
Abstract
Purpose
In life cycle assessment (LCA), eutrophication is commonly assessed using site-generic characterisation factors, despite being a site-dependent environmental impact. The purpose of this study was to improve the environmental relevance of marine eutrophication impact assessment in LCA, particularly regarding the impact assessment of waterborne nutrient emissions from Swedish agriculture.
Methods
Characterisation factors were derived using site-dependent data on nutrient transport for all agricultural soils in Sweden, divided into 968 catchment areas, and considering the Baltic Sea, the receiving marine compartment, as both nitrogen- and phosphorus-limited. These new characterisation factors were then applied to waterborne nutrient emissions from typical grass ley and spring barley cultivation in all catchments.
Results and discussion
The site-dependent marine eutrophication characterisation factors obtained for nutrient leaching from soils varied between 0.056 and 0.986 kg Neq/kg N and between 0 and 7.23 kg Neq/kg P among sites in Sweden. On applying the new characterisation factors to spring barley and grass ley cultivation at different sites in Sweden, the total marine eutrophication impact from waterborne nutrient emissions for these crops varied by up to two orders of magnitude between sites. This variation shows that site plays an important role in determining the actual impact of an emission, which means that site-dependent impact assessment could provide valuable information to life cycle assessments and increase the relevance of LCA as a tool for assessment of product-related eutrophication impacts.
Conclusions
Characterisation factors for marine eutrophication impact assessment at high spatial resolution, considering both the site-dependent fate of eutrophying compounds and specific nutrient limitations in the recipient waterbody, were developed for waterborne nutrient emissions from agriculture in Sweden. Application of the characterisation factors revealed variations in calculated impacts between sites in Sweden, highlighting the importance of spatial differentiation of characterisation modelling within the scale of the impact.
Similar content being viewed by others


Explore related subjects
Find the latest articles, discoveries, and news in related topics.Avoid common mistakes on your manuscript.
1 Introduction
Life cycle assessment (LCA) was originally developed as a site-independent tool, where the environments affected by the assessed impacts represent average or generic recipients (Potting and Hauschild 2006). For all impact categories where the impact is dependent on the activity location, spatial differentiation can be highly important to achieve representative assessment of the environmental impacts of a system, and failure to take spatial variation into account may give misleading results (Azevedo et al. 2013; Potting and Hauschild 2006). Spatial differentiation is therefore considered an important step to improve the LCA methodology (Hellweg and Milà i Canals 2014), and many recent publications explore different aspects of this topic (e.g. Anton et al. 2014; Cosme et al. 2015; Cosme and Hauschild 2017; Mutel et al. 2012; Scherer and Pfister 2015).
In recent years, LCA has emerged as a leading instrument for evaluating the climate impact of agricultural products and guiding policy in this field, for example in EU biofuel policy (Directive 2009/28/EC). However, for good policy advice on agriculture, climate impacts should not be studied in isolation, but in combination with other environmental impacts. Among these, eutrophication is a critical environmental impact for agricultural products, due to the high contribution of agriculture to eutrophication pressure. Eutrophication is one of the impact categories most frequently assessed in LCA and is also one of the impact categories representing a regional environmental impact, which means that site-differentiated life cycle impact assessment (LCIA) modelling may be required for accurate representation of impacts.
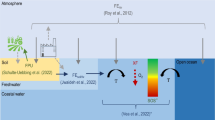
Eutrophication refers to ecosystem response to input of excess nutrients. Aquatic eutrophication causes excessive growth of algae, with secondary effects such as oxygen depletion and fish death, ultimately altering the ecosystem balance in the aquatic system (EC-JRC 2010). The main nutrients responsible for aquatic eutrophication are nitrogen (N) and phosphorus (P), and the fate and effect of these nutrients are site-dependent (Finnveden and Potting 1999). When N and P are applied to the soil as fertiliser, some is taken up by the crop, but some leaves the field as gaseous losses, or via erosion, surface run-off or leaching through the soil profile, eventually reaching a waterbody. However, a substantial proportion of the leached nutrients is removed from the subsequent water flow path by hydrological, chemical and biological processes; the combined effect of which is referred to as retention (Morrice et al. 1997). Previous studies have shown that retention varies considerably between catchments (Hejzlar et al. 2009; Tysmans et al. 2013). This indicates that the relevant scale for eutrophication assessment in LCA may not be country level.
In LCA, the connection between the emission and the indicator result is represented by a characterisation factor (CF) specific for the emitted substance and, in the case of site-dependent LCIA, also specific for the emission location. Despite the site dependency, eutrophication effects in LCA are often accounted for using a site-generic method which places the indicator at the point of emission, thus neglecting the fate of the nutrients after they leave the field and assuming that all leached N and P can cause eutrophication. This is commonly referred to as the CML method, after Guinée (2002). Some later LCA methodologies include fate in their derived eutrophication CFs, using models with continental or global coverage, e.g. ReCiPe 2008 (Struijs et al. 2013) and EDIP 2003 (Hauschild and Potting 2005). Major projects working in this direction (IMPACT World+ (http://www.impactworldplus.org) and LC-IMPACT (http://www.lc-impact.eu/)) are also currently underway. Parts of the marine eutrophication CFs for LC-IMPACT have been published, and they indicate considerable global variability in endpoint CFs (Cosme and Hauschild 2016; Cosme and Hauschild 2017; Cosme et al. 2015). The recently published ReCiPe 2017 methodology does not provide a characterisation model for marine eutrophication, due to lack of endpoint model (Huijbregts et al. 2016). Some other studies have focused on deriving CFs for a specific area, such as Finland (Seppälä et al. 2004); Brittany, France (Basset-Mens et al. 2006) and Galicia, Spain (Gallego et al. 2010). Their results indicate that site-dependent eutrophication assessment on a finer scale than that on country level would add valuable information to LCA, but those studies used data of variable quality and/or were limited in scope in terms of geography and nutrients. Seppälä et al. (2004) rely on expert judgement for determination of fate factors, Basset-Mens et al. (2006) only derive fate factors for nitrate, and Gallego et al. (2010) derive transport factors using data that was not specific for the region. Moreover, both Seppälä et al. (2004) and Gallego et al. (2010) derived one set of site-dependent CFs for the whole region, rather than several sets of CFs which could be used at different sites within the region.
When moving towards higher spatial resolution of marine eutrophication impact assessment, it becomes increasingly important to consider marine areas with characteristics deviating from the average conditions. One of these sites is the Baltic Sea, which is the world’s largest brackish ecosystem (Swedish EPA 2006) and severely affected by eutrophication (HELCOM 2009). Unlike most other marine environments, which are generally considered N-limited, the Baltic Sea is limited by both N and P, with variations among the sub-basins. Most of the Swedish rivers drain into the Baltic Sea and the retention mechanisms for leached nutrients in Sweden are affected by inland waters, which cover 12.7% of the country’s area (Eurostat 2016). The main sources of N and P to the sea basins surrounding Sweden are municipal wastewater treatment plants and agriculture (Brandt et al. 2009). In the present study, spatially differentiated characterisation factors for waterborne N and P emissions from agriculture were derived for Sweden.
The aim of this study was to increase the environmental relevance of marine eutrophication impact assessment in LCA, which was achieved through:
-
Developing CFs with high spatial resolution that can be used for site-dependent marine eutrophication impact assessment of agricultural nutrient losses from Swedish soils, with a focus on fate modelling;
-
Comparing these CFs with the currently available CFs and examining the implications of using CFs derived at different scales.
2 Methods
The purpose of the LCIA phase of an LCA is to convert inventory data (emissions and resource consumptions) into potential impacts expressed as a common category indicator, by applying substance-specific CFs for each elementary flow (Guinée 2002). This can be described as follows:
where Impact C is the impact on environmental impact category C of the emissions, emission inventory i,j is the amount of substance i crossing the system boundary between the technosphere and the environment at site j and CF i,j is the site-dependent CF for substance i emitted at site j.
In this study, the system boundary between the technosphere and the environment was set at the perimeter of the field and at plant root depth, as this is the limit at which nutrients can no longer be taken up by the crop they were intended to fertilise. The CFs were derived according to the characterisation model described in the following sections.
2.1 Characterisation model
The characterisation model presented in this study derives midpoint CFs (kg Neq/kg) for marine eutrophication, with focus on fate modelling. The following equation based on the adapted version of Seppälä et al. (2004) from Gallego et al. (2010) was used:
where transport factor i,j (−) describes the transport of substance i from the point of emission j to a marine recipient, nutrient limitation factor i,j (−) the sensitivity to substance i in the marine recipient corresponding to the point of emission j, and equivalency ratio i (kg Neq/kg) the algae growth potential of substance i. The marine recipient is given by the location of the point of emission.
Recent midpoint LCIA methods for emission-related impacts are designed using the concept CF = fate × exposure × effect (Pennington et al. 2004). A different terminology was chosen for this study, mainly due to the focus on substance transport from point of emission to recipient, and disregard of substance fate and effect in the recipient. However, the components of the CFs presented in this study can be related to this framework. The retention on the substance pathway, here represented by the transport factor, could be complemented by a component representing residence time in the recipient, to constitute a fate factor. This approach is adopted by Cosme and Hauschild (2017), who use watershed export fractions as a component of site-dependent marine eutrophication fate factors. The combination of our nutrient limitation factor and equivalency ratio could be seen as an exposure factor (following the terminology of e.g. Cosme et al. 2015), although it lacks a component to describe the recipients’ degree of sensitivity, but rather assumes that all nutrients that reach a recipient sensitive to that nutrient will cause eutrophication.
2.1.1 Transport factors
The transport factor (−) describes the proportion of leached nutrients that reaches a marine environment, and is calculated as follows:
where removal fraction i,j (−) is the fraction of emitted substance i removed from the substance flow by e.g. denitrification, organism uptake, sorption to soil particles or sedimentation in wetlands, lakes and streams. A direct emission to a marine environment will have a transport factor of 1, and emissions to soil, rivers and lakes will typically have a lower transport factor.
The removal fractions for the present study were obtained from Swedish Environmental Emissions Data (SMED 2015c), which in turn were derived to calculate and report Sweden’s nutrient loads to surrounding seas within HELCOMFootnote 1’s Fifth Pollution Load Compilation (PLC-5) (Brandt et al. 2009). The data consist of retention factors, defined as the fraction of the nutrients leaving the field that will not enter a marine environment (Brandt et al. 2009), and were interpreted as the removal fraction in the present study. The retention data were derived using the HBV-NP model for nutrient loads in Sweden, and the full modelling procedure is described in Brandt et al. (2009). The HBV-NP model aggregates point source emissions and diffuse sources from the area and simulates the N and P fractions’ transport in freshwater, using both process models and measured data. The retention mechanisms included in the model are denitrification, algae uptake, mineralisation, sedimentation and soil and groundwater retention. Other model parameters include weather data, hydrological data and free parameters used to calibrate the simulated nutrient concentrations to measured data. The calculation of diffuse nutrient loads from agriculture used in the derivation of the PLC-5 data is described in Johnsson et al. (2008).
The PLC-5 data are based on the definition that a waterborne nutrient emission occurs when the nutrients leave the root zone by either surface run-off or leaching through the soil profile (Johnsson et al. 2008), which means that the retention calculated in Brandt et al. (2009) is in accordance with the chosen system boundary. Retention factors are provided, for N and P separately, for 1093 catchment areas covering the whole of Sweden in the target year 2006, but flow-corrected for 1985–2004 to decrease the influence of weather fluctuations (Brandt et al. 2009). In the present study, the dataset was corrected to include only catchment areas which had land used for agricultural purposes in 2005 according to data from SMED (2015b), so that 968 catchments remained.
2.1.2 Nutrient limitation factors
Nutrient limitation is a fundamental concept in plant growth. Its significance is that plant growth in natural ecosystems generally is limited by one nutrient, since the plant requires nutrients in specific ratios. This means that addition of a limiting nutrient to a particular ecosystem promotes biomass growth, while addition of any other nutrient causes no detectable change (Smith et al. 1999).
In general, P is considered to limit primary production in freshwater systems and N is considered limiting in marine ecosystems. However, the limiting nutrient in brackish waters, such as the Baltic Sea, has been debated. Therefore, a literature review was performed to determine the dominant opinion on limiting nutrients in marine environments surrounding Sweden. A number of different data sources, all using different approaches for determining nutrient limitations in one or more of the sub-basins of the Baltic Sea, were reviewed. These included six research papers, four scientific reports and two reports from HELCOM. In the case of the HELCOM reports, the limiting nutrient was interpreted from the nutrient reduction targets set for different sub-basins of the Baltic Sea (see Fig. 1). The outcome of the literature review is presented in Table 1.

Overview of the marine sub-basins surrounding Sweden
Following the results of the literature review, the nutrient limitation factor was set to 1 for all N inputs to all sub-basins of the Baltic Sea. The reason for considering N as limiting even for the Bothnian Bay, which is unanimously referred to as P-limited in the literature included in the review, is that there is a net N flow from the Bothnian Bay to the Bothnian Sea (Savchuk 2005). For P, the limiting nutrient factor was set to 0 for inputs to Öresund and Kattegat, and 1 for all other sub-basins. In contrast to the N input to the Bothnian Bay, P in Öresund and Kattegat is less prone to be transported to a P-limited water body. This is because the net flow of P from Kattegat to the Baltic Sea is negative (Savchuk 2005) and the P limitation of Skagerrak primarily refers to estuarine areas (Swedish EPA 2008).
As the conclusions on nutrient limitations in the Baltic Sea sub-basins were uncertain, a sensitivity analysis was performed in which the nutrient limitation factor for all sub-basins surrounding Sweden was set to 1 for both N and P (Table 1). Since marine environments are generally considered strictly N-limited in other methodologies, a second sensitivity analysis was performed by setting the nutrient limitation factor for all sub-basins to 1 for N and 0 for P (Table 1).
2.1.3 Equivalency ratio
The equivalency ratio describes the potential for a nutrient to increase primary production if it reaches an environment where it is limiting for growth. In the basic characterisation models for eutrophication, such as CML 2001 (Guinée 2002), the CF is equal to the equivalency ratio. In the present study, the equivalency ratio was derived using the Redfield molar ratio (N:P = 16:1; Redfield 1958), which describes the nutrient composition of algae, and thereby the theoretical algae growth potential of different substances. In accordance with the ReCiPe method (Struijs et al. 2013), the category metric chosen was kg Neq. This resulted in equivalency ratios of 1 kg Neq/kg N and 7.23 kg Neq/kg P. The category metric can be recalculated into terms of a P compound or an N compound using the Redfield ratio, or oxygen demand for decomposition of biomass (138 moles of O2 per mole of algae biomass) (Guinée 2002).
2.2 Comparison with currently available characterisation factors
Globally or regionally weighted average CFs are commonly provided to allow for application of the method when spatial information on emissions is missing. In this study, a weighted country average CF was calculated and used for comparison with other LCIA methods. The local CFs derived were weighted according to the amount of agricultural land used in the particular area in 2005, based on data from SMED (2015b).
The purpose of the comparison was to indicate the quantitative level of agreement between the new CFs and commonly used midpoint CFs for marine eutrophication. Three established eutrophication methods stated to be applicable for emissions to soil were chosen for comparison: CML 2001 (Guinée 2002), EDIP 2003 (Hauschild and Potting 2005) and ReCiPe 2008 (Struijs et al. 2013). CML was chosen because it is still widely used, EDIP because it provides country-specific CFs and ReCiPe because it was recently recommended as the best available LCIA methodology for aquatic eutrophication (Hauschild et al. 2013). The emerging impact assessment methodology LC-IMPACT (http://www.lc-impact.eu/) would also be an appropriate candidate for comparison due to its comprehensiveness, but it was excluded from the quantitative comparison since the marine eutrophication characterisation factors are not published in full yet. Instead, it is discussed qualitatively in the Discussion section. Marine eutrophication CFs for emissions to soil are currently not available for IMPACT World+ (http://www.impactworldplus.org), and comparison was therefore not possible.
The CF for eutrophication in the CML 2001 baseline methodology (Guinée 2002) is equal to the equivalency ratio. CML 2001 does not discriminate between terrestrial, marine and freshwater eutrophication, so these values represent the total potential eutrophication impacts on all recipients. For EDIP 2003, the CF for Sweden was used. EDIP 2003 recommends not including P in the environmental impact category marine eutrophication (Hauschild and Potting 2005), but since a CF for P to marine environments is included in the EDIP 2003 report, it was included here for comparison. For ReCiPe 2008, the site-generic CF for fertiliser was used. Since the ReCiPe 2008 CF is developed to be applied to gross input of nutrients to the soil, it was corrected according to the correction factors provided in Struijs et al. (2013) to comply with the system boundary. The ReCiPe 2008 eutrophication methodology includes nutrient fate and considers marine recipients as strictly N-limited.
2.3 Relative importance of N and P
To estimate the relative contribution of N and P to the marine eutrophication impact of typical Swedish crop cultivation, the CFs were applied to realistic values on nutrient leaching from spring barley and grass ley. This was done using leaching data from Appendices 2 and 4 in Johnsson et al. (2008), which list average leaching per crop and field characteristics for 22 agricultural regions in Sweden. These were combined with data on soil type, phosphorus content and field inclination for 8559 sub-catchments, corresponding to sub-areas of the catchment areas for which transport factors were derived, and encompassing all parts of Sweden with current crop production (SMED 2015a). The obtained emission inventories for each sub-catchment were then multiplied by the site-dependent CFs following Eq. (1).
3 Results
3.1 Calculated characterisation factors
The calculated midpoint CFs represent the transport of waterborne nutrients from point of emission to coast and the limiting nutrient in the recipient ecosystem. The site-dependent CFs covering all of Sweden are presented in Fig. 2 for N emissions and in Fig. 3 for P emissions. These CFs varied between 0.056 and 0.986 kg Neq/kg N and between 0 and 7.23 kg Neq/kg P (all values presented in Online Resource 1). The CFs were generally higher in areas closer to the coast, but there were several deviations from this pattern. The CFs for P also tended to be higher in the north and were zero for the catchments that drain into Öresund and Kattegat, since the nutrient limitation factor for P is zero in these areas.

Site-dependent characterisation factors for waterborne N emissions from soil in Sweden. The unit is kg Neq at the recipient per kg N emitted at the source. Source Land use layers: Lantmäteriet, Creative Commons License

Site-dependent characterisation factors for waterborne P emissions from soil in Sweden. The unit is kg Neq at the recipient per kg P emitted at the source. Source Land use layers: Lantmäteriet, Creative Commons License
The resulting area-weighted CF for Sweden and the corresponding CFs from CML 2001, EDIP 2003 and ReCiPe 2008 are presented in Table 2. As can be seen, the CFs for N and P were lower than the CML values. The CF for N was similar to the EDIP and ReCiPe values, but the CF for P was several-fold higher than the EDIP CF.
3.2 Relative importance of N and P from crop cultivation
The marine eutrophication impacts calculated for 8559 sub-catchments in Sweden varied between 1.2 and 58 kg Neq/ha for spring barley and between 0.31 and 32 kg Neq/ha for grass ley (Fig. 4). Area-weighted total average was 21 kg Neq/ha for spring barley and 7.5 kg Neq/ha for grass ley. Nitrogen was the nutrient that accounted for the largest eutrophication impact (82% in spring barley cultivation and 75% in grass ley cultivation) under the simplified assumption that the same amount of land was cultivated in each sub-catchment. However, P had a larger impact than N in 2 and 15% of the sub-catchment for spring barley and grass ley, respectively.

Marine eutrophication impact of waterborne emission from spring barley (a) and grass ley (b) cultivation in Sweden
3.3 Sensitivity analysis of nutrient limitation
The sensitivity analysis showed that assuming N and P limitation in all sub-basins had a minor impact on the results, increasing calculated average marine eutrophication impact from spring barley and grass ley production by less than 10% (Table 3). Assuming only N limitation in all sub-basins reduced the calculated marine eutrophication impact by up to 25%.
4 Discussion
The results presented in this study highlight several issues concerning the implementation of spatially differentiated eutrophication assessment. The CFs with high spatial differentiation obtained in the analysis showed variation between sites within Sweden, providing an opportunity for gaining new insights on the impact behaviour of nutrients compared with using a generic or country average CF.
4.1 Spatial variation of characterisation factors
The variation in CFs within Sweden (Figs. 2 and 3) shows that using the country average as a site-dependent measure can mask potential differences in impacts from nutrient emissions at different sites. This means that country average CFs may give misleading LCIA results for site-dependent LCAs, supporting the claim by Bockstaller et al. (2008) that local impacts should ideally not be aggregated into higher scales than the actual impact.
A study on eutrophication CFs for airborne emissions within Sweden concluded that spatial differentiation did not add much variability (Finnveden and Nilsson 2005), so there may be a discrepancy in appropriate scale for airborne and waterborne nutrient emissions. However, Finnveden and Nilsson (2005) point out that distinguishing between environmental regions would be better than between countries. In the current study, the transport factor varied between 0.06 and 1 for N, and 0.007 and 1 for P, indicating that the transport pathway between the point of emission and the marine recipient is an important source of spatial variability in the CFs and therefore should not be neglected when deriving spatially differentiated fate factors. This was also highlighted by Basset-Mens et al. (2006) for nitrate transport in Brittany, France. Cosme and Hauschild (2017) used watershed export fractions with a spatial resolution of 5772 watersheds globally to model endpoint and damage CFs, and found soil-related fate factors to be responsible for most of the spatial differentiation of their damage model results. The scope of previous studies on site-dependent aquatic eutrophication assessment (e.g. Seppälä et al. (2004); Basset-Mens et al. (2006); Gallego et al. (2010) do not allow for direct comparison with the CFs obtained in the present study, but they generally support the conclusion that even country-specific CFs miss some of the local variation. Alternative spatial resolution for site-dependent eutrophication LCIA based on hydrological processes has been proposed by e.g. Basset-Mens et al. (2006) and Nitschelm et al. (2016). This is also in line with the approach in the EU Water Framework Directive (Directive 2000/60/EC), which considers water management at the river basin scale.
Identifying a relevant scale for LCIA models is not primarily about using the highest resolution, since factors such as uncertainty and possibility to interpret the results should also be considered (Mutel et al. 2012). The variability found within catchments in this study indicates that there are significant spatial variations even before the nutrients reach the marine recipient. This means that high-resolution fate factors can add valuable information to the assessment and that the appropriate scale for marine eutrophication impact assessment is likely to be small.
4.2 Comparison with other methodologies
The weighted average CFs were lower than the CML 2001 CFs, which was to be expected since the eutrophication impacts calculated by the CML method represent the maximum potential eutrophication impact on all recipients, including freshwater and terrestrial ecosystems. While the weighted average CF for N agreed well with the EDIP 2003 and ReCiPe CFs (Table 2), the average CF for P did not. A possible reason for this is that EDIP 2003 defines the inventory as the amount of nutrients available for leaving the topsoil after plant uptake and binding (Hauschild and Potting 2005), whereas we defined it as the nutrients leaving the root zone. This means that the system boundary between the technosphere and the environment was defined differently in the studies. The nutrients retained in the root zone would then be included in the inventory following EDIP 2003, while according to our definition, they would not be considered an emission until they had actually left the topsoil. This could make a significant difference, since P, unlike N, binds strongly to soil particles and therefore can accumulate in the topsoil (Hooda et al. 2001). Another factor possibly contributing to the discrepancy in CF values is that the model used for EDIP 2003 assumes fixed removal rates in soil and freshwater (Hauschild and Potting 2005). Fixed removal rates are based on the assumption that the fate of P is directly correlated with fertiliser application and that it is equal for all soils and freshwater systems. In reality, P leaching and retention are highly variable and depend on factors such as soil properties, soil management and previous P loads (Bergström et al. 2015). In contrast, the retention data for our transport factors are calibrated against field measurements (Brandt et al. 2009), so it appears that Swedish conditions deviate from the generic removal rates assumed for EDIP 2003 CFs. This highlights the importance of considering local characteristics when deriving regional CFs and somewhat contradicts use of generic models to derive regional CFs, since some local factors which may have a significant effect on the actual impact are difficult to capture in these models.
The combination of this study’s nutrient limitation factor and equivalency ratio (i.e. 1 kg Neq/kg N) can be compared with the exposure factors presented by (Cosme et al. 2015), which for the Baltic Sea equates to 0.807 kg Neq/kg N (recalculated using the Redfield ratio to match our units). Even though this comparison shows that our assumed exposure factor gives a similar result as the model by Cosme et al. (2015) which includes more processes, it should be noted that the Baltic Sea has the highest exposure factors among LMEs (Cosme et al. 2015), and thus, this assumption may be less valid for other LMEs (the lowest exposure factor equated to approximately 0.023). However, we want to highlight the fact that the results in this study show that it may be important to also include P limitation in marine eutrophication assessment and that differentiation in the marine environment on a smaller spatial scale than LMEs may be called for.
4.3 Applications and implications
Accurate impact assessment is becoming more important as the role of LCA in environmental assessment of agricultural products increases, and the potential of LCA as a decision support tool for farmers has been highlighted (Bartl et al. 2012; Fedele et al. 2014). As demonstrated by the variability in CFs in Figs. 2 and 3, higher spatial resolution should improve the correlation between LCIA results and actual impacts.
Eutrophication assessment in LCA using CFs with high spatial resolution would be especially informative when systems that include cultivation at different sites are compared, as in e.g. Korsaeth et al. (2014) and Lehuger et al. (2009), or for determining how to optimise the geographic placement or distribution of agricultural activities to minimise environmental impacts, as proposed by Nitschelm et al. (2016) and Page et al. (2014). Several case studies, e.g. Bessou et al. (2013) and Korsaeth et al. (2014), have attempted to spatialise LCA of crop production by using local emissions data, but are not using site-dependent impact CFs for eutrophication. Doing this may give a false impression of spatial differentiation, while in fact, significant spatial differences in environmental impacts are omitted. This indicates that site-dependent eutrophication CFs such as those presented in this study would be useful, but must then be available for the specific region. In addition, spatialised LCAs require systematic ways to handle the large amount of data in order to be a feasible approach for LCA practitioners, but methods to facilitate the application of regionalised impact assessment have been demonstrated (e.g. Mutel and Hellweg 2009; Mutel et al. 2012). There are also cases where CFs for a larger region, such as country or global scale, would be more suitable than regional CFs, e.g. LCAs of an average product or process from the region. It would then result in more accurate impact assessments if CFs at finer spatial scale were aggregated to represent average CFs at appropriate scale for the assessment, instead of CFs derived by site-generic methods. When these aggregated CFs are used, it should be made clear that they represent an average impact in the region, rather than representing the impact at all sites in the region.
On a global level, the displacement of N and P has been cited as one of the planetary boundaries where the safe operating space has been exceeded (Steffen et al. 2015), so there is a need for appropriate representation of these effects in environmental assessment tools such as LCA. For Sweden, climate change and lifestyle changes are expected to increase nutrient loads to the Baltic Sea (Hägg et al. 2014), giving further reason for action and increasing the need to prioritise between load-reducing measures. As better modelling of the consequences of different actions on watershed quality can help support policy improvements (Merot et al. 2009), recognising spatial variations in the relationship between nutrient loads and eutrophication effect can help improve the effectiveness of eutrophication-reducing measures and policies (Tysmans et al. 2013). For example, spatial differentiation was necessary to source-appoint the nutrient loads to the Baltic Sea in PLC-5 (Brandt et al. 2009). This spatial differentiation should also be considered when LCAs are used to connect the environmental pressure of agricultural products to the production location, as in e.g. Röös et al. (2011). In this context, our analysis of the relative importance of N and P highlights the applicability for single-site assessments when several eutrophying substances are involved, which is often the case for crop production where both N and P are emitted. Our analysis showed that spatialised CFs for eutrophication assessment could help identifying which nutrients to target when investing in measures that reduce waterborne nutrient emissions. Judging from the results, the eutrophication pressure on the seas surrounding Sweden is higher from N pollution than that from P pollution from spring barley and grass ley cultivation at most sites in Sweden.
4.4 Sensitivity of nutrient limitation factor
The sensitivity analysis (Table 3) showed that varying the assumed limiting nutrient in the Baltic Sea had a moderate effect on CFs and an even smaller effect on the average potential impacts of cultivation. This is because the calculated impact of N generally was larger than that of P for the crop cultivation situations assessed here. Although the sensitivity to P limitation was of minor importance for the country average impacts in the cases tested here, assumptions made on the limiting nutrient may still be an important factor for emissions in the specific areas where P impacts were more prominent than N impacts (2 and 15% of the sub-catchments for grass ley and spring barley, respectively) and in cases where the N:P ratio of the emissions are lower than for the average cultivation situations studied here.
For practical reasons, sub-basin resolution is not always a viable alternative for LCIA. However, nutrient limitation at the same level as the spatial differentiation of the CFs should be considered when regionalised LCIA methods are being developed. In the case of waterborne emissions from Sweden to the Baltic Sea, the results presented in this study (Table 3) suggest that assuming the entire Baltic Sea as N- and P-limited would give more accurate results than assuming only N limitation. Although N seems to be the primary limiting nutrient in most marine ecosystems, the Baltic Sea is not the only exception. Other studies report marine environments where both N and P limitation occurs and thus where P impacts could be important to include in marine eutrophication LCIA. These cases include tidal systems (Barba-Brioso et al. 2010) and estuaries such as Chesapeake Bay (Fisher et al. 1999) and the rias in Galicia, Spain (Gallego et al. 2010).
5 Conclusions and recommendations
Site-dependent CFs for marine eutrophication in Sweden, with a focus on fate modelling, were derived using data from national nutrient load reporting and consideration of site-specific nutrient limitation. The calculated CFs varied between 0.056 and 0.986 kg Neq/kg N and between 0 and 7.23 kg Neq/kg P among the 968 catchment areas included in the study. The spatial variation found among CFs indicates that high spatial resolution is needed when making site-dependent assessments of eutrophication impacts. The Swedish average of these site-dependent CFs compared well with CFs from existing methodologies that include fate for N, but not for P, where the Swedish average was substantially larger. However, calculated impacts from crop cultivation varied by up to two orders of magnitude between sites, which suggests that variation in impacts between regions may be substantial, and country averages therefore may not be appropriate for site-dependent eutrophication LCIA. Thus, we recommend that local characteristics are considered when deriving spatially differentiated CFs and that geographical resolution relevant to soil and water, which can be significantly smaller than country average, is applied when these CFs are presented.
The ongoing evolution of LCA from a site-generic towards site-dependent or even site-specific tool provides a more environmentally relevant basis for decision support, and opens up possibilities for LCA to answer new questions. This transformation requires LCIA methods that can support impact assessment on relevant spatial levels. For marine eutrophication impact assessment, this evolution has gone from the site-generic methods, via continental and country resolution, and towards currently emerging methods that apply spatial resolutions with more relevance to the scale of the impact. The spatial variation of substance transport presented in this study reinforces the motivation to continue the ongoing development of spatial differentiation in marine eutrophication assessment, and provides new insights on appropriate level of geographical resolution.
Notes
The Helsinki Commission (HELCOM, http://www.helcom.fi/) is the governing body for the multilateral Convention on the Protection of the Marine Environment of the Baltic Sea Area.
References
Andersson A, Hajdu S, Haecky P, Kuparinen J, Wikner J (1996) Succession and growth limitation of phytoplankton in the Gulf of Bothnia (Baltic Sea). Mar Biol 126:791–801
Anton A, Torrellas M, Nunez M, Sevigne E, Amores MJ, Munoz P, Montero JI (2014) Improvement of agricultural life cycle assessment studies through spatial differentiation and new impact categories: case study on greenhouse tomato production. Environ Sci Technol 48:9454–9462
Azevedo LB, Henderson AD, van Zelm R, Jolliet O, Huijbregts MAJ (2013) Assessing the importance of spatial variability versus model choices in life cycle impact assessment: the case of freshwater eutrophication in Europe. Environ Sci Technol 47:13565–13570
Barba-Brioso C, Quaranta G, Galan E, Fernandez-Caliani JC, Miras A (2010) The life cycle impact assessment applied to the Domingo Rubio tidal system by the study of seasonal variations of the aquatic eutrophication potential. Sci Total Environ 408:5897–5902
Bartl K, Verones F, Hellweg S (2012) Life cycle assessment based evaluation of regional impacts from agricultural production at the Peruvian coast. Environ Sci Technol 46:9872–9880
Basset-Mens C, Anibar L, Durand P, van der Werf HMG (2006) Spatialised fate factors for nitrate in catchments: modelling approach and implication for LCA results. Sci Total Environ 367:367–382
Bergström L et al (2015) Turnover and losses of phosphorus in Swedish agricultural soils: long-term changes, leaching trends, and mitigation measures. J Environ Qual 44:512–523
Bernes C (2005) Change beneath the surface: an in-depth look at Sweden’s marine environment. Swedish Environmental Protection Agency, Stockholm
Bessou C, Lehuger S, Gabrielle B, Mary B (2013) Using a crop model to account for the effects of local factors on the LCA of sugar beet ethanol in Picardy region, France. Int J Life Cycle Assess 18:24–36
Bockstaller C, Guichard L, Makowski D, Aveline A, Girardin P, Plantureux S (2008) Agri-environmental indicators to assess cropping and farming systems. A review. Agron Sustain Dev 28:139–149
Brandt M, Ejhed H, Rapp L (2009) Nutrient loads to the Swedish marine environment in 2006: Sweden’s report for HELCOM’s fifth pollution load compilation. Swedish Environmental Protection Agency, Stockholm
Cosme N, Hauschild MZ (2016) Effect factors for marine eutrophication in LCIA based on species sensitivity to hypoxia. Ecol Indic 69:453–462
Cosme N, Hauschild MZ (2017) Characterization of waterborne nitrogen emissions for marine eutrophication modelling in life cycle impact assessment at the damage level and global scale. Int J Life Cycle Assess. doi:10.1007/s11367-017-1271-5
Cosme N, Koski M, Hauschild MZ (2015) Exposure factors for marine eutrophication impacts assessment based on a mechanistic biological model. Ecol Model 317:50–63
Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 establishing a framework for community action in the field of water policy. OJL 327/1
Directive 2009/28/EC of the European Parliament and of the Council of 23 April 2009 on the promotion of the use of energy from renewable sources and amending and subsequently repealing Directives 2001/77/EC and 2003/30/EC. OJL 140/16
EC-JRC (2010) International Reference Life Cycle Data System (ILCD) Handbook—framework and requirements for life cycle impact assessment models and indicators. European Commission, Joint Research Center, Institute for Environment Sustainability. Publications Office of the European Union, Luxembourg
Eurostat (2016) Land cover statistics. http://ec.europa.eu/eurostat/statistics-explained/index.php/Land_cover_statistics. Accessed 16 February 2016
Fedele A, Mazzi A, Niero M, Zuliani F, Scipioni A (2014) Can the life cycle assessment methodology be adopted to support a single farm on its environmental impacts forecast evaluation between conventional and organic production? An Italian case study. J Clean Prod 69:49–59
Finnveden G, Nilsson M (2005) Site-dependent life-cycle impact assessment in Sweden. Int J Life Cycle Assess 10:235–239
Finnveden G, Potting J (1999) Eutrophication as an impact category. Int J Life Cycle Assess 4:311–314
Fisher TR et al (1999) Spatial and temporal variation of resource limitation in Chesapeake Bay. Mar Biol 133:763–778
Gallego A, Rodriguez L, Hospido A, Moreira MT, Feijoo G (2010) Development of regional characterization factors for aquatic eutrophication. Int J Life Cycle Assess 15:32–43
Guinée JB (2002) Handbook on life cycle assessment operational guide to the ISO standards. Kluwer Academic Publishers, Dordrecht, Boston
Hauschild M, Potting J (2005) Spatial differentiation in life cycle impact assessment—the EDIP2003 methodology. Danish Environmental Protection Agency, Copenhagen
Hauschild MZ et al (2013) Identifying best existing practice for characterization modeling in life cycle impact assessment. Int J Life Cycle Assess 18:683–697
Hejzlar J et al (2009) Nitrogen and phosphorus retention in surface waters: an inter-comparison of predictions by catchment models of different complexity. J Environ Monit 11:584–593
HELCOM (2007) HELCOM Baltic Sea action plan. Helsinki, Helsinki Commission
HELCOM (2009) Eutrophication in the Baltic Sea. An integrated thematic assessment of the effects of nutrient enrichment in the Baltic Sea region. Helsinki, Baltic Marine Environment Protection Commission
HELCOM (2013) HELCOM Copenhagen ministerial declaration. Helsinki Commission, Copenhagen
Hellweg S, Milà i Canals L (2014) Emerging approaches, challenges and opportunities in life cycle assessment. Science 344:1109–1113
Hooda PS, Truesdale VW, Edwards AC, Withers PJA, Aitken MN, Miller A, Rendell AR (2001) Manuring and fertilization effects on phosphorus accumulation in soils and potential environmental implications. Adv Environ Res 5:13–21
Huijbregts MAJ et al (2016) ReCiPe 2016: a harmonized life cycle impact assessment method at midpoint and endpoint level Report I: characterization RIVM Rapport 2016–0104
Hägg HE, Lyon SW, Wallstedt T, Morth CM, Claremar B, Humborg C (2014) Future nutrient load scenarios for the Baltic Sea due to climate and lifestyle changes. Ambio 43:337–351
Johnsson H, Larsson M, Lindsjö A, Mårtensson K, Persson K, Torstensson G (2008) Nutrient leaching from arable land in Sweden. Calculations of the normalized loads of nitrogen and phosphorus for 1995 and 2005 [in Swedish] (Swedish title: Läckage av näringsämnen från svensk åkermark – Beräkningar av normalläckage av kväve och fosfor för 1995 och 2005). Swedish Environmental Protection Agency, Stockholm
Kangro K, Olli K, Tamminen T, Lignell R (2007) Species-specific responses of a cyanobacteria-dominated phytoplankton community to artificial nutrient limitation in the Baltic Sea. Mar Ecol Prog Ser 336:15–27
Korsaeth A, Henriksen TM, Roer AG, Stromman AH (2014) Effects of regional variation in climate and SOC decay on global warming potential and eutrophication attributable to cereal production in Norway. Agric Syst 127:9–18
Lehuger S, Gabrielle B, Gagnaire N (2009) Environmental impact of the substitution of imported soybean meal with locally-produced rapeseed meal in dairy cow feed. J Clean Prod 17:616–624
Merot P, Aurousseau P, Gascuel-Odoux C, Durand P (2009) Innovative assessment tools to improve water quality and watershed management in farming areas. Integr Environ Assess Manag 5:158–166
Morrice JA, Valett HM, Dahm CN, Campana ME (1997) Alluvial characteristics, groundwater-surface water exchange and hydrological retention in headwater streams. Hydrol Process 11:253–267
Mutel CL, Hellweg S (2009) Regionalized life cycle assessment: computational methodology and application to inventory databases. Environmental Science & Technology 43:5797–5803
Mutel CL, Pfister S, Hellweg S (2012) GIS-based regionalized life cycle assessment: how big is small enough? Methodology and case study of electricity generation. Environ Sci Technol 46:1096–1103
Nausch M, Nausch G, Wasmund N (2004) Phosphorus dynamics during the transition from nitrogen to phosphate limitation in the central Baltic Sea. Mar Ecol Prog Ser 266:15–25
Nitschelm L, Aubin J, Corson MS, Viaud V, Walter C (2016) Spatial differentiation in life cycle assessment LCA applied to an agricultural territory: current practices and method development. J Clean Prod, 112, Part 4 :2472–2484
Page G, Ridoutt B, Bellotti B (2014) Location and technology options to reduce environmental impacts from agriculture. J Clean Prod 81:130–136
Pennington DW, Potting J, Finnveden G, Lindeijer E, Jolliet O, Rydberg T, Rebitzer G (2004) Life cycle assessment Part 2: current impact assessment practice. Environ Int 30:721–739
Potting J, Hauschild M (2006) Spatial differentiation in life cycle impact assessment: a decade of method development to increase the environmental realism of LCIA. Int J Life Cycle Assess 11:11–13
Redfield AC (1958) The biological control of chemical factors in the environment. Am Sci 46:230A–2221
Rolff C, Elfwing T (2015) Increasing nitrogen limitation in the Bothnian Sea, potentially caused by inflow of phosphate-rich water from the Baltic Proper. Ambio 44:601–611
Röös E, Sundberg C, Hansson PA (2011) Uncertainties in the carbon footprint of refined wheat products: a case study on Swedish pasta. Int J Life Cycle Assess 16:338–350
Savchuk OP (2005) Resolving the Baltic Sea into seven subbasins: N and P budgets for 1991–1999. J Mar Syst 56:1–15
Scherer L, Pfister S (2015) Modelling spatially explicit impacts from phosphorus emissions in agriculture. Int J Life Cycle Assess 20:785–795
Seppälä J, Knuuttila S, Silvo K (2004) Eutrophication of aquatic ecosystems—a new method for calculating the potential contributions of nitrogen and phosphorus. Int J Life Cycle Assess 9:90–100
SMED (2015a) Data: JordbrukMetadata_PLC5 http://www.smed.se/vatten/data/plc5. Accessed 17 November 2015
SMED (2015b) Data: Jordbruksgrödor_PLC5 http://www.smed.se/vatten/data/plc5. Accessed 17 November 2015
SMED (2015c) Data: PLC5. Retention_N_ P_normal_bakgrund_PLC5 http://www.smed.se/vatten/data/plc5. Accessed 17 November 2015
Smith VH, Tilman GD, Nekola JC (1999) Eutrophication: impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environ Pollut 100:179–196
Steffen W et al (2015) Planetary boundaries: guiding human development on a changing planet. Science. doi:10.1126/science.1259855
Struijs J, Beusen A, van Jaarsveld H, Huijbregts MAJ (2013) Eutrophication. In: Goedkoop M, Heijungs R, Huijbregts MAJ, De Schryver A, Struijs J, van Zelm R (eds) ReCiPe 2008. Report I: Characterisation. First edition (version 1.08)
Swedish EPA (2006) Eutrophication of Swedish seas: Final report Vol report 5509. Swedish Environmental Protection Agency, Stockholm
Swedish EPA (2008) Eutrophication of the seas along Sweden’s West Coast. Swedish Environmental Protection Agency, Stockholm
Tamminen T, Andersen T (2007) Seasonal phytoplankton nutrient limitation patterns as revealed by bioassays over Baltic Sea gradients of salinity and eutrophication. Mar Ecol Prog Ser 340:121–138
Tysmans DJJ, Lohr AJ, Kroeze C, Ivens W, van Wijnen J (2013) Spatial and temporal variability of nutrient retention in river basins: a global inventory. Ecol Indic 34:607–615
Vahtera E et al (2007) Internal ecosystem feedbacks enhance nitrogen-fixing cyanobacteria blooms and complicate management in the Baltic Sea. Ambio 36:186–194
Acknowledgements
The authors would like to thank Anders Larsolle at the Department of Energy and Technology, Swedish University of Agricultural Sciences, for the help with the visualisation of the characterisation factors.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Disclosure
This study was funded by the Swedish Research Council Formas through the project “Future Crop Production Relies on New Management Strategies for Soils” (grant no: 229-2013-82).
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Responsible editor: Ian Vázquez-Rowe
Electronic supplementary material
ESM 1
(PDF 605 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Henryson, K., Hansson, PA. & Sundberg, C. Spatially differentiated midpoint indicator for marine eutrophication of waterborne emissions in Sweden. Int J Life Cycle Assess 23, 70–81 (2018). https://doi.org/10.1007/s11367-017-1298-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11367-017-1298-7