (1999) and in Brandrud & Bendiksen (2001: Fig. 5, as C. camptoros), this species was mentioned as a Tilia-associated variant of the C. camptoros s. str. Subsequently, records of this species were published from a number of calcareous lime forests in Norway, in national reports (see, e.g., Brandrud et al., 2014b, 2016, 2020), always as C. camptoros. It is now recorded from 19 localities in SE Norway, from the Oslofjord area, north to Lake Mjøsa (sequence-verified from 4 collections). In S Sweden, it is known from approx. 20 localities, mainly from Gotland, Öland and near Lake Vänern, Kinnekulle (sequence-verified from 3 localities). In Norway and Sweden, it occurs mainly in calcareous Tilia forests or semi-open woodlands with Tilia, but apparently also in Corylus thickets without Tilia. These S Scandinavian Tilia-Corylus localities seem to host the largest European populations of the species. The calcareous Tilia forests are regarded as old relicts from warm periods (6000–8000 BP) after the ice age where such Tilia forests had a much wider distribution and were much more widespread in temperate Europe (cf. Brandrud 1999). Elsewhere in Europe, the species displays a western distribution, so far confirmed from a few sites in Germany, France, and Spain. In Germany and France, the species is found in rich Fagus forests, in shady sites with more leaf-litter/humus, making the soil more humid than many of the Tilia scree sites. Specimens from Fagus populations may look a little different, with a more pronounced hygrophanous marginal zone, and a less vivid redbrown oxidized, discoloured pileus (centre) (see Fig. 6g), but phylogenetically they are identical. The Spanish localities are alpine Dryas heaths in the eastern Pyrenees. In the Pyrenees, Fagus woodlands often constitute the uppermost subalpine forests, and it is likely that the species also has populations in the Fagus woodlands adjacent to the Dryas sites. In the exposed Dryas sites, the species becomes vividly redbrown-ochraceous brown. The species seems restricted to Europe, but should be expected to occur also further East in Europe, in association with Tilia or Fagus. Cortinarius marklundii forms a sister clade to the C. viridocoeruleus-C.camptoros-C. velicopius clade. The most similar species is C. viridocoeruleus (Fig. 1), from which it differs by 17 substitution and indel positions, representing 3% of sequence divergence in the ITS region.
Additional collections examined: France, Somme, Forêt de Creuse, calcareous broadleaved tree forest with Quercus, Fagus, 22 Oct 2012, X. Carteret, XC2012-150 (herb. X. Carteret); Ain, Valserhône, Mussel, loamy-calcareous Fagus forest, R. Fillion & A. Bidaud, 17 Oct 2011, AB11-10-202 (herb. R. Fillion & A. Bidaud). Germany, Sachsen-Anhalt, Uftrungen, Seeberge, calcareous Fagus forest, 10 Oct 2013, G. Schmidt-Stohn & G. Hensel, SSt13-150 (herb. G. Schmidt-Stohn); Mecklenburg-Vorpommern, Plau am See, Ziegeleiteich, loamy forest with Quercus, Carpinus, Corylus, and Fagus, 16 Oct 2019, G. Schmidt-Stohn, SSt19-145 (herb. G. Schmidt-Stohn); Thüringen, Bad Frankenhausen/Kyffhäuser, Ichstedter Lindenallee, Tilia on base rich ground, 9 Oct 2007, G. Schmidt-Stohn, SSt07-213 (herb. G. Schmidt-Stohn). Norway, Oppland, Gjøvik, Biri, Eriksrud NR, 4 Sept 2014, T.E. Brandrud, B. Dima, TEB358-14 (O), DB 5423 (ELTE); Akershus, Asker, Bjerkås NR (Bjerkås IV), 4 Sept 2017, T.E. Brandrud, B. Dima, TEB452-17 (O), DB6411 (ELTE); Telemark, Porsgrunn, Klevstrand, 9 Oct 2013, T.E. Brandrud, TEB890-13 (O). Spain, Berguedà, Saldes, Serra Encija, Refugi, 10 Sept 2018, J. Ballarà, JB-9736-18 (herb. J. Ballarà); Berguedà, Saldes, Serra Encija, Serrat Voltor, 2 Sept 2018, J. Ballarà, JB-9561-18 (herb. J. Ballarà). Sweden, Gotland, Vallstena, Alvena lindaräng, 29 Sept 1990, H. Marklund et al., CFP1024 / TEB 28-90 (S); Västergötland, Kinnekulle, H. Marklund et al., CFP874 (S); Öland, Ekerum, Halltorps Hage, lövängar with Tilia and Corylus, 8 Sept 2016, G. Schmidt-Stohn, SSt16-083 (herb. G. Schmidt-Stohn).
Cortinarius malavalii Bellanger & Ballarà, sp. nov.
Fig. 7a–c

Basidiocarps of the species in the /Camptori clade of the genus Cortinarius (voucher numbers and geographical origin in brackets): a, b, c C. malavalii (a JMB2012111303 holotype France, b JMB2011112620 France, c JMB 2012111301bis paratype France, not sequenced); d, e, f C. viridocaelestinus (d JA-CUSSTA 9531 holotype Spain, e FA 4768 paratype Spain, f FA 4689 paratype Spain, not sequenced). Photos: a, b, c J.-M. Bellanger; d, e, f F. Armada
MycoBank: MB 843230
Holotype: France, Hérault, Montpellier, CNRS “route de Mende” campus, under Quercus ilex, calcareous soil, 13 Nov 2012, J.-M. Bellanger, P. Geniez, JMB2012111303 (MPU927722), GenBank ON006474 (ITS).
Etymology: Dedicated to the French mycologist Jean-Claude Malaval, who devoted decades of his life to fungal diversity and keeps transmitting his vast knowledge to young naturalists.
Diagnosis: A rather stout member of the /Camptori clade, featuring green-olivaceous tinges on the pileus, contrasting with blue stipe when fresh, greyish to light lilac lamellae with concolorous edges, in Quercus woodlands of the Mediterranean area.
Description: Pileus 30–80 mm, hemispherical to convex, then expanded, often with a low umbo, hygrophanous, radially wrinkled with age, often bicoloured and marbled when fresh, dominated by green olivaceous tinges but distinctly warmer yellow orange towards the centre, dehydrating as more uniformly ochraceous colours upon rapid fading of greenish hues, often with rust brown spots or streaks. Lamellae moderately crowded, sinuate, grey to pale brown with faint lilac hues, edge crenulate, whitish. Stipe 40–75 × 9–18 mm, with conspicuous marginated bulb (15–32 mm), violaceous blue when young or after handling, towards base colouring orange brown. Veil not abundant but breaking into white, silky patches on the pileus surface, persisting in mature basidiocarps. Cortina abundant, whitish. Context firm, rubber-like as other species in the group, whitish in the pileus and stipe centre, blue violaceous in cortex, orange cream at base. Taste mild and odour faint. Macrochemical reactions: KOH reddish on the pileus, ochraceous cream on the context. Basidiospores [4, 4, 166], 8.8–11.2 × 5.4–6.6 μm (MV = 10.0 × 6.0 μm); variation of MVs: 9.4–10.4 × 5.6–6.2 μm; Q = 1.51–1.83 (MV = 1.67), brown, distinctly citriform, finely to medium strongly and densely verrucose, warts sometimes isolated but mostly somewhat interconnected, often clearly visible in the outline of the spores, suprahilar plage often clearly visible. Basidia 4-spored 32–36 × 8–10 μm. Lamellar edge with elements 7–10 μm wide, clavate to cylindrical. Pileipellis duplex, epicutis of gelified 2–5 μm wide, yellowish hyphae with claviform terminal elements. Hypoderm (subcutis) subcellular, of inflated elements 8–26 μm wide with yellow thick walls and a yellowish, intracellular pigmentation. Exsiccata brownish brick.
Habitat and distribution: Mediterranean calcareous Quercus ilex forests. Known from South France and Southeast Spain.
Comments: This species was probably described by Henry as C. versicolorum (Henry 1976). However, this binomial cannot safely be applied to the present taxon because it may also refer to C. violaceoserrulatus (see comments under C. viridocoeruleus). Unfortunately, the type material of C. versicolorum could not be found and sequenced to firmly link it to either of the three species currently represented in Quercus woodlands of the Montpellier area. Cortinarius malavalii differs mostly by greyish lamellae, with only faint lilac hues and concolorous smooth edges from C. violaceoserrulatus, which has conspicuously blue colour on lamellae that persists longer on edges when dehydrating. Based on our limited sampling of C. malavalii, the species also seems slightly stouter than C. violaceoserrulatus and differently coloured, more greenish-olivaceous and with less yellow hues on the pileus. Cortinarius viridocaelestinus grows in the same habitats but is so far only known from mountainous Q. ilex forests of southern Spain. Furthermore, C. viridocaelestinus produces shorter and semi-hypogeous basidiocarps with a combination of very distinctive features that should prevent any confusion: conspicuous and uniform green hues on the pileus, green translucent context contrasting with blue lamellae edges and abundant, thick veil remnants on the pileus. Cortinarius viridocoeruleus is more distantly related to C. malavalii (Fig. 1) but in the Mediterranean area, one collection of C. viridocoeruleus from the Montpellier area (TEB984-13, Fig. 5g) is morphologically very similar to C. malavalii, making confusions possible. Like most Camptori species, the greenish colours of C. malavalii rapidly vanish on drying, and turn ochraceous with brown to rusty streaks, and then the species becomes hard to identify. We believe collections illustrated by Consiglio (1995: 14–16) and Palazón (2001: 454) under the name C. viridicoeruleus may actually represent C. malavalii. Cortinarius malavalii occupies an isolated position in the phylogeny of the /Camptori clade, differing from its most similar relatives, C. marklundii, by 36 substitution and indel positions, and from Cortinarius sp. from USA (JF742658), by 35 substitution and indel positions, representing 6% of sequence divergence in the ITS region.
Additional collections examined: France, Hérault, Montpellier, CNRS “route de Mende” campus, under Quercus ilex, calcareous soil, 17 Nov 2011, J.-M. Bellanger, P. Geniez, JMB2011111701 (herb. J.-M Bellanger); ibid, 13 Nov 2012, JMB2012111303bis (paratype, herb. J.-M Bellanger); ibid, 14 Nov 2012, JMB2012111401 (herb. J.-M Bellanger); Vaucluse, Cadenet, La Royère, under Q. ilex and Q. pubescens, calcareous soil, 26 Nov 2011, J.-M. Bellanger, JMB2011112620 (herb. J.-M Bellanger). Spain, Barcelona, Berguedà, Montmajor, Mare de Deu del Bosc, under Q. ilex, 19 Oct 2014, J. Ballarà, JB-8503-14 (herb. J. Ballarà).
Cortinarius viridocaelestinus Armada & Bellanger, sp. nov.
Fig. 7d–f
MycoBank MB 843231
Holotype: Spain, Andalusia, Granada, Huétor de Santillán, Arroyo Palacios, under Quercus ilex, Q. faginea and a few scattered Pinus halepensis and P. pinaster, 1290–1320 m asl, F. Armada & M.-J. Díaz de Haro, 24 Nov 2018, JA-CUSSTA 9531 (JA), isotype FA4680 (herb. F. Armada), GenBank ON006501 (ITS).
Etymology: From the two Latin adjectives viridis, meaning green and caelestinus, meaning sky-blue and also referring to god’s kingdom.
Diagnosis: A semi-hypogeous member of the /Camptori clade, with green, hyaline hues on pileus and in the context, with initially sky-blue lamellae and a thick, conspicuous white veil.
Description: Pileus 32–50 mm, convex then applanate, often depressed towards the centre in mature specimens, viscid and covered by sticked clumps of earth, smooth to slightly wrinkled, hygrophanous, almost uniform pastel-green to glaucous bottle-green on young basidiocarps, persisting but mixed with brown or grey-brown hues when ageing, with paler margin. Lamellae crowded, adnate to sinuate, 5 mm wide, initially sky-blue then turning pinkish cream on faces when ageing, but the bluish tinges persisting longer on edges that are crenulate. Stipe 26–48 × 10–15 mm, with a rounded marginate bulb (up to 24 mm wide), fibrillose to almost smooth towards top, concolorous to lamellae but rapidly turning green-yellowish from the base. Mycelium white. Veil thick, membranous, white, persisting on the pileus centre as a unique patch or as a few smaller but conspicuous fragments, covering the outer surface and margin of the bulb by a thick white layer. Cortina abundant, whitish, rapidly collapsing along the whole stipe, turning ochraceous to rusty-brown from spores when ageing. Context firm, elastic like other species in the group, bottle-green hyaline in the pileus, bulb and stipe centre, whitish elsewhere, yellowing with age. Taste mild, not farinaceous, odour faint of pastry when cut. Macrochemical reactions: KOH pinkish-brown to foxy-brown on the pileus, foxy-brown on yellowing parts of the context. Basidiospores [1, 1, 45], 9.0–11.0 × 5.5–6.3 μm (MV = 10.0 × 5.9 μm); Q = 1.52–1.88 (MV = 1.70), brown, distinctly citriform, rarely amygdaloid, finely to medium strongly and densely verrucose, warts sometimes isolated but mostly interconnected, clearly visible in the outline of the spores, suprahilar plage often clearly visible. Basidia 4-spored 25–35 × 8.5–13.5 μm. Lamellar edge with marginal elements, 6–15 μm wide, clavate to cylindrical or more inflated to subglobose, shortly septate (2–3 segments), clamped. Pileipellis made of a layer of weakly gelified, 0.5–3 μm wide hyphae, topping poorly differentiated hyphae mixed with wider, up to 7–8 μm wide hyphae, with parietal to finely encrusted yellow pigmentation. Hypoderm elements sausage-like, up to 26 μm wide. Clamps present and numerous everywhere.
Habitat and distribution: So far only known from a single locality in montane Quercus woodlands near Granada, Spain. Late autumn.
Comments: Cortinarius viridocaelestinus is a remarkable species so far only known from two collections from the type locality. Nevertheless, we here describe it as new, because it displays a combination of morphological features that should prevent any confusion with other members of the /Camptori clade. Furthermore, it constitutes a phylogenetically unique clade, sister to C. violaceoserrulatus, from which it differs by 21 substitution and 3 indel positions. In macromorphological terms, the placement in /Camptori is straightforward because of the rubber-like context, the hygrophanous pileus, and the green and blue colours. However, the extent, persistence, and hyaline aspect of green hues on the pileus but also in the context, which is not blue, is unique to this species. The thick and conspicuous white veil and semi-hypogeous fruiting are also remarkable in the clade and should by themselves prevent any misidentification in the field. However, more collections of C. viridocaelestinus will be necessary to appreciate the stability of these spectacular features and better circumscribe its ecological requirements.
Additional collection examined: Spain, Andalusia, Granada, Huétor de Santillán, Arroyo Palacios, under Quercus ilex, Quercus faginea and a few scattered Pinus halepensis and Pinus pinaster, 1290–1320 m asl, 24 Nov 2018, F. Armada & M.-J. Díaz de Haro, FA 4689 (paratype, herb. F. Armada—not sequenced); Parque Natural Sierras de Tejeda, Almijara y Alhama, Játar, Loma del Cura, under Quercus ilex and a few scattered Pinus halepensis and Pinus pinaster with numerous Cistaceae, 1024 m, 6 Dec 2018, F. Armada & M.-J. Díaz de Haro, FA4768 (herb. F. Armada).
Key to species of the /Camptori clade
1 Associated with Abies, probably also Picea or Pinus ............................................................................ C. camptoros
1* Associated with frondose trees such as Quercus, Fagus, Tilia, Corylus ........................................................................ 2
2 Pileus predominantly ochraceous to yellow ochraceous, with ± thick whitish veil patches, MV of spores ≥ 9.7 × 5.5 μm (for Mediterranean coll. see also 5*) ............................................................... C. violaceoserrulatus
2* Pileus more redbrown, ochre brown, chestnut brown or predominantly violaceous blue to violaceous grey especially when fresh ............................................................................. 3
3 Pileus often distinctly innately fibrillose, initially predominantly violaceous blue to violaceous grey especially when fresh and wet, then greyish to greyish brown, often with a thin but clearly visible frosty veil, MV of spores ≤ 9.3 × 5.3 μm ....................................................................... C. viridocoeruleus
3* Pileus redbrown, ochre brown or chestnut brown to glaucous bottle-green, without such a clearly visible frosty veil but possibly with velum patches on pileus.................................... 4
4 Associated with deciduous trees, mainly Fagus, Tilia or also Dryas octopetala (Spain) from temperate regions, pileus glabrous and glossy, not innately fibrillose, first olivaceous grey, soon ochraceous redbrown and finally fulvous to chestnut brown ................................................................... C. marklundii
4* Associated with sclerophyllous Quercus species, so far only known from Mediterranean areas in France and Spain, pileus not glossy, often radially wrinkled or covered by sticked clumps of earth ......................................................... 5
5 Rather stout species with pileus up to 8 cm, pileus with green-olivaceous tinges contrasting with the blue stipe when fresh, pileus later more uniformly ochraceous, often with rust brown spots or streaks, without conspicuous veil remnants .............................................................................. C. malavalii
5* Rather small, semi-hypogeous, species with pileus 3–5 cm, covered by sticked clumps of earth, uniform pastel-green to glaucous bottle-green when young, then brown or grey-brown, with a thick, membranous, white, persisting veil on the pileus centre as a unique patch or smaller but conspicuous fragments ................................................ C. viridocaelestinus
Discussion
In the present study of the /Camptori clade, we describe four species as new in addition to the two already existing ones, C. camptoros and C. viridocoeruleus. All of the new species, except C. viridocaelestinus, have previously been included in the wide concept of C. camptoros or C. viridocoeruleus, respectively. Due to limited and overlapping morphological differentiation between the species, and a rather limited material available, insights from molecular genetic methods have been necessary to resolve the taxonomy, although the amplification and sequencing of the nrDNA ITS region in this group have been particularly difficult. Also, the ecological differentiation combined with molecular data has shown to be important taxonomic character. Although most species are southern and strictly associated to thermophilous, frondose trees, one species deviates from this pattern, being mainly associated with Abies alba in C or S Europe. Our phylogenetic analysis shows that all collections from coniferous trees (Abies alba, Picea abies and possibly also Pinus spp.) fall into one phylogenetic unit, together with the type of C. camptoros Brandrud & Melot. Originally, C. camptoros was treated in a wider concept including C. marklundii, which we show here is a species associated to deciduous trees. Looking at the original labels of the sequenced specimens assigned on morphological identification, the name C. viridocoeruleus appears in three different clades, representing C. viridocoeruleus (assessed by the holotype sequence) and the two new species C. violaceoserrulatus and C. malavalii. Therefore, C. viridocoeruleus has frequently been treated collectively. Our study also shows that C. viridocoeruleus is apparently rarer in S Europe than the other two species. However, at the European scale, C. viridocoeruleus and C. marklundii are apparently the most widespread Camptori species. Both are distributed from SW Europe north to the southernmost Scandinavia. It should be noted that none of the Camptori species known so far does occur in the vast boreal coniferous zone of N Europe. Whereas C. viridocoeruleus is preferentially a temperate Fagus forest species, C. marklundii has its major distribution in calcareous Tilia-Corylus forests of SE Norway and S/C Sweden, but occurs also in Fagus forests of Germany, and in (sub)alpine Dryas heaths of Spain. These Dryas populations might be old relict (remnant) populations from former Fagus forests, and the S Scandinavian populations are apparently old relict site of 6000–7000-year-old Tilia forests. Finally, C. viridocaelestinus is so far known only from the type locality and with its outstanding features, it seems easily distinguishable from all the other species.
However, it must be pointed out that our knowledge of the Camptori clade still remains limited. This study is the first large-scale attempt to disentangle this group. All the six currently known species are rare or very rare, and so we did not have enough collections of each species for more sound conclusions. Therefore, we would not be surprised if further species in this group will show up. But we hope that the present study will be a solid basis for future additions.
Availability of data and material
MycoBank, GenBank
Code availability
Not applicable
References
Ballarà J, Cadiñanos Aguirre JA, Calzada A, Campos JC, Esteve-Raventós F, Fernández Sasia R, Gutiérrez C, Macau N, Mahiques R, Mateos A, Moreno G, Ortega A, Pérez A, Pérez-De-Gregorio MA, Reyes JD, Santamaria N, Suárez E, Vila J (2011) Cortinarius Ibero-insulares 3. Fungi non delineati 58–59:1–236
Bidaud A, Moënne-Loccoz P, Reumaux P, Henry R (1993) Atlas des Cortinaires. Pars V. Éditions Fédération mycologique Dauphiné-Savoie, Marlioz
Bidaud A, Moënne-Loccoz P, Reumaux P, Carteret X (2008) Atlas des Cortinaires. Pars XVII (2). Éditions Fédération mycologique Dauphiné-Savoie, Marlioz, 1234 pp
Borchsenius F (2009) FastGap 1.2. Department of Bio-sciences, Aarhus University, Denmark. Published online at: http://www.aubot.dk/FastGap_home.htm. Accessed 16 Sept 2021
Borovička J, Bušek B, Mikšík M, Dvořák D, Jeppesen TS, Dima B, Albert L, Frøslev TG (2015) Cortinarius prodigiosus—a new species of the subgenus Phlegmacium from Central Europe. Mycol. Progress 14:29
Brandrud TE (1996) Cortinarius subgenus Phlegmacium section Phlegmacium in Europe: Descriptive part. Edinb J Bot 53(3):331–400
Brandrud TE (1999) Cortinarius subgenus Phlegmacium species associated with Tilia cordata (and Corylus avellana) in SE Norway: A relictual element? Journ JEC 1:83–88
Brandrud TE, Bendiksen E (2001) The Cortinarius species of calciphilous Tilia-Corylus and Quercus-Corylus woodlands of Fennoscandia, outposts of the temperate Fagus and Quercus-Carpinus forest types of C. Europe. Journ JEC 1:105–113
Brandrud TE, Melot J (1983) Cortinarius camptoros et Cortinarius patibilis, deux nouveaux Phlegmaciums des forêts de la montagne. Bull Soc Mycol France 99(2):220–237
Brandrud TE, Lindström H, Marklund H, Melot J, Muskos S (1990–2018) Cortinarius, Flora Photograpzica, vol 1–5 (English version), Härnösand
Brandrud TE, Bellù F, Frøslev TG, Schmidt-Stohn G (2013) Cortinarius subgenus Phlegmacium section Claricolores and the story about Cortinarius blattoi Mazza. Journ JEC 15:14–30
Brandrud TE, Dima B, Schmidt-Stohn G, Bellù F, Frøslev TG, Oertel B, Saar G, Soop K (2014a) Cortinarius subgenus Phlegmacium Sektion Multiformes in Europe. Journ JEC 16:162–199
Brandrud TE, Evju M, Skarpaas O (2014b) Nasjonal overvåking av kalklindeskog og kalklindeskogsopper. Beskrivelse av overvåkingsopplegg fra ARKO-prosjektet. NINA rapport 1057, pp. 37
Brandrud TE, Evju M, Blaalid R, Skarpaas O (2016) Nasjonal overvåking av kalklindeskog og kalklindeskogsopper. Resultat fra første overvåkingsomløp 2013−2015. - NINA Rapport 1297, pp. 128
Brandrud TE, Schmidt-Stohn G, Liimatainen K, Niskanen T, Frøslev TG, Soop K, Bojantchev D, Kytövuori I, Jeppesen TS, Bellù F, Saar G, Oertel B, Ali T, Thines M, Dima B (2018) Cortinarius sect. Riederi: taxonomy and phylogeny of the new section with Europaen and North American distribution. Mycol Progress 17:1323–1354
Brandrud TE, Brandrud MK, Dima B (2020) Nasjonal overvåking av kalklindeskog og kalklindeskogsopper. Resultater fra andre overvåkingsomløp, første år (2019). NINA rapport 1793. Norsk institutt for naturforskning
Chevassut G (1991) Cortinaires. Catalogue écologique des champignons supérieurs méditerranéens. Annales Féd Assoc Mycol Méditer 6:72
Chevassut G, Henry R (1975) Six Cortinaires méditerranéens nouveaux du chêne vert (Quercus ilex) dans le Bas-Languedoc (1). Doc Mycol 5(20):23–36
Clémençon H (1972) Zwei verbesserte Präparierlösungen für die mikroskopische Untersuchung von Pilzen. Z Pilzk 38:49–53
Consiglio G (1995) Contributo alla conoscenza dei macromiceti della regione Emilia-Romagna. Prima parte. Genere Cortinarius. Rev Micol 38(1):3–16
Consiglio G, Antonini D, Antonini M (2007) Sulla variabilità di Cortinarius viridicaeruleus. Journ JEC 9:84–94
Dima B, Lindström H, Liimatainen K, Olson A, Soop K, Kytövuori I, Dahlberg A, Niskanen T (2016) Typification of Friesian names in Cortinarius sections Anomali, Spilomei and Bolares, and description of two new species from northern Europe. Mycol Prog 15(9):903–919
Erb B, Matheis W (1983) Pilzmikroskopie. Kosmos, Gesellschaft der Naturfreunde, Fränkische Verlagshandlung, Stuttgart
Eyssartier G (2004) Notes sur quelques espèces de Cortinaires et Psathyrelles rares ou nouvelles. Fungi non delineati 28:1–55
Eyssartier G, Roux P (2011) Guide des champignons. Belin, Paris, 734 pp
Fernández Sasia R, Cadiñanos Aguirre JA (2000) Aportación al conocimiento de los Cortinarius en el norte de la península ibérica (I). Doc Mycol 30(119):63–73
Fernández-Brime S, Vila J, Ortega A (2014) Some new and interesting taxa of Cortinarius subgenus Phlegmacium from the European Mediterranean Basin. Mycologia 106(3):491–504
Frøslev TG (2007) Systematics of Cortinarius with special emphasis on section Calochroi. PhD thesis, University of Copenhagen
Frøslev TG, Jeppesen TS (2009) Three new species of Cortinarius subgenus Phlegmacium. Mycotaxon 106:469–477
Frøslev TG, Matheny PB, Hibbett DS (2005) Lower level relationships in the mushroom genus Cortinarius (Basidiomycota, Agaricales): a comparison of RPB1, RPB2 and ITS phylogenies. Mol Phyl Evol 37(2):602–618
Frøslev TG, Jeppesen TS, Læssøe T, Kjøller R (2007) Molecular phylogenetics and delimitation of species in Cortinarius section Calochroi (Basidiomycota, Agaricales) in Europe. Mol Phyl Evol 44:217–227
Frøslev TG, Kjøller R, Bruun HH, Ejrnæs R, Hansen AJ, Læssøe T, Heilmann-Clausen J (2019) Man against machine: Do fungal fruitbodies and eDNA give similar biodiversity assessments across broad environmental gradients? Biological Conservation 233:201–212
Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes—application to the identification of mycorrhizae and rusts. Mol Ecol 2:113–118
Garnica S, Weiß M, Oertel B, Oberwinkler F (2003) Phylogenetic relationships of European Phlegmacium species (Cortinarius, Agaricales). Mycologia 95(6):1155–1170
Garnica S, Weiß M, Oertel B, Oberwinkler F (2005) A framework for a phylogenetic classification in the genus Cortinarius (Basidiomycota, Agaricales) derived from morphological and molecular data. Can J Bot 83:1457–1477
Garnica S, Münzmay T, Saar G (2006) Cortinarius Subsektion Elegantiores in Europa. Journ JEC 8:79–101
Garnica S, Weiß M, Oertel B, Ammirati J, Oberwinkler F (2009) Phylogenetic relationships in Cortinarius, section Calochroi, inferred from nuclear DNA sequences. BMC Evol Biol 9:1
Garnica S, Schön ME, Abarenkov K, Riess K, Liimatainen K, Niskanen T, Dima B, Soop K, Frøslev TG, Jeppesen TS, Peintner U, Kuhnert-Finkernagel R, Brandrud TE, Saar G, Oertel B, Ammirati JF (2016) Determining threshold values for barcoding fungi: lessons from Cortinarius (Basidiomycota), a highly diverse and widespread ectomycorrhizal genus. FEMS Microbiol Ecol 92(4):fiw045
Gouy M, Guindon S, Gascuel O (2010) SeaView version 4: a multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol Biol Evol 27:221–224
Henry R (1976) Les Cortinaires purpurescents. Doc Mycol 7(25):25–55
Henry R (1985) Nouvelle étude de Cortinaires. Bull Soc Mycol Fr 101(1):1–54
Jeppesen TS, Frøslev TG, Brandrud TE (2012) Subgen. Phlegmacium (Fr.) Trog. In: Knudsen H, Vesterholt J (eds) Funga Nordica, 2nd revised edition. Agaricoid, boletoid, clavarioid, cyphelloid and gastroid genera. Nordsvamp, pp 782–826
Katoh K, Standley DM (2013) MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol 30:772–780
Krieglsteiner GJ, Gminder A (eds) (2010) Die Großpilze BadenWürttembergs. Band 5, Ständerpilze: Blätterpilze III. Ulmer, Stuttgart
Kumar S, Stecher G, Tamura K (2016) MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol Biol Evol 33(7):1870–1874
Liimatainen K, Niskanen T, Dima B, Kytövuori I, Ammirati JF, Frøslev TG (2014) The largest type study of Agaricales species to date: bringing identification and nomenclature of Phlegmacium (Cortinarius) into the DNA era. Persoonia 33:98–140
Liimatainen K, Kim JT, Pokorny L, Kirk PM, Dentinger B, Niskanen T (2022) Taming the beast: a revised classification of Cortinariaceae based on genomic data. Fungal Diversity. https://doi.org/10.1007/s13225-022-00499-9
Moser M (1961) Die Gattung Phlegmacium (Schleimköpfe). Die Pilze Mitteleuropas, Band IV. Bad Heilbrunn Obb., Julius Klinkhardt
Moser M (1967) Die Röhrlinge und Blätterpilze. In: Gams H, Kleine Kryptogamenflora 3 Aufl. 2 (b/2). Stuttgart
Nagy LG, Kocsubé S, Csanádi Z, Kovács GM, Petkovits T, Cs V, Papp T (2012) Re-mind the gap! Insertion—deletion data reveal neglected phylogenetic potential of the nuclear ribosomal internal transcribed spacer (ITS) of Fungi. PLoS ONE 7(11):e49794
Palazón F (2001) Setas para todos. Pirineos. Península Ibérica. Huesca, Editorial Pirineo, 653 pp
Peintner U, Moncalvo JM, Vilgalys R (2004) Toward a better understanding of the infrageneric relationships in Cortinarius. Mycologia 96(5):1042–1058
Ronquist F, Huelsenbeck JP (2003) MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19(12):1572–1574
Saar G, Brandrud TE, Dima B, Schmidt-Stohn G, Bellù F, Frøslev T, Oertel B, Soop K (2014) Cortinarius Untergattung Phlegmacium Sektion Purpurascentes in Europa. Journ JEC 16:140–161
Schmidt-Stohn G (2011) Fotografie ornamentierter Sporen mit der Methode des “Focus-Stacking”. Journ JEC 13:79–87
Schmidt-Stohn G, Saar G, Soop K, Brandrud TE, Dima B (2020) Cortinarius subgenus Phlegmacium subsection Varii in Europe. Journ JEC 22:27–49
Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, Levesque CA, Chen W and Fungal Barcoding Consortium (2012) Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc Natl Acad Sci USA 109:6241–6246
Silvestro D, Michalak I (2012) RaxmlGUI: a graphical front-end for RAxML. Org Divers Evol 12:335–337
Simmons MP, Ochoterena H, Carr TG (2001) Incorporation, relative homoplasy, and effect of gap characters in sequence-based phylogenetic analysis. Syst Biol 50:454–462
Soop K (2017) Cortinarius in Sweden, 15th edn. Éditions Scientrix
Soop K, Dima B, Cooper JA, Park D, Oertel B (2019a) A phylogenetic approach to a global supraspecific taxonomy of Cortinarius (Agaricales) with an emphasis on the southern mycota. Persoonia 42:261–290
Soop K, Schmidt-Stohn G, Dima B, Saar G, Brandrud TE (2019b) Cortinarius subgenus Phlegmacium section Aureocistophili in Europe. Journ JEC 21:33–54
Stamatakis A (2014) RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30(9):1312–1313
Stensrud Ø, Orr RJS, Reier-Røberg K, Schumacher T, Høiland K (2014) Phylogenetic relationships in Cortinarius with focus on North European species. Karstenia 54:57–71
Taschen E, Sauve M, Taudiere A, Parlade J, Selosse M, Richard F (2015) Whose truffle is this? Distribution patterns of ECM fungal diversity in Tuber melanosporum brûlés developed in multi-host Mediterranean plant communities. Environ Microbiol 17(8):2747–2761
White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols: a guide to methods and applications. Academic Press, New York, pp 315–322
Acknowledgements
We are very grateful to the Journées européennes du Cortinaire (J.E.C.), especially to president Oswald Rohner for the financial help of the sequencing work. We also like to thank André Bidaud (Vézeronces-Curtin, France), Xavier Carteret (Chaville, France), Francisco Javier Donaire Sánchez (Herbario JA-CUSSTA, Granada, Spain), Gunnar Hensel (Merseburg, Germany), Thomas Stjernegaard Jeppesen (Copenhagen, Denmark), Jacques Melot (Paris, France), Marco Maletti (Pesaro, Italy), Claudio Rossi (Casteldarne-Chienes, Italy), and Uwe Winkler (Konstanz, Germany) for providing exsiccata and/or photos. We are grateful to Kare Liimatainen and Tuula Niskanen (both Jodrell Laboratory, Royal Botanic Garden, Kew, United Kingdom) for providing us important sequences.
The work of Bálint Dima was supported by the ELTE Thematic Excellence Programme 2020 (TKP2020-IKA-05) financed by the National Research, Development and Innovation Office, Hungary.
Funding
Open access funding provided by Eötvös Loránd University. ELTE Thematic Excellence Programme 2020 (TKP2020-IKA-05) supported by the National Research, Development and Innovation Office.
The Journées européennes du Cortinaire (J.E.C.) funded most of the sequencing work.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design as well as to material preparation, data collection, and discussions. Molecular lab work was performed by B. Dima, J.M. Bellanger, and T.G. Frøslev. Phylogenetic analyses were performed by B. Dima. Macroscopic analyses were done by all authors. Microscopic analyses and statistics were done by G. Schmidt-Stohn. The manuscript was written by G. Schmidt-Stohn. J.M. Bellanger, T.E. Brandrud, and B. Dima. All authors commented on the manuscript as well as read and approved the final version.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Section Editor: Zhu-Liang Yang
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(FAS 43 kb)
Rights and permissions
This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schmidt-Stohn, G., Brandrud, T.E., Bellanger, JM. et al. Taxonomy and phylogeny of the phlegmacioid clade Camptori (Cortinarius s.l., Basidiomycota) in Europe with description of four new species. Mycol Progress 21, 55 (2022). https://doi.org/10.1007/s11557-022-01804-1
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11557-022-01804-1