
Abstract
Adenosine triphosphate (ATP) is coreleased with catecholamines from adrenal medullary chromaffin cells in response to sympathetic nervous system stimulation and may regulate these cells in an autocrine or paracrine manner. Increases in extracellular signal-regulated kinase (ERK) 1/2 phosphorylation were observed in response to ATP stimulation of bovine chromaffin cells. The signaling pathway involved in ATP-mediated ERK1/2 phosphorylation was investigated via Western blot analysis. ATP and uridine 5′-triphosphate (UTP) increased ERK1/2 phosphorylation potently, peaking between 5 and 15 min. The mitogen-activated protein kinase (MAPK/ERK)-activating kinase (MEK) inhibitor PD98059 blocked this response. UTP, which is selective for G-protein-coupled P2Y receptors, was the most potent agonist among several nucleotides tested. Adenosine 5′-O-(3-thio) triphosphate (ATPγS) and ATP were also potent agonists, characteristic of the P2Y2 or P2Y4 receptor subtypes, whereas agonists selective for P2X receptors or other P2Y receptor subtypes were weakly effective. The receptor involved was further characterized by the nonspecific P2 antagonists suramin and reactive blue 2, which each partially inhibited ATP-mediated ERK1/2 phosphorylation. Inhibitors of protein kinase C (PKC), protein kinase A (PKA), Ca2+/calmodulin-dependent protein kinase II (CaMKII), and phosphoinositide-3 kinase (PI3K) had no effect on ATP-mediated ERK1/2 phosphorylation. The Src inhibitor PP2, epidermal growth factor receptor (EGFR) inhibitor AG1478, and metalloproteinase inhibitor GM6001 decreased ATP-mediated ERK1/2 phosphorylation. These results suggest nucleotide-mediated ERK1/2 phosphorylation is mediated by a P2Y2 or P2Y4 receptor, which stimulates metalloproteinase-dependent transactivation of the EGFR.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Chromaffin cells are neuroendocrine cells that synthesize and secrete catecholamines in response to sympathetic nervous system stimulation and therefore participate in regulation of stress-modified parameters such as heart rate and blood pressure. A variety of additional agents are costored and released along with the catecholamines from the chromaffin granules, including neuropeptides such as the enkephalins and adenosine triphosphate (ATP) [1]. In addition to being released into the circulation, these agents may regulate chromaffin cell activity in an autocrine or a paracrine manner, allowing the cells to adjust to varying levels of stimulation.
The role of ATP in chromaffin cell function has not been well defined, even though it is secreted in high concentrations from chromaffin cells [1]. It has been suggested that ATP regulates chromaffin cell secretion, either positively [2] or negatively [3, 4], and that ATP regulates the function of voltage-dependent calcium channels [5]. ATP exerts its effects through either G-protein-coupled receptors, designated P2Y; or ion channels, designated P2X. These receptor types are further divided into subtypes, including P2Y1, P2Y2, P2Y4, P2Y6, and P2Y11–14 for the G-protein-coupled P2Y receptors and P2X1–7 for those that activate ion channels [6]. Previous work with chromaffin cells indicated ATP stimulation results in increases in inositol phosphates [7], cyclic adenosine monophosphate (cAMP) [8], and [Ca2+]i accumulation [7], likely via activation of a P2Y2 or P2Y4 receptor, both of which are present in bovine chromaffin cells (unpublished observations). The downstream effects of P2Y receptor stimulation by ATP in chromaffin cells are not known. The observed increases in signaling messengers may bring about the activation of multiple protein kinases or tyrosine kinases.
In several cell types, ATP signaling has been shown to activate extracellular signal-regulated kinase 1 and 2 (ERK1/2). P2Y receptors have been shown to couple to ERK1/2 activation via activation of protein kinases such as phosphoinositide-3 kinase (PI3K) [11] or protein kinase C (PKC) [10, 11]. P2Y receptor-mediated ERK1/2 has also been shown to be dependent on activation of tyrosine kinases such as Src or proline-rich tyrosine kinase (Pyk2) [9, 10] and/or on transactivation of the epidermal growth factor receptor (EGFR) [9]. Additionally, increases in cAMP in response to ATP may result in activation of protein kinase A (PKA), which has been shown to activate ERK1/2 [12]. Increases in [Ca2+]i in response to ATP may result in activation of Ca2+/calmodulin-dependent protein kinase II (CaMKII), which has also been shown to phosphorylate ERK1/2 [13]. Therefore, we elected to examine ATP-mediated ERK1/2 phosphorylation and the receptor subtype and signaling mechanism present in bovine chromaffin cells.
Materials and methods
Chromaffin cell isolation and cell culture
Bovine adrenal chromaffin cells (BACC) were isolated using a collagenase perfusion method as described previously [14, 15]. Cells were maintained on six-well plates at a density of 3 × 106 cells/well at 37°C with 5% CO2. Viability and purity were verified to be >95% by Trypan blue exclusion and neutral red staining, respectively.
Western blot analysis
Cells were incubated with agonist for 10 min, and inhibitor preincubations were 15 min, unless otherwise indicated. Cells were rinsed with phosphate buffered saline (PBS) and lysed with 200 μl of 50 mM N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid (HEPES) (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid), pH 7.2, containing 1 mM ethylenediaminetetraacetate (EDTA), 1 mM ethyleneglycoltetraacetic acid (EGTA), 0.2% triton X-100, 10 mM β-glycerol 2-phosphate disodium salt, 1 mM sodium orthovanadate, 1 mM benzamidine, 4 μg/ml leupeptin, 1 μM microcystin-LR, and 0.5 mM DTT. Lysates were centrifuged at 13,000 g, and protein concentrations of supernatants were determined with the Bio-Rad protein assay (Bio-Rad, Hercules, CA, USA). Loading buffer (2X) was added to cell lysates, consisting of 125 mM Tris(hydroxymethyl)aminomethane HCl (Research Organics, Cleveland, OH, USA), 4% sodium dodecylsulfate (SDS) (Research Organics, Cleveland, OH, USA), 20% glycerol, and 0.02% bromophenol blue. Samples were subsequently boiled and subjected to SDS-polyacrylamide gel electrophoresis (PAGE) on 10% Tris-HCl Criterion gels (BioRad, Hercules, CA, USA).
Protein was transferred to fluorescent-polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA, USA). Membranes were next blocked in Odyssey blocking buffer (LI-COR Biosciences, Lincoln, NE, USA) for 1 h, then incubated in Odyssey blocking buffer containing 0.2% Tween 20 (polyoxyethylene-sorbitan monolaurate) and mouse-anti-phospho-ERK 1/2 (Thr202/Tyr204) and rabbit-anti-ERK 1/2 (Cell Signaling Technology, Boston, MA, USA) primary antibodies. Blots were then incubated in Odyssey blocking buffer with 0.2% Tween 20 and 0.02% SDS and with the secondary antibodies goat-anti-mouse immunoglobulin (Ig)G Alexa Fluor680 (Molecular Probes, Eugene, OR, USA) and goat-anti-rabbit IgG IR800 (Rockland Immunochemicals, Gilbertsville, PA, USA). Membranes were developed with the Odyssey Infrared Imaging System, which utilizes two infrared channels (700 nm and 800 nm), allowing for detection of two target proteins simultaneously, in this case phosphorylated and total ERK1/2.
Statistics
Band integrated intensities were determined with Odyssey Imaging software. Phospho-ERK1/2 intensities were divided by total ERK1/2 intensities and normalized to fold increases over control. Data were analyzed with GraphPad Prism software, and one-way analysis of variance (ANOVA) was utilized to determine statistical significance. The decision was made to utilize ERK2 for graphical representations, as band intensities for phosphorylated ERK2 were stronger than phosphorylated ERK1, though results obtained were quantitatively similar for both.
Chemicals
Nucleotides and analogs were obtained from Sigma-Aldrich (St. Louis, MO, USA). PD98059 (2′-amino-3′-methoxylflavone), PMA (phorbol 12-myristate 13-acetate), suramin, and reactive blue 2 (RB2) were also obtained from Sigma-Aldrich. NF279 [8,8′-[Carbonylbis(imino-4,1-phenylenecarbonylimino-4,1-phenylenecarbonylimino)]bis-1,3,5-naphthalenetrisulfonic acid hexasodium salt] was purchased from Tocris (St. Louis, MO, USA). KT5720 [(9S,10S,12R)-2,3,9,10,11,12-Hexahydro-10-hydroxy-9-methyl-1-oxo-9,12-epoxy-1H-diindolo[1,2,3-fg:3′,2′,1′-kl]pyrrolo[3,4-i][1,6]benzodiazocine-10-carboxylic acid hexyl ester], Ro-31–8220 [2-[1-(3-(Amidinothio)propyl)-1H-indol-3-yl]-3-(1-methylindol-3-yl)maleimide methanesulfonate], LY294002 ]2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one], PP1 [4-amino-5-(4-methylphenyl)-7-(t-butyl)pyrazolo-D-3,4-pyrimidine], PP2 [4-amino-5-(4-chlorophenyl)-7-(t-butyl)pyrazolo-D-3,4-pyrimidine], AG1478 [4-(3-Chloroanilino)-6,7-dimethoxyquinazoline], and GM6001 [N-[(2R)-2-(hydroxamidocarbonylmethyl)-4-methylpentanoyl]-L-tryptophan methylamide] were purchased from Biomol (Philadelphia, PA, USA). Bis-I (bisindolylmaleimide-I), KN-92 [2-[N-(4-methoxybenzenesulfonyl)]amino-N-(4-chlorocinnamyl)-N-methylbenzylamine], KN-93 [2-[N-(2-hydroxyethyl)]-N-(4-methoxybenzenesulfonyl)]amino-N-(4-chlorocinnamyl)-N-methylbenzylamine], and wortmannin were purchased from Merck-Calbiochem Biosciences (Darmstadt, Germany). H89 [N-[2-(p-Bromocinnamylamino)ethyl]-5-isoquinolinesulfonamide] was obtained from Upstate (Charlottesville, VA, USA).Other reagents were obtained from either Sigma-Aldrich or Fisher Scientific.
Results
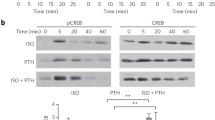
Western blot analyses with antibodies specific to activated ERK1/2 phosphorylated at Thr202/Tyr204 were utilized to examine the time course of ERK1/2 phosphorylation in response to nucleotide stimulation. Both ATP and UTP potently increased ERK1/2 phosphorylation, with a peak between 5 and 15 min (Fig. 1). ERK1/2 has been shown to be catalyzed only by MEK; therefore, the MEK inhibitor PD98059 was utilized to confirm the immediate upstream signaling event responsible for nucleotide-mediated ERK1/2 phosphorylation. PD98059 (10 μM) significantly decreased both ATP-mediated (∼90%) and UTP-mediated (∼70%, Fig. 2, Table 1) ERK1/2 phosphorylation.

Nucleotide-stimulated ERK1/2 phosphorylation is time dependent. BACCs were treated with ATP (100 μM) or UTP (100 μM) for 2 to 30 min. Blots are representative of three independent experiments performed in triplicate (n = 3); the upper band (phosphorylated or nonphosphorylated) is ERK1 = 44 kDa, and the lower band is ERK2 = 42 kDa. Blot intensities were measured with the Odyssey Imaging System; values are phosphorylated ERK2 intensity divided by total ERK2 intensity. Points on the graph represent mean ± standard error of the mean

MEK inhibition decreases ATP- and UTP-mediated ERK1/2 phosphorylation. BACCs were treated with PD98059 (10 μM) or dimethylsulfoxide for 15 min, followed by a 10-min stimulation with ATP (100 μM) or UTP (100 μM). Blots are representative of three independent experiments performed in triplicate (n = 3); the upper band (phosphorylated or nonphosphorylated) is ERK1 = 44 kDa and the lower band is ERK2 = 42 kDa. Blot intensities were measured with the Odyssey Imaging System; values are phosphorylated ERK2 intensity divided by total ERK2 intensity. Bars on the graph represent mean ± standard error of the mean. *** p < 0.001 vs. stimulator alone
Most studies designed to determine purinergic receptor subtypes use ligand potency studies due to the lack of available highly selective receptor subtype antagonists [16]. Correspondingly, we used a similar approach to characterize the receptor subtype involved in ERK1/2 phosphorylation. Examination of several purine analogs revealed a rank order of potency of UTP (EC50 = 1.6 μM)>ATPγS (6.5 μM)≥ATP (13 μM)>uridine diphosphate (UDP) (120 μM) = adenosine diphosphate (ADP) (220 μM) = 2-methylthio (ATP) 2-MeSATP (320 μM)>>α,β-methylene ATP (α,β-meATP) (Fig. 3), consistent with the involvement of a P2Y2 or P2Y4 receptor, as both UTP and ATP exhibit strong agonist action.

P2Y2 or P2Y4 receptor activation increases ERK1/2 phosphorylation. BACCs were treated with increasing concentrations of nucleotides and analogs for 10 min. Blots are representative of three independent experiments performed in triplicate (n = 3); the upper band (phosphorylated or nonphosphorylated) is ERK1 = 44 kDa, and the lower band is ERK2 = 42 kDa. Blot intensities were measured with the Odyssey Imaging System; values are phosphorylated ERK2 intensity divided by total ERK2 intensity. Points on the graph represent mean ± standard error of the mean. Con (control) refers to results obtained with unstimulated cells
The involvement of a P2 receptor in ERK1/2 phosphorylation was further supported using the nonselective P2 receptor antagonists suramin and RB2. Suramin (100 μM) significantly decreased ATP- or UTP-mediated ERK1/2 phosphorylation (∼60%, Fig. 4, Table 1). RB2 (100 μM) also decreased the effect of ATP- or UTP-stimulation on ERK1/2 phosphorylation (∼35%, Fig. 4, Table 1). The P2X-specific receptor agonist α,β-meATP had no effect on ERK1/2 phosphorylation at concentrations up to 100 μM (Fig. 3), eliminating the involvement of several of the P2X receptor subtypes. Moreover, UTP is selective for P2Y receptors, precluding the involvement of a P2X receptor in nucleotide-mediated ERK1/2 phosphorylation.

P2 receptor antagonists partially block ATP- and UTP-mediated ERK1/2 phosphorylation. BACCs were pretreated with or without suramin (100 μM) or reactive blue 2 (RB2, 100 μM) for 15 min, followed by a 10-min stimulation with or without ATP (100 μM) or UTP (100 μM). Blots are representative of three independent experiments performed in triplicate (n = 3); the upper band (phosphorylated or nonphosphorylated) is ERK1 = 44 kDa, and the lower band is ERK2 = 42 kDa. Blot intensities were measured with the Odyssey Imaging System; values are phosphorylated ERK2 intensity divided by total ERK2 intensity. Bars on graph represent mean ± standard error of the mean. *** p < 0.001 vs. stimulator alone
ATP stimulation of chromaffin cells has been shown to increase inositol phosphates [7], cAMP [8], and [Ca2+]i accumulation [7], which may lead to activation of several protein kinases, which may in turn be involved in ATP-mediated phosphorylation of ERK1/2. Therefore, a variety of inhibitors were used to examine which kinases are involved in ATP-mediated ERK1/2 phosphorylation. The PKA inhibitors H89 (10 μM) and KT5720 (100 nM) had no effect on ATP- or UTP-mediated ERK1/2 phosphorylation (Table 1). The broad-spectrum PKC inhibitors Ro-31–8220 (10 μM) and Bis-I (3.5 μM) also had no effect on ATP- or UTP-mediated ERK1/2 phosphorylation, although they were capable of blocking PMA-mediated ERK1/2 phosphorylation (∼60%, Table 1). Moreover, the CaMKII inhibitor KN93 (1 μM) and the PI3K inhibitors wortmannin (300 nM) and LY294002 (20 μM) had no effect on ATP- or UTP-mediated ERK1/2 phosphorylation (Table 1).
In addition to activation of PKs, P2Y receptors may utilize tyrosine kinases to activate ERK1/2 [10]. Therefore, the tyrosine kinase inhibitor PP2 was used to examine the role of Src family members in ATP-mediated ERK1/2 phosphorylation. PP2 (1 μM) decreased ATP- or UTP-mediated ERK2 phosphorylation (∼40%, Fig. 5, Table 1). In addition to inhibiting Src family members, PP2 is also a weak inhibitor of the EGFR. Also, G-protein-coupled receptor activation of Src family members may result in activation of ERK1/2 via activation of Ras [17, 18] or via transactivation of the EGFR. Therefore, the involvement of EGFR in ATP-mediated ERK1/2 phosphorylation was determined by treating cells with the EGFR inhibitor AG1478 (2.6 μM), which decreased ATP- and UTP-mediated ERK2 phosphorylation by about 70% (Fig. 6a, Table 1). EGF-mediated ERK1/2 phosphorylation was completely blocked by AG1478 (100%, p < 0.001, data not shown). Transactivation of the EGFR by G-protein-coupled receptors may be mediated by tyrosine kinases such as Src or via metalloproteinases, which release EGFR ligands such as heparin-binding EGF-like growth factor (HB-EGF) from the cell membrane [19]. The metalloproteinase inhibitor GM6001 (2.5 μM) decreased ATP- and UTP-mediated ERK1/2 phosphorylation by about 65% (Fig. 6b, Table 1).

Tyrosine kinase inhibition decreases ATP- and UTP-mediated ERK1/2 phosphorylation. BACCs were pretreated with PP2 (1 μM) or dimethylsulfoxide (DMSO) for 15 min, followed by a 10-min stimulation with or without ATP (100 μM) or UTP (100 μM). Blots are representative of three independent experiments performed in triplicate (n = 3); the upper band (phosphorylated or nonphosphorylated) is ERK1 = 44 kDa, and the lower band is ERK2 = 42 kDa. Blot intensities were measured with the Odyssey Imaging System; values are phosphorylated ERK2 intensity divided by total ERK2 intensity. Bars on the graphs represent mean ± standard error of the mean. *** p < 0.001 vs. stimulator alone

Nucleotide-mediated ERK1/2 phosphorylation is dependent on EGFR transactivation. BACCs were pretreated with or without AG1478 (2.6 μM) (a), GM6001 (2.5 μM) (b) and dimethylsulfoxide (DMSO) for 15 min, followed by a 10-min stimulation with or without ATP (100 μM) or UTP (100 μM). Blots are representative of three independent experiments performed in triplicate (n = 3); the upper band (phosphorylated or nonphosphorylated) is ERK1 = 44 kDa ,and the lower band is ERK2 = 42 kDa. Blot intensities were measured with the Odyssey Imaging System; values are phosphorylated ERK2 intensity divided by total ERK2 intensity. Bars on the graphs represent mean ± standard error of the mean. *** p < 0.001 vs. stimulator alone
Discussion
ATP and UTP potently increase ERK1/2 phosphorylation, with a peak between 5 and 15 min. This rapid peak in ERK1/2 phosphorylation in response to ATP would allow the cells to respond quickly to varying levels of stimulation. Although the physiological effects of ERK1/2 phosphorylation in these cells are unknown, possible actions requiring a rapid response include either the acute activation of proteins involved in catecholamine secretion and/or stimulation of protein expression important for exocytosis.
Ligand potency and inhibitor studies suggest either the P2Y2 or P2Y4 receptor subtype is responsible for nucleotide-mediated ERK1/2 phosphorylation, similar to data obtained for increases in inositol phosphates (unpublished observations). Both of these receptor subtypes are present in chromaffin cells, based on reverse transcriptase real-time polymerase chain reaction (PCR) data for P2Y2 and P2Y4, and appear to be expressed in these cells according to Western blot analysis with specific antibodies (unpublished observations.) Several lines of evidence suggest P2X receptors are not involved in the increase in ERK1/2 in response to nucleotide stimulation. First, UTP does not activate P2X ion channels but potently increases ERK1/2 phosphorylation. Additionally, α,βmeATP, an agonist selective for several P2X receptor subtypes, had no effect on ERK1/2 phosphorylation. The P2Y receptor involved is most likely either P2Y2 or P2Y4. UTP is highly selective for two P2Y receptor subtypes P2Y2 and P2Y4 and weakly effective on the P2Y6 receptor. The P2Y6 subtype can be ruled out because of the subtypes activated by UTP; only the P2Y2 and P2Y4 subtypes are also strongly activated by ATP. The weak effect of ADP, UDP and 2-MeSATP confirms this designation, as these agonists are specific for P2 receptor subtypes other than the P2Y2 or P2Y4 subtypes [6]. There are no available agonists or antagonists to distinguish between the P2Y2 and P2Y4 receptors. Even so, suramin and RB2 are commonly used to characterize these receptors in a given cell type, and their partial effectiveness is not contradictory to results found in other cell types for P2Y2 or P2Y4 receptors [20].
Nucleotides utilize multiple signaling pathways in different cell types to bring about increases in ERK1/2 phosphorylation. PKC and PI3K have been implicated in P2Y-mediated ERK1/2 phosphorylation [10, 11]. In PC12 cells, ERK1/2 phosphorylation in response to P2Y2 receptor activation has been shown to be both dependent [9, 10] and independent [21] of the small tyrosine kinase Pyk2. P2Y2 receptors have also been shown to contain an integrin-binding domain, arginine-glycine-aspartic acid (RGD), which is necessary for ERK1/2 activation in astrocytes [22, 23]. Additionally, P2Y2 receptors contain SH3-binding sites that associate with Src in astrocytoma cells [24] and astrocytes [23]. Also, in PC12 cells P2Y2 receptors have been shown to require EGFR transactivation to increase ERK1/2 phosphorylation [9].
Initially, we examined whether signaling pathways mediated by protein kinases were involved in ATP-mediated ERK1/2 phosphorylation. Previous studies determined that ATP-mediated stimulation of bovine chromaffin cells results in increases in inositol phosphates [7], cAMP [8], and [Ca2+]i accumulation [7]. P2Y2 or P2Y4 receptors couple to Gq to increase activation of PKC. Additionally, the observed increases in cAMP may result in activation of PKA, whereas increased [Ca2+]i may result in activation of CaMKII. However, inhibitors of each of these protein kinases had no effect on ATP- or UTP-mediated ERK1/2 phosphorylation, including the PKA inhibitors H89 and KT5720, PKC inhibitors Bis-I and Ro-81–3220, or the CaMKII inhibitor KN93. The PKC inhibitors Bis-I and Ro-81–3220 were capable of blocking PMA-mediated ERK1/2 phosphorylation, suggesting PKC can couple to ERK1/2 phosphorylation in these cells, and yet confirming the lack of involvement of this pathway in ATP-mediated ERK1/2 phosphorylation. We also examined the PI3K inhibitors wortmannin and LY294002, as this kinase has been implicated in P2Y-mediated ERK1/2 phosphorylation. These inhibitors also proved to be ineffective, suggesting the protein kinases examined were not responsible for ATP-mediated ERK1/2 phosphorylation.
We next examined the involvement of tyrosine kinases in ATP-mediated ERK1/2 phosphorylation, as these kinases have been shown to be involved in P2Y-mediated ERK1/2 phosphorylation. PP2 (1 μM) significantly decreased ATP- and UTP-mediated ERK1/2 phosphorylation. The reported IC50s for PP2-mediated inhibition of Src family members are in the nanomolar range (http://www.biomol.com), whereas the dose used in these studies is reported to cause weak inhibition of the EGFR [25]. Lower doses of PP2 had no effect on ATP- or UTP-mediated ERK1/2 phosphorylation (data not shown.) Therefore, at the effective dose used, it is not possible to conclude whether Src or EGFR inhibition was responsible for the decrease in ATP-mediated ERK1/2 phosphorylation. Additionally, activation of Src family members by G-protein-coupled receptors may increase ERK1/2 phosphorylation via activation of the renin angiotensin system (Ras) [17, 18] or via transactivation of the EGFR [26]. Thus, the involvement of EGFR in ATP-mediated ERK1/2 phosphorylation was investigated further. Inhibition of the EGFR with the specific inhibitor AG1478 decreased ATP- and UTP-mediated ERK1/2 phosphorylation, strongly suggesting EGFR transactivation is important for ATP-mediated ERK1/2 phosphorylation.
G-protein-coupled receptors may transactivate the EGFR via activation of tyrosine kinases such as Src, or via activation of metalloproteinases to generate EGFR ligands such as HB-EGF [19]. The role of metalloproteinases in ATP-mediated ERK1/2 phosphorylation was investigated with broad-spectrum inhibitor GM6001, which significantly decreased the response, suggesting that HB-EGF cleavage mediated by metalloproteinases in response to ATP may be responsible for transactivation of the EGFR and subsequently stimulation of ERK1/2 phosphorylation.
As expected, the MEK inhibitor PD98059 blocked ATP- and UTP-mediated ERK1/2 phosphorylation, confirming that MEK, the only known kinase upstream of ERK1/2, contributes to ERK1/2 phosphorylation. PD98059 blocked ATP-mediated ERK1/2 phosphorylation (∼90%) to a greater extent than UTP-mediated ERK1/2 phosphorylation (∼70%). This may be due to the fact that UTP is a more potent agonist and elicited a larger response than ATP for ERK1/2 phosphorylation. For the other inhibitors, the responses were very similar; however, none of the other inhibitors had such a pronounced effect on ATP-mediated ERK1/2 phosphorylation. Alternatively, ATP and UTP may activate multiple receptors with distinct signaling pathways that are variously more specific for ATP or UTP. Activation of ERK1/2 independent of MEK1 may involve inhibition of phosphatases.
Further studies are necessary to determine the consequence(s) of ERK1/2 phosphorylation in response to ATP stimulation of chromaffin cells. As chromaffin cells are nonproliferating, the stimulation of ERK1/2 phosphorylation by ATP and UTP may couple to regulation of gene transcription essential to exocytosis.
To our knowledge this is the first study demonstrating phosphorylation of ERK1/2 in response to ATP or UTP stimulation in bovine chromaffin cells. Our data show that the ERK1/2 phosphorylation response to ATP is mediated by either a P2Y2 or P2Y4 receptor. Protein kinases are not involved in nucleotide-mediated ERK1/2 phosphorylation, but rather, metalloproteinase-dependent transactivation of the EGFR is necessary for ATP-mediated ERK1/2 phosphorylation.
Abbreviations
- BACC:
-
bovine adrenal chromaffin cells
- ERK1/2:
-
extracellular signal-regulated kinase 1 and 2
- EGF:
-
epidermal growth factor
- EGFR:
-
EGF receptor
- MEK:
-
mitogen-activated protein kinase/ERK kinase
- [Ca2+]i :
-
cytosolic free Ca2+ concentration
- PKC:
-
protein kinase C
- PKA:
-
protein kinase A
- CaMKII:
-
Ca2+/calmodulin-dependent protein kinase II
- PI3K:
-
phosphoinositide-3 kinase
- 2-MeSATP:
-
2-methylthio ATP
- α,β-meATP:
-
α,β-methylene ATP
- HB-EGF:
-
heparin-binding EGF-like growth factor
- Pyk2:
-
proline-rich tyrosine kinase
- SH3:
-
Src homology 3
References
Viveros OH, Diliberto EJ Jr, Daniels AJ (1983) Biochemical and functional evidence for the cosecretion of multiple messengers from single and multiple compartments. Fed Proc 42:2923–2928
Reichsman F, Santos S, Westhead EW (1995) Two distinct ATP receptors activate calcium entry and internal calcium release in bovine chromaffin cells. J Neurochem 65:2080–2086
Ennion SJ, Powell AD, Seward EP (2004) Identification of the P2Y(12) receptor in nucleotide inhibition of exocytosis from bovine chromaffin cells. Mol Pharmacol 66:601–611
Chen XK, Wang LC, Zhou Y, Cai Q, Prakriya M, Duan KL, Sheng ZH, Lingle C, Zhou Z (2005) Activation of GPCRs modulates quantal size in chromaffin cells through G(betagamma) and PKC. Nat Neurosci 8:1160–1168
Ohta T, Kai T, Ito S (2004) Evidence for paracrine modulation of voltage-dependent calcium channels by amperometric analysis in cultured porcine adrenal chromaffin cells. Brain Res 1030:183–192
Burnstock G (2006) Pathophysiology and therapeutic potential of purinergic signaling. Pharmacol Rev 58:58–86
Zheng J, Zhang P, Toews M, Hexum TD (1997) Neuropeptide Y enhances ATP-induced formation of inositol phosphates in chromaffin cells. Biochem Biophys Res Commun 239:287–290
Zhang P, Zheng J, Bradley ME, Hexum TD (2001) ATP stimulated cyclic AMP formation in bovine chromaffin cells is enhanced by neuropeptide Y. Peptides 22:439–444
Soltoff SP (1998) Related adhesion focal tyrosine kinase and the epidermal growth factor receptor mediate the stimulation of mitogen-activated protein kinase by the G-protein-coupled P2Y2 receptor. Phorbol ester or [Ca2+]i elevation can substitute for receptor activation. J Biol Chem 273:23110–23117
Soltoff SP, Avraham H, Avraham S, Cantley LC (1998) Activation of P2Y2 receptors by UTP and ATP stimulates mitogen-activated kinase activity through a pathway that involves related adhesion focal tyrosine kinase and protein kinase C. J Biol Chem 273:2653–2660
Montiel M, de la Blanca EP, Jimenez E (2006) P2Y receptors activate MAPK/ERK through a pathway involving PI3K/PDK1/PKC-zeta in human vein endothelial cells. Cell Physiol Biochem 18:123–134
Schmitt JM, Stork PJ (2000) beta 2-adrenergic receptor activates extracellular signal-regulated kinases (ERKs) via the small G protein rap1 and the serine/threonine kinase B-Raf. J Biol Chem 275:25342–25350
Agell N, Bachs O, Rocamora N, Villalonga P (2002) Modulation of the Ras/Raf/MEK/ERK pathway by Ca(2+), and calmodulin. Cell Signal 14:649–654
Wilson SP, Kirshner N (1983) Preparation and maintenance of adrenal medullary chromaffin cell cultures. Methods Enzymol 103:305–312
Drakulich DA, Spellmon C, Hexum TD (2004) Effect of the ecto-ATPase inhibitor, ARL 67156, on the bovine chromaffin cell response to ATP. Eur J Pharmacol 485:137–140
Murthy KS, Makhlouf GM (1998) Coexpression of ligand-gated P2X and G protein-coupled P2Y receptors in smooth muscle. Preferential activation of P2Y receptors coupled to phospholipase C (PLC)-beta1 via Galphaq/11 and to PLC-beta3 via Gbetagammai3. J Biol Chem 273:4695–4704
Luttrell LM, Hawes BE, van BT, Luttrell DK, Lansing TJ, Lefkowitz RJ (1996) Role of c-Src tyrosine kinase in G protein-coupled receptor- and Gbetagamma subunit-mediated activation of mitogen-activated protein kinases. J Biol Chem 271:19443–19450
Dikic I, Tokiwa G, Lev S, Courtneidge SA, Schlessinger J (1996) A role for Pyk2 and Src in linking G-protein-coupled receptors with MAP kinase activation. Nature 383:547–550
Prenzel N, Zwick E, Daub H, Leserer M, Abraham R, Wallasch C, Ullrich A (1999) EGF receptor transactivation by G-protein-coupled receptors requires metalloproteinase cleavage of proHB-EGF. Nature 402:884–888
Burnstock G (2002) Potential therapeutic targets in the rapidly expanding field of purinergic signalling. Clin Med 2:45–53
Barsacchi R, Heider H, Girault J, Meldolesi J (1999) Requirement of pyk2 for the activation of the MAP kinase cascade induced by Ca(2+) (but not by PKC or G protein) in PC12 cells. FEBS Lett 461:273–276
Erb L, Liu J, Ockerhausen J, Kong Q, Garrad RC, Griffin K, Neal C, Krugh B, Santiago-Perez LI, Gonzalez FA, Gresham HD, Turner JT, Weisman GA (2001) An RGD sequence in the P2Y(2) receptor interacts with alpha(V)beta(3) integrins and is required for G(o)-mediated signal transduction. J Cell Biol 153:491–501
Weisman GA, Wang M, Kong Q, Chorna NE, Neary JT, Sun GY, Gonzalez FA, Seye CI, Erb L (2005) Molecular determinants of P2Y2 nucleotide receptor function: implications for proliferative and inflammatory pathways in astrocytes. Mol Neurobiol 31:169–183
Liu J, Liao Z, Camden J, Griffin KD, Garrad RC, Santiago-Perez LI, Gonzalez FA, Seye CI, Weisman GA, Erb L (2004) Src homology 3 binding sites in the P2Y2 nucleotide receptor interact with Src and regulate activities of Src, proline-rich tyrosine kinase 2, and growth factor receptors. J Biol Chem 279:8212–8218
Chen JK, Capdevila J, Harris RC (2000) Overexpression of C-terminal Src kinase blocks 14, 15-epoxyeicosatrienoic acid-induced tyrosine phosphorylation and mitogenesis. J Biol Chem 275:13789–13792
Daub H, Wallasch C, Lankenau A, Herrlich A, Ullrich A (1997) Signal characteristics of G protein-transactivated EGF receptor. EMBO J 16:7032–7044
Acknowledgment
We thank Matthew Beaver for excellent technical assistance and Robin Taylor for expert assistance with graphics. This work was supported by a grant to Terry D. Hexum from the American Heart Association (#0550030Z).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Luke, T.M., Hexum, T.D. UTP and ATP increase extracellular signal-regulated kinase 1/2 phosphorylation in bovine chromaffin cells through epidermal growth factor receptor transactivation. Purinergic Signalling 4, 323–330 (2008). https://doi.org/10.1007/s11302-008-9098-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11302-008-9098-y