
Abstract
The impact of climate change drivers on cultivated plants and pest insects has come into research focus. One of the most significant drivers is atmospheric carbon dioxide, which is converted into primary plant metabolites by photosynthesis. Increased atmospheric CO2 concentrations therefore affect plant chemistry. The chemical composition of non-volatile and volatile organic compounds of plants is used by insects to locate and identify suitable host plants for feeding and reproduction. We investigated whether elevated CO2 concentrations in the atmosphere affect the plant-pest interaction in a fruit crop of high economic importance in Europe. Therefore, potted pear trees were cultivated under specified CO2 conditions in a Free-Air Carbon dioxide Enrichment (FACE) facility at Geisenheim University in Germany for up to 14 weeks, beginning from bud swelling. We compared emitted volatiles from these pear trees cultivated for 7 and 14 weeks under two different CO2 levels (ambient: ca. 400 ppm and elevated: ca. 450 ppm CO2) and their impact on pest insect behavior. In total, we detected and analyzed 76 VOCs from pear trees. While we did not detect an overall change in VOC compositions, the relative release of single compounds changed in response to CO2 increase. Differences in VOC release were inconsistent over time (phenology stages) and between study years, indicating interactions with other climate parameters, such as temperature. Even though insect-plant interaction can rely on specific volatile compounds and specific mixtures of compounds, respectively, the changes of VOC patterns in our field study did not impact the host choice behavior of C. pyri females. In olfactometer trials, 64% and 60% of the females preferred the odor of pear trees cultivated under elevated CO2 for 7 and 14 weeks, respectively, over the odor from pear trees cultivated under ambient CO2. In binary-choice oviposition assays, C. pyri females laid most eggs on pears during April 2020; on average, 51.9 (± 51.3) eggs were laid on pears cultivated under eCO2 and 60.3 (± 48.7) eggs on aCO2.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Atmospheric carbon dioxide (CO2) concentrations are rising globally, mainly driven by economic growth and expansion of the human population (Feng et al. 2020). The greenhouse gas CO2 contributes to the global warming problem (IPCC 2014). Therefore, the reduction of anthropogenic CO2 emission was claimed as one of the most important goals by the Intergovernmental Panel on Climate Change (IPCC) in 2014 (IPCC 2014). While the increase of CO2 emission has slowed down in the European Union and the United States during the past decade, on a global level, the emission is still rising (Peters et al. 2020). Even though intensified production and cultivation practices of livestock and plants considerably contribute to the emission of greenhouse gases (Shakoor et al. 2021), agriculture itself is heavily affected by climate change, and its determinants are bearing new challenges for growers (Anderson et al. 2020). Changes of climate parameters and climate change drivers are impacting plant physiology as well as insect phenology, performance, and plant–insect interactions (Blande 2021; DeLucia et al. 2012; Forrest 2016; Jürgens and Bischoff 2017).
Regarding plant physiology, the global increase of carbon dioxide is of special interest, as plants utilize CO2 as carbon source to produce primary metabolites by photosynthesis. Therefore, higher CO2 concentrations have the potential to increase biomass production and crop yields and impact the nutritional quality of crops (Ainsworth and Long 2021; Dong et al. 2018; Wohlfahrt et al. 2018). In 2012, it was shown that growth under high CO2 concentrations increases photosynthetic rates in “Niitaka” pears (Pyrus pyrifolia) and resulted in higher soluble sugar contents in pear fruits (Han et al. 2012). A fertilizing effect of CO2 can increase the amount of primary metabolites, which are resources for production of secondary metabolites, such as volatile organic compounds (VOCs). Multifarious VOCs are released from plants. Characteristic plant VOC compositions are species, or even genotype and developmental stage specific, as well as inducible by biotic and abiotic stresses (Gallinger et al. 2020; Rid et al. 2019; Hickman et al. 2021). Thus, VOCs provide information regarding the identity and condition of the releasing plant to other organisms in their environment (Bouwmeester et al. 2019). Regarding the effect of CO2 on plant VOCs, it is shown that the exposition of oilseed rape plants to elevated CO2 concentrations (720 µl l−1) increased the constitutive emission of several terpenoids (Himanen et al. 2009). Such plant-derived VOCs are important for the intraspecific communication of plants. For insects, olfactory, besides visual, cues are important for the orientated movement (searching behavior) over distance (Deletre et al. 2016; Schoonhoven et al. 2005). Therefore, a change in the VOC composition emitted by plants under increased carbon dioxide concentrations can impact insect behavior (Chen et al. 2019).
Only a small number of studies have been carried out under field conditions because sophisticated facilities are needed to manipulate CO2 levels in open-field situations (Ainsworth and Long 2005, 2021). Especially, studies with slow-growing perennial plants are rare. To date, there is no study that has investigated the impact of CO2 concentrations on the interaction of pear trees and one of their most threatening pests, pear psyllids.
Psyllids or jumping plant lice belong to the order of Hemiptera and feed with their piercing-sucking mouth parts on the phloem tissue of their host plants. Psyllids are serious pests of several cultivated plants. Species in the genus Cacopsylla are known to be harmful to fruit trees, and the pear psyllid Cacopsylla pyri is a major pest in pear-growing regions all over Europe (Garcia-Chapa et al. 2005). C. pyri is damaging fruit trees directly and indirectly. Pear trees can suffer from massive psyllid infestations, due to direct feeding damage. In the event of mass occurrence, enormous amounts of honeydew, especially from nymphs, are excreted, serving as a medium for sooty mold fungus (Burckhardt 1994). Additionally, C. pyri is a vector of the phytopathogen “Candidatus Phytoplasma pyri”, a specialized cell wall–less bacterium colonizing the insect vector and the phloem tissue of pear trees (Carraro et al. 1998). While feeding on the phloem sap of host trees, psyllids can transmit “Ca. P. pyri” from plant to plant (Jarausch et al. 2019). This bacterium induces the so-called pear decline, a severe disease in pear-growing. Hence, C. pyri is of high economic relevance.
A number of studies on different species demonstrated that psyllids perceive plant volatiles and evaluated the role of plant chemical cues for host finding by psyllids (Alquézar et al. 2017; Coutinho-Abreu et al. 2014; Gallinger et al. 2020; George et al. 2016; Gross et al. 2019b; Kristoffersen et al. 2008; Mayer et al. 2008a, 2008b; Nehlin et al. 1994; Rid et al. 2016; Soroker et al. 2004; Yuvaraj et al. 2013). The detection of suitable host plants is crucial for survival and reproductive success of herbivorous insects. Therefore, we hypothesize that changes in volatile emission of host plants can impact the host preference of pear psyllids, which may impact their distribution and the dispersion of “Ca. P. pyri” in the future. Therefore, we firstly investigated whether increased CO2 concentrations affect the emission of VOCs from pear trees. This was realized under field conditions in a Free-Air Carbon dioxide Enrichment (FACE) facility where potted pear trees were placed under different CO2 levels before bud break. Further, we evaluated how changes in the VOC profiles impact the host choice of C. pyri females and thus potentially alter psyllid pest pressure in pear orchards under future conditions.
Materials and methods
Plants and insects
Two-year-old Pyrus communis cv. Williams Christ trees (rootstock: quince BA 29) obtained from a nursery (Jäger, Ladenburg, Germany) were used for the experiments. In both years (2019 and 2020), trees were potted in 25-L pots and pruned in February. Afterwards (2019: February 25, 2019, and 2020: March 2, 2020), the potted pear trees were placed in the inter-rows of the VineyardFACE facility at Geisenheim University, Germany, for a total period of 13 weeks in 2019 and 14 weeks in 2020. The FACE facility consisted of six rings; in three rings, an elevated concentration of CO2 (eCO2, approx. 450 ppm) was established, whereas in the other three rings, the ambient CO2 level (aCO2, approx. 400 ppm) was not modified (Fig. 1c). For a detailed description of the Geisenheim VineyardFACE facility, see publications by Reineke and Selim (2019) and Wohlfahrt et al. (2018). In 2019, three pear trees were placed at one edge of each ring, resulting in nine trees each for ambient and elevated CO2 concentration (Fig. 1a–c). In 2020, the number of pear trees per ring was increased to six, and the trees were positioned in the middle of each ring, resulting in 18 trees for each CO2 concentration (Fig. 1a). In 2022, half of the trees (three trees per ring) were removed from the FACE facility after 7 weeks and used for behavioral experiments with pear psyllids.

a Position of potted pear trees 2019 and 2020 in the inter-rows of the VineyardFACE facility at Geisenheim University, Germany. b Close-up of a FACE ring, consisting of 36 jets mounted at a height of 2.5 m equipped with fans for CO2 distribution. c Overview of the VineyardFACE experimental site, with three FACE rings assigned to the two CO2 concentrations, ambient CO2 (aCO2 ~ 400 ppm) and elevated CO2 (eCO2 ~ 450 ppm), and the associated CO2 tank
All pear trees were fertilized once at the beginning of the growing season with approx. 20-g slow-release fertilizer (Triabon 16–8-12 (+ 4 + TE), COMPO EXPERT GmbH, Münster, Germany). All plants were watered whenever necessary. Phenological growth stages of pear trees at the different sampling dates were classified following the BBCH identification key for pome fruits by Meier (2018): principal growth stage 3, shoot development; and secondary stages 1–9, shoots about 10–90% of final length.
Additional P. communis (cv. Williams Christ on Kirchensaller Mostbirne) were grown for the insect rearing and the no-choice oviposition assays, in an insect save screen house under ambient climatic conditions. Pear trees were fertilized with ~ 500-mL Triabon (COMPO Expert GmbH, Münster, Germany, 2 g/L) once (in March).
Adult C. pyri (summer form) were caught from P. communis trees at the experimental field of the Julius Kühn-Institut (JKI) in Dossenheim, Germany (49°27′00.0″N, 8°38′24.0″E), by beating tray method. A square beat sheet was placed under a branch of a pear tree; the branch was hit with a foam plastic-covered stick, causing the psyllids to fall on the sheet (Weintraub and Gross 2013). All psyllids were collected from the sheet. Species and sex was determined under a stereo microscope (Stemi 2000, Zeiss, Oberkochen, Germany). Captured C. pyri adults were either directly used for experiments (olfactometer experiments within 24 h) or kept in a climate chamber at 20 °C (photophase) and 16 °C (scotophase) (L16:D8) until they were used in oviposition experiments. Therefore, collected C. pyri adults were kept on P. communis (cv. Williams Christ) plants in screen cages (BugDorm, MegaView Science Co, Taiwan, 47.5 × 47.5 × 93 cm).
Volatile collection
Volatile emissions were sampled from potted P. communis trees in a non-destructive way in the field (FACE facility) at two phenological growth stages (shoot development) per year (2019: BBCH 32–33, May 7, 2019, and BBCH 38–39, June 4, 2019; 2020: BBCH 32–33, April 21, 2020, and BBCH 38–39, June 2, 2020). In 2019, the volatiles from nine trees per CO2 condition were sampled at both sampling dates. In 2020, the volatiles from 18 pear trees per CO2 condition were sampled at the first sampling date (BBCH 32–33, April 21, 2020); afterwards, half of the trees were removed from the FACE facility and used for behavioral studies, resulting in a total number of nine pear trees per CO2 condition that were sampled at the second BBCH stage (BBCH 38–39, June 2, 2020). Samples were taken with a mobile 6-channel headspace sampling device composed of six mass flow controllers (M + W Instruments GmbH, Leonhardsbuch, Germany) each controlling one vacuum pump (KNF Neuberger GmbH, Freiburg, Germany) as described by Rid et al. (2016). One single branch of each tree was carefully wrapped in polyethylene terephthalate oven bags (Toppits, Melitta, Minden, Germany). Clean air filter cartridges (ICAF 2X6, Sigma Scientific, Micanopy, USA) were used to filter the ambient air. The clean air was pumped through the oven bags with a flow of 1000 mL/min. Volatiles were trapped on stainless steel, prepacked sample tubes containing 200-mg Tenax TA35/60 sorbent (Markes, Neu-Isenburg, Germany). A total volume of 30 L was pumped over the matrix. Samples were analyzed by GC–MS analysis within 1 week.
GC–MS analysis
Volatile samples were analyzed with an automated thermal desorber (TurboMatrix™ ATD 650, PerkinElmer, Rodgau, Germany) connected to a gas chromatograph (Clarus R 680, PerkinElmer) coupled to a Perkin Elmer quadrupole inert mass selective detector (ATD-GC–MS). Sample tubes were desorbed for 10 min at 250 °C. A Tenax TA–filled cold trap was held at − 20 °C through the desorption followed by heating at a rate of 99 K/s to 250 °C and 1 min desorption time. A nonpolar Elite-5MS (Crossbond 5% diphenyl/95% dimethyl polysiloxane, PerkinElmer) capillary column (30 × 0.25-mm id × 0.25-μm film thickness) was used for GC separation of volatile compounds. Helium was used as carrier gas (Helium 6.0, Linde, Munich, Germany) with a column head pressure of 130 kPa. The GC temperature program was as follows: Initial oven temperature of 40 °C was held for 1 min, increased at a rate of 5 K/min to 180 °C, followed by a rate of 20 K/min to the final temperature of 280 °C, and held for 6 min. The GC inlet line temperature was 250 °C, and the ion source temperature was 180 °C. The quadrupole mass detector was operated in the electron impact (EI) mode at 70 eV. All data were obtained by collecting the full-scan mass spectra within the range of 35–350 m/z, resulting in the following number of chromatograms per year and BBCH stage used in statistical analysis, in 2019: naCO2BBCH32–33 = 8, neCO2BBCH32–33 = 7, naCO2BBCH38–39 = 9, and neCO2BBCH38–39 = 9, and in 2020: naCO2BBCH32–33 = 18, neCO2BBCH32–33 = 18, naCO2BBCH38–39 = 9, and neCO2BBCH38–39 = 9.
Identification and quantification with AMDIS
Chromatograms of volatile samples were analyzed using “Automated Mass spectral Deconvolution and Identification System” (AMDIS, V. 2.71; National Institute of Standards and Technology NIST, Boulder, CO). For the identification, the ion fragmentation patterns and retention indices of detected compounds were compared with standard compounds according to Gross et al. (2019a). Not identified compounds were annotated as unknowns. For quantification, the peak areas were integrated after deconvolution. Identification criteria were applied as follows: match factor had to be ≥ 80% and the relative retention index deviation ≤ 5% from reference value. The settings for deconvolution were component width, 32; adjacent peak subtraction, one; resolution, low; sensitivity, medium; shape requirements, low; level, very strong; maximum penalty, 20; and “no RI in library” 20. Components with a signal to noise ratio < 50 were excluded from the analysis.
Olfactometer assays
The preference of C. pyri females for odors of P. communis trees cultivated under different CO2 levels was evaluated in a dynamic Y-shaped olfactometer, after 7 weeks (BBCH 32–33) and 14 weeks (BBCH 38–39) of cultivation in the FACE facility. The Y-shaped glass tube was mounted on a board in an angle of 40° from the horizontal plane. The entrance arm length of the tube was 12.5 cm, the test arm length 8 cm, and the inner diameter 1 cm; test arms had an angle of 75°. A light source (LED-Lupenleuchte, Purelite, UK) was mounted 45 cm above the middle of the olfactometer, resulting to approx. 280 lx at the experimental area. All experiments were conducted between 09:00 a.m. and 6:00 p.m. at room temperature (20–26 °C and 30–35% RH). One potted pear tree per CO2 condition and growth stage was transferred from the FACE facility in Geisenheim to the JKI in Dossenheim. The trees were used for olfactometer assays 2 days in the following. One twig of each tree was carefully wrapped in oven plastic bags (Toppits, Melitta, Minden, Germany, 31 × 50 cm) and connected to the test arms. Charcoal-filtered and humidified air was pumped through the plastic bags into the test arms. The air flow of each test arm was adjusted to 75 mL/min (with a max. difference of 1 mL/min) by plastic valves and controlled by a flowmeter (MASS-STREAM, M + W Instruments, Allershausen, Germany) to ensure an equal airstream in both arms. The odors of P. communis (cv. Williams Christ on BA29) trees cultivated under elevated (approx. 450 ppm) or ambient (approx. 400 ppm) CO2 levels in the FACE facility were offered simultaneously or against clean air to C. pyri females. Psyllids were collected from the experimental field at the JKI and kept in reaction vials in the dark prior to olfactometer tests. Each psyllid was released from the reaction tube at the base of the entrance arm. To prevent side effects, the olfactometer was rotated 180° after five replicates. The number of females that entered one of the test arms (1 cm) and stayed for at least 30 s was counted. Psyllids that did not reach one of the test arms within 10 min were recorded as “no choice.” Olfactometers and accessories were cleaned with ethanol (70%) and heated at 230 °C (except plastic valves: 60 °C) for 2 h. Previous studies showed changes in psyllid behavior due to phytoplasma infections (Mayer et al. 2008a); therefore, infection status of C. pyri females was determined. Females from olfactometer assays were frozen at − 20 °C and tested for phytoplasma infection by DNA extraction followed by PCR and gel electrophoresis. Choices of females, which were infected with phytoplasmas, were excluded from the analysis. In total, the behavior of 46 to 52 females per comparison was observed.
Oviposition assays
To investigate the oviposition behavior of C. pyri, we conducted (“No-choice” section) no-choice experiments and (“Binary-choice” section) binary-choice experiments under the following conditions.
No-choice
No-choice oviposition assays were conducted to investigate a direct and short-term effect of CO2 on the oviposition preference of C. pyri females. Therefore, the assays were conducted in the presence of ambient (400 ppm) or strongly elevated (750 ppm) CO2 concentrations in a climate cabinet (Rumed P 1060, Rubarth Apparate GmbH, Laatzen, Germany) under controlled climate conditions at 20 °C during photophase and 16 °C during scotophase (L16:D8) and 60 ± 5% relative humidity. In total, thirty C. pyri females were placed on leaves of the additional planted P. communis (cv. Williams Christ on Kirchensaller Mostbirne) plants from the screen house per CO2 concentration. Each female was individually caged on one leaf with a gauze bag (10 × 12 cm). Two trees were used for each CO2 condition, and 15 females were caged at each tree (naCO2 = 30, neCO2 = 30). After 96 h, the leaves with the psyllids were detached from the pear trees and stored at − 20 °C until the number of eggs was counted under a stereomicroscope (Stemi 508, Carl Zeiss AG, Oberkochen, Germany). Oviposition assay was conducted in August 2019 with females from the third generation of reared C. pyri (as described in the”Plants and insects” section).
Binary-choice
Binary-choice assays were conducted with P. communis (cv. Williams Christ on BA29) trees grown under ambient (approx. 400 ppm) or elevated (approx. 450 ppm) CO2 levels. Therefore, eight of the potted pear trees per CO2 level were removed from the Geisenheim VineyardFACE facility (16 in total). The oviposition assays were conducted immediately after removing the trees from the facility in an insect save screen house at Geisenheim University. One intact leaf each from a tree grown under ambient and elevated CO2 concentrations was offered simultaneously, by inserting one intact leaf of each tree from opposite sides into a gauze sleeve (20 × 30 cm). In 2019 a single female (nBBCH38-39 = 47) and in 2020 one pair of C. pyri adults (one of each sex) was introduced in each bag (nBBCH32–33 = 49, nBBCH38–39 = 47). In both years, psyllids were allowed to oviposit for 5 days (120 h). Following this, the leaves were removed from the plants and frozen at − 20 °C. The number of eggs laid on each leaf was counted under a stereomicroscope (Stemi 508, Carl Zeiss AG, Oberkochen, Germany).
Statistical analysis
Statistical analysis was done in R version 4.0.3. (R Core Team 2020).
Volatile profiles: “vegan” package was used for multivariate analysis of volatile profiles from P. communis trees (Oksanen et al. 2020). A Bray–Curtis dissimilarity matrix was calculated from the compositional data set of volatiles with the vegdist function. Non-metric multidimensional scaling (NMDS) plots were used to visualize Bray–Curtis dissimilarities with the metaMDS function. Scaling was standardized by Wisconsin double standardization. Permutational multivariate analysis of variance (PERMANOVA) was used to analyze differences in volatile compositions between the CO2 treatments and sampling dates (BBCH stages), with the adonis function (N permutations = 10.000). Additionally, the dispersion of groups was tested for multivariate homogeneity. The relative amount of single volatiles in the profiles was compared between pears grown under aCO2 and eCO2 conditions with a nonparametric Mann–Whitney U-test.
Olfactometer assays: Preference for a host plant volatile profile was evaluated with the binominal test.
Oviposition: In binary-choice test, replicates where no eggs were laid were excluded from the analysis. In no-choice tests, zeros were not excluded, because they could represent avoidance behavior. Zero-inflated negative binomial (ZINB) models were fitted to evaluate the difference in number of eggs laid on leaves from pear trees grown under aCO2 and eCO2 conditions in the binary-choice experiment. The use of zero-inflated models provides the possibility to estimate the effect of covariates on the probability to encounter a zero in the count data. ZINB models were generated with the zeroinfl function from “pscl” package (Zeileis et al. 2008). The log-transformed total number of eggs per female was used as offset. We used AIC model selection to compare models that include or not include the combination of trees as factors. Integration of the tree combination as model factor did not increase the model fit and was therefore not included in the models.
Differences in oviposition choice depending on the growth condition (CO2 concentration) of the pear trees were calculated using estimated marginal means and 95% confidence intervals with the emmeans function from “emmeans” package (Lenth 2021).
Results
Volatile emission
In total, 76 components (Table S1) were annotated from the chromatograms and compared between the CO2 treatments at two growth stages.
In both years, the volatile composition emitted from pear trees at the two investigated phenological stages differed significantly (Fig. 2, (a) 2019: perMANOVA, F = 9.8859, P = 0.0002, N = 33; (b) 2020: perMANOVA, F = 13.98, P = 0.00009, N = 54). The CO2 concentration as well as the interaction between the phenological stage and the CO2 concentration had no significant effect on the discrimination between the volatile profiles (Fig. 2, (a) 2019: perMANOVA, F = 1.04, P = 0.361, N = 33; (b) 2020: perMANOVA, F = 0.96, P = 0.430, N = 54).

Non-metric multidimensional scaling (NMDS) plot based on Bray–Curtis dissimilarities calculated from proportional volatiles compositions of Pyrus communis trees grown under ambient (blue circles) or elevated (red triangles) CO2 conditions at two different phenological stages in the FACE system. a 2019 (2d-stress: 0.111; naCO2BBCH32–33 = 8, neCO2BBCH32–33 = 7, naCO2BBCH38–39 = 9, neCO2BBCH38–39 = 9) and b 2020 (2d-stress: 0.107; naCO2BBCH32–33 = 18, neCO2BBCH32–33 = 18, naCO2BBCH38–39 = 9, neCO2BBCH38–39 = 9)
Single volatiles were emitted in significantly different amounts from pear trees under the two CO2 concentrations. In 2019, pear trees at the early phenological stage (BBCH 32–33) released significantly less α- and β-caryophyllene (Fig. 3a, Mann–Whitney U, α-caryophyllene: Z = 2.08, P = 0.038, N = 15; β-caryophyllene: Z = 2.07, P = 0.038, N = 15) under eCO2 than aCO2 condition. Later, in the season (BBCH 38–39), the release of caryophyllene did not differ (Mann–Whitney U, α-caryophyllene: Z = 1.29, P = 0.199, N = 18; β-caryophyllene: Z = 0.22, P = 0.825, N = 18) between CO2 conditions. Pear trees under eCO2 conditions released significantly lower amounts of 2-carene (Fig. 3a, Mann–Whitney U, Z = 2.12, P = 0.034, N = 18) and 6-methyl-5-hepten-2-one (Fig. 3a, Mann–Whitney U, Z = 2.16, P = 0.031, N = 18) than under aCO2 conditions, whereas the emission of decane (Fig. 3a, Mann–Whitney U, Z = − 2.18, P = 0.029, N = 18) was increased under eCO2 compared to aCO2 conditions.

Box-Whisker plots with jittered raw values of relative amount of single volatile compounds from Pyrus communis trees grown under ambient (blue) or elevated (red) CO2 conditions in the FACE system. a 2019 at BBCH 32–33 (naCO2 = 8, naCO2 = 7) and BBCH 38–39 (naCO2 = 9, naCO2 = 9), b 2020 at BBCH 32–33 (naCO2 = 18, naCO2 = 18) and BBCH 38–39 (naCO2 = 9, naCO2 = 9). Medians are shown as lines, and whiskers extend to 1.5 times of the interquartile ranges
In 2020, pear trees at early growth stage (BBCH32–33) released significantly higher amounts of 1-tetradecane (Fig. 3b, Mann–Whitney U, Z = − 2.46, P = 0.014, N = 36) and γ-terpinene (Fig. 3b, Mann–Whitney U, Z = − 2.17, P = 0.03, N = 36) when grown under eCO2 than under aCO2 condition. Later, in the season (BBCH38–39), elevated CO2 levels lead to increased amounts of nonadecane (Fig. 3b, Mann–Whitney U, Z = − 2.03, P = 0.042, N = 18), 1-hexadecene (Fig. 3b, Mann–Whitney U, Z = − 2.17, P = 0.03, N = 18), γ-terpinene (Fig. 2b, Mann–Whitney U, Z = − 2.36, P = 0.018, N = 18), and one unidentified component RI1695 (Fig. 3b, Mann–Whitney U, Z = − 2.56, P = 0.010, N = 18) in the overall composition. The release of 1-octene (Fig. 3b, Mann–Whitney U, Z = 2.25, P = 0.024, N = 18), hexanal (Fig. 3b, Mann–Whitney U, Z = 2.34, P = 0.019, N = 18), and one unidentified component RI1098 (Fig. 3b, Mann–Whitney U, Z = 2.03, P = 0.043, N = 18) was reduced in odor profiles of pear trees cultivated under elevated compared to ambient CO2 concentration.
Olfactometer assays
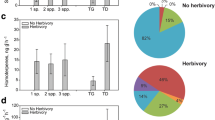
Cacopsylla pyri females did not significantly prefer odor blends from P. communis plants over clean air in olfactometer assays at BBCH 32–33 (Fig. 4a, binominal test, aCO2: P = 0.883, N = 46; eCO2: P = 0.48, N = 50) nor at BBCH 38–39 (Fig. 4b, binominal test, aCO2: P = 0.332, N = 52; eCO2: P = 1, N = 51). Additionally, females did not show a preference for odors from P. communis plants grown under elevated or ambient CO2 levels in the FACE environment (Fig. 4, (a) BBCH 32–33: binominal test, P = 0.065, N = 50; (b) BBCH 38–39: binominal test, P = 0.193, N = 48). When offered simultaneously, 64% of female C. pyri walked in the direction of the odor from eCO2 pears at BBCH 32–33 (Fig. 4a) and 60% at BBCH 38–39 (Fig. 4b). General motivation of females was high and ranged from 87 to 100% (Fig. 4).

Choice of C. pyri females in a Y-tube olfactometer for odor profiles of Pyrus communis trees grown under ambient (approx. 400 ppm, blue) or elevated (approx. 450 ppm, red) CO2 levels at a BBCH 32–33 and b BBCH 38–39 in 2020. Percentage of psyllids that made a choice (dark gray) and that did not (light gray) is presented as pie chart on the right (n.s. = not significant, binominal test)
Oviposition
Cacopsylla pyri females did not show significant oviposition preference for P. communis trees grown under ambient or elevated CO2 levels in no-choice and binary-choice bioassays (Table 1). In no-choice bioassay, C. pyri females laid on average 18.3 (± 29.9) eggs on pear leaves from ambient CO2 and 24.4 (± 30.7) eggs from elevated CO2 grown trees. Given a choice between two leaves from trees grown under ambient or elevated CO2 concentrations, C. pyri females laid on average 60.3 (± 48.7) eggs on aCO2 and 51.9 (± 51.3) eggs on eCO2 grown pear leaves at BBCH 32–33; 21.7 (± 30.9) and 21.7 (± 24.3) eggs on aCO2 and eCO2 grown leaves, respectively, at BBCH 38–39 in 2019; and 21.0 (± 24.4) and 16.1 (± 22.4) eggs on aCO2 and eCO2 grown leaves, respectively, at BBCH 38–39 in 2020. In general, C. pyri females laid twice as many eggs in experiments with pear trees in early growth stage (BBCH 32–33) in April 2020 compared to binary-choice experiments later in the season (BBCH 38–39; Table 1).
Discussion
Growing under elevated CO2 concentration altered the amount of single VOCs released from pear trees. Many herbivorous insects use specific blends of plant-emitted volatiles to identify and locate suitable host plants (Bruce and Pickett 2011). Therefore, the change in relative release of specific compounds in plant odors was expected to impact the behavior of the highly specialized pest insect C. pyri. Especially, terpenes and aldehydes are known to impact the behavior of different insect species. Thus, among other aldehydes, hexanal is shown to be perceived by Cacopsylla pruni, a psyllid species closely related to C. pyri (Gallinger et al. 2020). Contrary to expectations, the relative decrease of hexanal in the odor profile of pear trees had no impact on the host preference of C. pyri females. Neither had the relative increase of γ-terpinene. So far, it is unknown which compounds C. pyri can perceive (Gross et al. 2022), but in previous studies with Cacopsylla picta, there was evidence that young individuals are attracted by the terpene β-caryophyllene (Mayer et al. 2008a, b). Until now, the role of specific aldehydes and terpenes for host finding and choice of C. pyri remains unknown. The olfactometer experiments revealed no significant attraction of C. pyri females to pear odors, indicating that host selection of C. pyri may not be based on plant volatiles alone. Previous studies on other psyllid species highlighted the importance of visual cues on host selection (Farnier et al. 2018; George et al. 2020; Patt et al. 2011; Wenninger et al. 2009). Further research should be undertaken to investigate the role of visual cues and possible synergistic effects of olfactory and visual cues for host detection of C. pyri.
Nonetheless, changes in VOC profiles could impact other pest insects such as moths or aphids. Due to the complexity and specificity of interactions, no general predictions can be made at this point. Different effects of atmospheric CO2 were found concerning other pest insects and host plants. For the corn leaf aphid Rhopalosiphum maidis, it is shown that winged and wingless adults preferred the volatile profile of barley (Hordeum vulgare) seedlings grown under ambient compared to elevated CO2 concentrations in two choice olfactometer trials (Chen et al. 2019). In contrast, soybean aphid Aphis glycines do not discriminate odors from undamaged soybean grown under ambient or elevated CO2 concentrations (O’Neill et al. 2010). Predators and parasitoids likewise rely on olfactory cues to find their prey. Anthocorid bugs, the most significant predators of C. pyri, are attracted to α-farnesene and methyl salicylate (Scutareanu et al. 2003). These plant volatiles are induced by psyllid feeding (Scutareanu et al. 1997, 2003). We did not find an impact of CO2 on the release of these compounds in our current study. Nevertheless, the effect of climate change parameters on herbivore-induced plant VOCs in the pear-psyllid-predator system is an important issue for future research.
Additionally, we found a variable influence of CO2 on pear VOCs in the two years of our studies. These differences are likely based on interactions with further climatic conditions. Especially, heat and water stress should be considered. Indeed, a high supply with CO2 can lead to an increase in photosynthesis rates, whereas high temperatures can impair the activity of photosystem II and reduce the photosynthetic efficiency (Havaux et al. 1991). The photosynthetic capacity of apple leaves was significantly reduced during daytimes with high temperatures and light intensities (Mihaljević et al. 2020). Additionally, the pear trees were placed in the middle of the FACE rings in 2020, while they were placed along the edges in 2019. The CO2 distribution inside the rings is most homogeneous in the middle of the ring system (data not shown). The CO2 concentration may therefore have been more variable in 2019 than in 2020. Another aspect that should be taken in to consideration is the duration of exposure to elevated CO2 concentrations. In this study, pear trees were exposed to higher CO2 levels for a certain period. We choose to expose the trees before bud break and investigate the plant VOCs and psyllid preferences during leaf and shoot development under CO2 exposition. As this is a critical time in the development of plant as well as psyllid populations, due to limitations in time and space, we were not able to cultivate pear trees for several years. Therefore, we cannot exclude further changes or adaptations of VOC release from pear trees after CO2 exposition for longer time periods.
Changes in volatile emissions from pear trees may indicate changes in plant metabolism. Impacts on primary metabolites are likely to affect feeding behavior, reproduction success, and survival of psyllids (Gallinger and Gross 2018, 2020). A recent study with C. picta demonstrated a correlation of C:N ratio in the host phloem sap and a oviposition preference of C. picta females, highlighting the impact of phloem sap composition for psyllid oviposition choice (Görg et al. 2020). Contrary to our expectations, the results from the oviposition experiments in the current study are not indicating a biologically relevant change in phloem sap quality due to the exposition of the trees to elevated CO2 levels.
Another important aspect in the investigated system is the transmission of phytopathogens by C. pyri. Bosquee et al. (2018) showed a higher transmission efficiency of Potato virus Y for Myzus persicae reared under elevated CO2 compared to aphids reared under ambient CO2 concentrations. Thus, the impact on feeding behavior and phytoplasma transmission efficiency of psyllids should be further investigated with regard on the spread of “Ca. P. pyri”, a phytopathogen that is vectored by C. pyri.
Additionally, CO2 concentrations can affect psyllid fitness and development. A positive impact on insect performance under elevated CO2 concentration was demonstrated for A. glycines on soybeans, by increased numbers of aphids (Dermody et al. 2008). In accordance, Guo et al. (2014) found increased growth rates of pea aphids (Acyrthosiphon pisum) on Medicago truncatula grown under increased CO2 concentration. These effects were attributed to differences in feeding behavior of aphids, due to decreased plant resistance (Guo et al. 2014). This again highlights the importance of further investigations on the interaction between psyllid vectors, phytoplasmas, and their host plants under future climate conditions.
Availability of data and materials
Not applicable.
References
Ainsworth EA, Long SP (2005) What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol 165:351–371. https://doi.org/10.1111/j.1469-8137.2004.01224.x
Ainsworth EA, Long SP (2021) 30 years of free-air carbon dioxide enrichment (FACE): what have we learned about future crop productivity and its potential for adaptation? Global Change Biol 27:27–49. https://doi.org/10.1111/gcb.15375
Alquézar B, Volpe HXL, Magnani RF, de Miranda MP, Santos MA, Wulff NA, Bento JMS, Parra JRP, Bouwmeester H, Peña L (2017) β-caryophyllene emitted from a transgenic Arabidopsis or chemical dispenser repels Diaphorina citri, vector of Candidatus Liberibacters. Sci Rep 7:5639. https://doi.org/10.1038/s41598-017-06119-w
Anderson R, Bayer PE, Edwards D (2020) Climate change and the need for agricultural adaptation. Curr Opin Plant Biol 56:197–202. https://doi.org/10.1016/j.pbi.2019.12.006
Blande JD (2021) Effects of air pollution on plant–insect interactions mediated by olfactory and visual cues. Curr Opin Environ Sci Health 19:100228. https://doi.org/10.1016/j.coesh.2020.100228
Bosquee E, Boullis A, Bertaux M, Francis F, Verheggen FJ (2018) Dispersion of Myzus persicae and transmission of potato virus Y under elevated CO2 atmosphere. Entomol Exp Appl 166:380–385. https://doi.org/10.1111/eea.12661
Bouwmeester H, Schuurink RC, Bleeker PM, Schiestl F (2019) The role of volatiles in plant communication. Plant J 100:892–907. https://doi.org/10.1111/tpj.14496
Bruce TJA, Pickett JA (2011) Perception of plant volatile blends by herbivorous insects-finding the right mix. Phytochemistry 72:1605–1611. https://doi.org/10.1016/j.phytochem.2011.04.011
Burckhardt D (1994) Psylloid pests of temperate and subtropical crop and ornamental plants (Hemiptera, Psylloidea): a review. Trends in Agricultural Sciences, Entomology 2:173–186
Carraro L, Loi N, Ermacora P, Gregoris A, Osler R (1998) Transmission of pear decline by using naturally infected Cacopsylla pyri L. Acta Hortic 665–668. https://doi.org/10.17660/ActaHortic.1998.472.89
Chen Y, Martin C, Fingu Mabola JC, Verheggen F, Wang Z, He K, Francis F (2019) Effects of host plants reared under elevated CO2 concentrations on the foraging behavior of different stages of corn leaf aphids Rhopalosiphum maidis. Insects 10:182. https://doi.org/10.3390/insects10060182
Coutinho-Abreu IV, McInally S, Forster L, Luck R, Ray A (2014) Odor coding in a disease-transmitting herbivorous insect, the Asian citrus psyllid. Chem Senses 39:539–549. https://doi.org/10.1093/chemse/bju023
Deletre E, Schatz B, Bourguet D, Chandre F, Williams L, Ratnadass A, Martin T (2016) Prospects for repellent in pest control: current developments and future challenges. Chemoecology 26:127–142. https://doi.org/10.1007/s00049-016-0214-0
DeLucia EH, Nabity PD, Zavala JA, Berenbaum MR (2012) Climate change: resetting plant-insect interactions. Plant Physiol 160:1677–1685. https://doi.org/10.1104/pp.112.204750
Dermody O, O’Neill BF, Zangerl AR, Berenbaum MR, DeLucia EH (2008) Effects of elevated CO2 and O3 on leaf damage and insect abundance in a soybean agroecosystem. Arthropod-Plant Interactions 2:125–135. https://doi.org/10.1007/s11829-008-9045-4
Dong J, Gruda N, Lam SK, Li X, Duan Z (2018) Effects of elevated CO2 on nutritional quality of vegetables: a review. Front Plant Sci 9:924. https://doi.org/10.3389/fpls.2018.00924
Farnier K, Davies NW, Steinbauer MJ (2018) Not led by the nose: volatiles from undamaged Eucalyptus hosts do not influence psyllid orientation. Insects 9:166. https://doi.org/10.3390/insects9040166
Feng C, Zheng C-J, Shan M-L (2020) The clarification for the features, temporal variations, and potential factors of global carbon dioxide emissions. J Clean Prod 255:120250. https://doi.org/10.1016/j.jclepro.2020.120250
Forrest JR (2016) Complex responses of insect phenology to climate change. Curr Opin Insect Sci 17:49–54. https://doi.org/10.1016/j.cois.2016.07.002
Gallinger J, Gross J (2018) Unraveling the host plant alternation of Cacopsylla pruni—adults but not nymphs can survive on conifers due to phloem/xylem composition. Front Plant Sci 9:484. https://doi.org/10.3389/fpls.2018.00484
Gallinger J, Gross J (2020) Phloem metabolites of Prunus sp. rather than infection with Candidatus Phytoplasma prunorum influence feeding behavior of Cacopsylla pruni nymphs. J Chem Ecol 2020:1–15. https://doi.org/10.1007/s10886-020-01148-8
Gallinger J, Jarausch B, Jarausch W, Gross J (2020) Host plant preferences and detection of host plant volatiles of the migrating psyllid species Cacopsylla pruni, the vector of European Stone Fruit Yellows. J Pest Sci 93:461–475. https://doi.org/10.1007/s10340-019-01135-3
Garcia-Chapa M, Sabat J, Lavia A, Batlle A (2005) Role of Cacopsylla pyri in the epidemiology of pear decline in Spain. Eur J Plant Pathol 111:9–17. https://doi.org/10.1007/s10658-004-1981-y
George J, Robbins PS, Alessandro RT, Stelinski LL, Lapointe SL (2016) Formic and acetic acids in degradation products of plant volatiles elicit olfactory and behavioral responses from an insect vector. Chem Senses 41:325–338. https://doi.org/10.1093/chemse/bjw005
George J, Lapointe SL, Markle LT, Patt JM, Allan SA, Setamou M, Rivera MJ, Qureshi JA, Stelinski LL (2020) A multimodal attract-and-kill device for the Asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae). Insects 11:870. https://doi.org/10.3390/insects11120870
Görg LM, Gallinger J, Gross J (2020) The phytopathogen ‘Candidatus Phytoplasma mali’ alters apple tree phloem composition and affects oviposition behavior of its vector Cacopsylla picta. Chemoecology. https://doi.org/10.1007/s00049-020-00326-0
Gross J, Gallinger J, Rid M (2019a) Collection, identification, and statistical analysis of volatile organic compound patterns emitted by phytoplasma infected plants. Phytoplasmas. Humana Press, New York, NY, pp 333–343
Gross J, de Jorge BC, Gallinger J, Görg L, Maurer D, Rid M (2019b) The chemistry of multitrophic interactions in phytoplasma disease systems and advances in control of psyllid vectors with semiochemicals. Phytopathogenic Mollicutes 9:157–158
Gross J, Gallinger J, Görg L (2022) Interactions between phloem-restricted bacterial plant pathogens, their vector insects, host plants, and natural enemies, mediated by primary and secondary plant metabolites. Entomologia Generalis 42:185–215. https://doi.org/10.1127/entomologia/2021/1254
Guo H, Sun Y, Li Y, Liu X, Wang P, Zhu-Salzman K, Ge F (2014) Elevated CO2 alters the feeding behaviour of the pea aphid by modifying the physical and chemical resistance of Medicago truncatula. Plant, Cell Environ 37:2158–2168. https://doi.org/10.1111/pce.12306
Han J-H, Cho JG, Son I-C, Kim SH, Lee I-B, Choi IM, Kim D (2012) Effects of elevated carbon dioxide and temperature on photosynthesis and fruit characteristics of ‘Niitaka’ pear (Pyrus pyrifolia Nakai). Hortic Environ Biotechnol 53:357–361. https://doi.org/10.1007/s13580-012-0047-x
Havaux M, Greppin H, Strasser RJ (1991) Functioning of photosystems I and II in pea leaves exposed to heat stress in the presence or absence of light : analysis using in-vivo fluorescence, absorbance, oxygen and photoacoustic measurements. Planta 186:88–98. https://doi.org/10.1007/BF00201502
Hickman DT, Rasmussen A, Ritz K, Birkett MA, Neve P (2021) Review: allelochemicals as multi-kingdom plant defence compounds: towards an integrated approach. Pest Manag Sci 77:1121–1131. https://doi.org/10.1002/ps.6076
Himanen SJ, Nerg A-M, Nissinen A, Pinto DM, Stewart CN, Poppy GM, Holopainen JK (2009) Effects of elevated carbon dioxide and ozone on volatile terpenoid emissions and multitrophic communication of transgenic insecticidal oilseed rape (Brassica napus). New Phytol 181:174–186. https://doi.org/10.1111/j.1469-8137.2008.02646.x
IPCC (2014) Climate change 2014: Synthesis report. contribution of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change. Core Writing Team, Pachauri RK and Meyer LA (eds), IPCC, Geneva
Jarausch B, Tedeschi R, Sauvion N, Gross J, Jarausch W (2019) Psyllid vectors. In: Bertaccini A, Weintraub PG, Rao GP, Mori N (eds) Phytoplasmas: Plant Pathogenic Bacteria - II, vol 92. Springer Singapore, Singapore, pp 53–78
Jürgens A, Bischoff M (2017) Changing odour landscapes: the effect of anthropogenic volatile pollutants on plant–pollinator olfactory communication. Funct Ecol 31:56–64. https://doi.org/10.1111/1365-2435.12774
Kristoffersen L, Larsson MC, Anderbrant O (2008) Functional characteristics of a tiny but specialized olfactory system: olfactory receptor neurons of carrot psyllids (Homoptera: Triozidae). Chem Senses 33:759–769. https://doi.org/10.1093/chemse/bjn034
Lenth RV (2021) emmeans: estimated marginal means, aka least-squares means. R package. https://CRAN.R-project.org/package=emmeans
Mayer CJ, Vilcinskas A, Gross J (2008a) Pathogen-induced release of plant allomone manipulates vector insect behavior. J Chem Ecol 34:1518–1522. https://doi.org/10.1007/s10886-008-9564-6
Mayer CJ, Vilcinskas A, Gross J (2008b) Phytopathogen lures its insect vector by altering host plant odor. J Chem Ecol 34:1045–1049. https://doi.org/10.1007/s10886-008-9516-1
Meier U (2018) Growth stages of mono- and dicotyledonous plants: BBCH Monograph. https://doi.org/10.5073/20180906-074619
Mihaljević I, Lepeduš H, Šimić D, Viljevac Vuletić M, Tomaš V, Vuković D, Dugalić K, Teklić T, Skendrović Babojelić M, Zdunić Z (2020) Photochemical efficiency of photosystem II in two apple cultivars affected by elevated temperature and excess light in vivo. S Afr J Bot 130:316–326. https://doi.org/10.1016/j.sajb.2020.01.017
Nehlin G, Valterová I, Borg-Karlson AK (1994) Use of conifer volatiles to reduce injury caused by carrot psyllid, Trioza apicalis, Förster (Homoptera, Psylloidea). J Chem Ecol 20:771–783. https://doi.org/10.1007/BF02059612
O’Neill BF, Zangerl AR, DeLucia EH, Berenbaum MR (2010) Olfactory preferences of Popillia japonica, Vanessa cardui, and Aphis glycines for Glycine max grown under elevated CO2. Environ Entomol 39:1291–1301. https://doi.org/10.1603/EN09036
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2020) vegan: community ecology package. R package. https://CRAN.R-project.org/package=vegan
Patt JM, Meikle WG, Mafra-Neto A, Sétamou M, Mangan R, Yang C, Malik N, Adamczyk JJ (2011) Multimodal cues drive host-plant assessment in Asian citrus psyllid (Diaphorina citri). Environ Entomol 40:1494–1502. https://doi.org/10.1603/EN11149
Peters GP, Andrew RM, Canadell JG, Friedlingstein P, Jackson RB, Korsbakken JI, Le Quéré C, Peregon A (2020) Carbon dioxide emissions continue to grow amidst slowly emerging climate policies. Nat Clim Chang 10:3–6. https://doi.org/10.1038/s41558-019-0659-6
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Reineke A, Selim M (2019) Elevated atmospheric CO2 concentrations alter grapevine (Vitis vinifera) systemic transcriptional response to European grapevine moth (Lobesia botrana) herbivory. Sci Rep 9:2995. https://doi.org/10.1038/s41598-019-39979-5
Rid M, Mesca C, Ayasse M, Gross J (2016) Apple proliferation phytoplasma influences the pattern of plant volatiles emitted depending on pathogen virulence. Front Ecol Evol 3:152. https://doi.org/10.3389/fevo.2015.00152
Rid M, Markheiser A, Stein S, Hoffmann C, Gross J (2019) Volatiles of several grapevine cultivars emitted at different phenological stages linked to discriminatory ability of grapevine moths. J Plant Dis Prot 126:115–127. https://doi.org/10.1007/s41348-019-00214-y
Schoonhoven LM, van Loon JJA, Dicke M (2005) Insect-plant biology, 2nd edn. Oxford biology. Oxford Univ. Press, Oxford
Scutareanu P, Drukker B, Bruin J, Posthumus MA, Sabelis MW (1997) Volatiles from Psylla-infested pear trees and their possible involvement in attraction of anthocorid predators. J Chem Ecol 23:2241–2260. https://doi.org/10.1023/B:JOEC.0000006671.53045.16
Scutareanu P, Bruin J, Posthumus MA, Drukker B (2003) Constitutive and herbivore-induced volatiles in pear, alder and hawthorn trees. Chemoecology 13:63–74. https://doi.org/10.1007/s00049-002-0228-7
Shakoor A, Shakoor S, Rehman A, Ashraf F, Abdullah M, Shahzad SM, Farooq TH, Ashraf M, Manzoor MA, Altaf MM, Altaf MA (2021) Effect of animal manure, crop type, climate zone, and soil attributes on greenhouse gas emissions from agricultural soils—a global meta-analysis. J Clean Prod 278:124019. https://doi.org/10.1016/j.jclepro.2020.124019
Soroker V, Talebaev S, Harari AR, Wesley SD (2004) The role of chemical cues in host and mate location in the pear psylla Cacopsylla bidens (Homoptera: Psyllidae). J Insect Behav 17:613–626. https://doi.org/10.1023/B:JOIR.0000042544.35561.1c
Weintraub P, Gross J (2013) Capturing insect vectors of phytoplasmas. In: Dickinson M, Hodgetts J (eds) phytoplasma, vol 938. HumanaPress, Totowa, pp 61–72
Wenninger EJ, Stelinski LL, Hall DG (2009) Roles of Olfactory Cues, Visual Cues, and Mating Status in Orientation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) to Four Different Host Plants. Environ Entomol 36:225–234. https://doi.org/10.1603/022.038.0128
Wohlfahrt Y, Smith JP, Tittmann S, Honermeier B, Stoll M (2018) Primary productivity and physiological responses of Vitis vinifera L. cvs. under Free Air Carbon dioxide Enrichment (FACE). European Journal of Agronomy, 101, 149–162. Eur J Agron 101:149–162. https://doi.org/10.1016/J.EJA.2018.09.005
Yuvaraj JK, Andersson MN, Steinbauer MJ, Farnier K, Anderbrant O (2013) Specificity and sensitivity of plant odor-detecting olfactory sensory neurons in Ctenarytaina eucalypti (Sternorrhyncha: Psyllidae). J Insect Physiol 59:542–551. https://doi.org/10.1016/j.jinsphys.2013.03.004
Zeileis A, Kleiber C, Jackman S (2008) Regression models for count data in R. J Stat Softw 27:1–25
Acknowledgements
We thank Natalie Giesen, Thimo Braun, and Svenja Stein (JKI, Dossenheim, Germany) for excellent assistance in the lab and field. Felix Hergenhahn (JKI, Dossenheim, Germany) for care of the pear trees. We are grateful to Birgit Breckheimer, Winfried Schönbach, and Harald Schmidt (HGU, Geisenheim, Germany) for help with field experiments.
Funding
Open Access funding enabled and organized by Projekt DEAL. This work was supported by the federal Ministry of Food and Agriculture (BMEL) based on a decision of the Parliament of the Federal Republic of Germany via the Federal Office for Agriculture and Food (BLE) under the innovation support program (Grant number: 281B204316).
Author information
Authors and Affiliations
Contributions
M Rid-Moneta, C Becker, A Reineke, J Gallinger, and J Gross conceived the ideas and designed the experiments. M Rid-Moneta and J Gallinger conducted the experiments. J Gallinger analyzed the data and wrote the first draft of the manuscript, which was revisited and edited by M Rid-Moneta, C Becker, A Reineke, and J Gross.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Responsible Editor: Giovanni Benelli
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gallinger, J., Rid-Moneta, M., Becker, C. et al. Altered volatile emission of pear trees under elevated atmospheric CO2 levels has no relevance to pear psyllid host choice. Environ Sci Pollut Res 30, 43740–43751 (2023). https://doi.org/10.1007/s11356-023-25260-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-023-25260-w