
Abstract
A model for the mathematical study of immune response to breast cancer is proposed and studied, both analytically and numerically. It is a simplification of a complex one, recently introduced by two of the present authors. It serves for a compact study of the dynamical role in cancer promotion of a relatively recently described subgroup of regulatory B cells, which are evoked by the tumour.













Similar content being viewed by others
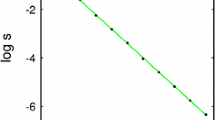

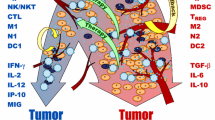
References
Abbas AK, Lichtman AH, Pillai S (2014) Cellular and molecular immunology E-book. Elsevier Health Sciences, Amsterdam
Aguirre-Ghiso JA (2007) Models, mechanisms and clinical evidence for cancer dormancy. Nat Rev Cancer 7(11):834–846. https://doi.org/10.1038/nrc2256
Al-Tuwairqi SM, Al-Johani NO, Simbawa EA (2020) Modeling dynamics of cancer virotherapy with immune response. Adv Differ Equ 1:1–26. https://doi.org/10.1186/s13662-020-02893-6
Biragyn A, Lee-Chang C, Bodogai M (2014) Generation and identification of tumor-evoked regulatory B cells. Methods Mol Biol (Clifton, NJ) 1190:271–289. https://doi.org/10.1007/978-1-4939-1161-5_19
Bitsouni V, Tsilidis V (2022) Mathematical modeling of tumor-immune system interactions: the effect of rituximab on breast cancer immune response. J Theor Biol 539, 111001. https://doi.org/10.1016/j.jtbi.2021.111001
Bray F, Ferlay J, Soerjomataram I et al (2018) Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J Clin 68(6):394–424. https://doi.org/10.3322/caac.21492
Bunimovich-Mendrazitsky S, Shochat E, Stone L (2007) Mathematical model of BCG immunotherapy in superficial bladder cancer. Bull Math Biol 69(6):1847–1870. https://doi.org/10.1007/s11538-007-9195-z
Byrne HM, Cox SM, Kelly C (2004) Macrophage-tumour interactions: in vivo dynamics. Discrete Contin Dyn Syst B 4(1):81. https://doi.org/10.3934/dcdsb.2004.4.81
Carol D, Rebecca S, Priti B et al (2014) Breast cancer statistics, 2013. CA A Cancer J Clin 64(1):52–62. https://doi.org/10.3322/caac.21203
Chraa D, Naim A, Olive D et al (2019) T lymphocyte subsets in cancer immunity: friends or foes. J Leukoc Biol 105(2):243–255. https://doi.org/10.1002/JLB.MR0318-097R
de Pillis LG, Radunskaya AE (2003) A mathematical model of immune response to tumor invasion. In: Computational fluid and solid mechanics 2003. Elsevier, pp 1661–1668, https://doi.org/10.1016/B978-008044046-0.50404-8
de Pillis LG, Renee Fister K, Gu W et al (2009) Mathematical model creation for cancer chemo-immunotherapy. Comput Math Methods Med 10(3):165–184. https://doi.org/10.1080/17486700802216301
de Pillis LG, Caldwell T, Sarapata E et al (2013) Mathematical modeling of regulatory T cell effects on renal cell carcinoma treatment. Discrete Contin Dyn Syst B 18:915–943. https://doi.org/10.3934/dcdsb.2013.18.915
den Breems NY, Eftimie R (2016) The re-polarisation of M2 and M1 macrophages and its role on cancer outcomes. J Theor Biol 390:23–39. https://doi.org/10.1016/j.jtbi.2015.10.034
Eftimie R, Bramson JL, Earn DJ (2011) Interactions between the immune system and cancer: a brief review of non-spatial mathematical models. Bull Math Biol 73(1):2–32. https://doi.org/10.1007/s11538-010-9526-3
Fentiman IS, Fourquet A, Hortobagyi GN (2006) Male breast cancer. The Lancet 367(9510):595–604. https://doi.org/10.1016/S0140-6736(06)68226-3
Ferlay J, Colombet M, Soerjomataram I et al (2019) Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int J Cancer 144(8):1941–1953. https://doi.org/10.1002/ijc.31937
Ghosh S, Banerjee S (2018) Mathematical modeling of cancer-immune system, considering the role of antibodies. Theory Biosci 137(1):67–78. https://doi.org/10.1007/s12064-018-0261-x
Gonzalez H, Hagerling C, Werb Z (2018) Roles of the immune system in cancer: from tumor initiation to metastatic progression. Genes Dev 32(19–20):1267–1284. https://doi.org/10.1101/gad.314617.118
Guo FF, Cui JW (2019) The role of tumor-infiltrating B cells in tumor immunity. J Oncol 2019:1–9. https://doi.org/10.1155/2019/2592419
He DH, Xu JX (2017) A mathematical model of pancreatic cancer with two kinds of treatments. J Biol Syst 25(01):83–104. https://doi.org/10.1142/S021833901750005X
Jordan D, Smith P, Smith P (2007) Nonlinear ordinary differential equations: an introduction for scientists and engineers. Oxford University Press, Oxford
Keohane E, Smith L, Walenga J (2015) Rodak’s Hematology - E-Book: Clinical Principles and Applications. Elsevier Health Sciences, Amsterdam
Khailaie S, Bahrami F, Janahmadi M et al (2013) A mathematical model of immune activation with a unified self-nonself concept. Front Immunol 4:474. https://doi.org/10.3389/fimmu.2013.00474
Kuznetsov VA, Makalkin IA, Taylor MA et al (1994) Nonlinear dynamics of immunogenic tumors: parameter estimation and global bifurcation analysis. Bull Math Biol 56(2):295–321. https://doi.org/10.1007/BF02460644
López AG, Seoane JM, Sanjuán MA (2014) A validated mathematical model of tumor growth including tumor-host interaction, cell-mediated immune response and chemotherapy. Bull Math Biol 76(11):2884–2906. https://doi.org/10.1007/s11538-014-0037-5
Mabarrack NHE, Turner NL, Mayrhofer G (2008) Recent thymic origin, differentiation, and turnover of regulatory T cells. J Leukoc Biol 84(5):1287–1297. https://doi.org/10.1189/jlb.0308201
Makhlouf AM, El-Shennawy L, Elkaranshawy HA (2020) Mathematical modelling for the role of CD4+ T cells in tumor-immune interactions. Comput Math Methods Med. https://doi.org/10.1155/2020/7187602
Nani F, Freedman H (2000) A mathematical model of cancer treatment by immunotherapy. Math Biosci 163(2):159–199. https://doi.org/10.1016/S0025-5564(99)00058-9
Narendra BL, Reddy KE, Shantikumar S et al (2013) Immune system: a double-edged sword in cancer. Inflamm Res 62(9):823–834. https://doi.org/10.1007/s00011-013-0645-9
Olkhanud PB, Baatar D, Bodogai M et al (2009) Breast cancer lung metastasis requires expression of chemokine receptor CCR4 and regulatory T cells. Can Res 69(14):5996–6004. https://doi.org/10.1158/0008-5472.CAN-08-4619
Olkhanud PB, Damdinsuren B, Bodogai M et al (2011) Tumor-evoked regulatory B cells promote breast cancer metastasis by converting resting CD4+ T cells to T-regulatory cells. Can Res 71(10):3505–3515. https://doi.org/10.1158/0008-5472.CAN-10-4316
Pang H, Yu Q, Guo B et al (2013) Frequency of regulatory T-cells in the peripheral blood of patients with pulmonary tuberculosis from Shanxi Province, China. PLoS ONE 8(6):65496. https://doi.org/10.1371/journal.pone.0065496
Perko L (2006) Differential equations and dynamical systems, 3rd edn. Berlin, Springer
Phan TA, Tian JP (2017) The role of the innate immune system in oncolytic virotherapy. Comput Math Methods Med. https://doi.org/10.1155/2017/6587258
Rackauckas C, Nie Q (2017) Differentialequations.jl—a performant and feature-rich ecosystem for solving differential equations in Julia. J Open Res Softw 5(1):15. https://doi.org/10.5334/jors.151
Rosser EC, Mauri C (2015) Regulatory B cells: origin, phenotype, and function. Immunity 42(4):607–612. https://doi.org/10.1016/j.immuni.2015.04.005
Segel L, Cohen I (2001) Design principles for the immune system and other distributed autonomous systems. Oxford University Press, Santa Fe Institute Studies on the Sciences of Complexity
Senekal NS, Mahasa KJ, Eladdadi A et al (2021) Natural killer cells recruitment in oncolytic virotherapy: a mathematical model. Bull Math Biol 83(7):1–51. https://doi.org/10.1007/s11538-021-00903-6
Sopik V, Narod SA (2018) The relationship between tumour size, nodal status and distant metastases: on the origins of breast cancer. Breast Cancer Res Treat 170(3):647–656. https://doi.org/10.1007/s10549-018-4796-9
Starr C, Taggart R, Evers C (2012) Biology: the unity and diversity of life. Cengage Learning
Szymańska Z (2003) Analysis of immunotherapy models in the context of cancer dynamics. Int J Appl Math Comput Sci 3(13):407–418
Vaghi C, Rodallec A, Fanciullino R et al (2020) Population modeling of tumor growth curves and the reduced gompertz model improve prediction of the age of experimental tumors. PLoS Comput Biol 16(2):1–24. https://doi.org/10.1371/journal.pcbi.1007178
Villasana M, Radunskaya A (2003) A delay differential equation model for tumor growth. J Math Biol 47(3):270–294. https://doi.org/10.1007/s00285-003-0211-0
Zhang Y, Wallace DL, De Lara CM et al (2007) In vivo kinetics of human natural killer cells: the effects of ageing and acute and chronic viral infection. Immunology 121(2):258–265
Acknowledgements
The authors would like to thank the reviewers for their thoughtful comments and efforts towards improving the present manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendices
Appendix
Appendix A Preliminary Results
In this section, we prove that IVP (we remind that this acronym is used for the initial value problem \(\left\{ (1),(2)\right\} \)) has a unique solution, which is non-negative for non-negative initial conditions and for positive parameter values. We also prove that our solution is global, i.e. it does not explode for some finite positive value of time t. The above conditions are necessary in order to assure that IVP yields biologically realistic results.
Proposition 14
(Uniqueness and non-negativity) For every \(\left( T_0,N_0,R_0,B_0\right) \in \mathbb {R}_{\ge 0}^4\), IVP has a unique (local) solution \(\left( T, N, R, B\right) :\left[ 0,\tau \right) \rightarrow \mathbb {R}_{\ge 0}^4\) for some \(\tau >0\).
Proof
It is easy to see that the conditions of the Picard-Lindelöf theorem are fulfilled, since every function of the right-hand side of system (1) is continuous, just like its partial derivative with respect to every variable. Thus, we have that there exists a unique solution to IVP. In fact, we can extended the solution and consider it in the maximal non-negative interval of existence.
Next, we prove the non-negativity of the solution. Rewriting (1a) in the following form
we notice that we have a Bernoulli equation for the variable T, and thus its solution is
which is non-negative, if \(T_0\) is non-negative.
Using the fact that \(\sigma > 0\), we turn our attention to (1b). We have that
and using Grönwall’s inequality, we have that
which means that \(N(t)\ge 0\), when \(N_0\ge 0\).
Using a similar method as above, from (1c) we get
which means that \(R(t)\ge 0\), when \(R_0\ge 0\).
We then use the separation of variables method to solve (1d) for the variable B. Its solution is
Clearly, if \(B_0\ge 0\), then \(B(t)\ge 0\). \(\square \)
Proposition 15
(Boundedness of T) The set \(\left[ 0,1/b\right] \) is positively invariant for the component T of the solution of IVP.
Proof
Since the solution of IVP is non-negative, we have from (1a) that
We assume the following two initial value problems:
Assuming that \(T_0 \le z(0) = 1/b \), and since \(q_1, q_2\) are Lipschitz functions on \(\mathbb {R}\) that satisfy the inequality \( q_1(T) \le q_2(T)\), from the comparison theorem we have that \(T(t) \le z(t)\) for t in the maximal non-negative interval of existence of the solution of IVP. Solving initial value problem (A3), yields \(z = 1/b\). Hence, \(T(t) \le 1/b\), with the assumption that the initial value of T is smaller or equal to 1/b. \(\square \)
Proposition 16
(Globality) If \(T_0 \le 1/b\), then the solution of IVP is global.
Proof
From (1d) and using the fact that \(T(t) \le 1/b\), we have that
and by using Grönwall’s inequality, we have that
Moreover, from (1c) and using the fact that \(\theta _RR \ge 0 \) we get
and by using Grönwall’s inequality, we have that
Finally, from (1b) and using the fact that \( \gamma RN \ge 0 \) we get
and by using Grönwall’s inequality, we have that
Since the solution is bounded on any compact non-negative interval, we deduce its (positive) globality. \(\square \)
Appendix B Parameter Estimation
Here we explain our reasoning behind our choice of parameters. Most of the parameters in our model have been chosen based on methods and data that can also be found in Bitsouni and Tsilidis (2022).
1.1 Appendix B.1 The Tumour
Based on the data fitting experiments conducted in Bitsouni and Tsilidis (2022), we chose the logistic function to model the breast cancer growth, with the tumour growth rate being \(a = 0.15\) day\(^{-1}\) and the inverse of the tumour carrying capacity being \(b = 1 \cdot 10^{-9}\) cell\(^{-1}\). The cell lines used for the estimation of these parameters are CN34BrM, MDA-231 and SUM1315.
1.2 Appendix B.2 The NK Cells
Healthy young adults have a total NK production rate of (15 ± 7.6) \(\cdot 10^6\) \(\text {cell} \cdot \text {litre}^{-1}\) \(\cdot \, \mathrm {day}^{-1}\), while healthy older adults have one of (7.3 ± 3.7) \(\cdot 10^6\) \(\text {cells} \cdot \text {litre}^{-1}\) \(\cdot \,\mathrm {day}^{-1}\) (Zhang et al. 2007). Since the average amount of blood in the human body is about 5 litre (Starr et al. 2012), the constant source of NK cells is in the range
The half-life of NK cells in humans is 1 to 2 weeks (Zhang et al. 2007) which, assuming exponential decay of NK cells, yields a range for \(\theta _N\) of \(({\frac{\ln 2}{14}}, {\frac{\ln 2}{7}})=(0.049, 0.099)\). Here, we choose an NK cells half-life of 11 days with a corresponding programmable NK death rate of
Approximately 4 to 29% of circulating lymphocytes are NK cells (Keohane et al. 2015). The average number of lymphocytes per microlitre is 1000 to 4800 cells (Abbas et al. 2014), and since the average human has an average of 5 litres of blood, we have that the total population of lymphocytes in a human is 5\(\cdot \)10\(^9\) to 24\(\cdot \)10\(^9\) cells. Therefore, the total population of NK cells in blood is \(N_{min}=\) 2\(\cdot \)10\(^8\) to \(N_{max}=\) 6.96\(\cdot \)10\(^9\) cells. At the healthy equilibrium, our model suggests the population of NK cells to be \(\frac{\sigma \theta _R}{\gamma \kappa +\theta _N \theta _R}\), which means that
Replacing the minimum value of \(\sigma \), and the maximum value of \(\theta _N\) and \(\kappa \) in the above inequality yields the minimum value of \(\gamma \), while replacing the maximum value of \(\sigma \), and the minimum value of \(\theta _N\) and \(\kappa \) (we derive a range for \(\kappa \) in Appendix B.3), yields the maximum value of \(\gamma \). Finally, after calculating the above two quantities, the resulting range for the parameter \(\gamma \) is
1.3 Appendix B.3 The Tregs
For the constant source of Tregs, our model suggests that a healthy organism has an average number of \( \kappa /\theta _R \) Tregs, since this is the coordinate which corresponds to Tregs in the healthy equilibrium. From Pang et al. (2013) we get that Tregs are 5 to 10% of the total CD4\(^+\) T cells population circulating in blood, while from Abbas et al. (2014) we get that the percentage of CD4\(^+\) T cells among the total population of circulating lymphocytes ranges from 50 to 60%. Hence, the percentage of Tregs among the total population of circulating lymphocytes is 2.5 to 6%. The average number of lymphocytes per microlitre is 1000 to 4800 cells (Abbas et al. 2014), and since the average human has an average of 5 litres of blood, we have that the total population of lymphocytes in a human is 5\(\cdot \)10\(^9\) to 24\(\cdot \)10\(^9\) cells. Therefore, the total population of Tregs in blood is 1.25\(\cdot \)10\(^8\) to 1.44\(\cdot \)10\(^9\) cells. Solving the equation
for \(\kappa \) and replacing the range of values for the total population of Tregs as found above and the value of \(\theta _R\) as found in the following paragraph, we finally get the constant source of Tregs to be in the interval
The half-life of Tregs is found to be about 18 days (Mabarrack et al. 2008). Thus, assuming Tregs follow exponential decay we have that
The rate of NK cell death due to Tregs, \(\gamma \), is assumed to be
1.4 Appendix B.4 The tBregs
As B cells are less studied than T cells and NK cells, we are not able to find the half-life of Bregs, let alone tBregs, as they are recently discovered. So, we estimate the rate of programmable tBreg cell death to be around \(\theta _B = 0.4 \text { day}^{-1}\).
The same holds for the rate of breast-cancer-induced tBreg activation, \(m_T\), which we assume it to be either \(m_T = 5.2 \cdot 10^{-15}\) cell\(^{-1}\) \(\cdot \) day\(^{-1}\) or \(m_T = 5 \cdot 10^{-10}\) cell\(^{-1}\) \(\cdot \) day\(^{-1}\).
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Bitsouni, V., Gialelis, N. & Tsilidis, V. A Mathematical Study of the Role of tBregs in Breast Cancer. Bull Math Biol 84, 112 (2022). https://doi.org/10.1007/s11538-022-01054-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11538-022-01054-y
Keywords
- Cancer immunology
- Tumour-evoked regulatory B cell
- Mathematical modelling
- Stability analysis
- Bifurcation analysis
- Numerical simulation